
Pollen Coat Proteomes of Arabidopsis thaliana, Arabidopsis lyrata, and Brassica oleracea Reveal Remarkable Diversity of Small Cysteine-Rich Proteins at the Pollen-Stigma Interface

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Extractions of Pollen Coat Proteins
2.3. Identification of Pollen Coat Proteins by Tandem Mass Spectrometry
2.4. Bioinformatic Analyses
3. Results
3.1. Isolation and Proteomic Profiling of the Pollen Coat Proteins from Arabidopsis thaliana, Arabidopsis lyrata, and Brassica oleracea
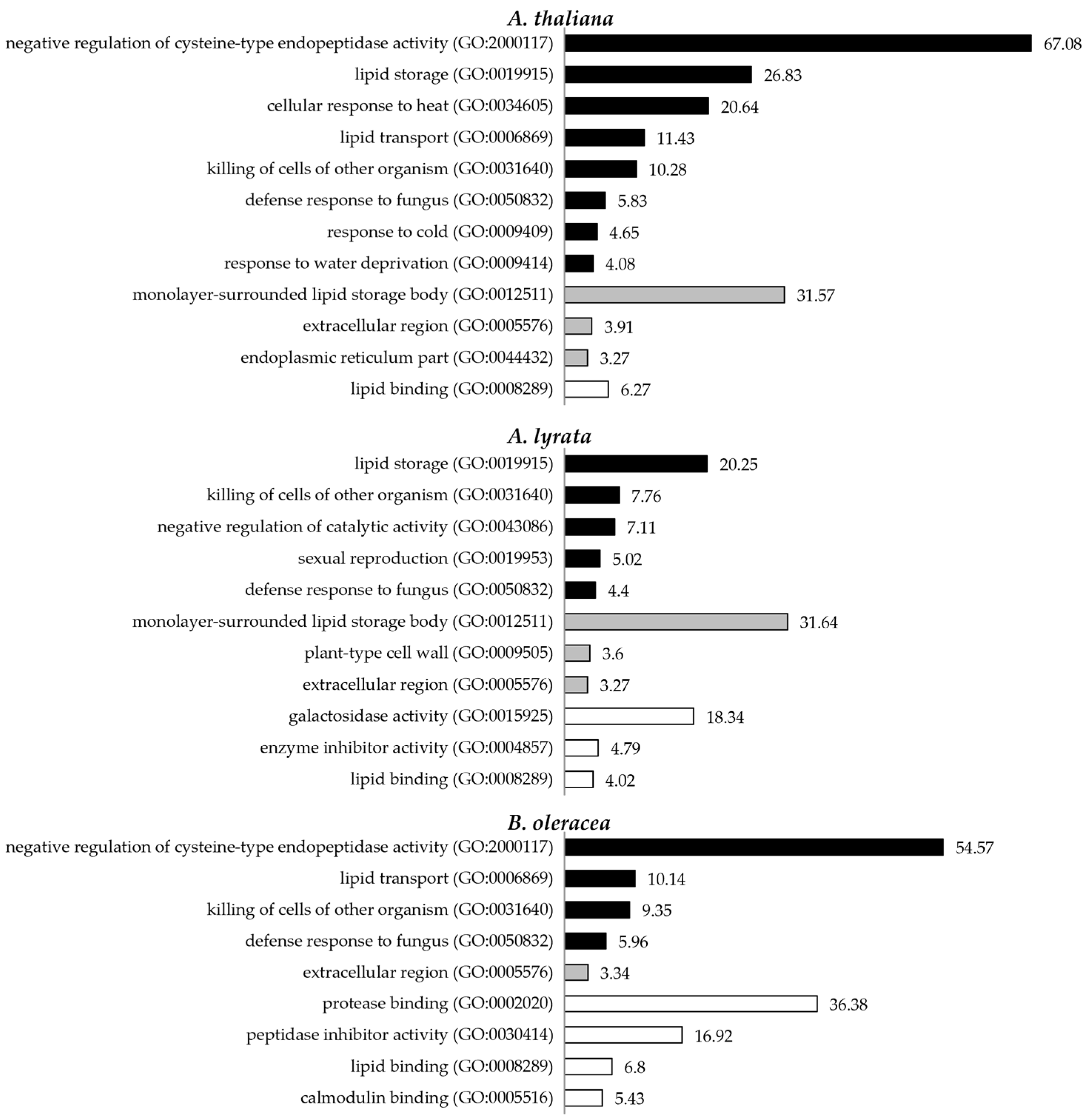
3.2. Pollen Coat Proteins Are Enriched in Functions Related to Signalling, the Cell Wall, and Lipid Metabolism
3.3. Small CRPs Are Highly Represented in the Pollen Coat of Members of the Brassicaceae
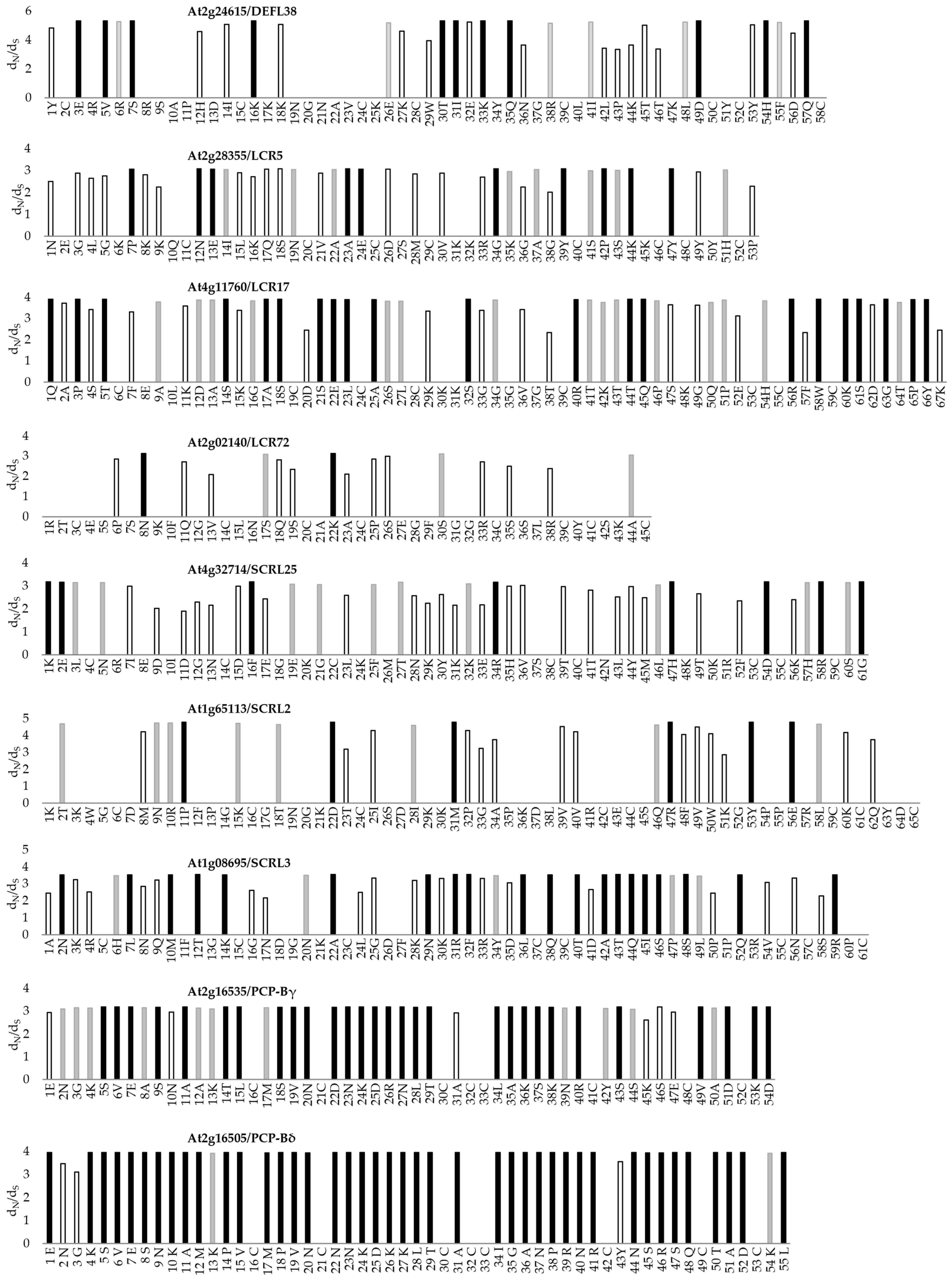
3.4. Evidence of Positive Selection in Regions of CRPs Identified in the Pollen Coat Proteomes
4. Discussion
4.1. Significant Expansion of the Known Pollen Coat Proteome in the Brassicaceae
4.2. Enrichment of the Proteins Associated with Signal Transduction, Cell Wall, and Lipid Metabolism in Pollen Coat Proteomes
4.3. Enrichment of CRPs in the Pollen Coat Provides Insights into the Evolutionary Link between CRP Signalling during Reproduction and Defence
4.4. Positive Selection on CRP-Encoding Genes Suggests Roles in Contributing to Species Barrier Formation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pacini, E.; Hesse, M. Pollenkitt—Its composition, forms and functions. Flora: Morphol. Distrib. Funct. Ecol. Plants. 2005, 200, 399–415. [Google Scholar] [CrossRef]
- Hesloph-Harrison, J. Tapetal origin of pollen-coat substances in lilium. New Phytol. 1968, 67, 779–786. [Google Scholar] [CrossRef]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Biogenesis and function of the lipidic structures of pollen grains. Sex. Plant Reprod. 1998, 11, 65–80. [Google Scholar] [CrossRef]
- Hernández-Pinzón, I.; Ross, J.H.E.; Barnes, K.A.; Damant, A.P.; Murphy, D.J. Composition and role of tapetal lipid bodies in the biogenesis of the pollen coat of Brassica napus. Planta 1999, 208, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Rejón, J.D.; Delalande, F.; Schaeffer-Reiss, C.; Alché, J.D.; Rodríguez-García, M.I.; Van Dorsselaer, A.; Castro, A.J. The pollen coat proteome: At the cutting edge of plant reproduction. Proteomes 2016, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, H.G.; Lewis, D. Formation of tryphine coating pollen grains of Raphanus, and its properties relating to self-incompatibility system. Proc. R. Soc. Lond. B Biol. 1973, 184, 149–165. [Google Scholar]
- Doughty, J.; Dixon, S.; Hiscock, S.J.; Willis, A.C.; Parkin, I.A.P.; Dickinson, H.G. PCP-A1, a defensin-like Brassica pollen coat protein that binds the S-locus glycoprotein, is the product of gametophytic gene expression. Plant Cell 1998, 10, 1333–1347. [Google Scholar] [CrossRef] [Green Version]
- Shiba, H.; Takayama, S.; Iwano, M.; Shimosato, H.; Funato, M.; Nakagawa, T.; Che, F.S.; Suzuki, G.; Watanabe, M.; Hinata, K.; et al. A pollen coat protein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiol. 2001, 125, 2095–2103. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Clarke, L.A.; Eason, R.J.; Parker, C.C.; Qi, B.X.; Scott, R.J.; Doughty, J. PCP-B class pollen coat proteins are key regulators of the hydration checkpoint in Arabidopsis thaliana pollen-stigma interactions. New Phytol. 2017, 213, 764–777. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y.; Shivanna, K.R. The receptive surface of the Angiosperm stigma. Ann. Bot. 1977, 41, 1233–1258. [Google Scholar] [CrossRef] [Green Version]
- Elleman, C.J.; Franklin-Tong, V.; Dickinson, H.G. Pollination in species with dry stigmas: The nature of the early stigmatic response and the pathway taken by pollen tubes. New Phytol. 1992, 121, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Abhinandan, K.; Sankaranarayanan, S.; Macgregor, S.; Goring, D.R.; Samuel, M.A. Cell–cell signaling during the Brassicaceae self-incompatibility response. Trends Plant Sci. 2021, 27, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, H. Dry stigmas, water and self-incompatibility in Brassica. Sex. Plant Reprod. 1995, 8, 1–10. [Google Scholar] [CrossRef]
- Safavian, D.; Goring, D.R. Secretory activity is rapidly induced in stigmatic papillae by compatible pollen, but inhibited for self-incompatible pollen in the Brassicaceae. PLoS ONE 2013, 8, e84286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doucet, J.; Lee, H.K.; Goring, D.R. Pollen Acceptance or Rejection: A tale of two pathways. Trends Plant Sci. 2016, 21, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Preuss, D.; Lemieux, B.; Yen, G.; Davis, R.W. A conditional sterile mutation eliminates surface components from Arabidopsis pollen and disrupts cell signaling during fertilization. Genes Dev. 1993, 7, 974–985. [Google Scholar] [CrossRef] [Green Version]
- Hülskamp, M.; Kopczak, S.D.; Horejsi, T.F.; Kihl, B.K.; Pruitt, R.E. Identification of genes required for pollen-stigma recognition in Arabidopsis thaliana. Plant J. 1995, 8, 703–714. [Google Scholar] [CrossRef]
- Fiebig, A.; Mayfield, J.A.; Miley, N.L.; Chau, S.; Fischer, R.L.; Preuss, D. Alterations in CER6, a gene identical to CUT1, differentially affect long-chain lipid content on the surface of pollen and stems. Plant Cell 2000, 12, 2001–2008. [Google Scholar] [CrossRef] [Green Version]
- Zinkl, G.M.; Zwiebel, B.I.; Grier, D.G.; Preuss, D. Pollen-stigma adhesion in Arabidopsis: A species-specific interaction mediated by lipophilic molecules in the pollen exine. Development 1999, 126, 5431–5440. [Google Scholar] [CrossRef]
- Mayfield, J.A.; Preuss, D. Rapid initiation of Arabidopsis pollination requires the oleosin-domain protein GRP17. Nat Cell Biol. 2000, 2, 128–130. [Google Scholar] [CrossRef]
- Updegraff, E.P.; Zhao, F.; Preuss, D. The extracellular lipase EXL4 is required for efficient hydration of Arabidopsis pollen. Sex. Plant Reprod. 2009, 22, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef] [PubMed]
- Doughty, J.; Wong, H.Y.; Dickinson, H.G. Cysteine-rich pollen coat proteins (PCPs) and their interactions with stigmatic S (incompatibility) and S-related proteins in Brassica: Putative roles in SI and pollination. Ann. Bot. 2000, 85, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Shiba, H.; Iwano, M.; Asano, K.; Hara, M.; Che, F.S.; Watanabe, M.; Hinata, K.; Isogai, A. Isolation and characterization of pollen coat proteins of Brassica campestris that interact with S locus-related glycoprotein 1 involved in pollen-stigma adhesion. Proc. Natl. Acad. Sci. USA 2000, 97, 3765–3770. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Shen, L.; Xiao, Y.; Vyshedsky, D.; Peng, C.; Sun, X.; Liu, Z.; Cheng, L.; Zhang, H.; Han, Z.; et al. Pollen PCP-B peptides unlock a stigma peptide-receptor kinase gating mechanism for pollination. Science 2021, 372, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Marshall, E.; Costa, L.M.; Gutierrez-Marcos, J. Cysteine-Rich Peptides (CRPs) mediate diverse aspects of cell–cell communication in plant reproduction and development. J. Exp. Bot. 2011, 62, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.-J.; Li, L.; Lan, Z.; Dresselhaus, T. Peptide signalling during the pollen tube journey and double fertilization. J. Exp. Bot. 2015, 66, 5139–5150. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.Y.; Duan, Q.; Li, C.; Liu, M.-C.J.; Wu, H.-M. Pollen–pistil interactions: It takes two to tangle but a molecular cast of many to deliver. Curr. Opin. Plant Biol. 2022, 69, 102279. [Google Scholar] [CrossRef]
- Noir, S.; Bräutigam, A.; Colby, T.; Schmidt, J.; Panstruga, R. A reference map of the Arabidopsis thaliana mature pollen proteome. Biochem. Biophys. Res. Commun. 2005, 337, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Li, L.; Chen, T.; Chong, K.; Xue, Y.; Wang, T. Proteomic analyses of Oryza sativa mature pollen reveal novel proteins associated with pollen germination and tube growth. Proteomics 2006, 6, 2504–2529. [Google Scholar] [CrossRef]
- Murphy, D.J.; Ross, J.H. Biosynthesis, targeting and processing of oleosin-like proteins, which are major pollen coat components in Brassica napus. Plant J. 1998, 13, 1–16. [Google Scholar] [PubMed]
- Mayfield, J.A.; Fiebig, A.; Johnstone, S.E.; Preuss, D. Gene families from the Arabidopsis thaliana pollen coat proteome. Science 2001, 292, 2482–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Cai, G.; Gong, F.; An, S.; Cresti, M.; Wang, W. Proteome profiling of maize pollen coats reveals novel protein components. Plant Mol. Biol. Rep. 2015, 33, 975–986. [Google Scholar] [CrossRef]
- Hohmann, N.; Wolf, E.M.; Lysak, M.A.; Koch, M.A. A time-calibrated road map of Brassicaceae species radiation and evolutionary history. The Plant Cell 2015, 27, 2770–2784. [Google Scholar] [CrossRef] [Green Version]
- Tsuchimatsu, T.; Goubet, P.M.; Gallina, S.; Holl, A.-C.; Fobis-Loisy, I.; Bergès, H.; Marande, W.; Prat, E.; Meng, D.; Long, Q.; et al. Patterns of polymorphism at the self-incompatibility locus in 1083 Arabidopsis thaliana genomes. Mol. Bio. Evo. 2017, 34, 1878–1889. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, A.G.; Doughty, J.; Dixon, S.; Elleman, C.; Hiscock, S.; Dickinson, H.G. The male determinant of self-incompatibility in Brassica oleracea is located in the pollen coating. Plant J. 1997, 12, 1351–1359. [Google Scholar] [CrossRef]
- Johnson-Brousseau, S.A.; McCormick, S. A compendium of methods useful for characterizing Arabidopsis pollen mutants and gametophytically-expressed genes. Plant J. 2004, 39, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Doughty, J.; Hedderson, F.; McCubbin, A.; Dickinson, H. Interaction between a coating-borne peptide of the Brassica pollen grain and stigmatic S (self-incompatibility)-locus-specific glycoproteins. Proc. Natl. Acad. Sci. USA 1993, 90, 467–471. [Google Scholar] [CrossRef] [Green Version]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Intra- and extracellular lipid composition and associated gene expression patterns during pollen development in Brassica napus. Plant J. 1997, 11, 549–562. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Logacheva, M.D.; Dmitriev, S.E.; Penin, A.A. RNA-seq analysis of an apical meristem time series reveals a critical point in Arabidopsis thaliana flower initiation. BMC Genom. 2015, 16, 466. [Google Scholar] [CrossRef] [Green Version]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Vanoosthuyse, V.; Miege, C.; Dumas, C.; Cock, J.M. Two large Arabidopsis thaliana gene families are homologous to the Brassica gene superfamily that encodes pollen coat proteins and the male component of the self-incompatibility response. Plant Mo. Bio. 2001, 46, 17–34. [Google Scholar] [CrossRef]
- Roxrud, I.; Lid, S.E.; Fletcher, J.C.; Schmidt, E.D.L.; Opsahl-Sorteberg, H.-G. GASA4, one of the 14-member Arabidopsis GASA family of small polypeptides, regulates flowering and seed development. Plant Cell Physiol 2007, 48, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.T.; Moskal, W.A., Jr.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Edstam, M.M.; Viitanen, L.; Salminen, T.A.; Edqvist, J. Evolutionary history of the non-specific lipid transfer proteins. Mol Plant 2011, 4, 947–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.H. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Bio. Evo. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Suen, D.F.; Wu, S.S.; Chang, H.C.; Dhugga, K.S.; Huang, A.H. Cell wall reactive proteins in the coat and wall of maize pollen: Potential role in pollen tube growth on the stigma and through the style. J. Biol. Chem. 2003, 278, 43672–43681. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M.A.; O’Leary, S.; Wu, S.; Gleddie, S.; Eudes, F.; Laroche, A.; Robert, L.S. A molecular and proteomic investigation of proteins rapidly released from triticale pollen upon hydration. Plant Mol. Biol. 2012, 79, 101–121. [Google Scholar] [CrossRef]
- Bosch, M.; Wang, L. Pollen-stigma interactions in Brassicaceae: Complex communication events regulating pollen hydration. J. Exp. Bot. 2020, 71, 2465–2468. [Google Scholar] [CrossRef]
- Foote, H.C.; Ride, J.P.; Franklin-Tong, V.E.; Walker, E.A.; Lawrence, M.J.; Franklin, F.C. Cloning and expression of a distinctive class of self-incompatibility (S) gene from Papaver rhoeas L. Proc. Natl. Acad. Sci. USA 1994, 91, 2265–2269. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, M.J.; de Graaf, B.H.J.; Hadjiosif, N.; Perry, R.M.; Poulter, N.S.; Osman, K.; Vatovec, S.; Harper, A.; Franklin, F.C.H.; Franklin-Tong, V.E. Identification of the pollen self-incompatibility determinant in Papaver rhoeas. Nature 2009, 459, 992–995. [Google Scholar] [CrossRef]
- Rajasekar, K.V.; Ji, S.; Coulthard, R.J.; Ride, J.P.; Reynolds, G.L.; Winn, P.J.; Wheeler, M.J.; Hyde, E.I.; Smith, L.J. Structure of SPH (self-incompatibility protein homologue) proteins: A widespread family of small, highly stable, secreted proteins. Biochem. J. 2019, 476, 809–826. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Silverstein, K.A.T.; Gao, L.; Walton, J.D.; Nallu, S.; Guhlin, J.; Young, N.D. Detecting small plant peptides using SPADA (Small Peptide Alignment Discovery Application). BMC Bioinform. 2013, 14, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, M.J.; Vatovec, S.; Franklin-Tong, V.E. The pollen S-determinant in Papaver: Comparisons with known plant receptors and protein ligand partners. J. Exp. Bot. 2010, 61, 2015–2025. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z. A superfamily of proteins with novel cysteine-rich repeats. Plant Physiol. 2001, 126, 473–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaattovaara, A.; Brandt, B.; Rajaraman, S.; Safronov, O.; Veidenberg, A.; Luklová, M.; Kangasjärvi, J.; Löytynoja, A.; Hothorn, M.; Salojärvi, J.; et al. Mechanistic insights into the evolution of DUF26-containing proteins in land plants. Commun. Bio. 2019, 2, 56. [Google Scholar] [CrossRef] [PubMed]
- Bourdais, G.; Burdiak, P.; Gauthier, A.; Nitsch, L.; Salojärvi, J.; Rayapuram, C.; Idänheimo, N.; Hunter, K.; Kimura, S.; Merilo, E.; et al. Large-scale phenomics identifies primary and fine-tuning roles for CRKs in responses related to oxidative stress. PLoS Genet. 2015, 11, e1005373. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Kim, Y.C.; Kwon, S.J.; Ryu, C.-M.; Park, O.K. The Arabidopsis cysteine-rich receptor-like kinase CRK36 regulates immunity through interaction with the cytoplasmic kinase BIK1. Front. Plant Sci. 2017, 8, 1856. [Google Scholar] [CrossRef] [Green Version]
- Yadeta, K.A.; Elmore, J.M.; Creer, A.Y.; Feng, B.; Franco, J.Y.; Rufian, J.S.; He, P.; Phinney, B.; Coaker, G. a cysteine-rich protein kinase associates with a membrane immune complex and the cysteine residues are required for cell death. Plant Physiol. 2017, 173, 771–787. [Google Scholar] [CrossRef] [Green Version]
- van Eerde, A.; Grahn, E.M.; Winter, H.C.; Goldstein, I.J.; Krengel, U. Atomic-resolution structure of the α-galactosyl binding Lyophyllum decastes lectin reveals a new protein family found in both fungi and plants. Glycobiology 2015, 25, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Tateno, H. SUEL-related lectins, a lectin family widely distributed throughout organisms. Biosci. Biotechnol. Biochem. 2010, 74, 1141–1144. [Google Scholar] [CrossRef] [PubMed]
- Trainotti, L.; Spinello, R.; Piovan, A.; Spolaore, S.; Casadoro, G. beta-Galactosidases with a lectin-like domain are expressed in strawberry. J. Exp. Bot. 2001, 52, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Hiscock, S.J.; Doughty, J.; Willis, A.C.; Dickinson, H.G. A 7-kDa pollen coating-borne peptide from Brassica napus interacts with S-locus glycoprotein and S-locus-related glycoprotein. Planta 1995, 196, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Stanchev, B.S.; Doughty, J.; Scutt, C.P.; Dickinson, H.; Croy, R.R. Cloning of PCP1, a member of a family of pollen coat protein (PCP) genes from Brassica oleracea encoding novel cysteine-rich proteins involved in pollen-stigma interactions. Plant J. 1996, 10, 303–313. [Google Scholar] [CrossRef]
- Silverstein, K.A.T.; Graham, M.A.; Paape, T.D.; VandenBosch, K.A. Genome organization of more than 300 defensin-like genes in Arabidopsis. Plant Physiol. 2005, 138, 600–610. [Google Scholar] [CrossRef] [Green Version]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: Classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Jauh, G.-Y.; Mollet, J.-C.; Eckard, K.J.; Nothnagel, E.A.; Walling, L.L.; Lord, E.M. A lipid transfer-like protein is necessary for lily pollen tube adhesion to an in vitro stylar matrix. Plant Cell 2000, 12, 151–164. [Google Scholar]
- Chae, K.; Kieslich, C.A.; Morikis, D.; Kim, S.-C.; Lord, E.M. A gain-of-function mutation of Arabidopsis lipid transfer protein 5 disturbs pollen tube tip growth and fertilization. Plant Cell 2009, 21, 3902–3914. [Google Scholar] [CrossRef] [Green Version]
- Paul, W.; Hodge, R.; Smartt, S.; Draper, J.; Scott, R. The isolation and characterisation of the tapetum-specific Arabidopsis thaliana A9 gene. Plant Mol. Biol. 1992, 19, 611–622. [Google Scholar] [CrossRef]
- Huang, M.D.; Chen, T.L.; Huang, A.H. Abundant type III lipid transfer proteins in Arabidopsis tapetum are secreted to the locule and become a constituent of the pollen exine. Plant Physiol. 2013, 163, 1218–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinovich, L.; Weiss, D. The Arabidopsis cysteine-rich protein GASA4 promotes GA responses and exhibits redox activity in bacteria and in planta. Plant J. 2010, 64, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Nahirñak, V.; Almasia, N.I.; Fernandez, P.V.; Hopp, H.E.; Estevez, J.M.; Carrari, F.; Vazquez-Rovere, C. Potato snakin-1 gene silencing affects cell division, primary metabolism, and cell wall composition. Plant Physiol. 2012, 158, 252–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, C.; Xu, H.; Ye, S.; Wang, S.; Li, L.; Zhang, S.; Wang, X. Gibberellic acid-stimulated arabidopsis6 serves as an integrator of gibberellin, abscisic acid, and glucose signaling during seed germination in Arabidopsis. Plant Physiol. 2015, 169, 2288–2303. [Google Scholar] [PubMed] [Green Version]
- Wu, T.; Cheng, C.; Zhong, Y.; Lv, Y.; Ma, Y.; Zhong, G. Molecular characterization of the gibberellin-stimulated transcript of GASA4 in Citrus. Plant Growth Regu. 2020, 91, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Smyth, D.R.; Bowman, J.L.; Meyerowitz, E.M. Early flower development in Arabidopsis. Plant Cell 1990, 2, 755–767. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Swanson, W.J.; Vacquier, V.D. The rapid evolution of reproductive proteins. Nat. Rev. Genet. 2002, 3, 137–144. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Blackman, B.K. Speciation genes in plants. Ann. Bot. 2010, 106, 439–455. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, R.; Yang, Z. Likelihood models for detecting positively selected amino acid sites and applications to the HIV-1 envelope gene. Genetics 1998, 148, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nielsen, R.; Goldman, N.; Pedersen, A.-M.K. Codon-substitution models for heterogeneous selection pressure at amino acid sites. Genetics 2000, 155, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Wong, W.S.W.; Nielsen, R. Bayes empirical Bayes inference of amino acid sites under positive selection. Mol. Bio. Evo. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, L.; Yang, Y.; Cui, J.; Chang, F.; Wang, Y.; Ma, H. Analysis of Arabidopsis floral transcriptome: Detection of new florally expressed genes and expansion of Brassicaceae-specific gene families. Front. Plant Sci. 2015, 5, 802. [Google Scholar] [CrossRef]
- Honys, D.; Twell, D. Comparative analysis of the Arabidopsis pollen transcriptome. Plant Physiol. 2003, 132, 640–652. [Google Scholar]
- Holmes-Davis, R.; Tanaka, C.K.; Vensel, W.H.; Hurkman, W.J.; McCormick, S. Proteome mapping of mature pollen of Arabidopsis thaliana. Proteomics 2005, 5, 4864–4884. [Google Scholar] [CrossRef]
- Marmagne, A.; Ferro, M.; Meinnel, T.; Bruley, C.; Kuhn, L.; Garin, J.; Barbier-Brygoo, H.; Ephritikhine, G. A high content in lipid-modified peripheral proteins and integral receptor kinases features in the Arabidopsis plasma membrane proteome. Mol. Cell. Proteom. 2007, 6, 1980–1996. [Google Scholar] [CrossRef] [Green Version]
- Ito, J.; Batth, T.S.; Petzold, C.J.; Redding-Johanson, A.M.; Mukhopadhyay, A.; Verboom, R.; Meyer, E.H.; Millar, A.H.; Heazlewood, J.L. Analysis of the Arabidopsis cytosolic proteome highlights subcellular partitioning of central plant metabolism. J. Proteome Res. 2011, 10, 1571–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolovski, N.; Rubtsov, D.; Segura, M.P.; Miles, G.P.; Stevens, T.J.; Dunkley, T.P.J.; Munro, S.; Lilley, K.S.; Dupree, P. Putative glycosyltransferases and other plant Golgi apparatus proteins are revealed by LOPIT proteomics. Plant Physiol. 2012, 160, 1037–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, X.; Liu, B. Tapetum-dependent male meiosis progression in plants: Increasing evidence emerges. Front. Plant Sci. 2020, 10, 1667. [Google Scholar] [CrossRef] [Green Version]
- Elleman, C.J.; Willson, C.E.; Sarker, R.H.; Dickinson, H.G. Interaction between the pollen tube and stigmatic cell wall following pollination in Brassica oleracea. New Phyto. 1988, 109, 111–117. [Google Scholar] [CrossRef]
- Elleman, C.J.; Dickinson, H.G. The role of the exine coating in pollen-stigma interactions in Brassica oleracea L. New Phytol. 1990, 114, 511–518. [Google Scholar] [CrossRef]
- Hiscock, S.J.; Allen, A.M. Diverse cell signalling pathways regulate pollen-stigma interactions: The search for consensus. New Phytol. 2008, 179, 286–317. [Google Scholar] [CrossRef]
- Rivera, A.M.; Wilburn, D.B.; Swanson, W.J. Domain expansion and functional diversification in vertebrate reproductive proteins. Mol. Bio. Evo. 2022, 39, msac105. [Google Scholar] [CrossRef]
- Qi, M.; Link, T.I.; Müller, M.; Hirschburger, D.; Pudake, R.N.; Pedley, K.F.; Braun, E.; Voegele, R.T.; Baum, T.J.; Whitham, S.A. A small cysteine-rich protein from the Asian Soybean Rust Fungus, Phakopsora pachyrhizi, suppresses plant immunity. PLOS Pathog. 2016, 12, e1005827. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Yin, Z.; Li, Z.; Wu, Y.; Huang, L. A small cysteine-rich protein from two kingdoms of microbes is recognized as a novel pathogen-associated molecular pattern. New Phytol. 2018, 222, 995–1011. [Google Scholar] [CrossRef]
- Wang, D.; Tian, L.; Zhang, D.-D.; Song, J.; Song, S.-S.; Yin, C.-M.; Zhou, L.; Liu, Y.; Wang, B.-L.; Kong, Z.-Q.; et al. Functional analyses of small secreted cysteine-rich proteins identified candidate effectors in Verticillium dahliae. Mol. Plant Pathol. 2020, 21, 667–685. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Yang, G.; Lin, H.; Liu, Y.; Li, P.; Zheng, A. A Novel, Small cysteine-rich effector, RsSCR10 in Rhizoctonia solani is sufficient to trigger plant cell death. Front. Microbiol. 2021, 12, 684923. [Google Scholar] [CrossRef]
- Snelders, N.C.; Petti, G.C.; van den Berg, G.C.M.; Seidl, M.F.; Thomma, B.P.H.J. An ancient antimicrobial protein co-opted by a fungal plant pathogen for in planta mycobiome manipulation. Proc. Nat. Acad. Sci. USA 2021, 118, e2110968118. [Google Scholar] [CrossRef]
- Wang, Y.-D.; Li, F.; Zhang, X.; Liu, T.-L.; Liang, W.-X.; Li, D.-L. PnSCR82, a small cysteine-rich secretory protein of Phytophthora nicotianae, can enhance defense responses in plants. J. Integr. Agric. 2022, 21, 751–761. [Google Scholar] [CrossRef]
- Nasrallah, J.B. Recognition and rejection of self in plant self-incompatibility: Comparisons to animal histocompatibility. Trends Immunol. 2005, 26, 412–418. [Google Scholar] [CrossRef]
- Sanabria, N.; Goring, D.; Nürnberger, T.; Dubery, I. Self/nonself perception and recognition mechanisms in plants: A comparison of self-incompatibility and innate immunity. New Phytol. 2008, 178, 503–514. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Márton, M.L. Micropylar pollen tube guidance and burst: Adapted from defense mechanisms? Curr. Opin. Plant Biol. 2009, 12, 773–780. [Google Scholar] [CrossRef]
- Bircheneder, S.; Dresselhaus, T. Why cellular communication during plant reproduction is particularly mediated by CRP signalling. J. Exp. Bot. 2016, 67, 4849–4861. [Google Scholar] [CrossRef]
- Lay, F.T.; Anderson, M.A. Defensins—Components of the innate immune system in plants. Curr. Protein Pept. Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.J.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Balandín, M.; Royo, J.; Gómez, E.; Muniz, L.M.; Molina, A.; Hueros, G. A protective role for the embryo surrounding region of the maize endosperm, as evidenced by the characterisation of ZmESR-6, a defensin gene specifically expressed in this region. Plant Mol. Biol. 2005, 58, 269–282. [Google Scholar] [CrossRef]
- Amien, S.; Kliwer, I.; Márton, M.L.; Debener, T.; Geiger, D.; Becker, D.; Dresselhaus, T. Defensin-like ZmES4 mediates pollen tube burst in maize via opening of the potassium channel KZM1. PLoS Biol. 2010, 8, e1000388. [Google Scholar] [CrossRef] [Green Version]
- Woriedh, M.; Merkl, R.; Dresselhaus, T. Maize EMBRYO SAC family peptides interact differentially with pollen tubes and fungal cells. J. Exp. Bot. 2015, 66, 5205–5216. [Google Scholar] [CrossRef] [Green Version]
- Kessler, S.A.; Shimosato-Asano, H.; Keinath, N.F.; Wuest, S.E.; Ingram, G.; Panstruga, R.; Grossniklaus, U. Conserved molecular components for pollen tube reception and fungal invasion. Science 2010, 330, 968–971. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Z.; Wu, D.; Yu, F. RALF–FERONIA signaling: Linking plant immune response with cell growth. Plant Commun. 2020, 1, 100084. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Li, L.; Luan, S. A receptor–channel trio conducts Ca2+ signalling for pollen tube reception. Nature 2022, 607, 534–539. [Google Scholar] [CrossRef]
- Mondragón-Palomino, M.; John-Arputharaj, A.; Pallmann, M.; Dresselhaus, T. Similarities between reproductive and immune pistil transcriptomes of Arabidopsis species. Plant Physiol. 2017, 174, 1559–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Dresselhaus, T.; Gu, H.; Qu, L.-J. Active role of small peptides in Arabidopsis reproduction: Expression evidence. J. Integr. Plant Biol. 2015, 57, 518–521. [Google Scholar] [CrossRef]
- Clark, N.L.; Aagaard, J.E.; Swanson, W.J. Evolution of reproductive proteins from animals and plants. Reprod. 2006, 131, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, C.S.; Yim, W.C.; Moon, J.-C.; Jung, J.H.; Lee, T.G.; Lim, S.D.; Cho, S.H.; Lee, K.K.; Kim, W.; Seo, Y.W.; et al. Evolution of non-specific lipid transfer protein (nsLTP) genes in the Poaceae family: Their duplication and diversity. Mol. Genet. Genomics 2008, 279, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, A.; Kimport, R.; Preuss, D. Comparisons of pollen coat genes across Brassicaceae species reveal rapid evolution by repeat expansion and diversification. Proc. Natl. Acad. Sci. USA 2004, 101, 3286–3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schein, M.; Yang, Z.H.; Mitchell-Olds, T.; Schmid, K.J. Rapid evolution of a pollen-specific oleosin-like gene family from Arabidopsis thaliana and closely related species. Mol. Bio. Evo. 2004, 21, 659–669. [Google Scholar] [CrossRef]
- Moyle, L.C.; Wu, M.; Gibson, M.J.S. Reproductive proteins evolve faster than non-reproductive proteins among Solanum species. Front. Plant Sci. 2021, 12, 635990. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Jiao, H.; Li, L.; Qiao, X.; Fabrice, M.R.; Wu, J.; Zhang, S. Expansion and evolutionary patterns of cysteine-rich peptides in plants. BMC Genom. 2017, 18, 610. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-L.; Zhao, X.; Lanz, C.; Weigel, D. Evolution of the S-Locus region in Arabidopsis relatives. Plant Physiol. 2011, 157, 937–946. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Description | A. t | A. l | B. o | |
|---|---|---|---|---|
| Signal transduction | 14-3-3-like protein | 4 | 2 | 0 |
| Cysteine-rich repeat secretory protein (CRRSP) | 7 | 7 | 14 | |
| F-box protein | 2 | 5 | 1 | |
| D-galactoside/L-rhamnose binding SUEL protein | 3 | 3 | 5 | |
| Kinase | 6 | 16 | 14 | |
| S-protein homolog (SPH) | 6 | 8 | 3 | |
| Small cysteine-rich protein (CRP) | 47 | 45 | 70 | |
| Lipid metabolism | Acyl-CoA-binding protein | 1 | 1 | 2 |
| Acyltransferase | 2 | 1 | 0 | |
| esterase/lipase (EXL) | 5 | 4 | 3 | |
| Oleosin/Glycine-rich protein (GRP) | 8 | 6 | 10 | |
| Glycosyltransferase | 1 | 2 | 0 | |
| Lipid binding protein | 1 | 1 | 2 | |
| Sterol carrier protein | 1 | 0 | 1 | |
| Cell wall-related | Acetylglucosaminyltransferase | 0 | 1 | 1 |
| Galactosidase | 1 | 5 | 1 | |
| Glucosidase | 1 | 1 | 0 | |
| Pectin acetylesterase | 0 | 1 | 1 | |
| Pectin lyase-like protein | 0 | 5 | 1 | |
| Pectin methylesterase inhibitor (PMEI) | 1 | 6 | 3 | |
| Pectinase | 1 | 0 | 1 | |
| Pectinesterase | 1 | 1 | 0 | |
| Xyloglucan endotransglucosylase | 0 | 1 | 1 | |
| Response to stress | Heat shock cognate protein | 0 | 1 | 2 |
| Late embryogenesis abundant protein (LEA) family protein | 4 | 5 | 2 | |
| Protein cold-regulated | 2 | 2 | 0 | |
| Proteinase inhibitor | 4 | 3 | 4 | |
| Redox | FAD-linked oxidase | 1 | 2 | 0 |
| Galactose oxidase | 0 | 1 | 1 | |
| Glutaredoxin | 2 | 2 | 1 | |
| Peroxidase | 1 | 6 | 4 | |
| Peroxygenase | 1 | 2 | 0 | |
| Proteolysis | Aspartic proteinase | 1 | 1 | 1 |
| Carboxypeptidase | 2 | 2 | 2 | |
| Cysteine proteinase | 2 | 2 | 0 | |
| Protease | 1 | 2 | 1 | |
| Proteasome | 2 | 0 | 1 | |
| CRP Classes (Subgroup Identifiers) | Species | No. of CRPs | Sizes | Cysteine Patterns |
|---|---|---|---|---|
| PCP-A/DEFL/LCR (CRP0000, 0260, 0520, 0560, 0570, 0580, 0650, 0700, 0710, 0960) | A. thaliana | 19 | 76–90 | CX(5-12)CX(4-8)CXXXCX(9-16)CX(3-14)CXCX(0-5)C |
| A. lyrata | 21 | 53–96 | CX(7-16)CX(4-8)CXXXCX(9-16)CX(4-13)CXC(1-8)C | |
| B. oleracea | 32 | 52–209 | CX(3-21)CX(4-11)CXXXCX(9-15)CX(4-12)CXCX(1-6)C | |
| PCP-B/PCP-BL (CRP5460, 5500, 5515) | A. thaliana | 4 | 74–82 | CXXXXCX(7-8)CXCCX(6-8)CX(6)CXXXC |
| A. lyrata | 2 | 74–77 | CXXXXCX(7-8)CXCCX(6-8)CX(6)CXXXC | |
| B. oleracea | 2 | 66–134 | CX(4-10)CX(8)CXCCX(6-9)CX(6)CXXXC | |
| SCR/SCRL (CRP0830) | A. thaliana | 10 | 87–98 | CX(9)CX(7)CX(13-19)CXCX(11-15)CXCX(3-7)C |
| A. lyrata | 13 | 79–105 | CX(9-10)CX(7-10)CX(13-19)CXCX(10-16)CXCX(3-7)C | |
| B. oleracea | 11 | 73–108 | CX(9-10)CX(7-8)CX(13-23)CXCX(11-27)CXCX(3-7)C | |
| nsLTP (CRP3860, 4000, 4380, 4670, 4820, 4900) | A. thaliana | 10 | 91–180 | CX(6-9)CX(13-16)CCX(8-19)CXCX(12-25)CX(6-15)C |
| A. lyrata | 7 | 93–119 | CX(6-9)CX(12-16)CCX(8-19)CXCX(17-24)CX(6-15)C | |
| B. oleracea | 20 | 90–265 | CX(6-9)CX(13-16)CCX(8-19)CXCX(12-25)CX(6-13)C | |
| GASA (CRP2700) | A. thaliana | 2 | 89–94 | CXXXCXXXCX(8)CXXXCXXCCXXCX(1-2)CX(11)CXCX(12)C |
| A. lyrata | 2 | 89–94 | CXXXCXXXCX(8)CXXXCXXCCXXCX(1-2)CX(11)CXCX(12)C | |
| B. oleracea | 2 | 89–91 | CXXXCXXXCX(8)CXXXCXXCCXXCXXCX(11)CXCX(12)C | |
| Others (CRP1850, 2865, 3600, 3670) | A. thaliana | 2 | 104–140 | - |
| A. lyrata | 0 | - | - | |
| B. oleracea | 3 | 73–165 | - |
| Accession (Description) | Gene Locus | Ns/ LA | dN/dS (M0) | 2ΔInL (M7 vs. M8) | Estimated Parameters (M8) | Positively Selected Sites |
|---|---|---|---|---|---|---|
| Q2V462(DEFL38) | AT2G24615 | 8/87 | 0.845 | 39.059 *** | 13.4% (ω = 3.217) | 23E, 25V, 29S, 36K, 50T, 51I, 53K, 55Q, 70D, 75H, 83Q, |
| Q8S8H3 (DEFL149/LCR5) | AT2G28355 | 9/62 | 0.885 | 16.300 ** | 19.1% (ω = 2.182) | 7P, 12N, 13E, 19Q, 20S, 25A, 26E, 28D, 36G, 41Y, 46P, 53K, 55Y |
| Q9T0E3 (DEFL151/LCR17) | AT4G11760 | 9/84 | 0.825 | 35.276 *** | 21.0% (ω = 2.063) | 4Q, 6P, 8T, 21S, 24A, 25S, 34S, 35E, 36L, 38A, 45S, 53R, 57T, 58Q, 73R, 75W, 77K, 78S, 80G, 82P, 83Y |
| Q9ZUL8 (DEFL10/LCR72) | AT2G02140 | 10/56 | 0.433 | 7.191 * | 9.9% (ω = 1.962) | 16N, 30K |
| P82644 (DEFL231/SCRL25) | AT4G32714 | 10/106 | 0.804 | 18.579 *** | 10.8% (ω = 2.423) | 32K, 33E, 47F, 66R, 79H, 86D, 90R, 96G |
| P82621 (DEFL226/SCRL2) | AT1G65113 | 9/65 | 0.860 | 29.623 *** | 5.9% (ω = 4.293) | 11P, 22D, 31M, 47R, 53Y, 56E |
| P82622 (DEFL228/SCRL3) | AT1G08695 | 10/86 | 0.887 | 39.072 *** | 11.2% (ω = 2.687) | 10N, 17L, 20M, 22T, 24K, 35A, 47N, 49R, 50F, 54L, 56Q, 59T, 61A, 62T, 63Q, 64I, 65S, 67S, 71Q, 82R |
| A8MR88 (PCP-Bγ/ESFL8) | AT2G16535 | 9/72 | 1.061 | 34.833 *** | 26.3% (ω = 2.147) | 5S, 6V, 7E, 9S, 11A, 17T, 18L, 21S, 22V, 23N, 25D, 26N, 27K, 29D, 30R, 31N, 32L, 33T, 38L, 41A, 42K, 43S, 44K, 46R, 49S, 52S, 55V, 57D, 59K, 60D |
| A8MQY8 (PCP-Bδ/ESFL9) | AT2G16505 | 10/85 | 1.357 | 74.217 *** | 22.0% (ω = 2.975) | 14E, 18K, 19S, 20V, 21E, 22S, 23N, 24K, 25A, 26M, 28P, 29V, 31M, 32P, 33V, 34N, 37N, 38N, 39K, 40D, 43K, 44K, 45L, 46T, 48A, 51I, 52G, 53A, 54N, 56P, 57R, 58N, 59R, 62N, 63S, 64R, 65S, 66Q, 68T, 69A, 70D, 73L |
| Q9C9N7 (LTP2.5) | AT1G66850 | 10/81 | 0.230 | 2.047 | 0.5% (ω = 3.321) | NA |
| Q2V3C1 (LTP1.9) | AT4G33355 | 10/97 | 0.405 | 5.103 | 5.8% (ω = 1.876) | NA |
| Q8LFM2 (GASA10) | AT5G59845 | 10/64 | 0.196 | 0.160 | 16.1% (ω = 1.015) | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Lau, Y.-L.; Fan, L.; Bosch, M.; Doughty, J. Pollen Coat Proteomes of Arabidopsis thaliana, Arabidopsis lyrata, and Brassica oleracea Reveal Remarkable Diversity of Small Cysteine-Rich Proteins at the Pollen-Stigma Interface. Biomolecules 2023, 13, 157. https://doi.org/10.3390/biom13010157
Wang L, Lau Y-L, Fan L, Bosch M, Doughty J. Pollen Coat Proteomes of Arabidopsis thaliana, Arabidopsis lyrata, and Brassica oleracea Reveal Remarkable Diversity of Small Cysteine-Rich Proteins at the Pollen-Stigma Interface. Biomolecules. 2023; 13(1):157. https://doi.org/10.3390/biom13010157
Chicago/Turabian StyleWang, Ludi, Yui-Leung Lau, Lian Fan, Maurice Bosch, and James Doughty. 2023. "Pollen Coat Proteomes of Arabidopsis thaliana, Arabidopsis lyrata, and Brassica oleracea Reveal Remarkable Diversity of Small Cysteine-Rich Proteins at the Pollen-Stigma Interface" Biomolecules 13, no. 1: 157. https://doi.org/10.3390/biom13010157