
Quantitative Trait Locus Mapping Combined with RNA Sequencing Reveals the Molecular Basis of Seed Germination in Oilseed Rape

 and
and
Abstract
:1. Introduction
2. Results
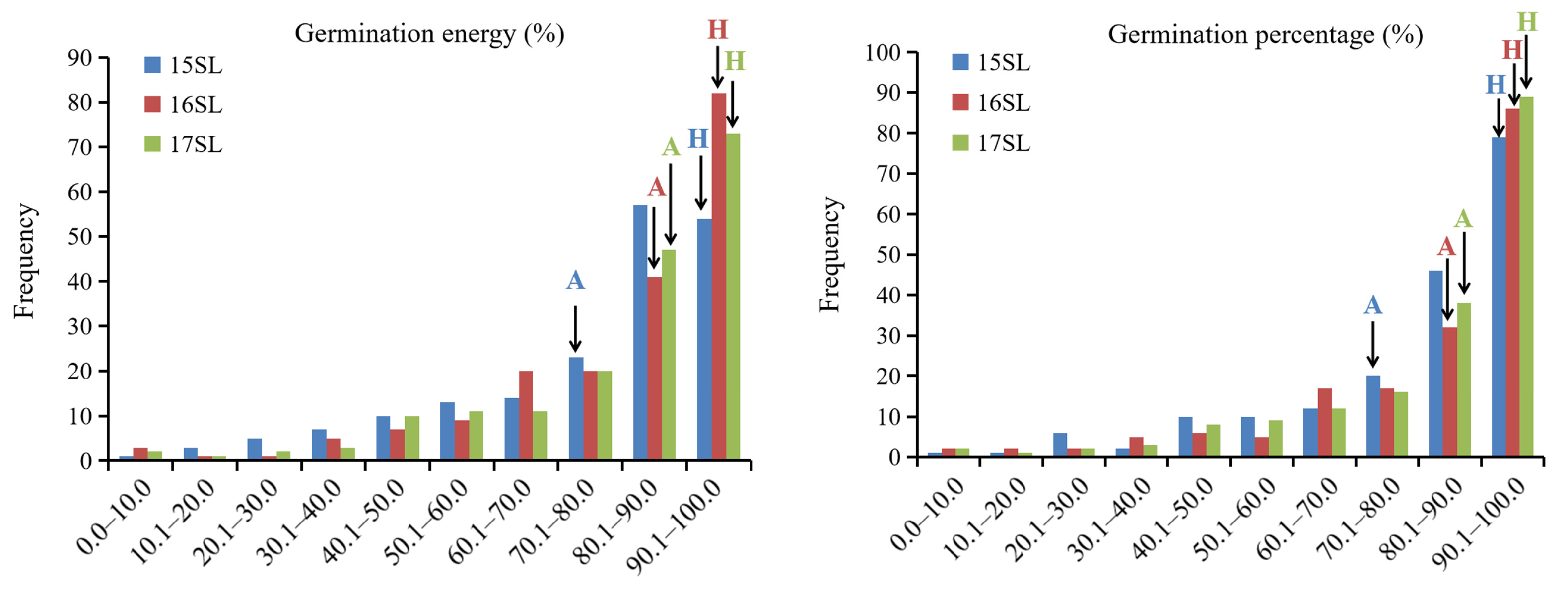
2.1. Phenotypic Variation in Germination Energy (GE) and Germination Percentage (GP)
2.2. QTL Mapping for GE and GP
2.3. Epistatic Interactions between QTLs
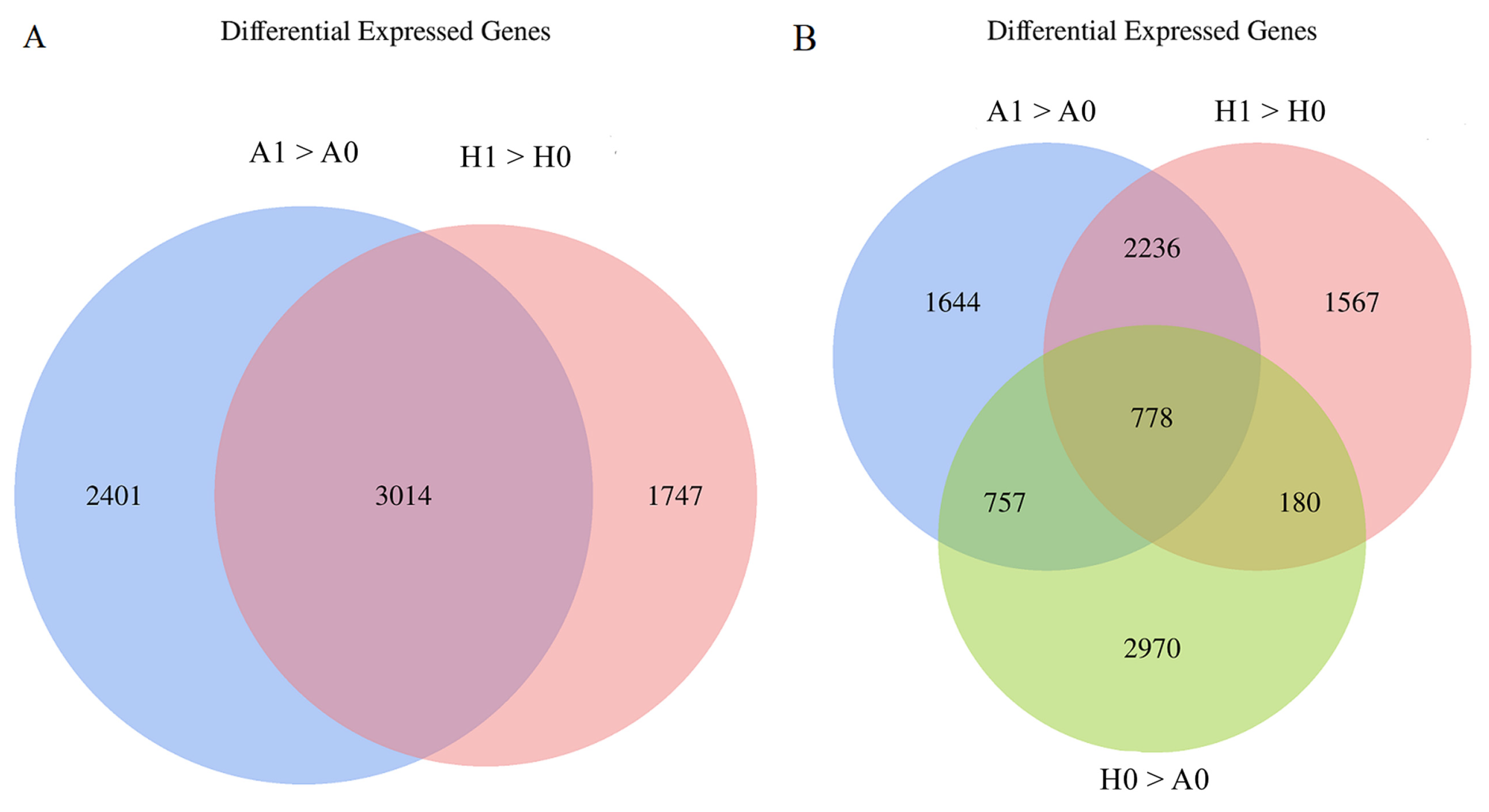
2.4. Transcriptomic Analysis of Seed Germination
2.4.1. Transcriptomic Basis of Oilseed Rape Seed Germination
2.4.2. Transcriptomic Basis of the Difference in Seed Germination between ‘APL01’ and ‘Holly’
2.5. Identification of Candidate Genes
3. Discussion
3.1. Major QTLs Couple with Minor QTLs to Co-Regulate the Germination of Brassica Napus Seeds
3.2. Epistatic Effects between QTLs Are Involved in Seed Germination
3.3. Material Metabolism and Energy Production Are the Biochemical Bases of Seed Germination in Brassica Napus
3.4. Differences in Sugar Metabolism, Protein Metabolism, Lipid Metabolism, Amino Acid Metabolism, the TCA Cycle, Electron Transfer, Signal Transduction, and DNA Metabolism Potentially Determine the Higher Germination Rate of ‘Holly’ Seeds
3.5. Genes Involved in Sugar Catabolism, Plant Hormone Signal Transduction, and Methionine Biosynthesis Are Promising Candidate Genes for Seed Germination in Brassica Napus
4. Materials and Methods
4.1. Plant Materials and Phenotyping
4.2. QTL Mapping
4.3. RNA Sequencing and Data Analysis
4.4. Mining Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar]
- Vanstraelen, M.; Benková, E. Hormonal interactions in the regulation of plant development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [Google Scholar]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar]
- Linkies, A.; Müller, K.; Morris, K.; Turecková, V.; Wenk, M.; Cadman, C.S.; Corbineau, F.; Strnad, M.; Lynn, J.R.; Finch-Savage, W.E.; et al. Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: A comparative approach using Lepidium sativum and Arabidopsis thaliana. Plant Cell 2009, 21, 3803–3822. [Google Scholar]
- Subbiah, V.; Reddy, K.J. Interactions between ethylene, abscisic acid and cytokinin during germination and seedling establishment in Arabidopsis. J. Biosci. 2010, 35, 451–458. [Google Scholar]
- Chiwocha, S.D.; Cutler, A.J.; Abrams, S.R.; Ambrose, S.J.; Yang, J.; Ross, A.R.; Kermode, A.R. The etr1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination. Plant J. 2005, 42, 35–48. [Google Scholar]
- Belin, C.; Megies, C.; Hauserova, E.; Lopez-Molina, L. Abscisic acid represses growth of the Arabidopsis embryonic axis after germination by enhancing auxin signaling. Plant Cell 2009, 21, 2253–2268. [Google Scholar]
- Preston, J.; Tatematsu, K.; Kanno, Y.; Hobo, T.; Kimura, M.; Jikumaru, Y.; Yano, R.; Kamiya, Y.; Nambara, E. Temporal expression patterns of hormone metabolism genes during imbibition of Arabidopsis thaliana seeds: A comparative study on dormant and non-dormant accessions. Plant Cell Physiol. 2009, 50, 1786–1800. [Google Scholar]
- Lee, S.; Kim, S.; Park, C. Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis. New Phytol. 2010, 188, 626–637. [Google Scholar]
- Rajjou, L.; Belghazi, M.; Huguet, R.; Robin, C.; Moreau, A.; Job, C.; Job, D. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol. 2006, 141, 910–923. [Google Scholar]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar]
- Ma, W.; Guan, X.; Li, J.; Pan, R.; Wang, L.; Liu, F.; Ma, H.; Zhu, S.; Hu, J.; Ruan, Y.-L.; et al. Mitochondrial small heat shock protein mediates seed germination via thermal sensing. Proc. Natl. Acad. Sci. USA 2019, 116, 4716–4721. [Google Scholar]
- Barba-Espín, G.; Nicolas, E.; Almansa, M.S.; Cantero-Navarro, E.; Albacete, A.; Hernández, J.A.; Díaz-Vivancos, P. Role of thioproline on seed germination: Interaction ROS-ABA and effects on antioxidative metabolism. Plant Physiol. Biochem. 2012, 59, 30–36. [Google Scholar]
- Bahin, E.; Bailly, C.; Sotta, B.; Kranner, I.; Corbineau, F.; Leymarie, J. Crosstalk between reactive oxygen species and hormonal signalling pathways regulates grain dormancy in barley. Plant Cell Environ. 2011, 34, 980–993. [Google Scholar]
- Ishibashi, Y.; Koda, Y.; Zheng, S.-H.; Yuasa, T.; Iwaya-Inoue, M. Regulation of soybean seed germination through ethylene production in response to reactive oxygen species. Ann. Bot. 2013, 111, 95–102. [Google Scholar]
- Diaz-Vivancos, P.; Barba-Espín, G.; Hernández, J.A. Elucidating hormonal/ROS networks during seed germination: Insights and perspectives. Plant Cell Rep. 2013, 32, 1491–1502. [Google Scholar]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; VandeKerckhove, J.; Job, D. Importance of methionine biosynthesis for Arabidopsis seed germination and seedling growth. Physiol. Plant. 2002, 116, 238–247. [Google Scholar]
- Ju, C.; Kong, D.; Lee, Y.; Ge, G.; Song, Y.; Liu, J.; Kwak, J.M. Methionine synthase 1 provides methionine for activation of the GLR3.5 Ca2+ channel and regulation of germination in Arabidopsis. J. Exp. Bot. 2020, 71, 178–187. [Google Scholar]
- Kim, J.Y.; Kwak, K.J.; Jung, H.J.; Lee, H.J.; Kang, H. MicroRNA402 affects seed germination of Arabidopsis thaliana under stress conditions via targeting DEMETER-LIKE Protein3 mRNA. Plant Cell Physiol. 2010, 51, 1079–1083. [Google Scholar]
- Nakabayashi, K.; Okamoto, M.; Koshiba, T.; Kamiya, Y.; Nambara, E. Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: Epigenetic and genetic regulation of transcription in seed. Plant J. 2005, 41, 697–709. [Google Scholar]
- Min, S.S.; Dong, G.K.; Sun, H.L. Isolation and characterization of a jasmonic acid carboxyl methyltransferase gene from hot pepper (Capsicum annuum L.). J. Plant Biol. 2005, 48, 292–297. [Google Scholar]
- Zhao, N.; Ferrer, J.-L.; Ross, J.; Guan, J.; Yang, Y.; Pichersky, E.; Noel, J.P.; Chen, F. Structural, biochemical, and phylogenetic analyses suggest that indole-3-acetic acid methyltransferase is an evolutionarily ancient member of the SABATH family. Plant Physiol. 2008, 146, 455–467. [Google Scholar]
- Varbanova, M.; Yamaguchi, S.; Yang, Y.; McKelvey, K.; Hanada, A.; Borochov, R.; Yu, F.; Jikumaru, Y.; Ross, J.; Cortes, D.; et al. Methylation of gibberellins by Arabidopsis GAMT1 and GAMT2. Plant Cell 2007, 19, 32–45. [Google Scholar]
- Ravanel, S.; Gakière, B.; Job, D.; Douce, R. The specific features of methionine biosynthesis and metabolism in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 7805–7812. [Google Scholar]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar]
- Ventura, L.; Donà, M.; Macovei, A.; Carbonera, D.; Buttafava, A.; Mondoni, A.; Rossi, G.; Balestrazzi, A. Understanding the molecular pathways associated with seed vigor. Plant Physiol. Biochem. 2012, 60, 196–206. [Google Scholar]
- Corbineau, F. Markers of seed quality: From present to future. Seed Sci. Res. 2012, 22, S61–S68. [Google Scholar]
- Bønsager, B.C.; Shahpiri, A.; Finnie, C.; Svensson, B. Proteomic and activity profiles of ascorbate-glutathione cycle enzymes in germinating barley embryo. Phytochemistry 2010, 71, 1650–1656. [Google Scholar]
- Bethke, P.C.; Libourel, I.G.L.; Jones, R.L. Nitric oxide reduces seed dormancy in Arabidopsis. J. Exp. Bot. 2006, 57, 517–526. [Google Scholar]
- Basnet, R.K.; Duwal, A.; Tiwari, D.N.; Xiao, D.; Monakhos, S.; Bucher, J.; Visser, R.G.F.; Groot, S.P.C.; Bonnema, A.B.; Maliepaard, C.A. Quantitative trait locus analysis of seed germination and seedling vigour in Brassica rapa reveals QTL hotspots and epistatic interactions. Front. Plant Sci. 2015, 6, 1032. [Google Scholar]
- Hatzig, S.V.; Frisch, M.; Breuer, F.; Nesi, N.; Ducournau, S.; Wagner, M.-H.; Leckband, G.; Abbadi, A.; Snowdon, R.J. Genome-wide association mapping unravels the genetic control of seed germination and vigor in Brassica napus. Front. Plant Sci. 2015, 6, 221. [Google Scholar]
- Gu, J.; Chao, H.; Gan, L.; Guo, L.; Zhang, K.; Li, Y.; Wang, H.; Raboanatahiry, N.; Li, M. Proteomic dissection of seed germination and seedling establishment in Brassica napus. Front. Plant Sci. 2016, 7, 1482. [Google Scholar]
- Nguyen, T.C.T.; Abrams, S.R.; Friedt, W.; Snowdon, R.J. Quantitative trait locus analysis of seed germination, seedling vigour and seedling-regulated hormones in Brassica napus. Plant Breed. 2018, 137, 388–401. [Google Scholar]
- Boter, M.; Calleja-Cabrera, J.; Carrera-Castaño, G.; Wagner, G.; Hatzig, S.V.; Snowdon, R.J.; Legoahec, L.; Bianchetti, G.; Bouchereau, A.; Nesi, N.; et al. An integrative approach to analyze seed germination in Brassica napus. Front. Plant Sci. 2019, 10, 1342. [Google Scholar]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 5th ed.; Worth Publishers: New York, NY, USA, 2008; pp. 238–272. [Google Scholar]
- Körner, M. Identifizierung und Erste Charakterisierung Einer Neuen Triacylglyceridlipase-Familie aus Arabidopsis thaliana. Ph.D. Thesis, Martin-Luther-Universität Halle-Wittenberg, Sachsen-Anhalt, Germany, 2005. [Google Scholar]
- Eastmond, P.J. Glycerol-insensitive Arabidopsis mutants: gli1 seedlings lack glycerol kinase, accumulate glycerol and are more resistant to abiotic stress. Plant J. 2004, 37, 617–625. [Google Scholar]
- Penfield, S.; Rylott, E.L.; Gilday, A.D.; Graham, S.; Larson, T.R.; Graham, I.A. Reserve mobilization in the Arabidopsis endosperm fuels hypocotyl elongation in the dark, is independent of abscisic acid, and requires PHOSPHOENOLPYRUVATE CARBOXYKINASE1. Plant Cell 2004, 16, 2705–2718. [Google Scholar]
- Holdsworth, M.J.; Bentsink, L.; Soppe, W.J.J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol. 2008, 179, 33–54. [Google Scholar]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Ubeda-Tomás, S.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.-A.; Taylor, I.; et al. The N-end rule pathway promotes seed germination and establishment through removal of ABA sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar]
- Obroucheva, N.V.; Antipova, O.V. Physiology of the initiation of seed germination. Russ. J. Plant Physiol. 1997, 44, 250–264. [Google Scholar]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Good, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant J. 2007, 49, 1108–1121. [Google Scholar]
- Rocha, M.; Licausi, F.; Araújo, W.L.; Nunes-Nesi, A.; Sodek, L.; Fernie, A.R.; van Dongen, J.T. Glycolysis and the tricarboxylic acid cycle are linked by alanine aminotransferase during hypoxia induced by waterlogging of Lotus japonicus. Plant Physiol. 2010, 152, 1501–1513. [Google Scholar]
- Bettey, M.; Finch-Savage, W.E. Respiratory enzyme activities during germination in Brassica seed lots of differing vigour. Seed Sci. Res. 1996, 6, 165–174. [Google Scholar]
- Wakao, S.; Andre, C.; Benning, C. Functional analyses of cytosolic glucose-6-phosphate dehydrogenases and their contribution to seed oil accumulation in Arabidopsis. Plant Physiol. 2008, 146, 277–288. [Google Scholar]
- Smiri, M.; Chaoui, A.; El Ferjani, E. Respiratory metabolism in the embryonic axis of germinating pea seed exposed to cadmium. J. Plant Physiol. 2009, 166, 259–269. [Google Scholar]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol. 2001, 126, 835–848. [Google Scholar]
- Fu, Q.; Wang, B.-C.; Jin, X.; Li, H.-B.; Han, P.; Wei, K.-H.; Zhang, X.-M.; Zhu, Y.-X. Proteomic analysis and extensive protein identification from dry, germinating Arabidopsis seeds and young seedlings. J. Biochem. Mol. Biol. 2005, 38, 650–660. [Google Scholar]
- Müller, K.; Job, C.; Belghazi, M.; Job, D.; Leubner-Metzger, G. Proteomics reveal tissue-specific features of the cress (Lepidium sativum L.) endosperm cap proteome and its hormone-induced changes during seed germination. Proteomics 2010, 10, 406–416. [Google Scholar]
- Rajjou, L.; Gallardo, K.; Debeaujon, I.; Vandekerckhove, J.; Job, C.; Job, D. The effect of α-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination. Plant Physiol. 2004, 134, 1598–1613. [Google Scholar]
- Hong, Y.; Zhao, J.; Guo, L.; Kim, S.-C.; Deng, X.; Wang, G.; Zhang, G.; Li, M.; Wang, X. Plant phospholipases D and C and their diverse functions in stress responses. Prog. Lipid Res. 2016, 62, 55–74. [Google Scholar]
- Li, G.; Lin, F.; Xue, H.-W. Genome-wide analysis of the phospholipase D family in Oryza sativa and functional characterization of PLDβ1 in seed germination. Cell Res. 2007, 17, 881–894. [Google Scholar]
- Sagar, S.; Singh, A. Emerging role of phospholipase C mediated lipid signaling in abiotic stress tolerance and development in plants. Plant Cell Rep. 2021, 40, 2123–2133. [Google Scholar]
- Uraji, M.; Katagiri, T.; Okuma, E.; Ye, W.; Hossain, M.A.; Masuda, C.; Miura, A.; Nakamura, Y.; Mori, I.C.; Shinozaki, K.; et al. Cooperative function of PLDδ and PLDα1 in abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol. 2012, 159, 450–460. [Google Scholar]
- Yang, P.J. Cloning of Phospholipase Dγ (CbPLDγ) Gene in Chorispora bungeana and Its Function Responding to Drought Stress. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2018. [Google Scholar]
- Ariizumi, T.; Hauvermale, A.L.; Nelson, S.K.; Hanada, A.; Yamaguchi, S.; Steber, C.M. Lifting DELLA repression of Arabidopsis seed germination by nonproteolytic gibberellin signaling. Plant Physiol. 2013, 162, 2125–2139. [Google Scholar]
- Weissenbock, G. Flavonoid metabolism during seed germination. Ber. Dtsch. Bot. Gesellscaft 1970, 63, 131–135. [Google Scholar]
- Liu, R.; Lu, J.; Xing, J.; Du, M.; Wang, M.; Zhang, L.; Li, Y.; Zhang, C.; Wu, Y. Transcriptome and metabolome analyses revealing the potential mechanism of seed germination in Polygonatum cyrtonema. Sci. Rep. 2021, 11, 12161. [Google Scholar]
- Steber, C.M.; Mccourt, P. A role for brassinosteroids in germination in Arabidopsis. Plant Physiol. 2001, 125, 763–769. [Google Scholar]
- Liu, F.; Xiang, N.; Hu, J.G.; Shijuan, Y.; Xie, L.; Brennan, C.S.; Huang, W.; Guo, X. The manipulation of gene expression and the biosynthesis of Vitamin C, E and folate in light-and dark-germination of sweet corn seeds. Sci. Rep. 2017, 7, 7484. [Google Scholar]
- Umezawa, T.; Sugiyama, N.; Anderson, J.; Takahashi, F.; Ishihama, Y.; Peck, S.; Shinozaki, K. Protein phosphorylation network in abscisic acid signaling. In Plant and Microbe Adaptations to Cold in a Changing World, 1st ed.; Imai, R., Yoshida, M., Matsumoto, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 1, pp. 155–164. [Google Scholar]
- Han, C.; Yang, P.; Sakata, K.; Komatsu, S. Quantitative proteomics reveals the role of protein phosphorylation in rice embryos during early stages of germination. J. Proteome Res. 2014, 13, 1766–1782. [Google Scholar]
- Yang, Q.; Sang, S.; Chen, Y.; Wei, Z.; Wang, P. The role of Arabidopsis inositol polyphosphate kinase AtIPK2β in glucose suppression of seed germination and seedling development. Plant Cell Physiol. 2018, 59, 343–354. [Google Scholar]
- Pope, P. A Molecular and Biochemical Analysis of Inositol Metabolism in Cotyledons of Phaseolus vulgaris and Arabidopsis thaliana. Ph.D. Thesis, University of Alabama, Tuscaloosa, AL, USA, 2000. [Google Scholar]
- Yang, X.-Y.; Chen, Z.-W.; Xu, T.; Qu, Z.; Pan, X.-D.; Qin, X.-H.; Ren, D.-T.; Liu, G.-Q. Arabidopsis kinesin KP1 specifically interacts with VDAC3, a mitochondrial protein, and regulates respiration during seed germination at low temperature. Plant Cell 2011, 23, 1093–1106. [Google Scholar]
- Botha, F.C.; Potgieter, G.P.; Botha, A.-M. Respiratory metabolism and gene expression during seed germination. Plant Growth Regul. 1992, 11, 211–224. [Google Scholar]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar]
- Coulon, D.; Faure, L.; Grison, M.; Pascal, S.; Wattelet-Boyer, V.; Clark, J.; Le Guedard, M.; Testet, E.; Bessoule, J.-J. LPIAT, a lyso-phosphatidylinositol acyltransferase, modulates seed germination in Arabidopsis thaliana through PIP signalling pathways and is involved in hyperosmotic response. Int. J. Mol. Sci. 2020, 21, 1654. [Google Scholar]
- Ravindran, P.; Yong, S.Y.; Mohanty, B.; Kumar, P.P. An LRR-only protein regulates abscisic acid-mediated abiotic stress responses during Arabidopsis seed germination. Plant Cell Rep. 2020, 39, 909–920. [Google Scholar]
- Chaudhry, F. Genetic and Biochemical Analyses of Arabidopsis thaliana Glycosyl Hydrolase Family 43 Enzymes A and B. Ph.D. Thesis, Purdue University, West Lafayette, IN, USA, 2007. [Google Scholar]
- Diaz, I.; Vicente-Carbajosa, J.; Abraham, Z.; Martinez, M.; Isabel-La Moneda, I.; Carbonero, P. The GAMYB protein from barley interacts with the DOF transcription factor BPBF and activates endosperm-specific genes during seed development. Plant J. 2002, 29, 453–464. [Google Scholar]
- Kobayashi, F.; Takumi, S.; Nakamura, C. Increased freezing tolerance in an ABA-hypersensitive mutant of common wheat. J. Plant Physiol. 2008, 165, 224–232. [Google Scholar]
- Wang, X.; Yu, K.; Li, H.; Peng, Q.; Chen, F.; Zhang, W.; Chen, S.; Hu, M.; Zhang, J. High-density SNP map construction and QTL identification for the apetalous character in Brassica napus L. Front. Plant Sci. 2015, 6, 1164. [Google Scholar]
- Yu, K.; Wang, X.; Chen, F.; Peng, Q.; Chen, S.; Li, H.; Zhang, W.; Fu, S.; Hu, M.; Long, W.; et al. Quantitative trait transcripts mapping coupled with expression quantitative trait loci mapping reveal the molecular network regulating the apetalous characteristic in Brassica napus L. Front. Plant Sci. 2018, 9, 89. [Google Scholar]
- Chen, F.; Zhang, W.; Yu, K.; Sun, L.; Gao, J.; Zhou, X.; Peng, Q.; Fu, S.; Hu, M.; Long, W.; et al. Unconditional and conditional QTL analyses of seed fatty acid composition in Brassica napus L. BMC Plant Biol. 2018, 18, 49. [Google Scholar]
- Yu, K.; Wang, X.; Li, W.; Sun, L.; Peng, Q.; Chen, F.; Zhang, W.; Guan, R.; Zhang, J. Identification and physical mapping of QTLs associated with flowering time in Brassica napus L. Euphytica 2019, 215, 152. [Google Scholar]
- Yu, K.; Zhang, W.; Guo, Y.; Zheng, M.; Chen, F.; Sun, C.; Hu, M.; Tian, E.; Wang, X.; Zhang, J. Integrating unconditional and conditional QTLs to dissect the genetic basis of stem mechanical strength in Brassica napus L. Euphytica 2021, 217, 34. [Google Scholar]
- Li, H.; Ye, G.; Wang, J. A modified algorithm for the improvement of composite interval mapping. Genetics 2007, 175, 361–374. [Google Scholar]
- Arcade, A.; Labourdette, A.; Falque, M.; Mangin, B.; Chardon, F.; Charcosset, A.; Joets, J. BioMercator: Integrating genetic maps and QTL towards discovery of candidate genes. Bioinformatics 2004, 20, 2324–2326. [Google Scholar]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar]



{kind=link}
{kind=link}
| Trait | Seed Lot | Parents | RIL Lines | Skewness | Kurtosis | Heritability | ||
|---|---|---|---|---|---|---|---|---|
| APL01 | Holly | Range | Mean ± SD | |||||
| Germination energy | 15SL | 72.0 | 92.0 | 0.0–100.0 | 77.1 ± 21.1 | −1.26 | 0.95 | 80.2% |
| 16SL | 82.0 | 96.0 | 0.0–100.0 | 81.3 ± 20.5 | −1.71 | 2.80 | ||
| 17SL | 90.0 | 98.0 | 4.0–100.0 | 81.4 ± 19.8 | −1.57 | 2.27 | ||
| Germination percentage | 15SL | 72.0 | 92.0 | 0.0–100.0 | 81.1 ± 19.9 | −1.52 | 1.86 | 80.0% |
| 16SL | 82.0 | 100.0 | 0.0–100.0 | 82.5 ± 20.4 | −1.76 | 2.93 | ||
| 17SL | 90.0 | 100.0 | 6.0–100.0 | 83.9 ± 19.4 | −1.75 | 2.83 | ||
| Trait | Consensus QTL | Identified QTL | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| QTL | Chr. | Pos. | CI | QTL | Chr. | Pos. | CI | LOD | PVE (%) | Add. | Seed Lot | |
| Germination energy | qGE.A4 | A4 | 3 | 1.5–4.5 | iqGE/17SL.A4 | A4 | 3 | 1.5–4.5 | 11.42 | 4.50 | 0.09 | 17SL |
| qGE.C2-1 | C2 | 81 | 77.5–81.5 | iqGE/17SL.C2-1 | C2 | 81 | 77.5–81.5 | 9.27 | 3.34 | 0.08 | 17SL | |
| qGE.C2-2 | C2 | 87 | 84.5–91.5 | iqGE/17SL.C2-2 | C2 | 87 | 84.5–91.5 | 5.32 | 2.20 | −0.06 | 17SL | |
| qGE.C3 | C3 | 29 | 28.5–29.5 | iqGE/15SL.C3 | C3 | 29 | 28.5–30.5 | 3.95 | 8.28 | 0.06 | 15SL | |
| iqGE/16SL.C3 | C3 | 29 | 28.5–30.5 | 4.04 | 10.59 | 0.06 | 16SL | |||||
| iqGE/17SL.C3 | C3 | 29 | 28.5–29.5 | 28.65 | 13.61 | 0.16 | 17SL | |||||
| qGE.C7 | C7 | 93 | 92.5–93.5 | iqGE/17SL.C7 | C7 | 93 | 92.5–93.5 | 5.61 | 2.04 | −0.06 | 17SL | |
| Germination percentage | qGP.A4 | A4 | 2 | 1.5–3.5 | iqGP/17SL.A4 | A4 | 2 | 1.5–3.5 | 3.63 | 2.39 | 0.04 | 17SL |
| qGP.A5 | A5 | 105 | 104.5–107.5 | iqGP/17SL.A5 | A5 | 105 | 104.5–107.5 | 4.38 | 2.48 | 0.04 | 17SL | |
| qGP.C2-1 | C2 | 81 | 79.5–81.5 | iqGP/17SL.C2-1 | C2 | 81 | 79.5–81.5 | 19.64 | 2.58 | 0.10 | 17SL | |
| qGP.C2-2 | C2 | 87 | 85.5–88.5 | iqGP/17SL.C2-2 | C2 | 87 | 85.5–88.5 | 16.06 | 2.31 | −0.09 | 17SL | |
| qGP.C3 | C3 | 29 | 28.5–29.5 | iqGP/15SL.C3 | C3 | 29 | 28.5–30.5 | 3.61 | 8.67 | 0.06 | 15SL | |
| iqGP/16SL.C3 | C3 | 29 | 28.5–30.5 | 3.42 | 10.35 | 0.06 | 16SL | |||||
| iqGP/17SL.C3 | C3 | 29 | 28.5–29.5 | 63.76 | 16.37 | 0.24 | 17SL | |||||
| qGP.C4 | C4 | 93 | 92.5–93.5 | iqGP/17SL.C4 | C4 | 93 | 92.5–93.5 | 43.64 | 9.28 | −0.18 | 17SL | |
| Trait | Chr._1 | Pos._1 | CI_1 | Chr._2 | Pos._2 | CI_2 | LOD | PVE (%) | Add._1 | Add._2 | Add-by-Add | Seed Lot |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Germination energy | C2 | 85 | 83.5–86.5 | C2 | 90 | 88.5–91.5 | 5.73 | 5.29 | 0.08 | −0.08 | 0.17 | 16SL |
| C3 | 100 | 98.5–101.5 | C3 | 105 | 103.5–106.5 | 7.16 | 3.39 | 0.07 | −0.06 | 0.18 | 16SL | |
| C5 | 25 | 23.5–26.5 | C5 | 30 | 28.5–31.5 | 6.43 | 4.59 | −0.11 | 0.11 | 0.12 | 16SL | |
| A9 | 65 | 63.5–66.5 | C4 | 40 | 38.5–41.5 | 5.08 | 11.01 | 0.02 | 0.02 | −0.05 | 17SL | |
| A2 | 40 | 38.5–41.5 | C6 | 35 | 33.5–36.5 | 5.07 | 10.04 | −0.02 | 0.01 | −0.07 | 15SL | |
| C7 | 95 | 93.5–96.5 | C8 | 45 | 43.5–46.5 | 6.74 | 13.01 | −0.01 | 0.00 | 0.08 | 15SL | |
| Germination percentage | A2 | 60 | 58.5–61.5 | A2 | 65 | 63.5–66.5 | 6.88 | 2.98 | −0.14 | 0.14 | 0.13 | 16SL |
| C2 | 80 | 78.5–81.5 | C2 | 85 | 83.5–86.5 | 6.19 | 3.30 | 0.14 | −0.14 | 0.11 | 16SL | |
| C3 | 100 | 98.5–101.5 | C3 | 105 | 103.5–106.5 | 8.43 | 2.41 | 0.06 | −0.05 | 0.19 | 16SL | |
| C4 | 140 | 138.5–141.5 | C4 | 145 | 143.5–146.5 | 5.34 | 2.47 | 0.14 | −0.17 | 0.14 | 16SL | |
| C5 | 25 | 23.5–26.5 | C5 | 30 | 28.5–31.5 | 8.28 | 3.35 | −0.12 | 0.12 | 0.13 | 16SL | |
| C6 | 75 | 73.5–76.5 | C6 | 80 | 78.5–81.5 | 6.62 | 3.29 | −0.13 | 0.14 | 0.11 | 16SL | |
| C9 | 20 | 18.5–21.5 | C9 | 25 | 23.5–26.5 | 6.32 | 2.83 | −0.14 | 0.12 | 0.16 | 16SL | |
| C5 | 25 | 23.5–26.5 | C5 | 30 | 28.5–31.5 | 6.00 | 10.10 | −0.11 | 0.11 | 0.12 | 15SL | |
| C7 | 95 | 93.5–96.5 | C8 | 45 | 43.5–46.5 | 7.05 | 3.60 | −0.01 | 0.00 | 0.08 | 15SL |
| Comparison Group | Number of Genes |
|---|---|
| A1 > A0 | 5415 |
| H1 > H0 | 4761 |
| H0 > A0 | 4685 |
| Gene_ID | Sequence Variation | Differential Expression | Arabidopsis Homologous Genes | Function Description |
|---|---|---|---|---|
| BnaC03g10300D | yes | no | AT5G20840 | Phosphoinositide phosphatase family protein; involved in phosphoinositides biosynthesis. |
| BnaC03g10310D | yes | no | AT5G20850 | RAS associated with diabetes protein 51 (RAD51); involved in DNA repair. |
| BnaC03g10320D | yes | no | AT5G20870 | O-Glycosyl hydrolases family 17 protein; involved in carbohydrate hydrolysis. |
| BnaC03g10330D | yes | no | AT5G20885 | RING/U-box superfamily protein; involved in protein ubiquitination. |
| BnaC03g10340D | yes | no | AT5G20890 | TCP-1/cpn60 chaperonin family protein; involved in protein folding. |
| BnaC03g10350D | yes | no | AT5G20910 | RING/U-box superfamily protein; involved in protein ubiquitination. |
| BnaC03g10370D | yes | no | AT5G20940 | Glycosyl hydrolase family protein; involved in carbohydrate hydrolysis. |
| BnaC03g10380D | yes | H1 > H0 | AT5G20970 | HSP20-like chaperones superfamily protein; involved in ROS generation. |
| BnaC03g10390D | yes | no | AT5G20980 | Methionine synthase 3 (MS3); involved in methionine biosynthesis. |
| BnaC03g10400D | yes | no | AT5G20990 | B73; involved in molybdenum cofactor biosynthesis. |
| BnaC03g10410D | yes | no | AT5G21010 | BTB-POZ and MATH domain 5 (BPM5); involved in protein ubiquitination. |
| BnaC03g10420D | yes | no | AT5G21050 | Hyccin; involved in phosphatidylinositol phosphate biosynthetic process. |
| BnaC03g10430D | yes | no | -- | Unknown protein |
| BnaC03g10440D | yes | no | AT5G21080 | Uncharacterized protein |
| BnaC03g10460D | yes | no | AT5G21090 | Leucine-rich repeat (LRR) family protein; involved in ABA signaling. |
| BnaC03g10470D | yes | no | AT5G21130 | Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family |
| BnaC03g10480D | yes | no | AT5G21160 | LA RNA-binding protein |
| BnaC03g10490D | yes | no | AT1G32210 | DEFENDER AGAINST APOPTOTIC DEATH 1 (ATDAD1) |
| BnaC03g10590D | yes | no | AT5G22791 | F-box family protein |
| BnaC03g10620D | yes | no | AT5G22040 | Ubiquitin carboxyl-terminal hydrolase; involved in protein deubiquitination. |
| BnaC03g10680D | yes | no | AT5G05760 | Syntaxin of plants 31 (SYP31); involved in cytokinesis. |
| BnaC03g10800D | no | (H0 > A0) & (A1 > A0) & (H1 > H0) | AT5G22310 | Trichohyalin-like protein; functions in protein binding. |
| BnaC03g10840D | yes | no | AT2G42380 | BZIP34; FUNCTIONS IN: involved in the regulation of ABA signaling response. |
| BnaC03g10920D | yes | no | AT5G22670 | F-box/RNI-like/FBD-like domains-containing protein |
| BnaC03g10950D | no | H0 > A0 | AT5G22740 | Cellulose synthase-like A02 (CSLA02); involved in mucilage biosynthesis. |
| BnaC03g10960D | yes | no | AT5G22750 | RAD5; involved in DNA repair. |
| BnaC03g11030D | no | (H0 > A0) & (A1 > A0) & (H1 > H0) | AT5G22880 | Histone B2 (HTB2); involved in nucleosome assembly. |
| BnaC03g11050D | no | H0 > A0 | AT5G22920 | CHY-type/CTCHY-type/RING-type Zinc finger protein; involved in regulation of stomatal opening. |
| BnaC03g11080D | yes | no | AT5G22950 | SNF7 family protein; involved in vesicle-mediated transport. |
| BnaC03g11220D | yes | no | AT3G45390 | Concanavalin A-like lectin protein kinase family protein; involved in protein phosphorylation. |
| BnaC03g11310D | yes | no | AT5G60210 | ROP interactive partner 5 (RIP5) |
| BnaC03g11340D | yes | no | AT5G60200 | TARGET OF MONOPTEROS 6 (TMO6); involved in seed germination. |
| BnaC03g11350D | yes | no | AT5G60190 | Cysteine proteinases superfamily protein; involved in proteolysis. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, K.; He, Y.; Li, Y.; Li, Z.; Zhang, J.; Wang, X.; Tian, E. Quantitative Trait Locus Mapping Combined with RNA Sequencing Reveals the Molecular Basis of Seed Germination in Oilseed Rape. Biomolecules 2021, 11, 1780. https://doi.org/10.3390/biom11121780
Yu K, He Y, Li Y, Li Z, Zhang J, Wang X, Tian E. Quantitative Trait Locus Mapping Combined with RNA Sequencing Reveals the Molecular Basis of Seed Germination in Oilseed Rape. Biomolecules. 2021; 11(12):1780. https://doi.org/10.3390/biom11121780
Chicago/Turabian StyleYu, Kunjiang, Yuqi He, Yuanhong Li, Zhenhua Li, Jiefu Zhang, Xiaodong Wang, and Entang Tian. 2021. "Quantitative Trait Locus Mapping Combined with RNA Sequencing Reveals the Molecular Basis of Seed Germination in Oilseed Rape" Biomolecules 11, no. 12: 1780. https://doi.org/10.3390/biom11121780