
The Readthrough Isoform AQP4ex Is Constitutively Phosphorylated in the Perivascular Astrocyte Endfeet of Human Brain

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Multiple Alignment of Predicted Aquaporin Transcripts from Different Vertebrates and Mammals
2.2. Cell Cultures and Transfection
2.3. Antibodies
2.4. Human Brain Biopsies
2.5. Immunofluorescence
2.5.1. Human Brain Tissue
2.5.2. Mammalian Cells
2.6. Sample Preparation for SDS-PAGE
2.7. Sample Preparation for BN-PAGE
2.8. SDS-PAGE and Western Blot Analysis
3. Results
3.1. Multiple Alignment Analysis of AQP4 Extension
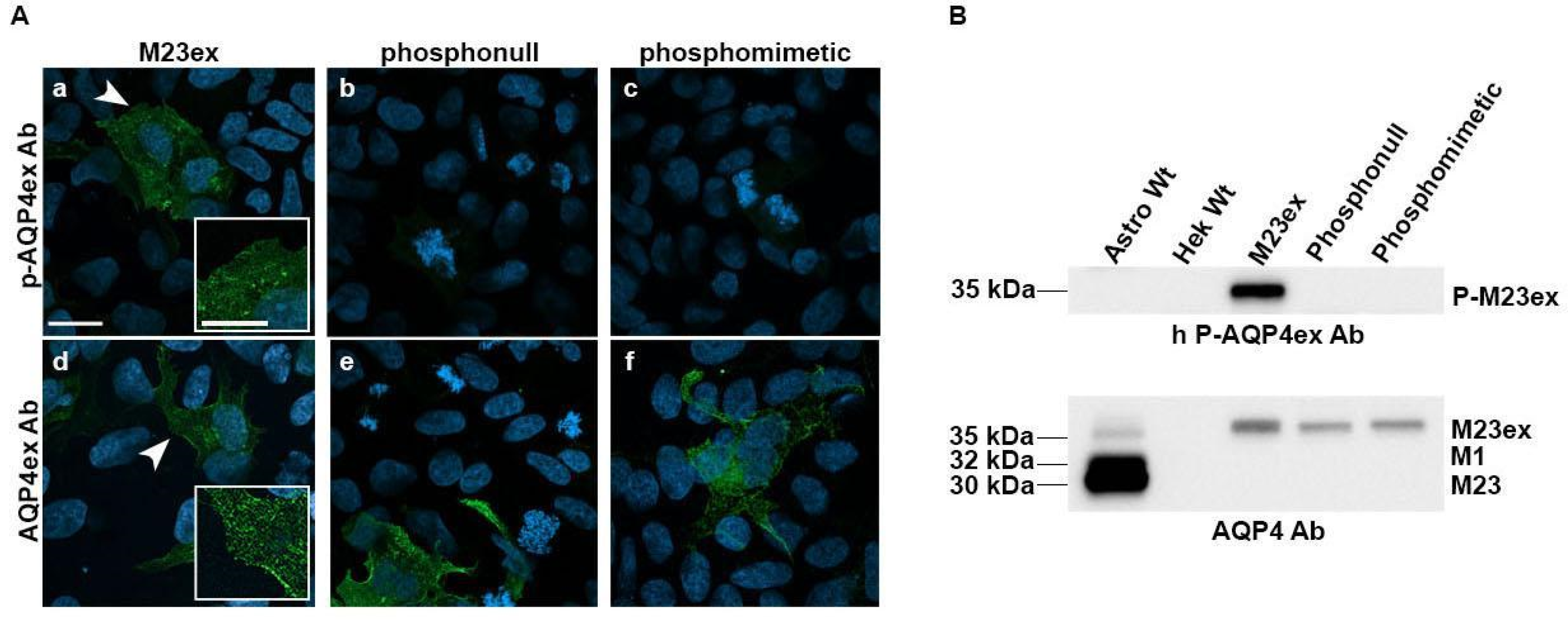
3.2. Characterization of Antibodies on p-Ser335 Using Transfected Cells
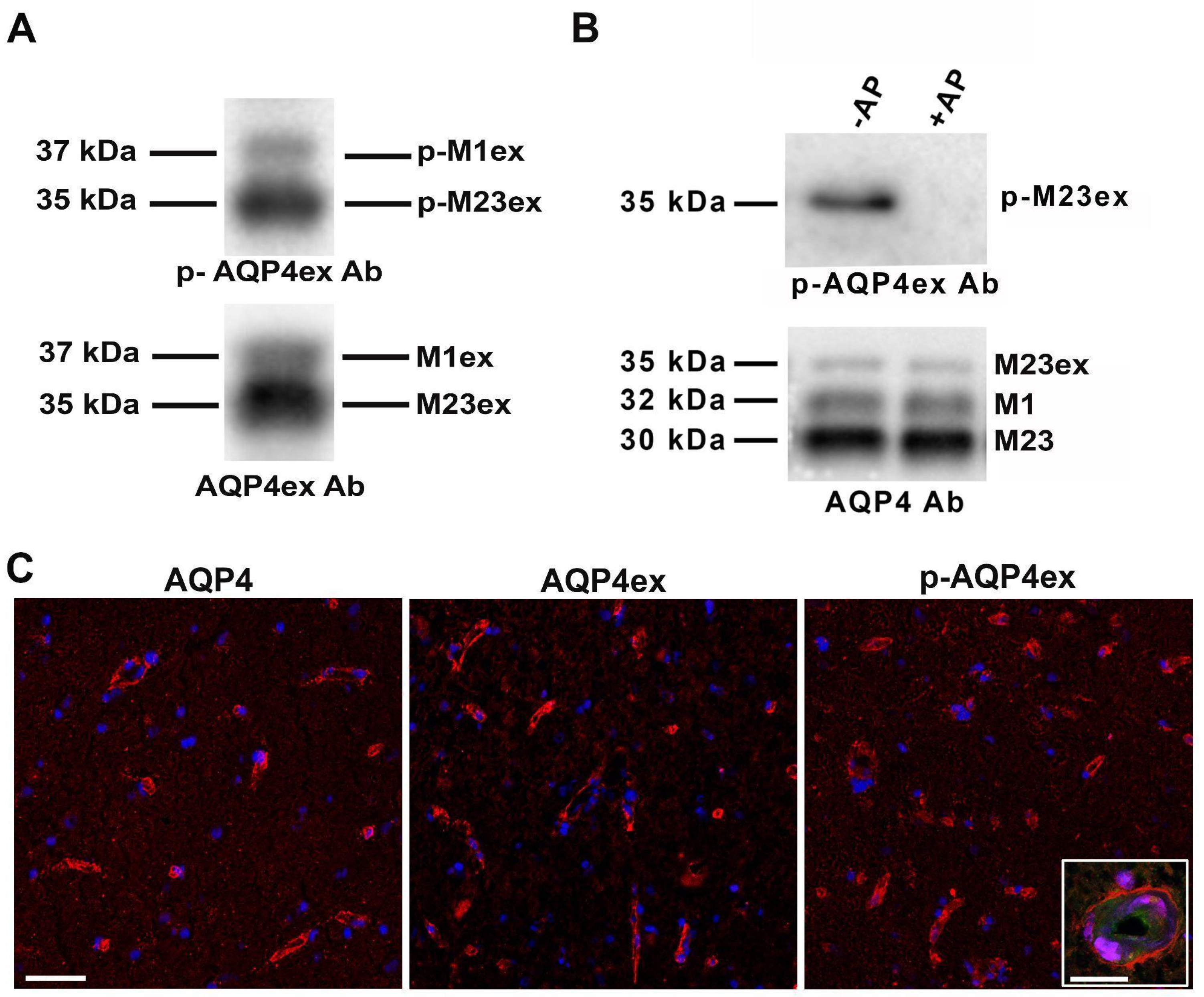
3.3. p-AQP4ex Expression in Human Brain
3.4. p-AQP4ex Supramolecular Organization in Human Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frigeri, A.; Nicchia, G.P.; Svelto, M. Aquaporins as targets for drug discovery. Curr. Pharm. Des. 2007, 13, 2421–2427. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Nico, B.; Camassa, L.; Mola, M.; Loh, N.; Dermietzel, R.; Spray, D.; Svelto, M.; Frigeri, A. The role of aquaporin-4 in the blood-brain barrier development and integrity: Studies in animal and cell culture models. Neuroscience 2004, 129, 935–945. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Krishna, S.; Verkman, A.S. Aquaporin water channels and brain edema. Mt. Sinai J. Med. 2002, 69, 242–248. [Google Scholar] [PubMed]
- Iliff, J.J.; Chen, M.J.; Plog, B.A.; Zeppenfeld, D.M.; Soltero, M.; Yang, L.; Singh, I.; Deane, R.; Nedergaard, M. Impairment of glymphatic pathway function promotes tau pathology after traumatic brain injury. J. Neurosci. 2014, 34, 16180–16193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.; Pisani, F.; Nicchia, G.P.; Svelto, M.; Frigeri, A. Evidences for a leaky scanning mechanism for the synthesis of the shorter M23 protein isoform of aquaporin-4: Implication in orthogonal array formation and neuromyelitis optica antibody interaction. J. Biol. Chem. 2010, 285, 4562–4569. [Google Scholar] [CrossRef] [Green Version]
- Jin, B.J.; Rossi, A.; Verkman, A.S. Model of aquaporin-4 supramolecular assembly in orthogonal arrays based on heterotetrameric association of M1-M23 isoforms. Biophys. J. 2011, 100, 2936–2945. [Google Scholar] [CrossRef] [Green Version]
- Nicchia, G.P.; Rossi, A.; Mola, M.; Pisani, F.; Stigliano, C.; Basco, D.; Mastrototaro, M.; Svelto, M.; Frigeri, A. Higher order structure of aquaporin-4. Neuroscience 2010, 168, 903–914. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Cogotzi, L.; Rossi, A.; Basco, D.; Brancaccio, A.; Svelto, M.; Frigeri, A. Expression of multiple AQP4 pools in the plasma membrane and their association with the dystrophin complex. J. Neurochem. 2008, 105, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- Nicchia, G.P.; Ficarella, R.; Rossi, A.; Giangreco, I.; Nicolotti, O.; Carotti, A.; Pisani, F.; Estivill, X.; Gasparini, P.; Svelto, M.; et al. D184E mutation in aquaporin-4 gene impairs water permeability and links to deafness. Neuroscience 2011, 197, 80–88. [Google Scholar] [CrossRef]
- Wolburg, H.; Wolburg-Buchholz, K.; Fallier-Becker, P.; Noell, S.; Mack, A.F. Structure and functions of aquaporin-4-based orthogonal arrays of particles. Int. Rev. Cell Mol. Biol. 2011, 287, 1–41. [Google Scholar] [PubMed]
- Loughran, G.; Chou, M.-Y.; Ivanov, I.P.; Jungreis, I.; Kellis, M.; Kiran, A.M.; Baranov, P.V.; Atkins, J.F. Evidence of efficient stop codon readthrough in four mammalian genes. Nucleic Acids Res. 2014, 42, 8928–8938. [Google Scholar] [CrossRef] [Green Version]
- De Bellis, M.; Pisani, F.; Mola, M.G.; Rosito, S.; Simone, L.; Buccoliero, C.; Trojano, M.; Nicchia, G.P.; Svelto, M.; Frigeri, A. Translational readthrough generates new astrocyte AQP4 isoforms that modulate supramolecular clustering, glial endfeet localization, and water transport. Glia 2017, 65, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, C.; Abbrescia, P.; Valente, O.; Nicchia, G.P.; Banitalebi, S.; Amiry-Moghaddam, M.; Trojano, M.; Frigeri, A. Tissue Distribution of the Readthrough Isoform of AQP4 Reveals a Dual Role of AQP4ex Limited to CNS. Int. J. Mol. Sci. 2020, 21, 1531. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, C.; Buccoliero, C.; Mola, M.G.; Abbrescia, P.; Nicchia, G.P.; Trojano, M.; Frigeri, A. AQP4ex is crucial for the anchoring of AQP4 at the astrocyte end-feet and for neuromyelitis optica antibody binding. Acta Neuropathol. Commun. 2019, 7, 51. [Google Scholar] [CrossRef]
- Nesverova, V.; Törnroth-Horsefield, S. Phosphorylation-Dependent Regulation of Mammalian Aquaporins. Cells 2019, 8, 82. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P. The origins of protein phosphorylation. Nat. Cell Biol. 2002, 4, E127–E130. [Google Scholar] [CrossRef]
- Vandebroek, A.; Yasui, M. Regulation of AQP4 in the Central Nervous System. Int. J. Mol. Sci. 2020, 21, 1603. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, D.L.; Barrett, T.; Benson, D.A.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Edgar, R.; Federhen, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2007, 35, D5–D12. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. The Clustal Omega Multiple Alignment Package. Methods Mol. Biol. 2021, 2231, 3–16. [Google Scholar]
- Nicchia, G.P.; Frigeri, A.; Liuzzi, G.M.; Svelto, M. Inhibition of aquaporin-4 expression in astrocytes by RNAi determines alteration in cell morphology, growth, and water transport and induces changes in ischemia-related genes. FASEB J. 2003, 17, 1508–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, O.; Messina, R.; Ingravallo, G.; Bellitti, E.; Zimatore, D.S.; de Gennaro, L.; Abbrescia, P.; Pati, R.; Palazzo, C.; Nicchia, G.P.; et al. Alteration of the translational readthrough isoform AQP4ex induces redistribution and downregulation of AQP4 in human glioblastoma. Cell Mol. Life Sci. 2022, 79, 140. [Google Scholar] [CrossRef] [PubMed]
- Pisani, F.; Sparaneo, A.; Tortorella, C.; Ruggieri, M.; Trojano, M.; Mola, M.G.; Nicchia, G.P.; Frigeri, A.; Svelto, M. Aquaporin-4 autoantibodies in Neuromyelitis Optica: AQP4 isoform-dependent sensitivity and specificity. PLoS ONE 2013, 8, e79185. [Google Scholar] [CrossRef] [PubMed]
- Nicchia, G.P.; Mastrototaro, M.; Rossi, A.; Pisani, F.; Tortorella, C.; Ruggieri, M.; Lia, A.; Trojano, M.; Frigeri, A.; Svelto, M. Aquaporin-4 orthogonal arrays of particles are the target for neuromyelitis optica autoantibodies. Glia 2009, 57, 1363–1373. [Google Scholar] [CrossRef]
- Dephoure, N.; Zhou, C.; Villen, J.; Beausoleil, S.A.; Bakalarski, C.E.; Elledge, S.J.; Gygi, S.P. A quantitative atlas of mitotic phosphorylation. Proc. Natl. Acad. Sci. USA 2008, 105, 10762–10767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadohira, I.; Abe, Y.; Nuriya, M.; Sano, K.; Tsuji, S.; Arimitsu, T.; Yoshimura, Y.; Yasui, M. Phosphorylation in the C-terminal domain of Aquaporin-4 is required for Golgi transition in primary cultured astrocytes. Biochem. Biophys. Res. Commun. 2008, 377, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Gunnarson, E.; Zelenina, M.; Aperia, A. Regulation of brain aquaporins. Neuroscience 2004, 129, 947–955. [Google Scholar] [CrossRef]
- Fazzina, G.; Amorini, A.M.; Marmarou, C.R.; Fukui, S.; Okuno, K.; Dunbar, J.G.; Glisson, R.; Marmarou, A.; Kleindienst, A. The protein kinase C activator phorbol myristate acetate decreases brain edema by aquaporin 4 downregulation after middle cerebral artery occlusion in the rat. J. Neurotrauma 2010, 27, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Assentoft, M.; Kaptan, S.; Fenton, R.A.; Hua, S.Z.; De Groot, B.L.; Macaulay, N. Phosphorylation of rat aquaporin-4 at Ser(111) is not required for channel gating. Glia 2013, 61, 1101–1112. [Google Scholar] [CrossRef] [Green Version]
- Carmosino, M.; Procino, G.; Svelto, M. Na+-K+-2Cl- cotransporter type 2 trafficking and activity: The role of interacting proteins. Biol. Cell 2012, 104, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Madrid, R.; Le Maout, S.; Barrault, M.; Janvier, K.; Benichou, S.; Mérot, J. Polarized trafficking and surface expression of the AQP4 water channel are coordinated by serial and regulated interactions with different clathrin-adaptor complexes. EMBO J. 2001, 20, 7008–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; Macdonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799.e19. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.; Beltrao, P. Evolution of protein kinase substrate recognition at the active site. PLoS Biol. 2019, 17, e3000341. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, D.; Lake, A.M.; Yang, W.; Yang, C.; Wesseling, H.; Guise, A.; Uncu, C.; Dalal, J.S.; Kraft, A.W.; Lee, J.-M.; et al. Cell-Type-Specific Profiling of Alternative Translation Identifies Regulated Protein Isoform Variation in the Mouse Brain. Cell Rep. 2019, 26, 594–607.e7. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (ng/mL) | NC | 1000.00 | 500.00 | 250.00 | 125.00 | 62.50 | 31.25 | 15.62 | 7.81 | 3.90 | 1.95 | Blank | / | / | |
| Diluition | 1:1000 | 1:1000 | 1:2000 | 1:4000 | 1:8000 | 1:16,000 | 1:32,000 | 1:64,000 | 1:128,000 | 1:256,000 | 1:512,000 | Blank | Titer | Coating | |
| Sample | |||||||||||||||
| Specific Antibody | 0.155 | 2.810 | 2.789 | 2.748 | 2.733 | 2.672 | 2.604 | 2.442 | 2.077 | 1.630 | 1.167 | 0.064 | >1:512,000 | A | |
| 0.118 | 0.528 | 0.336 | 0.200 | 0.128 | 0.094 | 0.076 | 0.075 | 0.074 | 0.067 | 0.06 | 0.059 | 1:8000 | B | ||
| “Pan” Antibody | / | 2.765 | 2.705 | 2.684 | 2.683 | 2.579 | 2.368 | 2.147 | 1.691 | 1.196 | 0.799 | 0.070 | >1:512,000 | A | |
| / | 2.786 | 2.659 | 2.613 | 2.584 | 2.491 | 2.327 | 2.045 | 1.547 | 1.127 | 0.707 | 0.066 | >1:512,000 | B | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pati, R.; Palazzo, C.; Valente, O.; Abbrescia, P.; Messina, R.; Surdo, N.C.; Lefkimmiatis, K.; Signorelli, F.; Nicchia, G.P.; Frigeri, A. The Readthrough Isoform AQP4ex Is Constitutively Phosphorylated in the Perivascular Astrocyte Endfeet of Human Brain. Biomolecules 2022, 12, 633. https://doi.org/10.3390/biom12050633
Pati R, Palazzo C, Valente O, Abbrescia P, Messina R, Surdo NC, Lefkimmiatis K, Signorelli F, Nicchia GP, Frigeri A. The Readthrough Isoform AQP4ex Is Constitutively Phosphorylated in the Perivascular Astrocyte Endfeet of Human Brain. Biomolecules. 2022; 12(5):633. https://doi.org/10.3390/biom12050633
Chicago/Turabian StylePati, Roberta, Claudia Palazzo, Onofrio Valente, Pasqua Abbrescia, Raffaella Messina, Nicoletta Concetta Surdo, Konstantinos Lefkimmiatis, Francesco Signorelli, Grazia Paola Nicchia, and Antonio Frigeri. 2022. "The Readthrough Isoform AQP4ex Is Constitutively Phosphorylated in the Perivascular Astrocyte Endfeet of Human Brain" Biomolecules 12, no. 5: 633. https://doi.org/10.3390/biom12050633