
The Type III Effectome of the Symbiotic Bradyrhizobium vignae Strain ORS3257

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Plasmid Construction and Mutagenesis
2.3. Transcriptome Analysis
2.4. Proteome Analysis
2.5. Western Blot Analysis
2.6. Symbiotic Tests
2.7. Outer Membrane Vesicles Observation by Transmission Electron Microscopy
3. Results
3.1. Transcriptome Identification of the Genes under the Control of TtsI and the Flavonoid Genistein in Bradyrhizobium vignae ORS3257
3.1.1. T3SS Machinery and Predicted T3SS Effectors Genes Regulated by TtsI
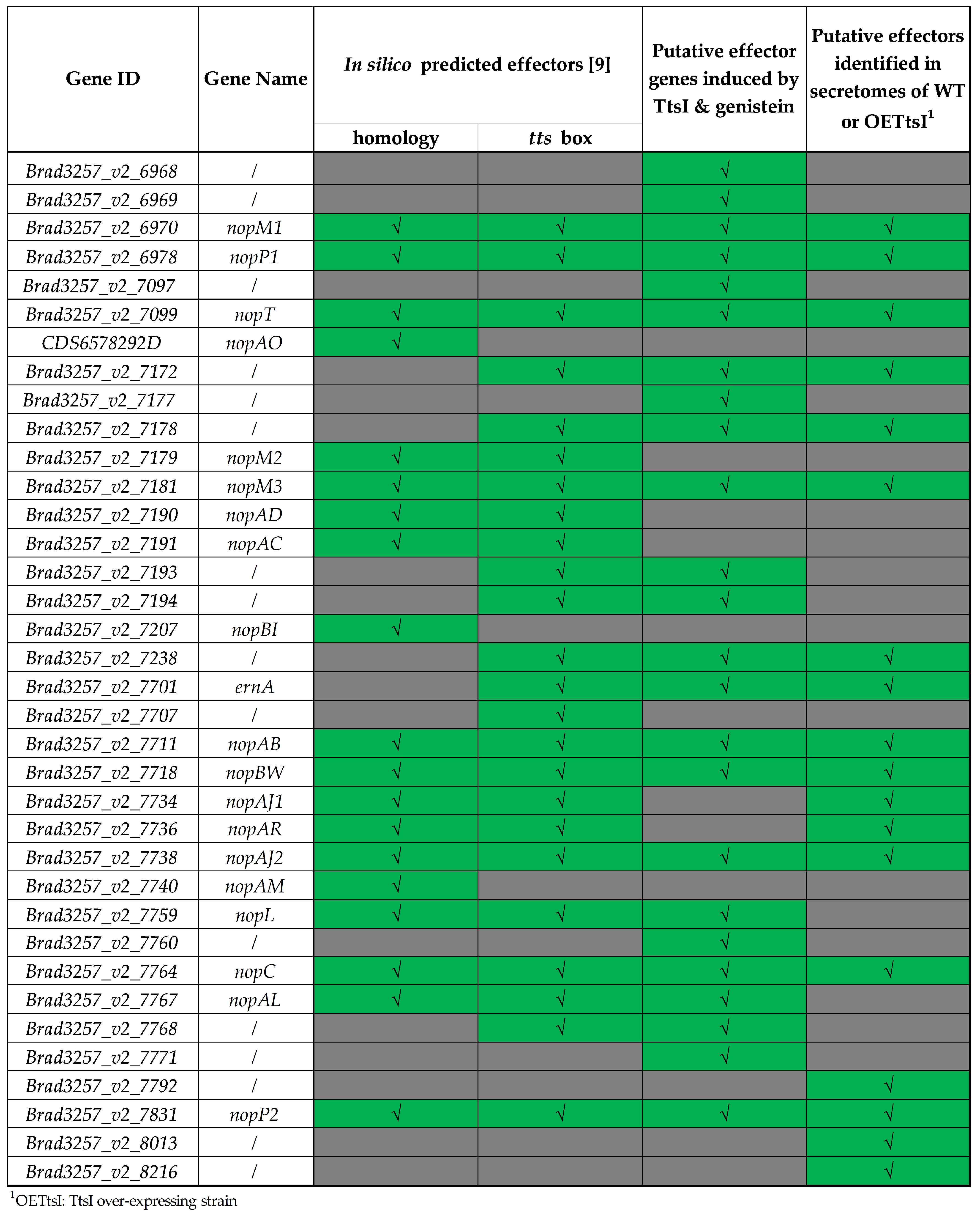
3.1.2. Identification of New Putative Effectors by Transcriptome Analysis
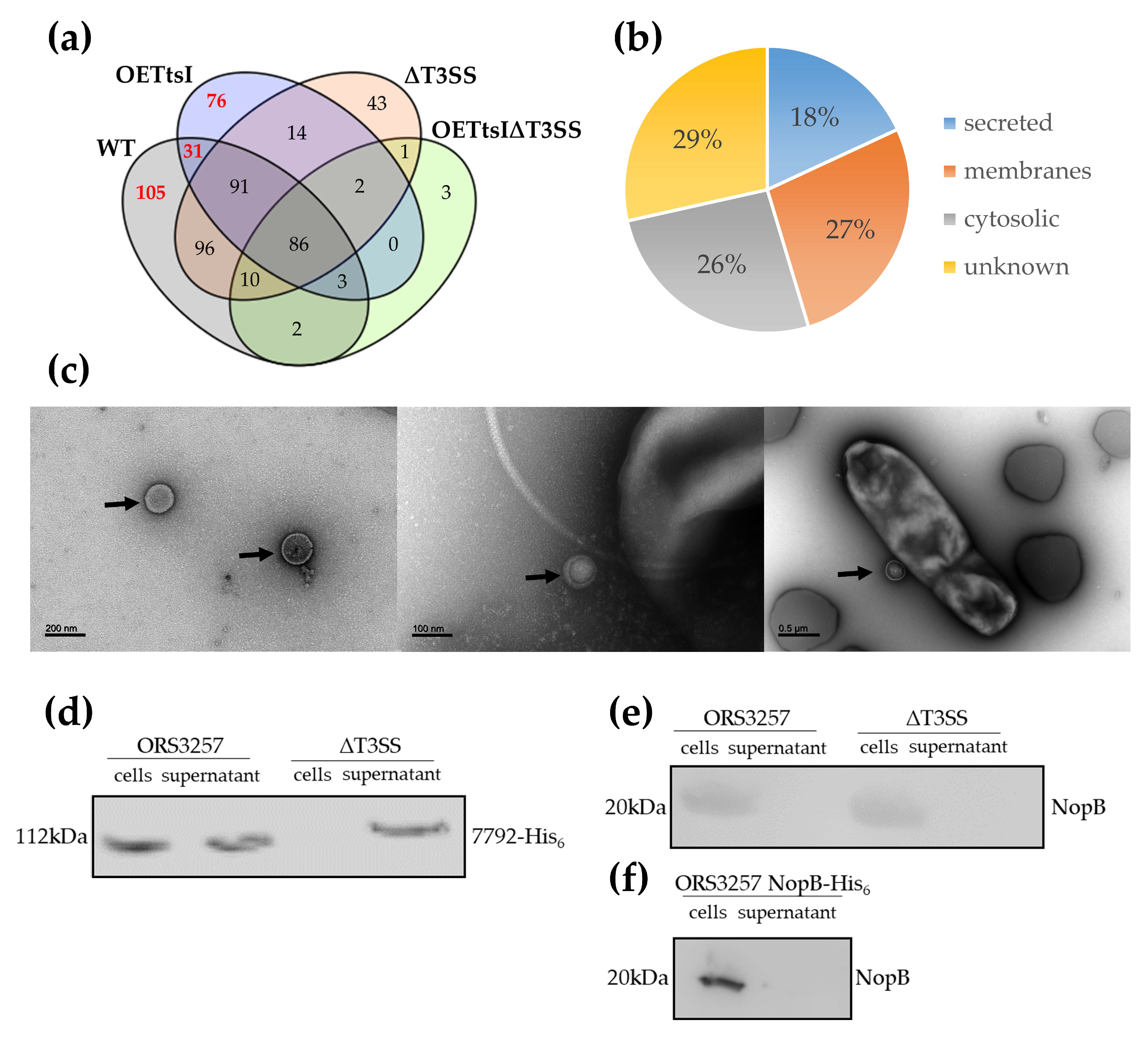
3.2. T3SS-Dependent Secretome in Bradyrhizobium vignae ORS3257
3.2.1. Cytosolic and Membrane Proteins Are Present in the Supernatant
3.2.2. The Expression Level of the T3SS Apparatus Modulates Protein Production and Secretion in Bradyrhizobium vignae ORS3257
3.2.3. T3SS-Dependent Secreted Proteins by Bradyrhizobium vignae ORS3257 and Identification of New Putative Effectors
3.2.4. Validation of T3SS-Effector Candidates Identified by Omics Using Western-Blotting
3.2.5. Secretion of T3SS-Needle Components
3.3. A Mutant in the BRAD3257_v2_7792 Effector Is Not Impaired in Symbiosis with Aeschynomene indica
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-gonzález, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 325, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ormeño-Orrillo, E.; Martínez-Romero, E. A genomotaxonomy view of the Bradyrhizobium genus. Front. Microbiol. 2019, 10, 1334. [Google Scholar] [CrossRef]
- Lerouge, P.; Roche, P.; Faucher, C.; Maillet, F.; Truchet, G.; Promé, J.C.; Dénarié, J. Symbiotic host-specificity of Rhizobium meliloti is determined by a sulphated and acylated glucosamine oligosaccharide signal. Nature 1990, 344, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Moulin, L.; Vallenet, D.; Barbe, V.; Cytryn, E.; Avarre, J.C.; Jaubert, M.; Simon, D.; Cartieaux, F.; Prin, Y.; et al. Legumes symbioses: Absence of nod genes in photosynthetic bradyrhizobia. Science 2007, 316, 1307–1312. [Google Scholar] [CrossRef]
- Okazaki, S.; Kaneko, T.; Sato, S.; Saeki, K. Hijacking of leguminous nodulation signaling by the rhizobial type III secretion system. Proc. Natl. Acad. Sci. USA 2013, 110, 17131–17136. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, S.; Tittabutr, P.; Teulet, A.; Thouin, J.; Fardoux, J.; Chaintreuil, C.; Gully, D.; Arrighi, J.F.; Furuta, N.; Miwa, H.; et al. Rhizobium-legume symbiosis in the absence of Nod factors: Two possible scenarios with or without the T3SS. ISME J. 2016, 10, 64–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratu, S.T.N.; Teulet, A.; Miwa, H.; Masuda, S.; Nguyen, H.P.; Yasuda, M.; Sato, S.; Kaneko, T.; Hayashi, M.; Giraud, E.; et al. Rhizobia use a pathogenic-like effector to hijack leguminous nodulation signalling. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Teulet, A.; Busset, N.; Fardoux, J.; Gully, D.; Chaintreuil, C.; Cartieaux, F.; Jauneau, A.; Comorge, V.; Okazaki, S.; Kaneko, T.; et al. The rhizobial type III effector ErnA confers the ability to form nodules in legumes. Proc. Natl. Acad. Sci. USA 2019, 116, 21758–21768. [Google Scholar] [CrossRef] [Green Version]
- Teulet, A.; Gully, D.; Rouy, Z.; Camuel, A.; Koebnik, R.; Giraud, E.; Lassalle, F. Phylogenetic distribution and evolutionary dynamics of nod and T3SS genes in the genus Bradyrhizobium. Microb. Genom. 2020, 6, mgen000407. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Mota, L.J.; Sorg, I.; Cornelis, G.R. Type III secretion: The bacteria-eukaryotic cell express. FEMS Microbiol. Lett. 2005, 252, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Deslandes, L.; Rivas, S. Catch me if you can: Bacterial effectors and plant targets. Trends Plant Sci. 2012, 17, 644–655. [Google Scholar] [CrossRef]
- Tampakaki, A.P. Commonalities and differences of T3SSs in rhizobia and plant pathogenic bacteria. Front. Plant Sci. 2014, 5, 114. [Google Scholar] [CrossRef]
- Peng, Y.; Van Wersch, R.; Zhang, Y. Convergent and divergent signaling in PAMP-triggered immunity and effector-triggered immunity. Mol. Plant Microbe Interact. 2018, 31, 403–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraud, E.; Lavergne, J.; Verméglio, A. Characterization of bacteriophytochromes from photosynthetic bacteria: Histidine kinase signaling triggered by light and redox sensing. Methods Enzymol. 2010, 471, 135–159. [Google Scholar] [CrossRef]
- Renier, A.; Maillet, F.; Fardoux, J.; Poinsot, V.; Giraud, E.; Nouwen, N. Photosynthetic Bradyrhizobium sp. strain ORS285 synthesizes 2-O-methylfucosylated lipochitooligosaccharides for nod gene–dependent interaction with Aeschynomene plants. Mol. Plant Microbe Interact. 2011, 24, 1440–1447. [Google Scholar] [CrossRef] [Green Version]
- Gully, D.; Czernic, P.; Cruveiller, S.; Mahé, F.; Longin, C.; Vallenet, D.; François, P.; Nidelet, S.; Rialle, S.; Giraud, E.; et al. Transcriptome profiles of Nod factor-independent symbiosis in the tropical legume Aeschynomene evenia. Sci. Rep. 2018, 8, 10934. [Google Scholar] [CrossRef]
- Le Quéré, A.; Gully, D.; Teulet, A.; Navarro, E.; Gargani, D.; Fardoux, J.; Cruveiller, S.; Neyra, M.; Giraud, E.; Krasova Wade, T. Complete genome sequence of Bradyrhizobium sp. strain ORS3257, an efficient nitrogen-fixing bacterium isolated from cowpea in Senegal. Microbiol. Resour. Announc. 2019, 8, e01449-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, S.; Zehner, S.; Hempel, J.; Lang, K.; Michael, G. Genetic organization and functional analysis of the type III secretion system of Bradyrhizobium elkanii. FEMS Microbiol. Lett. 2009, 295, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaldi, K.; Gourion, B.; Fardoux, J.; Hannibal, L.; Cartieaux, F.; Boursot, M.; Vallenet, D.; Chaintreuil, C.; Prin, Y.; Nouwen, N.; et al. Large-scale transposon mutagenesis of photosynthetic Bradyrhizobium sp. strain ORS278 reveals new genetic loci putatively important for nod-independent symbiosis with Aeschynomene indica. Mol. Plant. Microbe Interact. 2010, 23, 760–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.H.; Urbach, J.M.; Law, T.F.; Arnold, L.W.; Hu, A.; Gombar, S.; Grant, S.R.; Ausubel, F.M.; Dangl, J.L. A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae. Proc. Natl. Acad. Sci. USA 2005, 102, 2549–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Campos, S.B.; Deakin, W.J.; Broughton, W.J.; Passaglia, L.M.P. Roles of flavonoids and the transcriptional regulator TtsI in the activation of the type III secretion system of Bradyrhizobium elkanii SEMIA587. Microbiology 2011, 157, 627–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, J.; Stacey, G. Nodulation gene regulation in Bradyrhizobium japonicum: A unique integration of global regulatory circuits. Appl. Environ. Microbiol. 2003, 69, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Poueymiro, M.; Cazalé, A.C.; François, J.M.; Parrou, J.L.; Peeters, N.; Genin, S. A Ralstonia solanacearum type III effector directs the production of the plant signal metabolite trehalose-6-phosphate. mBio 2014, 5, e02065-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, A.T. Outer membrane vesicles ( OMVs ) of gram-negative bacteria: A perspective update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Breakefield, X.; Budnik, V.; Buzas, E.; Bond, V.C.; et al. Vesiclepedia: A compendium for extracellular vesicles with Ccontinuous community annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gal, J.E.; Lara-tejero, M.; Marlovits, T.C.; Wagner, S. Bacterial type III secretion systems: Specialized nanomachines for protein delivery into target cells. Annu. Rev. Microbiol. 2014, 68, 415–438. [Google Scholar] [CrossRef] [Green Version]
- Marie, C.; Deakin, W.J.; Viprey, V.; Kopciñska, J.; Golinowski, W.; Krishnan, B.; Perret, X.; Broughton, W.J. Characterization of Nops, nodulation outer proteins, secreted via the type III secretion system of NGR234. Mol. Plant Microbe Interact. 2003, 16, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Lonjon, F.; Turner, M.; Rengel, D.; Lohou, D.; van de Kerkhove, Q.; Cazalé, A.-C.; Peeters, N.; Genin, S.; Vailleau, F. Comparative secretome analysis of Ralstonia solanacearum type 3 secretion-associated mutants reveals a fine control of effector delivery, essential for bacterial pathogenicity. Mol. Cell. Proteom. 2016, 15, 598–613. [Google Scholar] [CrossRef] [Green Version]
- Hempel, J.; Zehner, S.; Göttfert, M.; Patschkowski, T. Analysis of the secretome of the soybean symbiont Bradyrhizobium japonicum. J. Biotechnol. 2009, 140, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Deakin, W.J.; Marie, C.; Saad, M.M.; Krishnan, H.B.; Broughton, W.J. NopA is associated with cell surface appendages produced by the type III secretion system of Rhizobium sp. strain NGR234. Mol. Plant Microbe Interact. 2005, 18, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, M.M.; Staehelin, C.; Broughton, W.J.; Deakin, W.J. Protein-protein interactions within type III secretion system-dependent pili of Rhizobium sp. strain NGR234. J. Bacteriol. 2008, 190, 750–754. [Google Scholar] [CrossRef] [Green Version]
- Saad, M.M.; Kobayashi, H.; Marie, C.; Brown, I.R.; Mansfield, J.W.; Broughton, W.J.; Deakin, W.J. NopB, a type III secreted protein of Rhizobium sp. strain NGR234, is associated with pilus-like surface appendages. J. Bacteriol. 2005, 187, 1173–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotson, A.; Chosed, R.; Shu, H.; Orth, K.; Mudgett, M.B. Xanthomonas type III effector XopD targets SUMO-conjugated proteins in planta. Mol. Microbiol. 2003, 50, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Staehelin, C.; Krishnan, H.B. Nodulation outer proteins: Double-edged swords of symbiotic rhizobia. Biochem. J. 2015, 470, 263–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvitko, B.H.; Park, D.H.; Velásquez, A.C.; Wei, C.F.; Russell, A.B.; Martin, G.B.; Schneider, D.J.; Collmer, A. Deletions in the repertoire of pseudomonas syringae pv. tomato DC3000 type III secretion effector genes reveal functional overlap among effectors. PLoS Pathog. 2009, 5, e1000388. [Google Scholar] [CrossRef]
- van Engelenburg, S.B.; Palmer, A.E. Imaging type-III secretion reveals dynamics and spatial segregation of Salmonella effectors. Nat. Methods. 2010, 7, 325–330. [Google Scholar] [CrossRef]
- Rehman, H.M.; Cheung, W.L.; Wong, K.S.; Xie, M.; Luk, C.Y.; Wong, F.L.; Li, M.W.; Tsai, S.N.; To, W.T.; Chan, L.Y.; et al. High-throughput mass spectrometric analysis of the whole proteome and secretome from Sinorhizobium fredii strains CCBAU25509 and CCBAU45436. Front. Microbiol. 2019, 10, 2569. [Google Scholar] [CrossRef]
- Songwattana, P.; Noisangiam, R.; Teamtisong, K.; Prakamhang, J.; Teulet, A.; Tittabutr, P.; Piromyou, P.; Boonkerd, N.; Giraud, E.; Teaumroong, N. Type 3 secretion system (T3SS) of Bradyrhizobium sp. DOA9 and its roles in legume symbiosis and rice endophytic association. Front. Microbiol. 2017, 8, 1810. [Google Scholar] [CrossRef] [PubMed]
- Kimbrel, J.A.; Thomas, W.J.; Jiang, Y.; Creason, A.L.; Thireault, C.A.; Sachs, J.L.; Chang, J.H. Mutualistic co-evolution of type III effector genes in Sinorhizobium fredii and Bradyrhizobium japonicum. PLoS Pathog. 2013, 9, e1003204. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.P.; Ratu, S.T.N.; Yasuda, M.; Göttfert, M.; Okazaki, S. InnB, a novel type III effector of Bradyrhizobium elkanii USDA61, controls symbiosis with vigna species. Front. Microbiol. 2018, 9, 3155. [Google Scholar] [CrossRef] [Green Version]
- Süß, C.; Hempel, J.; Zehner, S.; Krause, A.; Patschkowski, T.; Göttfert, M. Identification of genistein-inducible and type III-secreted proteins of Bradyrhizobium japonicum. J. Biotechnol. 2006, 126, 69–77. [Google Scholar] [CrossRef]
- Altindis, E.; Fu, Y.; Mekalanos, J.J. Proteomic analysis of Vibrio cholerae outer membrane vesicles. Proc. Natl. Acad. Sci. USA 2014, 111, E1548–E1556. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, V.K.; Vorhölter, F.J.; Niehaus, K.; Watt, S.A. Analysis of outer membrane vesicle associated proteins isolated from the plant pathogenic bacterium Xanthomonas campestris pv. campestris. BMC Microbiol. 2008, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Marie, C.; Deakin, W.J.; Ojanen-Reuhs, T.; Diallo, E.; Reuhs, B.; Broughton, W.J.; Perret, X. TtsI, a key regulator of Rhizobium species NGR234 is required for type III-dependent protein secrection and synthesis of rhamnose-rich polysaccharides. Mol. Plant Microbe Interact. 2004, 17, 958–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Sukovich, D.J.; Ciccarelli, L.; Mayr, J.; Fernandez-Rodriguez, J.; Mirsky, E.A.; Tucker, A.C.; Gordon, D.B.; Marlovits, T.C.; Voigt, C.A. Control of type III protein secretion using a minimal genetic system. Nat. Commun. 2017, 8, 14737. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.; Eardley, L.; Hotson, A.; Cao, Y.; Mudgett, M.B. Characterization of the Xanthomonas AvrXv4 effector, a SUMO protease translocated into plant cells. Mol. Plant Microbe Interact. 2004, 17, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Tsurumaru, H.; Hashimoto, S.; Okizaki, K.; Kanesaki, Y.; Yoshikawa, H.; Yamakawa, T. A putative type III secretion system effector encoded by the MA20_12780 gene in Bradyrhizobium japonicum Is-34 causes incompatibility with Rj4 genotype soybeans. Appl. Environ. Microbiol. 2015, 81, 5812–5819. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Q.W.; Bai, J.; Cai, J.; Huang, Q.Y.; Wang, Y.; Liang, Y.; Zhong, Z.; Wagner, C.; Xie, Z.P.; Staehelin, C. NopD of Bradyrhizobium sp. XS1150 possesses SUMO protease activity. Front. Microbiol. 2020, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Faruque, O.M.; Hiwa, H.; Uasuda, M.; Fuji, Y.; Kneko, T.; Sato, S.; Okazaki, S. Identification of Bradyrhizobium elkanii genes involved in incompatibility with soybean plants carrying the Rj4 allele. Appl. Environ. Microbiol. 2015, 81, 6710–6717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratu, S.T.N.; Hirata, A.; Kalaw, C.O.; Yasuda, M.; Tabuchi, M.; Okazaki, S. Multiple domains in the rhizobial type III effector Bel2-5 determine symbiotic efficiency with soybean. Front. Plant Sci. 2021, 12, 689064. [Google Scholar] [CrossRef] [PubMed]
- Krasova-Wade, T.; Ndoye, I.; Braconnier, S.; Sarr, B.; De Lajudie, P.; Neyra, M. Diversity of indigeneous bradyrhizobia associated with three cowpea cultivars (Vigna unguiculata (L.) Walp.) grown under limited and favorable water conditions in Senegal (West Africa). Afr. J. Biotechnol. 2003, 2, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Grönemeyer, J.L.; Hurek, T.; Bünger, W.; Reinhold-Hurek, B. Bradyrhizobium vignae sp. nov., a nitrogen-fixing symbiont isolated from effective nodules of Vigna and Arachis. Int. J. Syst. Evol. Microbiol. 2016, 66, 62–69. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busset, N.; Gully, D.; Teulet, A.; Fardoux, J.; Camuel, A.; Cornu, D.; Severac, D.; Giraud, E.; Mergaert, P. The Type III Effectome of the Symbiotic Bradyrhizobium vignae Strain ORS3257. Biomolecules 2021, 11, 1592. https://doi.org/10.3390/biom11111592
Busset N, Gully D, Teulet A, Fardoux J, Camuel A, Cornu D, Severac D, Giraud E, Mergaert P. The Type III Effectome of the Symbiotic Bradyrhizobium vignae Strain ORS3257. Biomolecules. 2021; 11(11):1592. https://doi.org/10.3390/biom11111592
Chicago/Turabian StyleBusset, Nicolas, Djamel Gully, Albin Teulet, Joël Fardoux, Alicia Camuel, David Cornu, Dany Severac, Eric Giraud, and Peter Mergaert. 2021. "The Type III Effectome of the Symbiotic Bradyrhizobium vignae Strain ORS3257" Biomolecules 11, no. 11: 1592. https://doi.org/10.3390/biom11111592