
Identification of Chalcone Isomerase Family Genes and Roles of CnCHI4 in Flavonoid Metabolism in Camellia nitidissima

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of CHI Family Genes in C. nitidissima
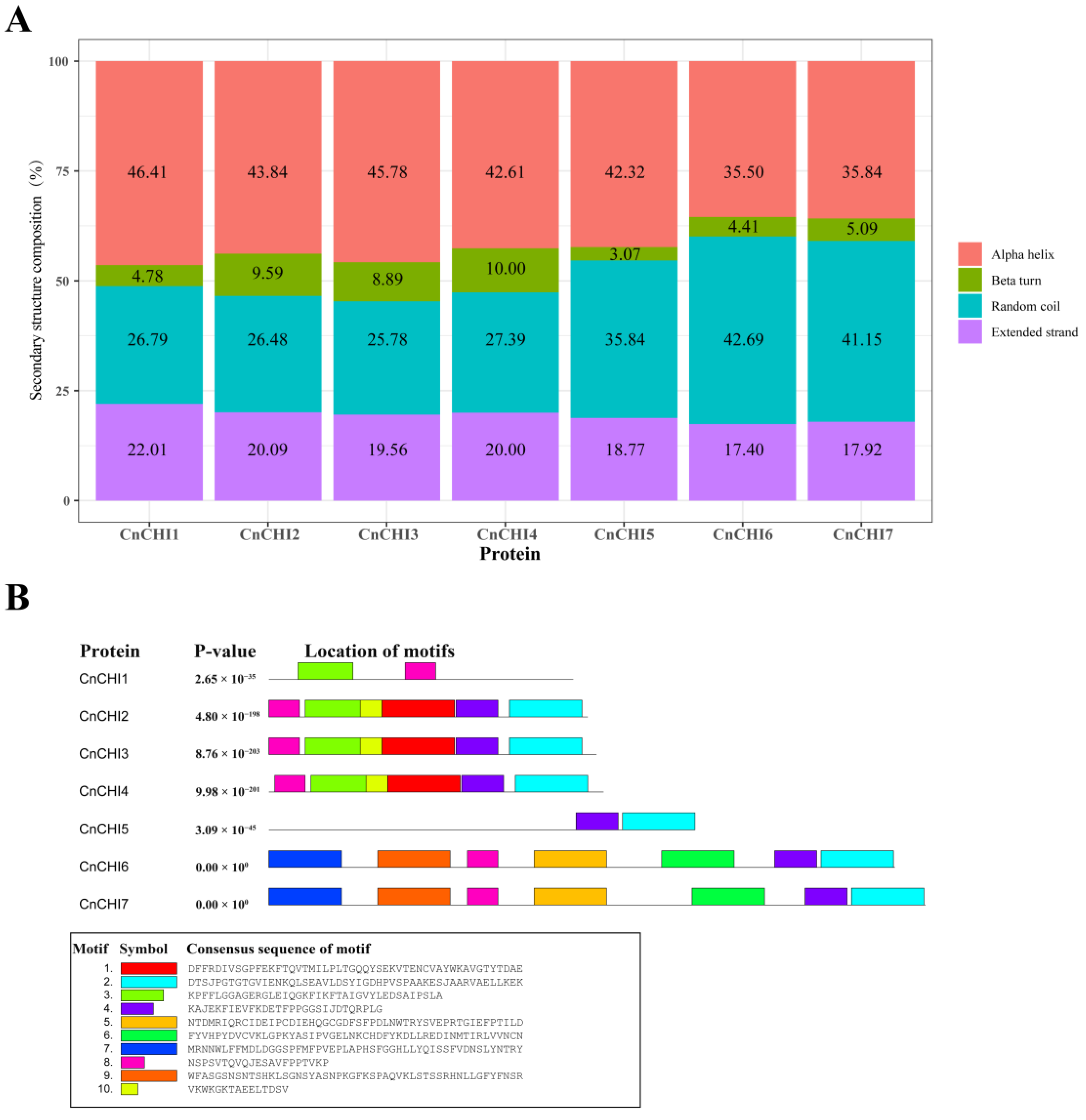
2.2. Structural and Biochemical Analyses of the CHI Proteins of C. nitidissima
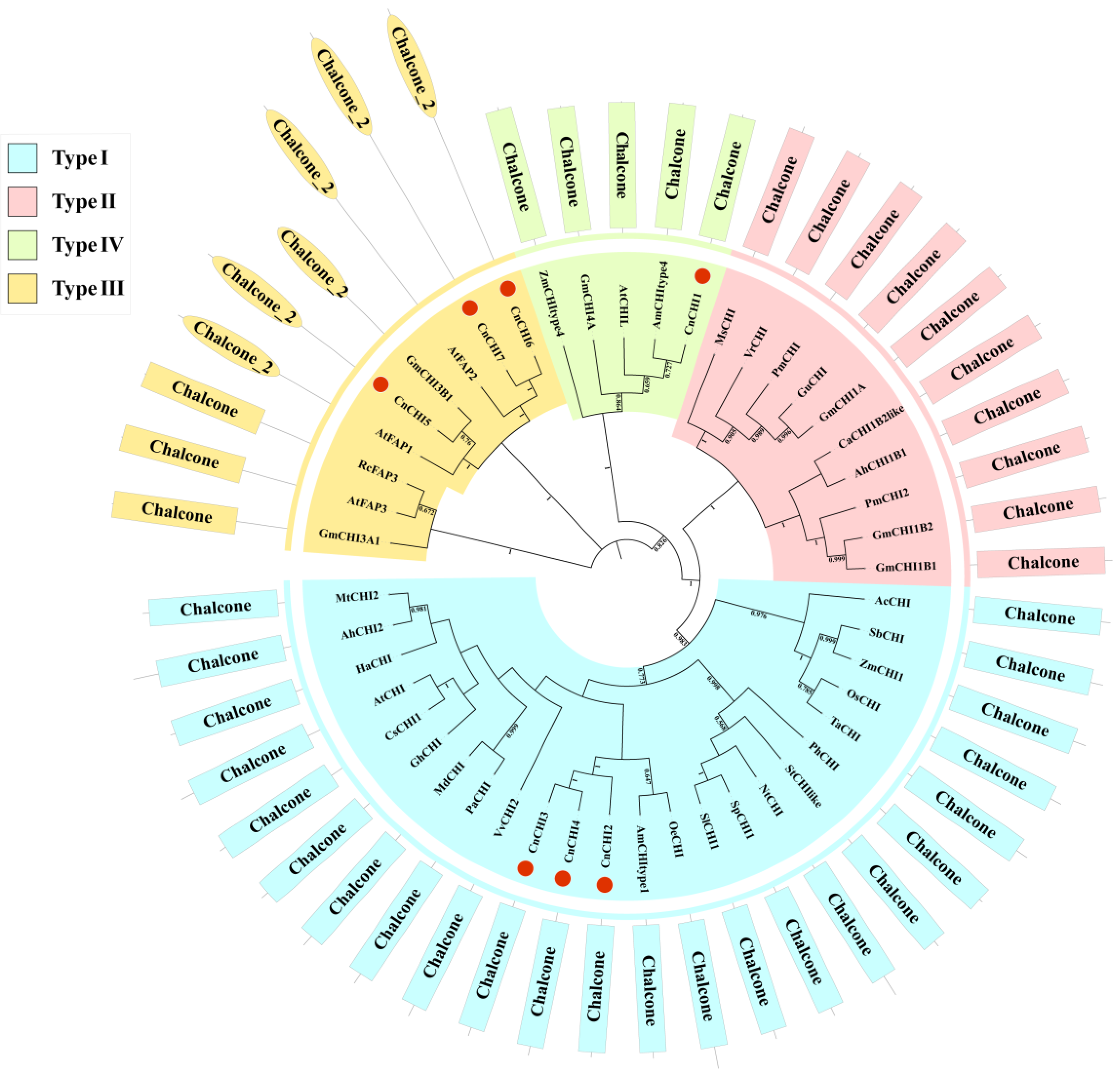
2.3. Phylogenetic Analysis and Multiple Sequence Alignment of CnCHI Proteins
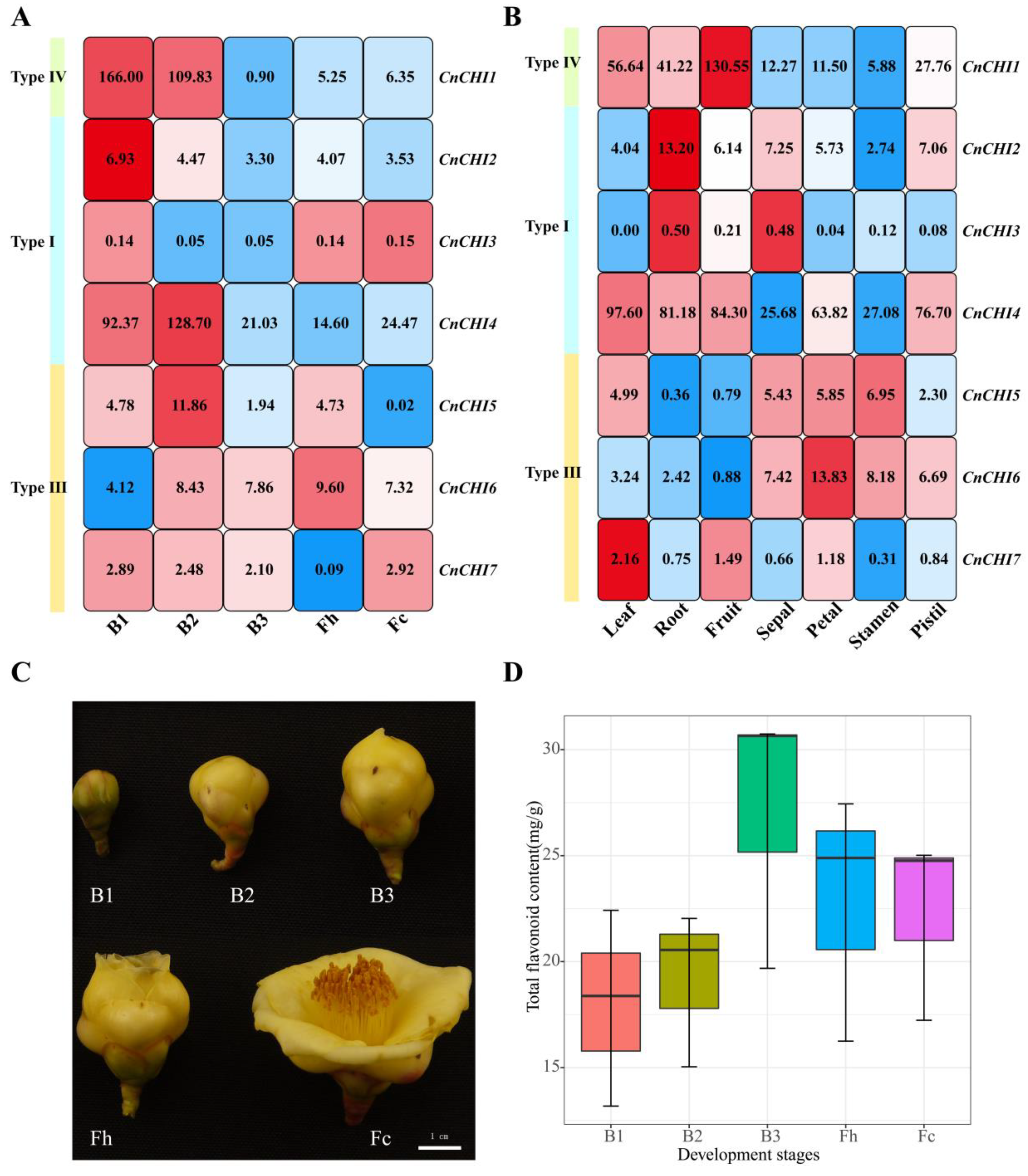
2.4. Plant Materials and Sample Preparation
2.5. Determination of Total Flavonoid Content (TFC)
2.6. Genes Isolation
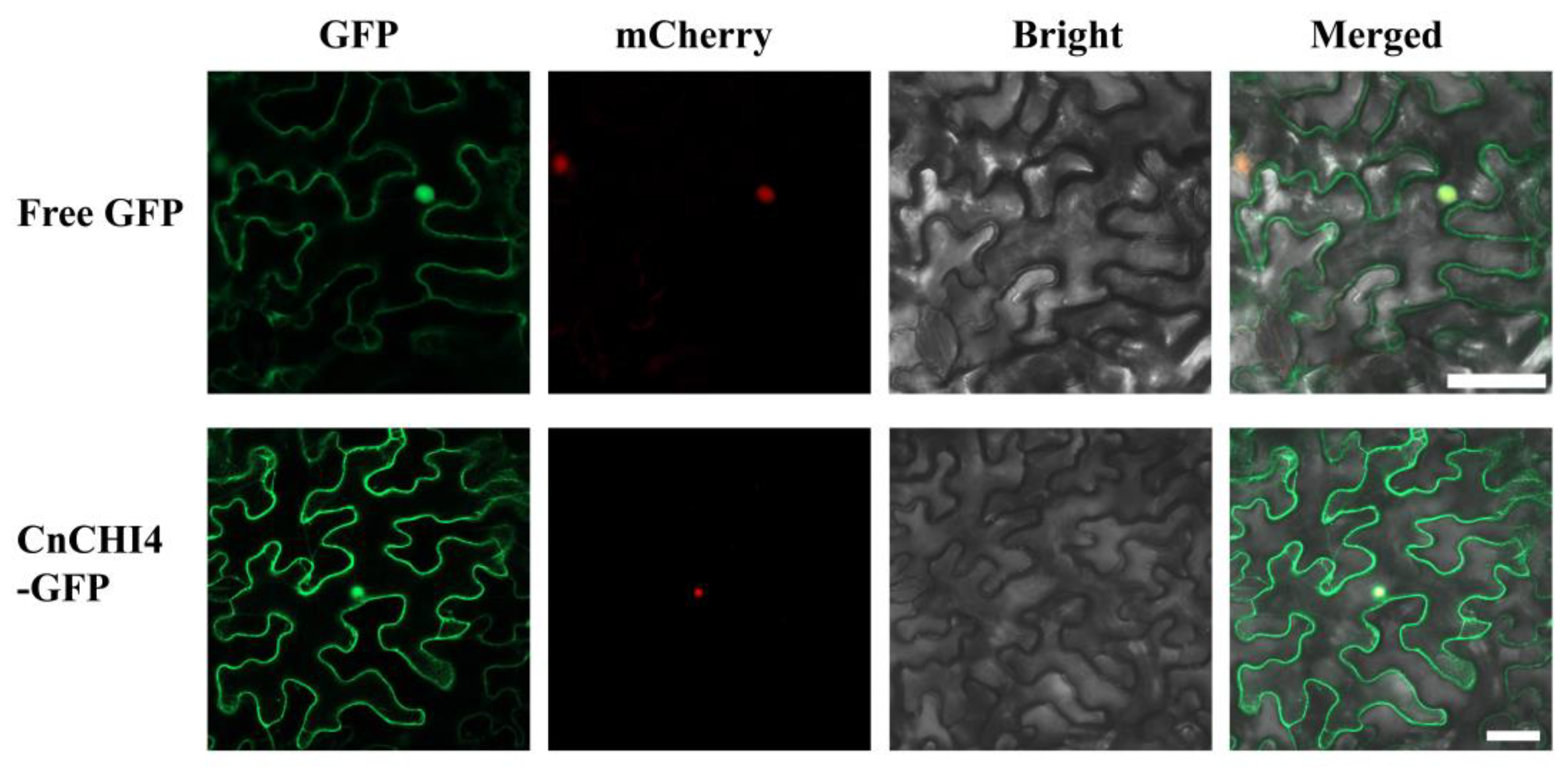
2.7. Analysis of Subcellular Localization
2.8. CnCHI4 Overexpression in N. tabacum
2.9. Transient Overexpression of CnCHI4 in C. nitidissima
2.10. qRT-PCR Analysis
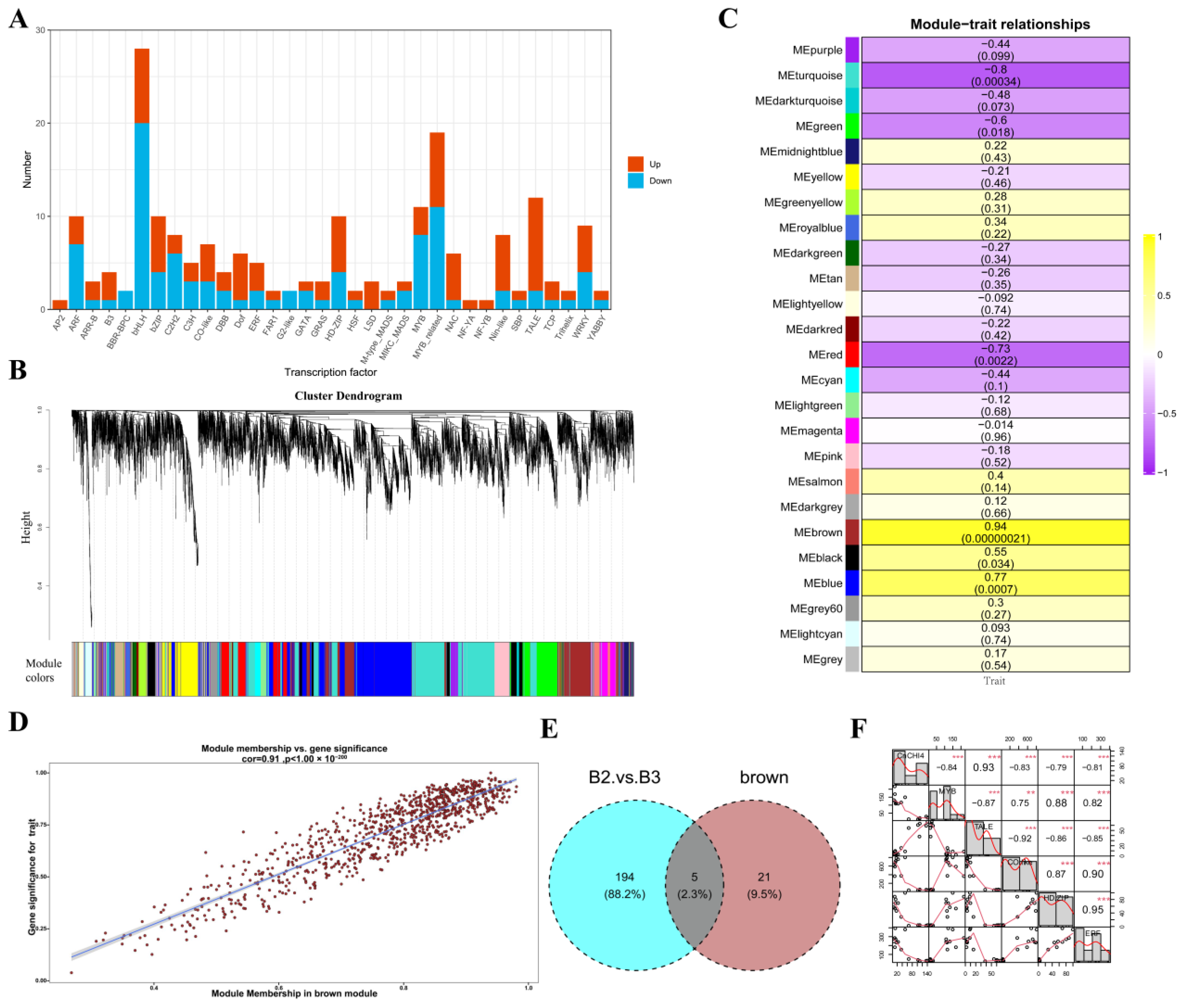
2.11. Weighted Gene Co-Expression Network Analysis (WGCNA) and Identification of Differentially Expressed Genes (DEGs)
2.12. Analyses of Promoter Clones and Cis-Elements
2.13. Dual-Luciferase Assay
2.14. Yeast One-Hybrid Assay
2.15. Statistical Analyses
3. Results
3.1. Identification and Characterization of CnCHI Proteins
3.2. Phylogenetic Relationships, Multiple Sequence Alignment, and Classification of CnCHIs
3.3. Analysis of CnCHI Expression and Measurement of TFC
3.4. CnCHI4 Is Localized in the Cytoplasm and Nucleus
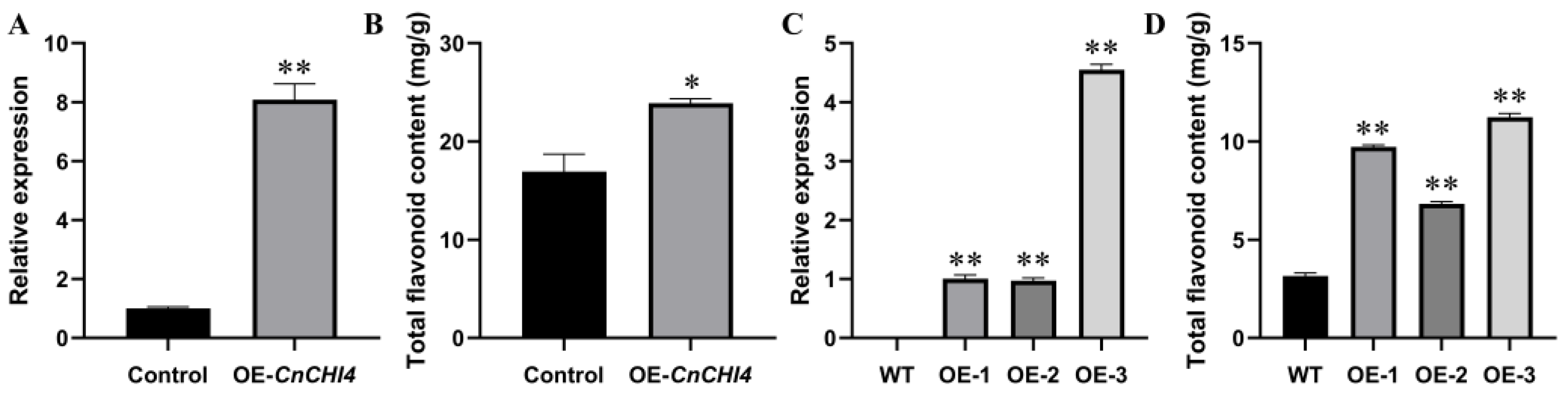
3.5. Overexpression of CnCHI4 in C. nitidissma and N. tabacum
3.6. Identification of Key Regulatory Factors of CnCHI4
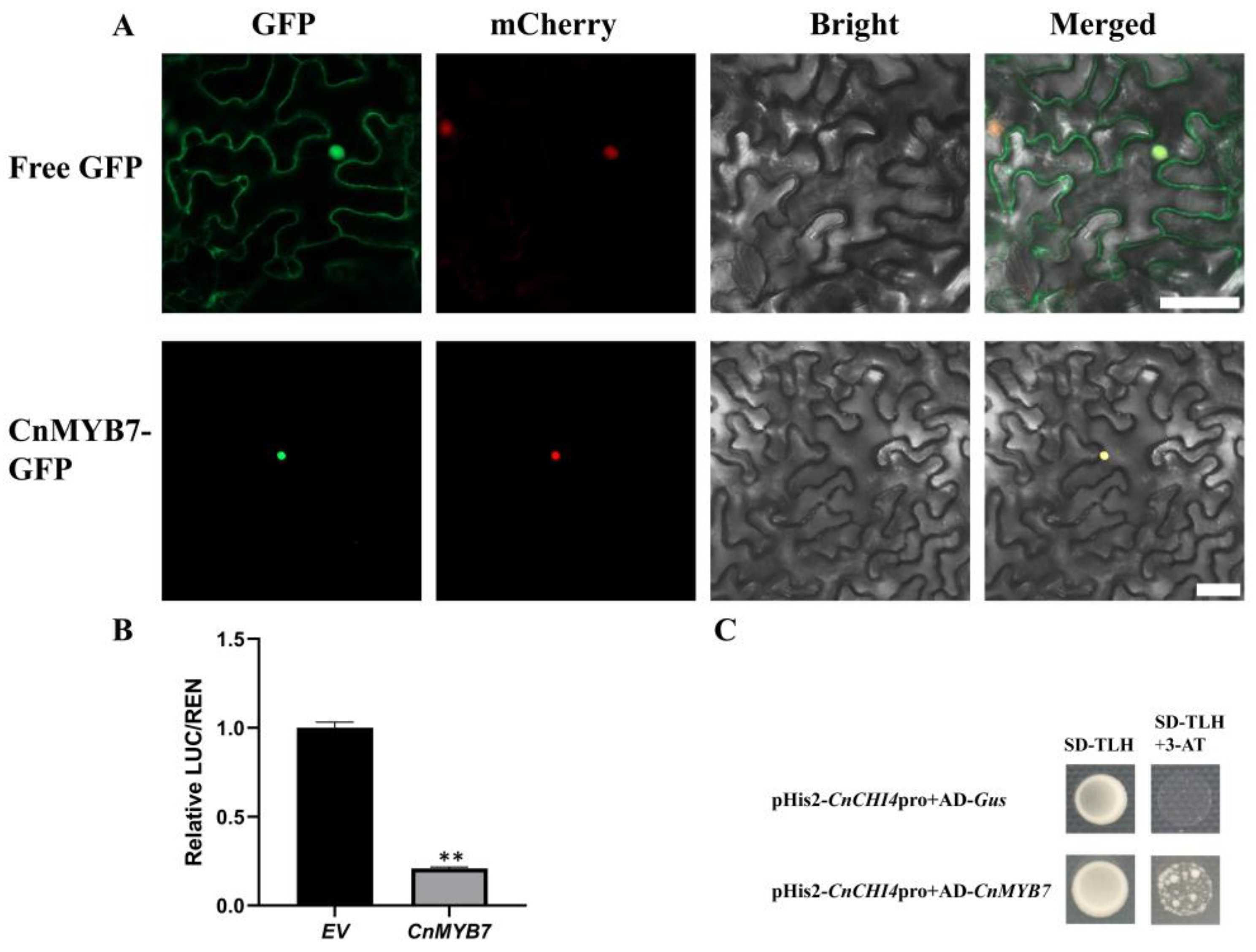
3.7. CnMYB7 was Localized in the Nucleus
3.8. CnMYB7 Bound and Inhibited the CnCHI4 Promoter
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, A.; Zhang, X.; Han, X.; Zhang, Y.; Gao, S.; Liu, C.; Lou, H. Identification of chalcone isomerase in the basal land plants reveals an ancient evolution of enzymatic cyclization activity for synthesis of flavonoids. New Phytol. 2018, 217, 909–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Dobritsa, A.A.; Lei, Z.; Nishikawa, S.-I.; Urbanczyk-Wochniak, E. LAP5 and LAP6 encode anther-specific proteins with similarity to chalcone synthase essential for pollen exine development in Arabidopsis. Plant Physiol. 2010, 153, 937–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Guo, H.; Song, X.; Zhang, D.; Long, Y.; Xing, Z. Adaptive Evolution of Chalcone Isomerase Superfamily in Fagaceae. Biochem. Genet. 2021, 59, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Hosseinzade, A.; Sadeghi, O.; Biregani, A.N. Immunomodulatory Effects of Flavonoids: Possible Induction of T CD4+ Regulatory Cells Through Suppression of mTOR Pathway Signaling Activity. Front. Immunol. 2019, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Chao, N.; Wang, R.; Hou, C.; Yu, T. Functional characterization of two chalcone isomerase (CHI) revealing their responsibility for anthocyanins accumulation in mulberry. Plant Physiol. Biochem. 2021, 161, 65–73. [Google Scholar] [CrossRef]
- Waki, T.; Mameda, R.; Nakano, T.; Yamada, S. A conserved strategy of chalcone isomerase-like protein to rectify promiscuous chalcone synthase specificity. Nat. Commun. 2020, 11, 870. [Google Scholar] [CrossRef]
- Sun, W.; Shen, H.; Xu, H.; Tang, X.; Tang, M.; Ju, Z.; Yi, Y. Chalcone Isomerase a Key Enzyme for Anthocyanin Biosynthesis in Ophiorrhiza japonica. Front. Plant Sci. 2019, 10, 865. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Jin, Z.; Qu, W.; Zhao, D. Cloning of a cDNA encoding the Saussurea medusa chalcone isomerase and its expression in transgenic tobacco. Plant Physiol. Biochem. 2006, 44, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Przysiecka, Ł.; Książkiewicz, M.; Wolko, B.; Naganowska, B. Structure, expression profile and phylogenetic inference of chalcone isomerase-like genes from the narrow-leafed lupin (Lupinus angustifolius L.) genome. Front. Plant Sci. 2015, 6, 268. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Gao, Y.; Liu, F.; He, B.; Jia, X.; Meng, F.; Zhang, H.; Guo, M. Integrating molecular characterization and metabolites profile revealed CtCHI1′s significant role in Carthamus tinctorius L. BMC Plant Biol. 2019, 19, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, J.-Y.; Kasthuri, G.M.; Park, J.-Y.; Kang, W.-J. An overexpression of chalcone reductase of Pueraria montana var. lobata alters biosynthesis of anthocyanin and 5’-deoxyflavonoids in transgenic tobacco. Biochem. Biophys. Res. Commun. 2003, 303, 326–331. [Google Scholar]
- Kim, S.; Jones, R.; Yoo, K.S.; Pike, L.M. Gold color in onions (Allium cepa): A natural mutation of the chalcone isomerase gene resulting in a premature stop codon. Mol. Genet. Genom. 2004, 272, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; Ric, D.V.C.H.; Van Tunen, A.J.; Verhoeyen, M.E. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Ngaki, M.N.; Louie, G.V.; Philippe, R.N.; Manning, G. Evolution of the chalcone-isomerase fold from fatty-acid binding to stereospecific catalysis. Nature 2012, 485, 530–533. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Takagi, K.; Fukuchi-Mizutani, M.; Ishiguro, K. A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation. Plant J. 2014, 78, 294–304. [Google Scholar] [CrossRef]
- Jiang, W.; Yin, Q.; Wu, R.; Zheng, G.; Liu, J.; Dixon, R.A.; Pang, Y. Role of a chalcone isomerase-like protein in flavonoid biosynthesis in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 7165–7179. [Google Scholar] [CrossRef] [Green Version]
- Ban, Z.; Qin, H.; Mitchell, A.J. Noncatalytic chalcone isomerase-fold proteins in Humulus lupulus are auxiliary components in prenylated flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E5223–E5232. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Wang, X.; Zhang, X. An apple MYB transcription factor regulates cold tolerance and anthocyanin accumulation and undergoes MIEL1-mediated degradation. Plant Biotechnol. J. 2020, 18, 337–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W.; Wang, H.; Li, M. The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant Biotechnol. J. 2016, 14, 2120–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrtens, F.; Kranz, H.; Bednarek, P. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Pei, X.; Zhang, H.; Li, X. MYB-Mediated Regulation of Anthocyanin Biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Han, D.; Huang, B.; Li, Y.; Dang, Q. The MdAP2-34 modulates flavonoid accumulation in apple (Malus domestica Borkh.) by regulating MdF3′H. Postharvest Biol. Technol. 2022, 192, 111994. [Google Scholar] [CrossRef]
- He, D.; Li, X.; Sai, X.; Wang, L.; Li, S.; Xu, Y. Camellia nitidissima C.W. Chi: A review of botany, chemistry, and pharmacology. Phytochem. Rev. 2017, 17, 327–349. [Google Scholar] [CrossRef]
- Hou, X.; Du, H.; Yang, R.; Qi, J. The antitumor activity screening of chemical constituents from Camellia nitidissima Chi. Int. J. Mol. Med. 2018, 41, 2793–2801. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2018, 38, 107316. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Zhu, Y.; Ni, S.; Chen, J.; Feng, X.; Zhang, Y.; Li, S.; Zhu, H.; Wen, Y. De novo assembly of the Camellia nitidissima transcriptome reveals key genes of flower pigment biosynthesis. Front. Plant Sci. 2017, 8, 1545. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, Q.; Chen, Y.; Zhu, Y.; Zhou, X.; Li, B. Full-length transcriptome sequencing provides insights into flavonoid biosynthesis in Camellia nitidissima Petals. Gene 2023, 850, 146924. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L. Study on Flower Color Formation Metabolism Mechanism and Key Genes Function of Camellia nitidissima. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2021. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Horsch, R.; Fry, J.; Hoffmann, N.; Wallroth, M.; Eichholtz, D.; Rogers, S.; Fraley, R. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yu, S.; Feng, Y.; Mo, R.; Wang, K.; Fan, M.; Fan, Z.; Yin, H.; Li, J.; Li, X. Comparative Transcriptome and Pigment Analyses Reveal Changes in Gene Expression Associated with Flavonol Metabolism in Yellow Camellia. Forests 2022, 13, 1094. [Google Scholar] [CrossRef]
- Zhu, J.; Zhao, W.; Li, R. Identification and Characterization of Chalcone Isomerase Genes Involved in Flavonoid Production in Dracaena cambodiana. Front. Plant Sci. 2021, 12, 616396. [Google Scholar] [CrossRef]
- Chen, W.H.; Li, P.F.; Chen, M.K.; Lee, Y.I.; Yang, C.H. Forever Young Flower Negatively Regulates Ethylene Response DNA-Binding Factors by Activating an Ethylene-Responsive Factor to Control Arabidopsis Floral Organ Senescence and Abscission. Plant Physiol. 2015, 168, 1666–1683. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef] [Green Version]
- Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure and mechanism of the evolutionarily unique plant enzyme chalcone isomerase. Nat. Struct. Biol. 2000, 7, 786–791. [Google Scholar]
- Lam, P.Y.; Wang, L.; Lui, A.C.W. Deficiency in flavonoid biosynthesis genes CHS, CHI, and CHIL alters rice flavonoid and lignin profiles. Plant Physiol. 2022, 188, 1993–2011. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, Y.S.; Kim, Y.; Thwe, A.A. Molecular characterization of flavonoid biosynthetic genes and accumulation of baicalin, baicalein, and wogonin in plant and hairy root of Scutellaria lateriflora. Saudi J. Biol. Sci. 2018, 25, 1639–1647. [Google Scholar] [CrossRef] [Green Version]
- Lambais, M.R.; Mehdy, M.C. Suppression of endochitinase, β-1,3-endoglucanase, and chalcone isomerase expression in bean vesicular-arbuscular mycorrhizal roots under different soil phosphate conditions. Mol. Plant-Microbe Interact. 1993, 6, 75. [Google Scholar] [CrossRef]
- Luo, X.; Bai, Y.; Gao, F.; Li, C.; Wu, Q. Gene cloning and expression level of chalcone isomerase during florescence and content of flavonoids in Fagopyrum dibotrys. Chin. Tradit. Herb. Drugs 2013, 44, 1481–1485. [Google Scholar]
- Nishihara, M.; Nakatsuka, T.; Yamamura, S. Flavonoid components and flower color change in transgenic tobacco plants by suppression of chalcone isomerase gene. FEBS Lett. 2005, 579, 6074–6078. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Roberts, K.; Martin, C. Temporal and spatial control of expression of anthocyanin biosynthetic genes in developing flowers of Antirrhinum majus. Plant J. 2010, 2, 425–434. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, X.; Gong, Q.; Cao, J. Three AP2/ERF family members modulate flavonoid synthesis by regulating type IV chalcone isomerase in citrus. Plant Biotechnol. J. 2021, 19, 671–688. [Google Scholar] [CrossRef]
- Winkel, J.B.S. Metabolic Channeling in Plants. Annu. Rev. Plant Biol. 2004, 55, 85–107. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Zhang, K.; Sun, Z.; Yan, M. LNK1 and LNK2 Corepressors Interact with the MYB3 Transcription Factor in Phenylpropanoid Biosynthesis. Plant Physiol. 2017, 174, 1348–1358. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D. The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lu, W.; Ran, L.; Dou, L. R2R3-MYB transcription factor MYB6 promotes anthocyanin and proanthocyanidin biosynthesis but inhibits secondary cell wall formation in Populus tomentosa. Plant J. 2019, 99, 733–751. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Khaldun, A.B.M.; Lv, H.; Du, L. Isolation and functional characterization of a R2R3-MYB regulator of the anthocyanin biosynthetic pathway from Epimedium sagittatum. Plant Cell Rep. 2016, 35, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Osbourn, A.; Ma, P. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Misra, P.; Trivedi, P.K. Constitutive expression of Arabidopsis MYB transcription factor, AtMYB11, in tobacco modulates flavonoid biosynthesis in favor of flavonol accumulation. Plant Cell Rep. 2015, 34, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Misra, P.; Bhambhani, S.; Bhatia, C. Expression of Arabidopsis MYB transcription factor, AtMYB111, in tobacco requires light to modulate flavonol content. Sci. Rep. 2014, 4, 5018. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene ID | Amino Acid Length (aa) | pI | Molecular Weight (KDa) | Grand Average of Hydropathicity | Instability Index | Signal Peptide |
|---|---|---|---|---|---|---|---|
| CnCHI1 | F01.PB9454 | 209 | 4.83 | 23,193.41 | −0.148 | 41.12 | no |
| CnCHI2 | F01.PB6499 | 219 | 5.39 | 23,711.19 | −0.148 | 41.12 | no |
| CnCHI3 | F01.PB58445 | 225 | 5.25 | 24,264.77 | −0.097 | 42.96 | no |
| CnCHI4 | F01.PB37720 | 230 | 5.25 | 24,735.25 | −0.116 | 47.73 | no |
| CnCHI5 | F01.PB54606 | 293 | 8.73 | 32,154.61 | −0.189 | 48.40 | no |
| CnCHI6 | F01.PB71691 | 431 | 8.6 | 47,966.94 | −0.097 | 43.83 | no |
| CnCHI7 | F01.PB53164 | 452 | 8.6 | 50,117.33 | −0.093 | 42.57 | no |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Li, J.; Peng, T.; Ni, S.; Feng, Y.; Wang, Q.; Wang, M.; Chu, X.; Fan, Z.; Li, X.; et al. Identification of Chalcone Isomerase Family Genes and Roles of CnCHI4 in Flavonoid Metabolism in Camellia nitidissima. Biomolecules 2023, 13, 41. https://doi.org/10.3390/biom13010041
Yu S, Li J, Peng T, Ni S, Feng Y, Wang Q, Wang M, Chu X, Fan Z, Li X, et al. Identification of Chalcone Isomerase Family Genes and Roles of CnCHI4 in Flavonoid Metabolism in Camellia nitidissima. Biomolecules. 2023; 13(1):41. https://doi.org/10.3390/biom13010041
Chicago/Turabian StyleYu, Suhang, Jiyuan Li, Ting Peng, Sui Ni, Yi Feng, Qiushi Wang, Minyan Wang, Xian Chu, Zhengqi Fan, Xinlei Li, and et al. 2023. "Identification of Chalcone Isomerase Family Genes and Roles of CnCHI4 in Flavonoid Metabolism in Camellia nitidissima" Biomolecules 13, no. 1: 41. https://doi.org/10.3390/biom13010041