
Ribose Intake as Food Integrator: Is It a Really Convenient Practice?
by
 , , , and
, , , and
Roberta Moschini
1,2 ,
,
Francesco Balestri
1,2,
Mario Cappiello
1,2,
Giovanni Signore
1,2,
Umberto Mura
1,* and
Antonella Del-Corso
1,2 1
Biochemistry Unit, Department of Biology, University of Pisa, Via San Zeno, 51, 56127 Pisa, Italy
2
Interdepartmental Research Center Nutrafood “Nutraceuticals and Food for Health”, University of Pisa, 56124 Pisa, Italy
*
Author to whom correspondence should be addressed.
Biomolecules 2022, 12(12), 1775; https://doi.org/10.3390/biom12121775
Submission received: 21 October 2022
/
Revised: 7 November 2022
/
Accepted: 22 November 2022
/
Published: 29 November 2022
Abstract
:Reports concerning the beneficial effects of D-ribose administration in cardiovascular and muscle stressful conditions has led to suggestions for the use of ribose as an energizing food supplement for healthy people. However, this practice still presents too many critical issues, suggesting that caution is needed. In fact, there are many possible negative effects of this sugar that we believe are underestimated, if not neglected, by the literature supporting the presentation of the product to the market. Here, the risks deriving from the use of free ribose as ATP source, forcing ribose-5-phosphate to enter into the pentose phosphate pathway, is emphasized. On the basis of the remarkable glycation capacity of ribose, the easily predictable cytotoxic effect of the molecule is also highlighted.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Nucleotides, besides being relevant precursor in the nucleic acid (NA) synthesis, represent the most advanced goal of metabolic evolution in terms of ready to use chemical energy storage; they contain two phosphoanhydridic bonds, whose cleavage, under proper catalytic action, may provide the driving force of endergonic reactions needed for cell life. We can mention, just as an example, the UTP role in the generation of UDP-glucose, the specific precursor for the glucose storage as glycogen or the extraordinary concerted function of GTP as energy donor in the ribosomal machinery for protein synthesis and, lastly, the ubiquitous ATP involvement as energy supplier in innumerable and so different energy requiring processes. Indeed, ATP is synonymous of cell energy and the phosphorylation status of the adenylic nucleotides (i.e., AMP, ADP, and ATP) is taken as an index (i.e., the energy charge) of the energetic status of the cell. Thus, the attention of researchers (as well as entrepreneurs) in finding new strategies to favor the replenishment of the nucleotides in general and, in particular, of adenylyl nucleotides (both in terms of their levels and of their highest phosphorylation status) is understandable. We are so used to seeing the ribose scaffold in relevant biological molecules as nucleotides and nucleic acids that not thinking that D-ribose enters as a precursor of their synthesis will sound almost inconceivable. However, it seems that such an apparent “ready to use” precursor for nucleotide synthesis is mainly disregarded in the metabolic implant, being glucose, through its phosphorylation at C6, the preferred precursor of the ribofuranosidic scaffold of nucleotides [1]. Nevertheless, as underlined below, D-ribose intake has been proposed as a strategic approach to increase levels of ATP. If we simply consider the widespread “commercial propaganda” about the beneficial effects of D-ribose in the human metabolism, we may be convinced that this metabolite is essential for an energy-filled, healthy, and enjoyable life. Emphasis on this aspect appears in many commercial advertisements, highlighting the fact that ribose, being part of nucleotides, has the potential to enhance ATP availability. Thus, ribose intake is suggested to energize healthy subjects undergoing physical exercise. However, with a basic insight into metabolic processes, it is not difficult to challenge the commercial presentation of this “fantastic” food integrator, even if allegedly supported by literature references [2,3,4,5,6]. The main concern arising from many of these presentations is related to the ambiguity on the goal of the proposed treatment and the inadequacy in presenting what we can define as the “unfriendly features” of ribose. In fact, it is rather ambiguous whether ribose intake should improve the energy charge or the nucleotide pool level or both. On the other hand, the possible adverse effects linked to the reactivity of the ribose molecule are essentially neglected. Thus, we want to point out that, as presented, the energy injection that should occur in healthy active people (children, runners, body builders, sportsmen), deriving from D-ribose, doesn’t have, yet, any strong scientific base allowing the proposal and promotion for indiscriminate consumption of the sugar. Conversely, the risk of damage arising from circulating ribose is firmly standing. As potential customers rather than as scientists, we were surprisingly worried in learning of the actions undertaken by ribose Producers at the level of European Commission to advocate for the integration of this molecule in the formulations of a variety of edible products, from energy bars and drinks, to biscuits, flavored drinks, fruit juice, vegetable juice, and yogurt [7]. We do not know whether such actions are still proceeding, but certainly the topic is worth of attention by EU governments. Thus, in this review, some considerations are made on what is loudly claimed being advantageous, and what may be subtly dangerous regarding a ribose dietary intake by healthy people.
2. Ribose Metabolism and Nucleotides Synthesis
The preferential choice of the ribose scaffold as a building block for biomolecules is self-evident simply by observing its recurring presence in molecules, such as nucleotides and nucleic acids, which are so relevant for cell life. The reliability of molecules is considered as a factor possibly influencing their recruitment as privileged precursors of metabolic pathways. This is the indubitable case of the pyranosidic structure of glucose, whose reliability [8] but also its minimal, even though possible, glycating activity [9], led glucose to be chosen as the privileged precursor of bioenergetics pathways. The opposite is the case, following the same reasoning approach, for the use of ribose as metabolic precursor for nucleotide synthesis. Due to the structural unreliability of ribose and due to its the remarkable glycation ability, this sugar should have appeared ab initio too risky in supporting a relevant goal as the nucleotide synthesis. Then, the pentose phosphate pathway (PPP), jointly to its function of replenishment of reducing equivalents (i.e., NADPH formation), is widely recognized as the mainstream of ribose-5-phosphate (R5P) synthesis [10], through the use of glucose-6 phosphate as precursor. This unavoidable precursor of de novo nucleotide synthesis is an intermediate of the reversible reaction network of PPP. These reactions modulate the overall PPP by recycling the C3 to C6 glucose-derived carbon atoms into the main glycolytic/gluconeogenic flux. Thus, due to their reversibility, they represent the access door of glycolysis intermediates (i.e., glyceraldehyde-3-phosphate and fructose-6-phosphate) to R5P generation [11].
R5P would also directly enter PPP by the ATP dependent phosphorylation of D-ribose catalyzed by ribokinase (RK). This enzyme, which belongs to the PfkB family of carbohydrate kinases [12] has been characterized in human [13,14] and displays most of the features of the E. coli RK in terms of phosphate and metal ion activation. Nevertheless, the human enzyme exhibits a reduced efficiency with respect to bacterial RK, with KM values for D-ribose of at least one order of magnitude higher (approximately 2 mM versus 0.2 mM) [15,16]. Furthermore, a substrate inhibition by ribose is observed, which may limit R5P formation [13]. Much more evident is the lack of efficiency of RK when compared to adenosine kinase (AdoK), another member of the PfkB carbohydrate kinase family. This enzyme shares with RK structural elements, regulatory properties, and the ability to recognize as substrate the C5-hydroxyl group of the ribofuranoside scaffold (here linked to the purine base). The KM value of the human AdoK for adenosine is in the range of µM or sub-µM levels (0.1 to 3 µM), from two to three order of magnitude lower than of RK for D-ribose [17,18,19]. Besides possible peculiar differences between these strongly related family members, the poor effectiveness of RK may certainly be ascribed to the unreliability of the D-ribose, mostly present in solution in its pyranosidic form (80%) [20,21], which is less adequate as substrate for RK action [1]. In any case, R5P may eventually be formed upon RK action. This is the route invoked by ribose “supporters” to rationalize the beneficial features of exogenously supplemented ribose entering the glycolytic/gluconeogenic pathway and, most importantly, the nucleotides synthesis. In this regard it is worth noting that the need of this ATP-dependent activation step for ribose to enter metabolic routes excludes any possible advantage of ribose with respect to glucose as ergogenic nutrient. On the other hand, with some exceptions, ribose alone is unable to sustain cell growth [22,23,24]. Therefore, ribose would exclusively assume the possible role of precursor of nucleotides synthesis.
Now, the relevant questions are why and when this metabolic opportunity should be forced and, most importantly, its price.
3. The Claim of Ribose Intake as an Ergogenic Practice
The ergogenic effect of ribose comes out from studies in which the ability of the cell to warrant the homeostasis of ATP is somehow compromised. This is due to either a pathological status or an excessive energy expenditure. Evidence has been reported from studies on animals and humans for which ribose treatment may be helpful in cardiovascular diseases. An increase in the rate of ATP recovery in myocardium after ribose intravenous injection was observed in rats undergoing oxygen reperfusion after myocardial ischemia [25]. A similar effect was reported in a canine model for myocardial ischemia, in which the level of ATP, accounting for 50% of the pre-ischemic level, was restored after 24 h to 85% in animals subjected to ribose infusion [26]. Moreover, human subjects suffering of cardiopathic dysfunctions such as ischemia, cardiomyopathy, and hypertrophy, were beneficially affected by ribose administration [27,28,29].
D-ribose intake was successfully used to ameliorate symptoms in the case of fibromyalgia and chronic fatigue syndrome [2,30]. In a case report of a patient affected by myoadenilate deaminase deficiency, a recovery of ATP depletion was observed after ribose treatment [31]. This observation was, however, not supported by other case reports referring to the same pathological status [32].
With such evidence, the rational base of the reports claiming the beneficial effect of an external supplement of ribose generally referred to a failure of the oxidative branch of PPP, unable anymore to replenish R5P levels adequate for the de novo synthesis and “salvage” of nucleotides. This problem is very stimulating and worth being experimentally furthered to give answers to the above questions concerning why and when ribose intake may be beneficial.
The benefits that ribose may offer in a number of pathological situations has been exhaustively reviewed [33]. However, no research has been carried out regarding some problems that the ribose treatment may, at least theoretically, promote. The word “glycation”, highly expected to occur talking on ribose, does not appear all over the text. Much more attention was finally posed on this aspect in a very recent review on this topic [34].
Besides some attempts tailored to verify the potential usefulness of ribose intake to ameliorate the distress in pathological conditions, ribose treatment was widely tested to prove the advantages it could have for healthy people to ameliorate their physical performance. There is evidence that the rate enhancement, if any, of ATP resynthesis occurring after repeated physical exercise does not imply any improvement in the performance score [35,36,37,38]. However, a number of reports on healthy subjects underline the advantage of taking ribose [39,40,41,42]. It is in this literature frame that the idea to administer ribose to healthy people as energizer emerged, giving the scientific support for commercial spots to include ribose as a dietary integrator. While we will return later on this argument, we believe useful at this point, since the ribose intake is connected to the PPP, to have an insight on this pathway.
4. Getting Closer to PPP
The relevance of PPP as a source of reducing power (i.e., NADPH) and nucleotides precursor (i.e., R5P) is an assessed point for biochemistry students. However, how critical is this pathway for the proper function of cellular and organism health is probably underestimated [43].
4.1. The Oxidative Branch of PPP
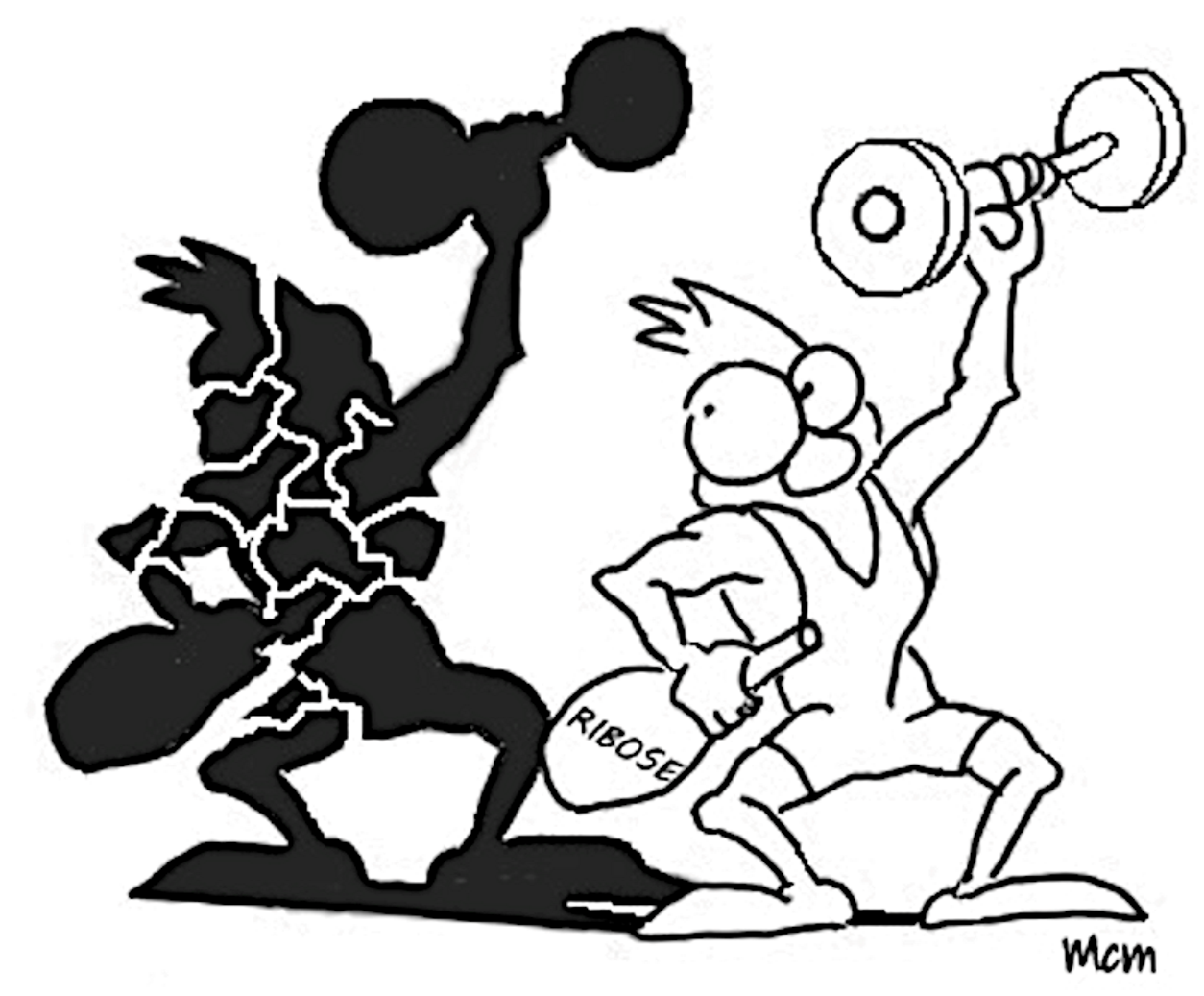
Looking at the oxidative irreversible part of PPP, apparently devoted to NADPH replenishment (Figure 1), the failure of glucose-6-phosphate dehydrogenase (G6PDH) activity, the enzyme which catalyzes the first and limiting step of the pathway, may determine dramatic pathological situations [44]. The deficiency of this enzyme derives from a genetic disorder, especially pronounced in specific world regions, characterized by several point mutations with a wide phenotypic heterogeneity [45,46].
Such a deficiency leads to a number of severe pathophysiologic states, such as neonatal jaundice [47], chronic non-spherocytic haemolytic anaemia [48], predisposition to haemolytic crisis induced by drugs or infections [49,50,51]. The pathology defined as “favism”, in which compounds present in fava beans, such as divicine and isouramil are inducers of haemolysis, is emblematic [52]. The haemolysis linked to G6PDH deficiency may also be triggered in diabetes, in myocardial infarction, or even during extreme physical exercise [53,54,55]. Cardio circulatory dysfunctions also have been related to deficiency of G6PDH activity. Also in this case, the effect was ascribed to a decrease of NADPH levels, which leads to the loss of modulation of the redox status of endothelial cells, which in turn induces ROS accumulation [56,57,58]. Alterations at level of 6-phosphogluco-δ lactone dehydrogenase (6PGLDH), which catalyses the second irreversible step of PPP, are rarer. Deficiency of this enzyme have been reported to be associated to a reduction of red blood cell resistance [59,60]. However, possibly since it is not a limiting step for G6P flux into PPP, the loss of approximately 50% to 80% in 6PGLDH activity does not determine severe consequences as it occurs for the G6PDH deficiency.
When 6-phosphogluco-δ lactone lactonase (6PGL) deficiency occurs on top of G6PDH deficiency, the situation may become more critical. In fact, in polymorphic variants of G6PDH in which no chronic hemolysis takes place, the concomitant lack of 6PGL may induce hemolysis [61]. Also, overexpression of G6PDH is associated with pathological states. This is the case, for instance, of the metabolic support offered by the over activity of the enzyme to cancer cell growth [62,63].
4.2. The Non-Oxidative, Reversible Branch of PPP
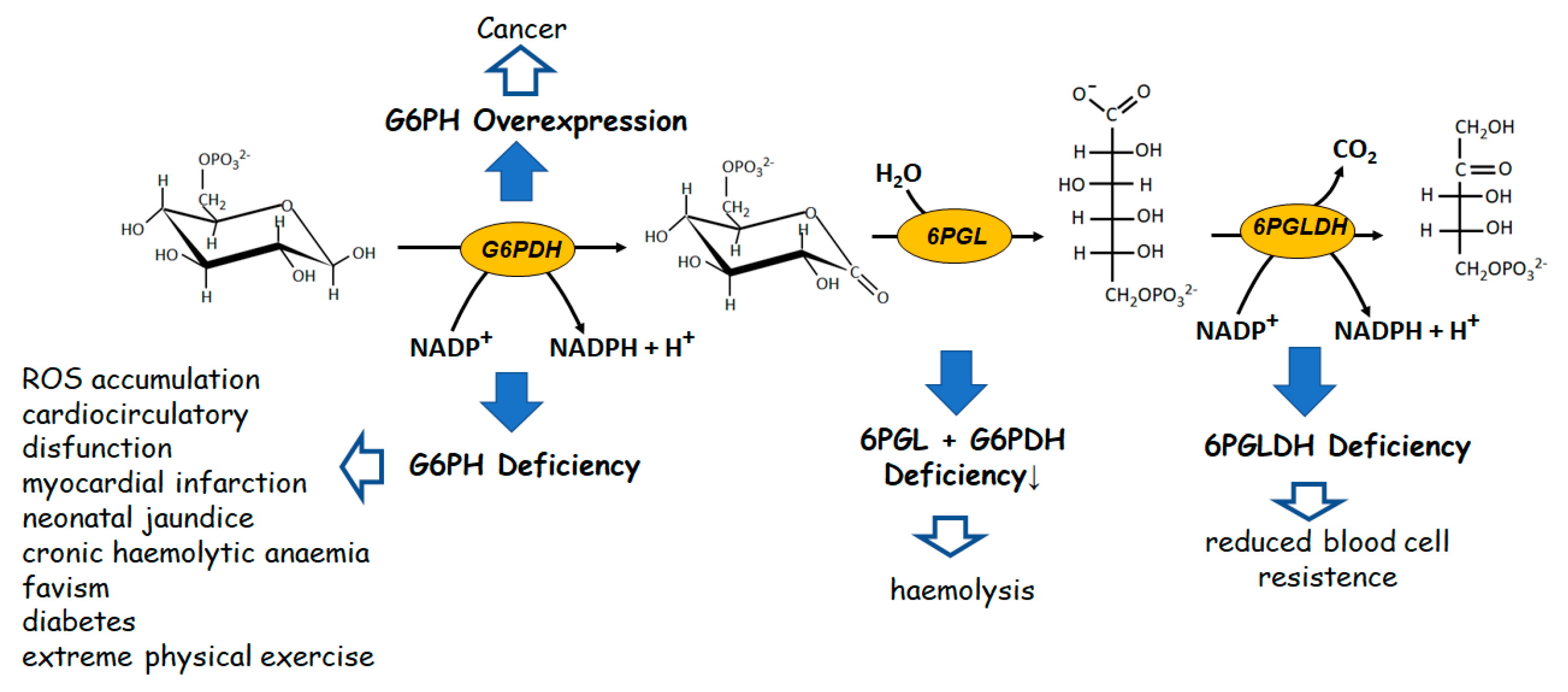
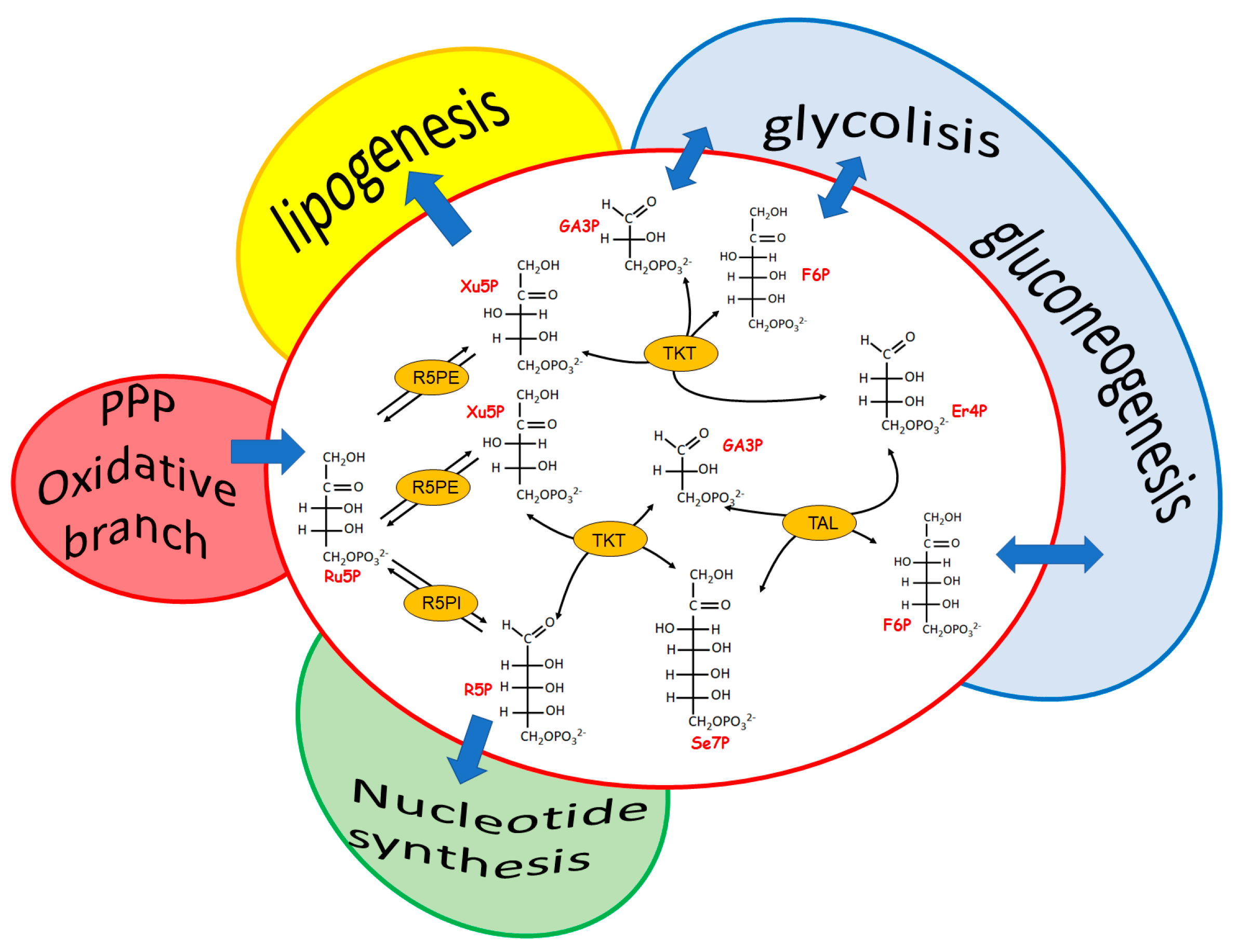
The most evident role of the reversible branch of PPP is to modulate recovery of pentose sugars to feed, through glucose-6P resynthesis, the oxidative PPP branch when reducing power, rather than R5P-linked nucleotide synthesis, is required. This pathway consists of four enzymes: two isomerases and two transferases. The isomerases refer to a ribulose-5-phosphate isomerase (Ru5PI), converting ribulose-5P (Ru5P) to R5P and a Ru5P epimerase (Ru5PE) converting Ru5P to xylulose-5P (Xu5P). The transferases refer to a transketolase (TKT) and a transaldolase (TAL). TKT is a thiaminepyrophosphate-dependent enzyme able to transfer a two-carbon group (a glycolaldehyde residue) from an α-ketosugar to an aldose, as is the case of the reactions between Xu5P and R5P, generating glyceraldehyde-3P (Ga3P) and sedoeptulose-7P (Se7P), and between Xu5P and erythrose-4P (Er4P), generating Ga3P and F6P. Finally, TAL catalyzes the transfer of a three carbon group (dihydroxyacetone residue) from an α-ketosugar (i.e., F6P and Se7P) to an aldose (i.e., Ga3P and Er4P) (Figure 2).
All of the reactions of this branch of PPP are reversible. However, nothing is more wrong than thinking of this pathway as a simple sealed container of processes at equilibrium. In fact, an articulated signaling network affecting the level of the involved enzymes occurs, making this pathway a relevant metabolic checkpoint for the cell health, in which different intermediates are functional links between the formation of NADPH and R5P and the different metabolic needs (Figure 2). Both TKT and TAL, which are differently represented in different tissues [64], catalyze relevant regulatory steps able to fulfil the main goals of PPP (i.e., to guarantee reducing power for anabolic demand and antioxidation protection and to allow nucleotides synthesis). The interplay of the enzymes involved in the two branches of PPP appears peculiar for different cells systems, leading, when altered, to cell type-specific pathologies [62,65]. Indeed, both the deficiency and the overexpression of TKT and TAL appeared to be linked to pathological states characterized by cell proliferation and cell death.
Transketolase-like 1, one of the mutated transketolase transcript isoforms of TKT, was found overexpressed in a variety of tumors. Gastric, uterine, colorectal, endometrial, renal, thyroid and liver cancer [66,67,68,69,70,71,72] are characterized by an elevated level of the enzyme, which was suggested as a marker of poor prognosis of the tumor and as potential antitumor target. TAL and G6PDH, are over expressed in different hepatomas [62]. The overexpression of these enzymes, which catalyze limiting steps of the two branches of the PPP, should fulfil the increased need of nucleotide synthesis for tumor cells. This would be accomplished through the channeling of glucose towards R5P synthesis from both the oxidative and the reversible arms of the pathway. The former would proceed through NADP+ reduction and the latter would favour, through the TKT-dependent generation of Er4P, the utilization of the glycolytic intermediates F6P and Ga3P [64]. Indeed, the interconnection between G6PDH and TAL appears to occur both on a structural and a functional basis, being the two enzymes in human neutrophils involved in a supramolecular structure, which appears to control the overall flux of the PPP [73]. This event led to hypothesize that the two enzymes are part of a functional strategic device devoted to equilibrate the two arms of the PPP. In such a competitive game, however, as predicted by a theoretical analysis of factors affecting the fluxes of G6P metabolism in red blood cells [74], TAL overexpression appears to act as a dominating factor in determining an overall decrease in NADPH production and thus a loss of antioxidant ability [64]. On the other hand, in Jurkat human leukemic T cells, the overexpression of TAL is accompanied by a deficiency of both G6PDH and 6PGDH. In this case, no competition between the two PPP branches occurs and, as predictable, a concomitant significant loss of NADPH, reduced levels of GSH and high rate of ROS formation were observed. These cells are prone to apoptotic events induced by H2O2 and NO, by signaling factors as tumor necrosis factor-α, anti-FAS monoclonal antibodies or simply by serum deprivation [75].
As it occurs for overexpression of TAL, the activation of the enzyme through phosphorylation resulted in a depletion of NADPH and induction of oxidative stress in fibroblasts from patients affected by Xeroderma pigmentosum or transformed by SV40. In these cell systems, the apparent involvement of TAL in the homeostasis of cell reduced state is concomitant with a low activity of catalase, another antioxidant enzyme whose activity could be restored by NADPH supplementation [76].
Not only hyperactivity, but also a reduced activity of TKT and TAL has a significant impact on cell function. A reduced activity of TKT was shown in neurodegenerative diseases, diabetes mellitus and cancer [77,78] and associated in diabetes to an increase in circulating ribose and glycation phenomena. Indeed, while the complete deficiency of TK and G6PDH is lethal, TAL deficiency may be tolerated by some cell types and tissues, even though the human organism cannot survive in the absence of this enzyme. TAL deficiency is associated to a variety of different pathologies, as multiple sclerosis and rheumatoid arthritis as well as different tumors [79]. It is remarkable how the modulation of NADPH and ROS levels, exerted by TAL activity, affects the mitochondrial trans-membrane potential [79]. In this regard, both TKT and, more incisively, TAL deficiency were reported to activate in vivo the mitochondrial unfolded protein response (UPRmt) in Caenorhabditis elegans. In particular, it was shown how TAL deficiency determines, through complex oxidative stress and a starvation-like responses, mitochondrial morphology changes, impairment of mitochondrial respiration and decreased fats level, and increases the longevity of the animal. To conclude, the impact of TAL activity in modulating NADPH levels and controlling the ROS production mainly associated to mitochondrial oxidative processes, appears the result of the ability of the enzyme to favour R5P accumulation. In the case of TAL overexpression, R5P accumulation would occur through the recruitment of glycolytic precursors. In the case of TAL deficiency, the impairment of the recycle of R5P into G6P, limits the efficiency of the oxidative branch of PPI. In this respect, the accumulation of neurotoxic molecules, as erythritol, arabitol and ribitol, concomitant with a marked decline of NADPH levels, led to propose the involvement in the process of aldose reductase [80], a NADPH-dependent enzyme for which four and five carbon atom aldoses, including ribose, are indeed better substrates than glucose [81].
We can complete the survey on the reversible branch of PPP by looking at the remaining two enzymes, namely Ru5PI and Ru5PE.These enzymes, through their action on Ru5P, connect the two branches of the pathway. In fact, the two reaction products, Xu5P and R5P, can be addressed towards G6P resynthesis, thus allowing the complete utilization of the glucose molecule and then the maximization of NADPH generation. On the other hand, R5P is the indubitable precursor for nucleotide synthesis either de novo, through further activation of R5P to PRPP, or salvage, through its isomerization to R1P by phosphopentomutase [82,83]. As mentioned above, also in this case, the flux of intermediates of the reversible branch of PPP is not functionally isolated. In fact, the epimerization of Ru5P to Xu5P affects lipogenesis, being Xu5P (together with glucose and fructose-1,6-bisphosphate) the inducer of lipogenic genes through the activation of a protein serine/threonine phosphatase acting on the ChREBP (Carbohydrate-responsive element binding protein) transcription factor [84,85,86]. Also, the efficiency of the isomerization reaction between Ru5P and R5P may enter in this metabolic control, which may dangerously evolve to cancer when significantly stressed, as reported, for instance, for Ru5P-isomerase overexpression in human hepatocellular carcinoma [87,88]. In this regard, we may conclude underlining that a body of experimental evidence merge in the conclusion that dysregulation of the PPP promotes carcinogenesis [89].
All of these considerations evidence that an intervention at any point on PPP, which alters the level of an intermediate, may affect a complex body of processes whose relevance in being damaging or even beneficial on health has not yet been fully clarified [90]. Thus, ribose intake, forcing the generation of R5P by a high substrate concentration, besides pushing toward nucleotide synthesis, would influence PPP with not yet clearly assessed consequences. If this practice may be in principle understandable or even acceptable in severe pathological situations as described above, in which “possible” damage is overcome by “ascertained” damage linked to the pathological status, it is not anymore understandable when it is applied to healthy, even “tired” subjects. On the basis of the above considerations, it should be clear when and why the ribose supply might be beneficial. “When”, in the case the generation of pentose phosphates through the G6PDH/6PGDH couple is knockdown by a failure of the oxidative PPP and/or in the case of a failure of the reversible PPP branch in recruiting glycolysis intermediate for R5P synthesis. “Why”, in the attempt to counteract an emergency. It should be obvious, however, that the treatment should occur under a strict medical control and for a proper limited time. This is due to the fact that the metabolic and functional “price” of such a therapeutic approach may be high (see below).
5. The Shady Side of the Ribose Intake
A relevant and deleterious feature of ribose is its glycative potential. When the question of protein glycation cannot be avoided, “ribose lovers” minimize the ribose features presenting this molecule as a natural sugar which, as it occurs for glucose, may induce protein glycation phenomena. This is true, but what is usually missed in the presentation of the molecule is that ribose is one of the sugars most effective in determining glycation reactions [91,92,93,94,95,96]. Indeed, protein glycation protocols are devised which make use of ribose to enhance glycation phenomena. Just as an example, in order to evaluate the influence of glycosylation on the progression of spontaneous osteoarthritis, the elective method to increase cartilage glycation was a topic injection of ribose in the knee of the animals [97]. Moreover, pentosidine, initially suggested as an index of protein glycation linked to dysmetabolism of pentoses [98] and nowadays considered one of the general biomarkers of advanced glycation end products (AGEs) [99], was firstly discovered as a product of the non-enzymatic reaction between ribose and lysine or arginine, and then formed in vitro by the reaction of ribose and other pentoses with human collagen [100].
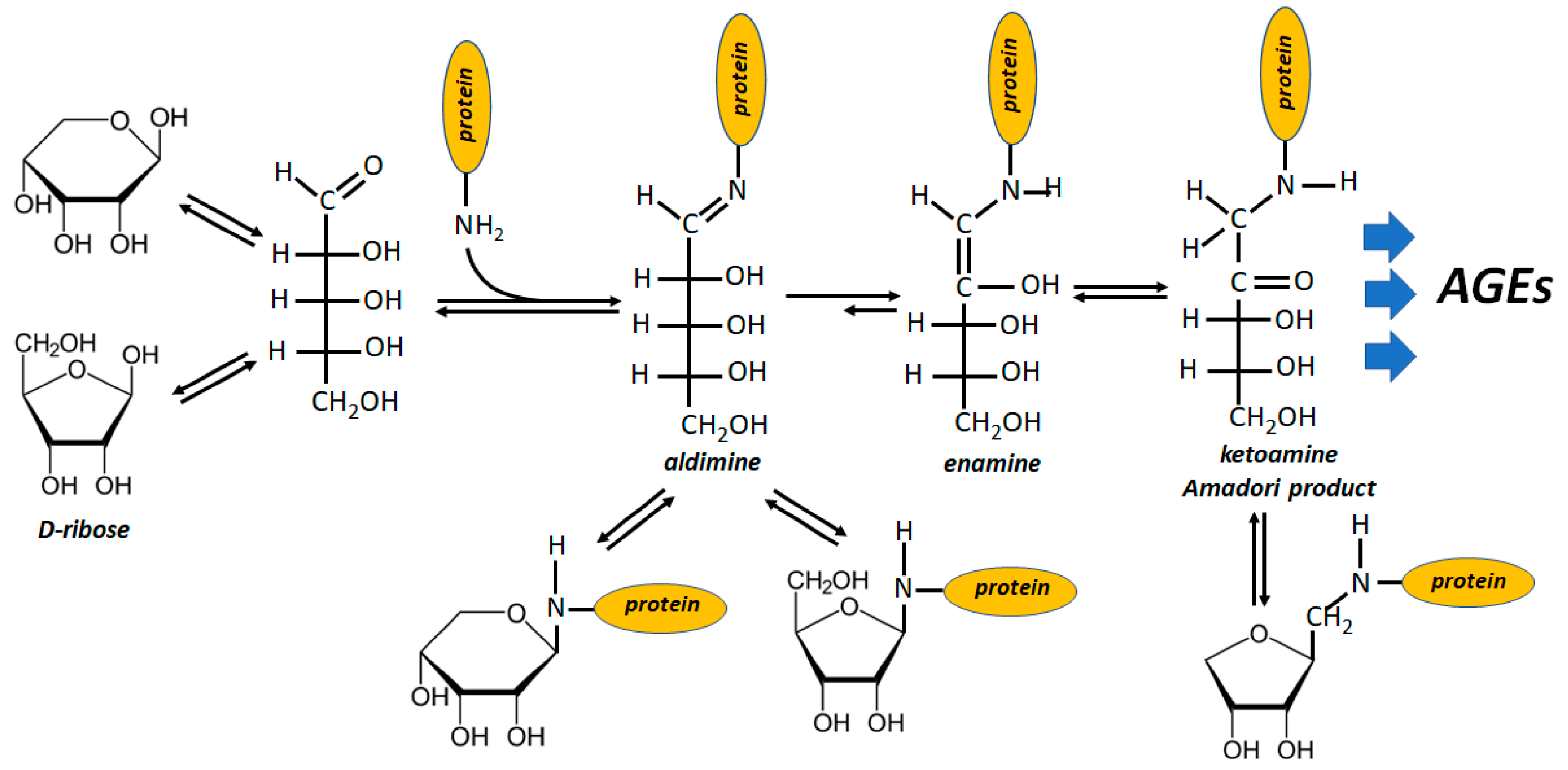
Protein glycation starts from the reaction of the free aldehydic group of sugars with protein amino residues and the special glycation ability of ribose stands on the high level of free aldehydic form which accounts for 20 to 30 times that of glucose [101] (Figure 3). In fact, R5P is unable to form a pyranosidic structure and displays an even higher availability of free aldehydic groups with respect to ribose [102,103]. It is not surprising, therefore, that also R5P is an efficient inducer of the Maillard reaction sequence (even more active than ribose, being 100-fold faster than glucose) [104]. Moreover, R5P enters the glycation mechanism with its phosphate group, which absolves a catalytic function in the products formation, promoting, at the same time, oxidative stress [103,105,106].
Many proteins have been found to be extremely susceptible to glycation by ribose, a process which evolves with generation of deleterious AGEs [91]. The glycating action on hemoglobin [92,107] and myoglobin [108,109] is remarkable. The deleterious effects induced by ribose glycation on other functional proteins are also widely documented. Thus, ribose-induced glycated serum was found to be toxic towards cultured pancreatic β-cells in terms of viability and insulin secretion [110]. Ribose, besides targeting albumin [93,111,112], was shown to actively glycate other plasma proteins, such as fibrinogen [113], low density lipoprotein [114,115,116], and immunoglobulin-G [96]. Also, neuro degenerative disorders find in AGEs generation a potent eliciting factor [117,118]. Thus, neuronal Tau-protein, α-synuclein, β2-microglobulin, following glycation, contribute to the development of several common neurodegenerative diseases [119,120,121,122,123,124]. In addition, cognitive impairments observed in animal models and in diabetic patients has been ascribed to glycation and AGEs accumulation linked to ribose [93,125,126,127,128,129,130]. Diabetic nephropathy also appears linked to glycation processes and AGEs accumulation [131,132,133]. Collagen is another possible glycation target with consequences in the proper function of bones and tendons [134,135,136,137]. It is rather difficult to find tissues or pathological situations in which the deleterious effect of not enzymatic glycation and AGEs accumulation can be excluded. Also, DNA and DNA related enzymes have been found to be glycated by ribose, with consequences that, speaking generally, are difficult to predict [116,138,139,140,141,142].
The source of the above underlined phenomena should be ascribed to the level of circulating ribose. The blood level of free ribose is rather low, resulting in fasting condition 50 times lower than glucose. Whether this may be related to the efficient preservation of the hemiacetal furanosidic scaffold of the molecule (i.e., R1P, as it occurs in the nucleotide salvage pathways), or to its high reactivity or both, is a debatable and open question. What appears indubitable, however, is the evident plan of nature to avoid this cytotoxic molecule standing around in the organism and, at the same time, fulfilling the need of the ribose scaffold making use of glucose (a more reliable and safer molecule) [1,8].
As for glucose, the ribose levels in blood and urine increase in diabetes, a fact shown to correlate with haemoglobin glycation and with a deficiency of TKT [107,143]. This evidence strongly supports the view of ribose as a relevant factor in the development of diabetes and related complications [95,107,144].
It is worth noting that the above list of references is only part of the vast field of reports dealing with the effects associated with glycation phenomena and AGEs accumulation induced by ribose.
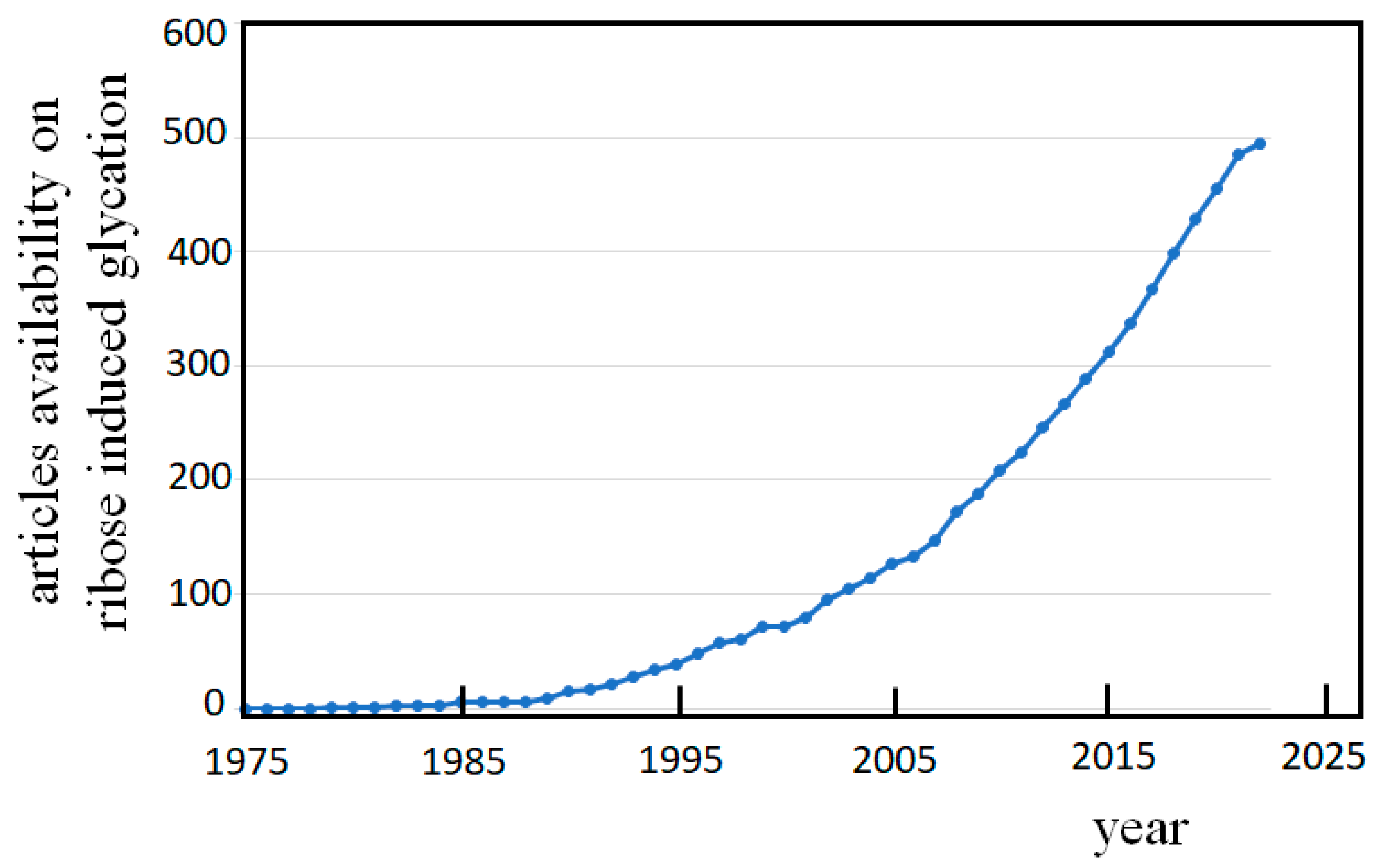
The number of publications on the damaging inference of these events during the past forty years, as emerges simply searching for the topic “glycation and ribose” in PubMed®, (Figure 4) clearly shows that the consciousness is arising within the scientific community regarding the risky phenomenon of ribose glycation.
Thus, it is somehow disturbing that even nowadays there are papers dealing with the effects of ribose administration in human in which the words “glycation” or “AGEs” are lacking, especially when the conclusion is suggestive of the good and safe practice for healthy people to assume ribose to feel better. On the other hand, a similar lack of consciousness is evident when the risky phenomenon of ribose-glycation is experimentally faced. Emblematic in this respect are the results coming from administration of ribose to horses [4]. This paper, which is frequently cited as a reference in commercial advertisement inserts supporting ribose as a dietary integrator, reports that the treated animals felt better and no glycation phenomena were observed. In conclusion, ribose intake is satisfactory for the horses and should be good also for human.
Considering the glycation ability of ribose, we do believe that such an unexpected result is, at least, too hazardous in suggesting a safe use of ribose in humans.
Unfortunately, the claim for a safe and desirable use of ribose as food integrator is not so infrequent as conclusion statement in the literature dealing with ribose intake. So, the feasibly harmful action of ribose remains underestimated in the commercial world.
6. Conclusions
Ribose is a cytotoxic molecule whose availability in the cell is apparently kept under rigorous control. Clear evidence shows the benefits of intervention of ribose in peculiar distress situations as it may occur in ischemia, cardiomyopathy, and hypertrophy. However, if one is to consider ribose as a potential therapeutic tool in conditions in which the molecule may relieve the patient from discomfort, one must think to administer ribose as an integrator to healthy people. In the former case, we face, under a rigorous medical control, a damaging distress situation in which both patient and physician are aware of the balance between benefits and side effects. On the contrary, in the case of the use of ribose as food integrator, based on the suggestion to become more energetic and stronger, the users are driven only by the unlimited desire of humans to feel better. Ribose is therefore a quite interesting and intriguing molecule, but much more experimental effort is needed to safely enjoy of it, if even it will be the case.
Author Contributions
All authors contributed to the drafting of the manuscript and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by University of Pisa, Fondi di Ateneo 2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Maria Chiara Mura for drawing the Graphical Abstract.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Del-Corso, A.; Cappiello, M.; Moschini, R.; Balestri, F.; Mura, U.; Ipata, P.L. The Furanosidic Scaffold of d-Ribose: A Milestone for Cell Life. Biochem. Soc. Trans. 2019, 47, 1931–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitelbaum, J.E.; Johnson, C.; St Cyr, J. The Use of D-Ribose in Chronic Fatigue Syndrome and Fibromyalgia: A Pilot Study. J. Altern. Complement. Med. 2006, 12, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Neutel, J.; Homer, K.; Tempero, K.; Shah, A.; Khankari, R. Evaluation of D-Ribose Pharmacokinetics, Dose Proportionality, Food Effect, and Pharmacodynamics after Oral Solution Administration in Healthy Male and Female Subjects. J. Clin. Pharmacol. 2014, 54, 546–554. [Google Scholar] [CrossRef]
- Sinatra, S.T.; Caiazzo, C. (D)-Ribose Supplementation in the Equine: Lack of Effect on Glycated Plasma Proteins Suggesting Safety in Humans. J. Am. Coll. Nutr. 2015, 34, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.E.; Hiebert, J.B.; Thimmesch, A.; Pierce, J.T.; Vacek, J.L.; Clancy, R.L.; Sauer, A.J.; Pierce, J.D. Understanding D-Ribose and Mitochondrial Function. Adv. Biosci. Clin. Med. 2018, 6, 1–5. [Google Scholar] [CrossRef]
- Alzoubi, K.; Ismail, Z.; Al-Essa, M.; Alshogran, O.; Abutayeh, R.; Abu-Baker, N. Pharmacokinetic Evaluation of D-Ribose after Oral and Intravenous Administration to Healthy Rabbits. CPAA 2018, 10, 73–78. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA); Turck, D.; Bresson, J.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.; et al. Safety of D-ribose as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2018, 16, e05265. [Google Scholar] [CrossRef] [Green Version]
- Del-Corso, A.; Cappiello, M.; Moschini, R.; Balestri, F.; Mura, U. How the Chemical Features of Molecules May Have Addressed the Settlement of Metabolic Steps. Metabolomics 2018, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Bunn, H.F.; Higgins, P.J. Reaction of Monosaccharides with Proteins: Possible Evolutionary Significance. Science 1981, 213, 222–224. [Google Scholar] [CrossRef]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.-M.; Krüger, A.; Tauqeer Alam, M.; et al. The Return of Metabolism: Biochemistry and Physiology of the Pentose Phosphate Pathway. Biol. Rev. Camb. Philos. Soc. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- Williams, J.F.; Blackmore, P.F. Non-Oxidative Synthesis of Pentose 5-Phosphate from Hexose 6-Phosphate and Triose Phosphate by the L-Type Pentose Pathway. Int. J. Biochem. 1983, 15, 797–816. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Sander, C.; Valencia, A. Convergent Evolution of Similar Enzymatic Function on Different Protein Folds: The Hexokinase, Ribokinase, and Galactokinase Families of Sugar Kinases: Convergent Evolution of Sugar Kinases. Protein Sci. 1993, 2, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; van Koeverden, P.; Singh, B.; Gupta, R.S. Identification and Characterization of Human Ribokinase and Comparison of Its Properties with E. coli Ribokinase and Human Adenosine Kinase. FEBS Lett. 2007, 581, 3211–3216. [Google Scholar] [CrossRef] [Green Version]
- Quiroga-Roger, D.; Babul, J.; Guixé, V. Role of Monovalent and Divalent Metal Cations in Human Ribokinase Catalysis and Regulation. Biometals 2015, 28, 401–413. [Google Scholar] [CrossRef]
- Chuvikovsky, D.; Esipov, R.; Skoblov, Y.; Chupova, L.; Muravyova, T.; Miroshnikov, A.; Lapinjoki, S.; Mikhailopulo, I. Ribokinase from E. coli: Expression, Purification, and Substrate Specificity. Bioorganic Med. Chem. 2006, 14, 6327–6332. [Google Scholar] [CrossRef] [PubMed]
- Maj, M.C.; Gupta, R.S. The Effect of Inorganic Phosphate on the Activity of Bacterial Ribokinase. J. Protein Chem. 2001, 20, 139–144. [Google Scholar] [CrossRef]
- Yamada, Y.; Goto, H.; Ogasawara, N. Adenosine Kinase from Human Liver. Biochim. Biophys. Acta 1981, 660, 36–43. [Google Scholar] [CrossRef]
- Schnebli, H.P.; Hill, D.L.; Bennett, L.L. Purification and Properties of Adenosine Kinase from Human Tumor Cells of Type H. Ep. No. 2. J. Biol. Chem. 1967, 242, 1997–2004. [Google Scholar] [CrossRef] [PubMed]
- Palella, T.D.; Andres, C.M.; Fox, I.H. Human Placental Adenosine Kinase: Kinetic Mechanism and Inhibition. J. Biol. Chem. 1980, 255, 5264–5269. [Google Scholar] [CrossRef]
- Angyal, S.; Bethell, G.; Cowley, D.; Pickles, V. Equilibria between Pyranoses and Furanoses. IV. 1-Deoxyhexuloses and 3-Hexuloses. Aust. J. Chem. 1976, 29, 1239. [Google Scholar] [CrossRef]
- Angyal, S.; Bethell, G. Conformational Analysis in Carbohydrate Chemistry: III. The 13C N.M.R. Spectra of the Hexuloses. Aust. J. Chem. 1976, 29, 1249. [Google Scholar] [CrossRef]
- Mahoney, J.R.; Sako, E.Y.; Seymour, K.M.; Marquardt, C.A.; Foker, J.E. A Comparison of Different Carbohydrates as Substrates for the Isolated Working Heart. J. Surg. Res. 1989, 47, 530–534. [Google Scholar] [CrossRef]
- Chang, R.S. Genetic study of human cells in vitro. J. Exp. Med. 1960, 111, 235–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitzer, L.J.; Wice, B.M.; Kennell, D. The Pentose Cycle. Control and Essential Function in HeLa Cell Nucleic Acid Synthesis. J. Biol. Chem. 1980, 255, 5616–5626. [Google Scholar] [CrossRef]
- Zimmer, H.G.; Ibel, H. Ribose Accelerates the Repletion of the ATP Pool during Recovery from Reversible Ischemia of the Rat Myocardium. J. Mol. Cell. Cardiol. 1984, 16, 863–866. [Google Scholar] [CrossRef] [PubMed]
- St Cyr, J.A.; Bianco, R.W.; Schneider, J.R.; Mahoney, J.R.; Tveter, K.; Einzig, S.; Foker, J.E. Enhanced High Energy Phosphate Recovery with Ribose Infusion after Global Myocardial Ischemia in a Canine Model. J. Surg. Res. 1989, 46, 157–162. [Google Scholar] [CrossRef]
- Pliml, W.; von Arnim, T.; Stäblein, A.; Hofmann, H.; Zimmer, H.G.; Erdmann, E. Effects of Ribose on Exercise-Induced Ischaemia in Stable Coronary Artery Disease. Lancet 1992, 340, 507–510. [Google Scholar] [CrossRef]
- Bayram, M.; St. Cyr, J.A.; Abraham, W.T. d-Ribose Aids Heart Failure Patients with Preserved Ejection Fraction and Diastolic Dysfunction: A Pilot Study. Ther. Adv. Cardiovasc. Dis. 2015, 9, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Shecterle, L.M.; Terry, K.R.; St Cyr, J.A. Potential Clinical Benefits of D-Ribose in Ischemic Cardiovascular Disease. Cureus 2018, 10, e2291. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, A.; Henriksson, K.G.; Larsson, J. Reduced High-Energy Phosphate Levels in the Painful Muscles of Patients with Primary Fibromyalgia. Arthritis Rheum. 1986, 29, 817–821. [Google Scholar] [CrossRef]
- Patten, B.M. Beneficial Effect of D-Ribose in Patient with Myoadenylate Deaminase Deficiency. Lancet 1982, 1, 1071. [Google Scholar] [CrossRef] [PubMed]
- Lecky, B.R.F. Failure of D-Ribose in Myoadenylate Deaminase Deficiency. Lancet 1983, 321, 193. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.F.; Pepine, C.J. D-Ribose as a Supplement for Cardiac Energy Metabolism. J. Cardiovasc. Pharmacol. Ther. 2000, 5, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Xiao, Y.; Zhang, L.; Fang, J.; Yang, N.; Zhang, Z.; Nasser, M.I.; Qin, H. D-ribose: Potential Clinical Applications in Congestive Heart Failure and Diabetes, and Its Complications (Review). Exp. Ther. Med. 2021, 21, 496. [Google Scholar] [CrossRef]
- Kreider, R.B.; Melton, C.; Greenwood, M.; Rasmussen, C.; Lundberg, J.; Earnest, C.; Almada, A. Effects of Oral D-Ribose Supplementation on Anaerobic Capacity and Selected Metabolic Markers in Healthy Males. Int. J. Sport. Nutr. Exerc. Metab. 2003, 13, 76–86. [Google Scholar] [CrossRef]
- Berardi, J.M.; Ziegenfuss, T.N. Effects of Ribose Supplementation on Repeated Sprint Performance in Men. J. Strength Cond. Res. 2003, 17, 47–52. [Google Scholar] [CrossRef]
- Hellsten, Y.; Skadhauge, L.; Bangsbo, J. Effect of Ribose Supplementation on Resynthesis of Adenine Nucleotides after Intense Intermittent Training in Humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R182–R188. [Google Scholar] [CrossRef]
- Op’t Eijnde, B.; Van Leemputte, M.; Brouns, F.; Van Der Vusse, G.J.; Labarque, V.; Ramaekers, M.; Van Schuylenberg, R.; Verbessem, P.; Wijnen, H.; Hespel, P. No Effects of Oral Ribose Supplementation on Repeated Maximal Exercise and de Novo ATP Resynthesis. J. Appl. Physiol. 2001, 91, 2275–2281. [Google Scholar] [CrossRef]
- Raue, U.; Gallagher, P.M.; Williamson, D.L.; Godard, M.P.; Trappe, S.W. Effects of Ribose Supplementation on Performance During Repeated High-Intensity Cycle Sprints. Med. Sci. Sports Exerc. 2001, 33, S44. [Google Scholar] [CrossRef]
- Van Gammeren, D.; Falk, D.; Antonio, J. The Effects of Four Weeks of Ribose Supplementation on Body Composition and Exercise Performance in Healthy, Young, Male Recreational Bodybuilders: A Double-Blind, Placebo-Controlled Trial. Curr. Ther. Res. 2002, 63, 486–495. [Google Scholar] [CrossRef]
- Seifert, J.G.; Brumet, A.; St Cyr, J.A. The Influence of D-Ribose Ingestion and Fitness Level on Performance and Recovery. J. Int. Soc. Sports Nutr. 2017, 14, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Qiu, J.; Cai, T.; Yi, L.; Benardot, D.; Zou, M. Effect of D-Ribose Supplementation on Delayed Onset Muscle Soreness Induced by Plyometric Exercise in College Students. J. Int. Soc. Sports Nutr. 2020, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Patra, K.C.; Hay, N. The Pentose Phosphate Pathway and Cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.; Cheng, M.; Chiu, D.T. Glucose-6-Phosphate Dehydrogenase—From Oxidative Stress to Cellular Functions and Degenerative Diseases. Redox Rep. 2007, 12, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, T.J.; D’Urso, M.; Battistuzzi, G.; Estrada, M.; Foulkes, N.S.; Martini, G.; Calabro, V.; Poggi, V.; Giordano, R.; Town, M. Diverse Point Mutations in the Human Glucose-6-Phosphate Dehydrogenase Gene Cause Enzyme Deficiency and Mild or Severe Hemolytic Anemia. Proc. Natl. Acad. Sci. USA 1988, 85, 5171–5175. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Manzo, S.; Marcial-Quino, J.; Vanoye-Carlo, A.; Serrano-Posada, H.; Ortega-Cuellar, D.; González-Valdez, A.; Castillo-Rodríguez, R.; Hernández-Ochoa, B.; Sierra-Palacios, E.; Rodríguez-Bustamante, E.; et al. Glucose-6-Phosphate Dehydrogenase: Update and Analysis of New Mutations around the World. Int. J. Mol. Sci. 2016, 17, 2069. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.H.; Bhutani, V.K.; Brown, A.K. System-Based Approach to Management of Neonatal Jaundice and Prevention of Kernicterus. J. Pediatr. 2002, 140, 396–403. [Google Scholar] [CrossRef]
- Filosa, S.; Cai, W.; Galanello, R.; Cao, A.; De Mattia, D.; Schettini, F.; Martini, G. A Novel Single-Base Mutation in the Glucose 6-Phosphate Dehydrogenase Gene Is Associated with Chronic Non-Spherocytic Haemolytic Anaemia. Hum. Genet. 1994, 94, 560–562. [Google Scholar] [CrossRef]
- Luzzatto, L.; Poggi, V. Chapter 17: Glucose-6-Phosphate Dehydrogenase Deficiency. In Nathan and Oski’s Hematology of Infancy and Childhood; Orkin, S., Ed.; Elsevier: Philadelphia, PA, USA, 2009. [Google Scholar]
- Manganelli, G.; Fico, A.; Martini, G.; Filosa, S. Discussion on Pharmacogenetic Interaction in G6PD Deficiency and Methods to Identify Potential Hemolytic Drugs. Cardiovasc. Hematol. Disord. Drug Targets 2010, 10, 143–150. [Google Scholar] [CrossRef]
- Liu, W.-L.; Li, F.; He, Z.-X.; Jiang, H.-Y.; Ai, R. Glucose-6-Phosphate Dehydrogenase Qingzhen: Identification of a Novel Splice Mutation (IVS5-1 G>A). Pediatr. Blood Cancer 2012, 58, 825–826. [Google Scholar] [CrossRef]
- Luzzatto, L.; Arese, P. Favism and Glucose-6-Phosphate Dehydrogenase Deficiency. N. Engl. J. Med. 2018, 378, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutler, E. Glucose-6-Phosphate Dehydrogenase Deficiency: A Historical Perspective. Blood 2008, 111, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisels, M.J. Neonatal Jaundice. Pediatr. Rev. 2006, 27, 443–454. [Google Scholar] [CrossRef]
- Carette, C.; Dubois-Laforgue, D.; Gautier, J.-F.; Timsit, J. Diabetes Mellitus and Glucose-6-Phosphate Dehydrogenase Deficiency: From One Crisis to Another. Diabetes Metab. 2011, 37, 79–82. [Google Scholar] [CrossRef]
- Leopold, J.A.; Cap, A.; Scribner, A.W.; Stanton, R.C.; Loscalzo, J. Glucose-6-phosphate Dehydrogenase Deficiency Promotes Endothelial Oxidant Stress and Decreases Endothelial Nitric Oxide Bioavailability. FASEB J. 2001, 15, 1771–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Brenner, D.A.; Cui, L.; Lim, C.C.; Wang, B.; Pimentel, D.R.; Koh, S.; Sawyer, D.B.; Leopold, J.A.; Handy, D.E.; et al. Glucose-6-Phosphate Dehydrogenase Modulates Cytosolic Redox Status and Contractile Phenotype in Adult Cardiomyocytes. Circ. Res. 2003, 93, e9–e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Cui, L.; Brenner, D.A.; Wang, B.; Handy, D.E.; Leopold, J.A.; Loscalzo, J.; Apstein, C.S.; Liao, R. Increased Myocardial Dysfunction After Ischemia-Reperfusion in Mice Lacking Glucose-6-Phosphate Dehydrogenase. Circulation 2004, 109, 898–903. [Google Scholar] [CrossRef] [Green Version]
- Caprari, P.; Caforio, M.P.; Cianciulli, P.; Maffi, D.; Pasquino, M.T.; Tarzia, A.; Amadori, S.; Salvati, A.M. 6-Phosphogluconate Dehydrogenase Deficiency in an Italian Family. Ann. Hematol. 2001, 80, 41–44. [Google Scholar] [CrossRef]
- Vives Corrons, J.L.; Colomer, D.; Pujades, A.; Rovira, A.; Aymerich, M.; Merino, A.; Aguilar i Bascompte, J.L. Congenital 6-Phosphogluconate Dehydrogenase (6PGD) Deficiency Associated with Chronic Hemolytic Anemia in a Spanish Family. Am. J. Hematol. 1996, 53, 221–227. [Google Scholar] [CrossRef]
- Beutler, E.; Kuhl, W.; Gelbart, T. 6-Phosphogluconolactonase Deficiency, a Hereditary Erythrocyte Enzyme Deficiency: Possible Interaction with Glucose-6-Phosphate Dehydrogenase Deficiency. Proc. Natl. Acad. Sci. USA 1985, 82, 3876–3878. [Google Scholar] [CrossRef]
- Kowalik, M.A.; Columbano, A.; Perra, A. Emerging Role of the Pentose Phosphate Pathway in Hepatocellular Carcinoma. Front. Oncol. 2017, 7, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Sun, H.; Zhang, S.; Shan, C. The Multiple Roles of Glucose-6-Phosphate Dehydrogenase in Tumorigenesis and Cancer Chemoresistance. Life 2022, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Morris, H.P.; Weber, G. Behavior of Transaldolase (EC 2.2. 1.2) and Transketolase (EC 2.2. 1.1) Activities in Normal, Neoplastic, Differentiating, and Regenerating Liver. Cancer Res. 1976, 36, 3189–3197. [Google Scholar] [PubMed]
- Samland, A.K.; Sprenger, G.A. Transaldolase: From Biochemistry to Human Disease. Int. J. Biochem. Cell. Biol. 2009, 41, 1482–1494. [Google Scholar] [CrossRef]
- Ahopelto, K.; Böckelman, C.; Hagström, J.; Koskensalo, S.; Haglund, C. Transketolase-like Protein 1 Expression Predicts Poor Prognosis in Colorectal Cancer. Cancer Biol. Ther. 2016, 17, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, J.-H.; Guo, C.-K.; Cai, P. Gene Silencing of TKTL1 by RNAi Inhibits Cell Proliferation in Human Hepatoma Cells. Cancer Lett. 2007, 253, 108–114. [Google Scholar] [CrossRef]
- Krockenberger, M.; Engel, J.B.; Schmidt, M.; Kohrenhagen, N.; Häusler, S.F.M.; Dombrowski, Y.; Kapp, M.; Dietl, J.; Honig, A. Expression of Transketolase-like 1 Protein (TKTL1) in Human Endometrial Cancer. Anticancer Res. 2010, 30, 1653–1659. [Google Scholar]
- Langbein, S.; Frederiks, W.M.; zur Hausen, A.; Popa, J.; Lehmann, J.; Weiss, C.; Alken, P.; Coy, J.F. Metastasis Is Promoted by a Bioenergetic Switch: New Targets for Progressive Renal Cell Cancer. Int. J. Cancer 2008, 122, 2422–2428. [Google Scholar] [CrossRef]
- Zerilli, M.; Amato, M.C.; Martorana, A.; Cabibi, D.; Coy, J.F.; Cappello, F.; Pompei, G.; Russo, A.; Giordano, C.; Rodolico, V. Increased Expression of Transketolase-like-1 in Papillary Thyroid Carcinomas Smaller than 1.5 Cm in Diameter Is Associated with Lymph-Node Metastases. Cancer 2008, 113, 936–944. [Google Scholar] [CrossRef]
- Chen, H.; Yue, J.-X.; Yang, S.-H.; Ding, H.; Zhao, R.-W.; Zhang, S. Overexpression of Transketolase-like Gene 1 Is Associated with Cell Proliferation in Uterine Cervix Cancer. J. Exp. Clin. Cancer Res. 2009, 28, 43. [Google Scholar] [CrossRef] [Green Version]
- Ahopelto, K.; Laitinen, A.; Hagström, J.; Böckelman, C.; Haglund, C. Transketolase-Like Protein 1 and Glucose Transporter 1 in Gastric Cancer. Oncology 2020, 98, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-B.; Espinoza, J.; Romero, R.; Petty, H.R. Transaldolase Is Part of a Supramolecular Complex Containing Glucose-6-Phosphate Dehydrogenase in Human Neutrophils That Undergoes Retrograde Trafficking during Pregnancy. Metabolism 2005, 54, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Ni, T.-C.; Savageau, M.A. Application of Biochemical Systems Theory to Metabolism in Human Red Blood Cells. J. Biol. Chem. 1996, 271, 7927–7941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banki, K.; Hutter, E.; Colombo, E.; Gonchoroff, N.J.; Perl, A. Glutathione Levels and Sensitivity to Apoptosis Are Regulated by Changes in Transaldolase Expression. J. Biol. Chem. 1996, 271, 32994–33001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachaise, F.; Martin, G.; Drougard, C.; Perl, A.; Vuillaume, M.; Wegnez, M.; Sarasin, A.; Daya-Grosjean, L. Relationship between Posttranslational Modification of Transaldolase and Catalase Deficiency in UV-Sensitive Repair-Deficient Xeroderma Pigmentosum Fibroblasts and SV40-Transformed Human Cells. Free Radic. Biol. Med. 2001, 30, 1365–1373. [Google Scholar] [CrossRef]
- Coy, J.F.; Dressler, D.; Wilde, J.; Schubert, P. Mutations in the Transketolase-like Gene TKTL1: Clinical Implications for Neurodegenerative Diseases, Diabetes and Cancer. Clin. Lab. 2005, 51, 257–273. [Google Scholar]
- Zhao, J.; Zhong, C.-J. A Review on Research Progress of Transketolase. Neurosci. Bull. 2009, 25, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Perl, A. The Pathogenesis of Transaldolase Deficiency. IUBMB Life 2007, 59, 365–373. [Google Scholar] [CrossRef]
- Perl, A.; Hanczko, R.; Telarico, T.; Oaks, Z.; Landas, S. Oxidative Stress, Inflammation and Carcinogenesis Are Controlled through the Pentose Phosphate Pathway by Transaldolase. Trends Mol. Med. 2011, 17, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Balestri, F.; Cappiello, M.; Moschini, R.; Rotondo, R.; Abate, M.; Del-Corso, A.; Mura, U. Modulation of Aldose Reductase Activity by Aldose Hemiacetals. Biochim. Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 2329–2339. [Google Scholar] [CrossRef] [Green Version]
- Tozzi, M.G.; Camici, M.; Mascia, L.; Sgarrella, F.; Ipata, P.L. Pentose Phosphates in Nucleoside Interconversion and Catabolism. FEBS J. 2006, 273, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Maliekal, P.; Sokolova, T.; Vertommen, D.; Veiga-da-Cunha, M.; Van Schaftingen, E. Molecular Identification of Mammalian Phosphopentomutase and Glucose-1,6-Bisphosphate Synthase, Two Members of the Alpha-D-Phosphohexomutase Family. J. Biol. Chem. 2007, 282, 31844–31851. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, T.; Kawaguchi, T.; Wadzinski, B.E.; Uyeda, K. Xylulose 5-Phosphate Mediates Glucose-Induced Lipogenesis by Xylulose 5-Phosphate-Activated Protein Phosphatase in Rat Liver. Proc. Natl. Acad. Sci. USA 2003, 100, 5107–5112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iizuka, K.; Wu, W.; Horikawa, Y.; Takeda, J. Role of Glucose-6-Phosphate and Xylulose-5-Phosphate in the Regulation of Glucose-Stimulated Gene Expression in the Pancreatic β Cell Line, INS-1E. Endocr. J. 2013, 60, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Iizuka, K. The Roles of Carbohydrate Response Element Binding Protein in the Relationship between Carbohydrate Intake and Diseases. Int. J. Mol. Sci. 2021, 22, 12058. [Google Scholar] [CrossRef]
- Chou, Y.-T.; Chen, L.-Y.; Tsai, S.-L.; Tu, H.-C.; Lu, J.-W.; Ciou, S.-C.; Wang, H.-D.; Yuh, C.-H. Ribose-5-Phosphate Isomerase A Overexpression Promotes Liver Cancer Development in Transgenic Zebrafish via Activation of ERK and β-Catenin Pathways. Carcinogenesis 2019, 40, 461–473. [Google Scholar] [CrossRef]
- Ciou, S.; Chou, Y.; Liu, Y.; Nieh, Y.; Lu, J.; Huang, S.; Chou, Y.; Cheng, L.; Lo, J.; Chen, M.; et al. Ribose-5-phosphate Isomerase A Regulates Hepatocarcinogenesis via PP2A and ERK Signaling. Int. J. Cancer 2015, 137, 104–115. [Google Scholar] [CrossRef]
- Jin, L.; Zhou, Y. Crucial Role of the Pentose Phosphate Pathway in Malignant Tumors (Review). Oncol. Lett. 2019, 17, 4213–4221. [Google Scholar] [CrossRef] [Green Version]
- Ge, T.; Yang, J.; Zhou, S.; Wang, Y.; Li, Y.; Tong, X. The Role of the Pentose Phosphate Pathway in Diabetes and Cancer. Front. Endocrinol. 2020, 11, 365. [Google Scholar] [CrossRef]
- Wei, Y.; Han, C.S.; Zhou, J.; Liu, Y.; Chen, L.; He, R.Q. D-Ribose in Glycation and Protein Aggregation. Biochim. Biophys. Acta 2012, 1820, 488–494. [Google Scholar] [CrossRef]
- Siddiqui, Z.; Ishtikhar, M.; Moinuddin, N.; Ahmad, S. D-Ribose Induced Glycoxidative Insult to Hemoglobin Protein: An Approach to Spot Its Structural Perturbations. Int. J. Biol. Macromol. 2018, 112, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, L.; Wang, Y.; Wei, Y.; Xu, Y.; He, T.; He, R. D-Ribose Contributes to the Glycation of Serum Protein. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 2285–2292. [Google Scholar] [CrossRef] [PubMed]
- Feroz, A.; Khaki, P.S.S.; Khan, M.S.; Bano, B. Protein Aggregation as a Consequence of Non-Enzymatic Glycation: Therapeutic Intervention Using Aspartic Acid and Arginine. Int. J. Biol. Macromol. 2020, 163, 1844–1858. [Google Scholar] [CrossRef] [PubMed]
- Mou, L.; Hu, P.; Cao, X.; Chen, Y.; Xu, Y.; He, T.; Wei, Y.; He, R. Comparison of Bovine Serum Albumin Glycation by Ribose and Fructose in Vitro and in Vivo. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2022, 1868, 166283. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Al-Shaghdali, K.; Rehman, S.; Khan, M.Y.; Rafi, Z.; Faisal, M.; Alatar, A.A.; Tahir, I.K.; Khan, S.; Ahmad, S.; et al. Nonenzymatic Glycosylation of Isolated Human Immunoglobulin-G by D-ribose. Cell Biochem. Funct. 2022, 40, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Willett, T.L.; Kandel, R.; De Croos, J.N.A.; Avery, N.C.; Grynpas, M.D. Enhanced Levels of Non-Enzymatic Glycation and Pentosidine Crosslinking in Spontaneous Osteoarthritis Progression. Osteoarthr. Cartil. 2012, 20, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Sell, D.R.; Monnier, V.M. End-Stage Renal Disease and Diabetes Catalyze the Formation of a Pentose-Derived Crosslink from Aging Human Collagen. J. Clin. Investig. 1990, 85, 380–384. [Google Scholar] [CrossRef]
- Dyer, D.G.; Blackledge, J.A.; Thorpe, S.R.; Baynes, J.W. Formation of Pentosidine during Nonenzymatic Browning of Proteins by Glucose. Identification of Glucose and Other Carbohydrates as Possible Precursors of Pentosidine in Vivo. J. Biol. Chem. 1991, 266, 11654–11660. [Google Scholar] [CrossRef]
- Sell, D.R.; Monnier, V.M. Structure Elucidation of a Senescence Cross-Link from Human Extracellular Matrix. J. Biol. Chem. 1989, 264, 21597–21602. [Google Scholar] [CrossRef]
- Dworkin, J.P.; Miller, S.L. A Kinetic Estimate of the Free Aldehyde Content of Aldoses. Carbohydr. Res. 2000, 329, 359–365. [Google Scholar] [CrossRef]
- Pierce, J.; Serianni, A.S.; Barker, R. Anomerization of Furanose Sugars and Sugar Phosphates. J. Am. Chem. Soc. 1985, 107, 2448–2456. [Google Scholar] [CrossRef]
- Munanairi, A.; O’Banion, S.K.; Gamble, R.; Breuer, E.; Harris, A.W.; Sandwick, R.K. The Multiple Maillard Reactions of Ribose and Deoxyribose Sugars and Sugar Phosphates. Carbohydr. Res. 2007, 342, 2575–2592. [Google Scholar] [CrossRef]
- Sandwick, R.; Johanson, M.; Breuer, E. Maillard Reactions of Ribose 5-Phosphate and Amino Acids. Ann. N. Y. Acad. Sci. 2005, 1043, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Gersten, R.A.; Gretebeck, L.M.; Hildick-Smith, G.; Sandwick, R.K. Maillard Reaction of Ribose 5-Phosphate Generates Superoxide and Glycation Products for Bovine Heart Cytochrome c Reduction. Carbohydr. Res. 2010, 345, 2499–2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fica-Contreras, S.M.; Shuster, S.O.; Durfee, N.D.; Bowe, G.J.K.; Henning, N.J.; Hill, S.A.; Vrla, G.D.; Stillman, D.R.; Suralik, K.M.; Sandwick, R.K.; et al. Glycation of Lys-16 and Arg-5 in Amyloid-β and the Presence of Cu2+ Play a Major Role in the Oxidative Stress Mechanism of Alzheimer’s Disease. J. Biol. Inorg. Chem. 2017, 22, 1211–1222. [Google Scholar] [CrossRef]
- Chen, X.; Su, T.; Chen, Y.; He, Y.; Liu, Y.; Xu, Y.; Wei, Y.; Li, J.; He, R. D -Ribose as a Contributor to Glycated Haemoglobin. EBioMedicine 2017, 25, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokiej, M.; Livermore, A.T.; Harris, A.W.; Onishi, A.C.; Sandwick, R.K. Ribose Sugars Generate Internal Glycation Cross-Links in Horse Heart Myoglobin. Biochem. Biophys. Res. Commun. 2011, 407, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; You, Y.; Gao, S.-Q.; Tang, S.; Chen, L.; Wen, G.-B.; Lin, Y.-W. Identification of the Protein Glycation Sites in Human Myoglobin as Rapidly Induced by D-Ribose. Molecules 2021, 26, 5829. [Google Scholar] [CrossRef]
- Viviani, G.L.; Puddu, A.; Sacchi, G.; Garuti, A.; Storace, D.; Durante, A.; Monacelli, F.; Odetti, P. Glycated Fetal Calf Serum Affects the Viability of an Insulin-Secreting Cell Line in Vitro. Metabolism 2008, 57, 163–169. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, L.; Chen, J.; Ge, L.; He, R.Q. Rapid Glycation with D-Ribose Induces Globular Amyloid-like Aggregations of BSA with High Cytotoxicity to SH-SY5Y Cells. BMC Cell. Biol. 2009, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Basak, P.; Pramanik, A.; Majumder, R.; Ghosh, A.; Hazra, S.; Guria, M.; Bhattacharyya, M.; Banik, S.P. Ribosylation Induced Structural Changes in Bovine Serum Albumin: Understanding High Dietary Sugar Induced Protein Aggregation and Amyloid Formation. Heliyon 2020, 6, e05053. [Google Scholar] [CrossRef] [PubMed]
- Khanam, A.; Alouffi, S.; Rehman, S.; Ansari, I.A.; Shahab, U.; Ahmad, S. An in Vitro Approach to Unveil the Structural Alterations in d-Ribose Induced Glycated Fibrinogen. J. Biomol. Struct. Dyn. 2021, 39, 5209–5223. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Akhter, F.; Moinuddin; Shahab, U.; Khan, M.S. Studies on Glycation of Human Low Density Lipoprotein: A Functional Insight into Physico-Chemical Analysis. Int. J. Biol. Macromol. 2013, 62, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Akhter, F.; Salman Khan, M.; Faisal, M.; Alatar, A.A.; Ahmad, S. Detection of Circulating Auto-Antibodies Against Ribosylated-LDL in Diabetes Patients. J. Clin. Lab. Anal. 2017, 31, e22039. [Google Scholar] [CrossRef]
- Akhter, F.; Khan, M.S.; Ahmad, S. Acquired Immunogenicity of Calf Thymus DNA and LDL Modified by D-Ribose: A Comparative Study. Int. J. Biol. Macromol. 2015, 72, 1222–1227. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Irace, G.; Sirangelo, I. Role of Glycation in Amyloid: Effect on the Aggregation Process and Cytotoxicity. In Exploring New Findings on Amyloidosis; Fernandez-Escamilla, A.M., Ed.; IntechOpen: London, UK, 2016; ISBN 978-953-51-2618-8. [Google Scholar]
- Javed, M.; Ahmad, M.I.; Javed, H.; Naseem, S. D-Ribose and Pathogenesis of Alzheimer’s Disease. Mol. Biol. Rep. 2020, 47, 2289–2299. [Google Scholar] [CrossRef]
- Chen, L.; Wei, Y.; Wang, X.; He, R. D-Ribosylated Tau Forms Globular Aggregates with High Cytotoxicity. Cell. Mol. Life Sci. 2009, 66, 2559–2571. [Google Scholar] [CrossRef]
- Kong, F.-L.; Cheng, W.; Chen, J.; Liang, Y. D-Ribose Glycates Β2-Microglobulin to Form Aggregates with High Cytotoxicity through a ROS-Mediated Pathway. Chem. Biol. Interact. 2011, 194, 69–78. [Google Scholar] [CrossRef]
- Wei, Y.; Han, C.; Wang, Y.; Wu, B.; Su, T.; Liu, Y.; He, R. Ribosylation Triggering Alzheimer’s Disease-like Tau Hyperphosphorylation via Activation of CaMKII. Aging Cell 2015, 14, 754–763. [Google Scholar] [CrossRef]
- Wu, B.; Wang, Y.; Shi, C.; Chen, Y.; Yu, L.; Li, J.; Li, W.; Wei, Y.; He, R. Ribosylation-Derived Advanced Glycation End Products Induce Tau Hyperphosphorylation Through Brain-Derived Neurotrophic Factor Reduction. J. Alzheimer Dis. 2019, 71, 291–305. [Google Scholar] [CrossRef]
- Farzadfard, A.; König, A.; Petersen, S.V.; Nielsen, J.; Vasili, E.; Dominguez-Meijide, A.; Buell, A.K.; Outeiro, T.F.; Otzen, D.E. Glycation Modulates Alpha-Synuclein Fibrillization Kinetics: A Sweet Spot for Inhibition. J. Biol. Chem. 2022, 298, 101848. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, Y.; Wang, X.; He, R. Ribosylation Rapidly Induces α-Synuclein to Form Highly Cytotoxic Molten Globules of Advanced Glycation End Products. PLoS ONE 2010, 5, e9052. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Lu, Y.; Wei, Y.; Wu, B.; Liu, Y.; He, R. D-Ribosylation Induces Cognitive Impairment through RAGE-Dependent Astrocytic Inflammation. Cell Death Dis. 2014, 5, e1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Lu, Y.; Wei, Y.; Liu, Y.; He, R. D-Ribose Induces Cellular Protein Glycation and Impairs Mouse Spatial Cognition. PLoS ONE 2011, 6, e24623. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Chen, Y.; Xu, Y.; He, T.; Wei, Y.; He, R. D-Ribose Is Elevated in T1DM Patients and Can Be Involved in the Onset of Encephalopathy. Aging 2019, 11, 4943–4969. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, H.; Zhang, H.; Li, R.; Zhang, Q.; Luo, D.; Cai, X.; Li, M. Serum Oxidized Low Density Lipoprotein Serves as a Mediator for the Inverse Relationship between Serum D-Ribose and Cognitive Performance in Type 2 Diabetic Patients. Free Radic. Biol. Med. 2021, 171, 91–98. [Google Scholar] [CrossRef]
- Xu, K.; Wang, M.; Zhou, W.; Pu, J.; Wang, H.; Xie, P. Chronic D-Ribose and D-Mannose Overload Induce Depressive/Anxiety-like Behavior and Spatial Memory Impairment in Mice. Transl. Psychiatry 2021, 11, 90. [Google Scholar] [CrossRef]
- Wu, B.; Wei, Y.; Wang, Y.; Su, T.; Zhou, L.; Liu, Y.; He, R. Gavage of D-Ribose Induces Aβ-like Deposits, Tau Hyperphosphorylation as Well as Memory Loss and Anxiety-like Behavior in Mice. Oncotarget 2015, 6, 34128–34142. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Li, G.; Zhang, Q.; Ritter, J.; Li, W.; Li, P.-L. D-Ribose Induces Podocyte NLRP3 Inflammasome Activation and Glomerular Injury via AGEs/RAGE Pathway. Front. Cell Dev. Biol. 2019, 7, 259. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Wang, X.; Zhang, N.; Fu, H.; Li, W. D-Ribose Induces Nephropathy through RAGE-Dependent NF-ΚB Inflammation. Arch. Pharm. Res. 2018, 41, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhao, S.; Hong, J.; Li, W.; Wang, X. Protective Effects of Kaempferol on D-Ribose-Induced Mesangial Cell Injury. Oxidative Med. Cell. Longev. 2019, 2019, 7564207. [Google Scholar] [CrossRef] [PubMed]
- Vashishth, D.; Gibson, G.J.; Khoury, J.I.; Schaffler, M.B.; Kimura, J.; Fyhrie, D.P. Influence of Nonenzymatic Glycation on Biomechanical Properties of Cortical Bone. Bone 2001, 28, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Gong, H.; Cen, H.; Shi, P.; Zhang, R.; Li, Z.; Bi, X. Influence of Non-Enzymatic Glycation on the Mechanical Properties of Cortical Bone. J. Mech. Behav. Biomed. Mater. 2021, 119, 104553. [Google Scholar] [CrossRef]
- Lee, J.M.; Veres, S.P. Advanced Glycation End-Product Cross-Linking Inhibits Biomechanical Plasticity and Characteristic Failure Morphology of Native Tendon. J. Appl. Physiol. 2019, 126, 832–841. [Google Scholar] [CrossRef]
- De Caro, L.; Terzi, A.; Fusaro, L.; Altamura, D.; Boccafoschi, F.; Bunk, O.; Giannini, C. Time Scale of Glycation in Collagen of Bovine Pericardium-Derived Bio-Tissues. IUCrJ 2021, 8, 1024–1034. [Google Scholar] [CrossRef]
- Kajimoto, Y.; Matsuoka, T.; Kaneto, H.; Watada, H.; Fujitani, Y.; Kishimoto, M.; Sakamoto, K.; Matsuhisa, M.; Kawamori, R.; Yamasaki, Y.; et al. Induction of Glycation Suppresses Glucokinase Gene Expression in HIT-T15 Cells. Diabetologia 1999, 42, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Waris, S.; Pischetsrieder, M.; Saleemuddin, M. DNA Damage by Ribose: Inhibition at High Ribose Concentrations. Indian J. Biochem. Biophys. 2010, 47, 148–156. [Google Scholar]
- Akhter, F.; Salman Khan, M.; Shahab, U.; Moinuddin; Ahmad, S. Bio-Physical Characterization of Ribose Induced Glycation: A Mechanistic Study on DNA Perturbations. Int. J. Biol. Macromol. 2013, 58, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Chandra, G.K.; Eklouh-Molinier, C.; Fere, M.; Angiboust, J.-F.; Gobinet, C.; Van-Gulick, L.; Jeannesson, P.; Piot, O. Probing in Vitro Ribose Induced DNA-Glycation Using Raman Microspectroscopy. Anal. Chem. 2015, 87, 2655–2664. [Google Scholar] [CrossRef]
- Dinda, A.K.; Tripathy, D.R.; Dasgupta, S. Glycation of Ribonuclease A Affects Its Enzymatic Activity and DNA Binding Ability. Biochimie 2015, 118, 162–172. [Google Scholar] [CrossRef]
- Su, T.; Xin, L.; He, Y.-G.; Wei, Y.; Song, Y.-X.; Li, W.-W.; Wang, X.-M.; He, R.-Q. The Abnormally High Level of Uric D-Ribose for Type-2 Diabetics. Acta Agron. Sin. 2013, 40, 816. [Google Scholar] [CrossRef]
- Siddiqui, Z.; Faisal, M.; Alatar, A.R.; Ahmad, S. Prevalence of Auto-Antibodies against D-Ribose-Glycated-Hemoglobin in Diabetes Mellitus. Glycobiology 2019, 29, 409–418. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The oxidative branch of PPP. Alterations in enzymatic activities are connected and/or ob–served in the indicated pathological and stressful conditions, as mentioned in the text.
Figure 1.
The oxidative branch of PPP. Alterations in enzymatic activities are connected and/or ob–served in the indicated pathological and stressful conditions, as mentioned in the text.

Figure 2.
The reversible branch of PPP. The links among the reversible branch of PPP and relevant metabolic pathways are highlighted. Ru5PI, Ru5P-isomerase; Ru5PE, Ru5P-epimerase; TKT, trans–ketolase; TAL, transaldolase.
Figure 2.
The reversible branch of PPP. The links among the reversible branch of PPP and relevant metabolic pathways are highlighted. Ru5PI, Ru5P-isomerase; Ru5PE, Ru5P-epimerase; TKT, trans–ketolase; TAL, transaldolase.

Figure 3.
Schematic representation of AGEs generation upon protein glycation induced by D-ribose. The same reactions progression may apply to R5P, except for the pyranosidic structure of the sugar phosphate, for the cyclic N-substituted-1-C- ribopyranoside and then for the cyclic N-substituted-1-C-ribulofuranoside.
Figure 3.
Schematic representation of AGEs generation upon protein glycation induced by D-ribose. The same reactions progression may apply to R5P, except for the pyranosidic structure of the sugar phosphate, for the cyclic N-substituted-1-C- ribopyranoside and then for the cyclic N-substituted-1-C-ribulofuranoside.

Figure 4.
Articles availability along the past forty years. Numbers emerge from the PubMed® data base, searching for: “glycation and ribose” on August 2022.
Figure 4.
Articles availability along the past forty years. Numbers emerge from the PubMed® data base, searching for: “glycation and ribose” on August 2022.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moschini, R.; Balestri, F.; Cappiello, M.; Signore, G.; Mura, U.; Del-Corso, A. Ribose Intake as Food Integrator: Is It a Really Convenient Practice? Biomolecules 2022, 12, 1775. https://doi.org/10.3390/biom12121775
AMA Style
Moschini R, Balestri F, Cappiello M, Signore G, Mura U, Del-Corso A. Ribose Intake as Food Integrator: Is It a Really Convenient Practice? Biomolecules. 2022; 12(12):1775. https://doi.org/10.3390/biom12121775
Chicago/Turabian StyleMoschini, Roberta, Francesco Balestri, Mario Cappiello, Giovanni Signore, Umberto Mura, and Antonella Del-Corso. 2022. "Ribose Intake as Food Integrator: Is It a Really Convenient Practice?" Biomolecules 12, no. 12: 1775. https://doi.org/10.3390/biom12121775
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.