
VEGF as a Direct Functional Regulator of Photoreceptors and Contributing Factor to Diabetes-Induced Alteration of Photoreceptor Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
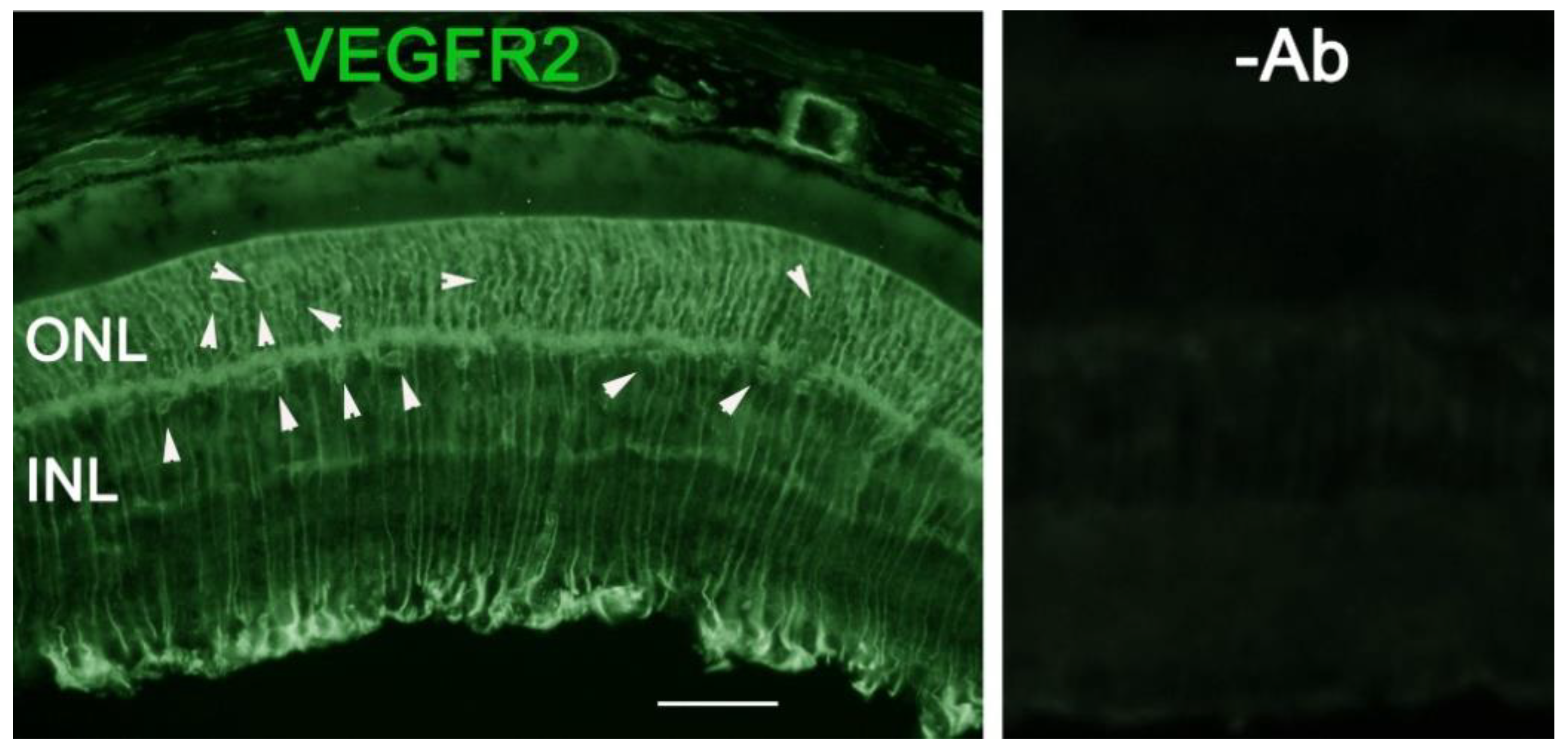
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Electroretinography (ERG)
2.3. Immunohistochemistry
2.4. Statistical Analaysis
3. Results
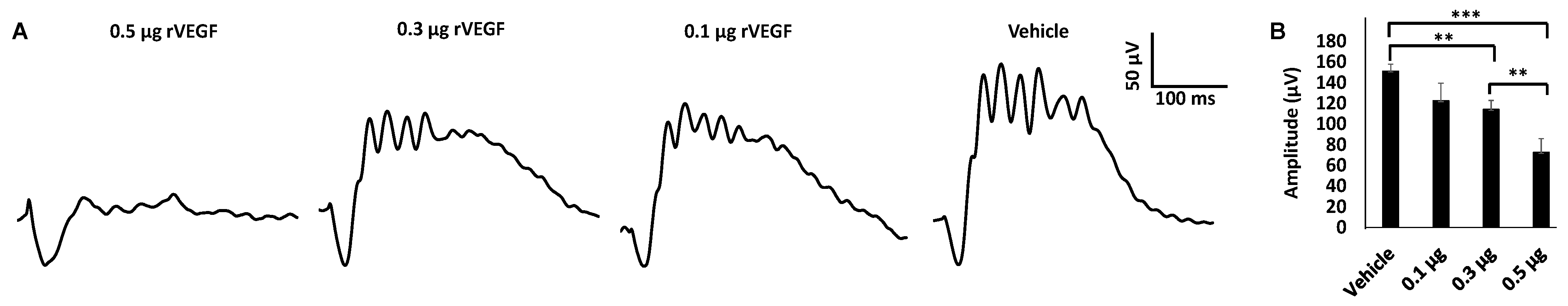
3.1. Effect of VEGF on Phtoreceptor Function
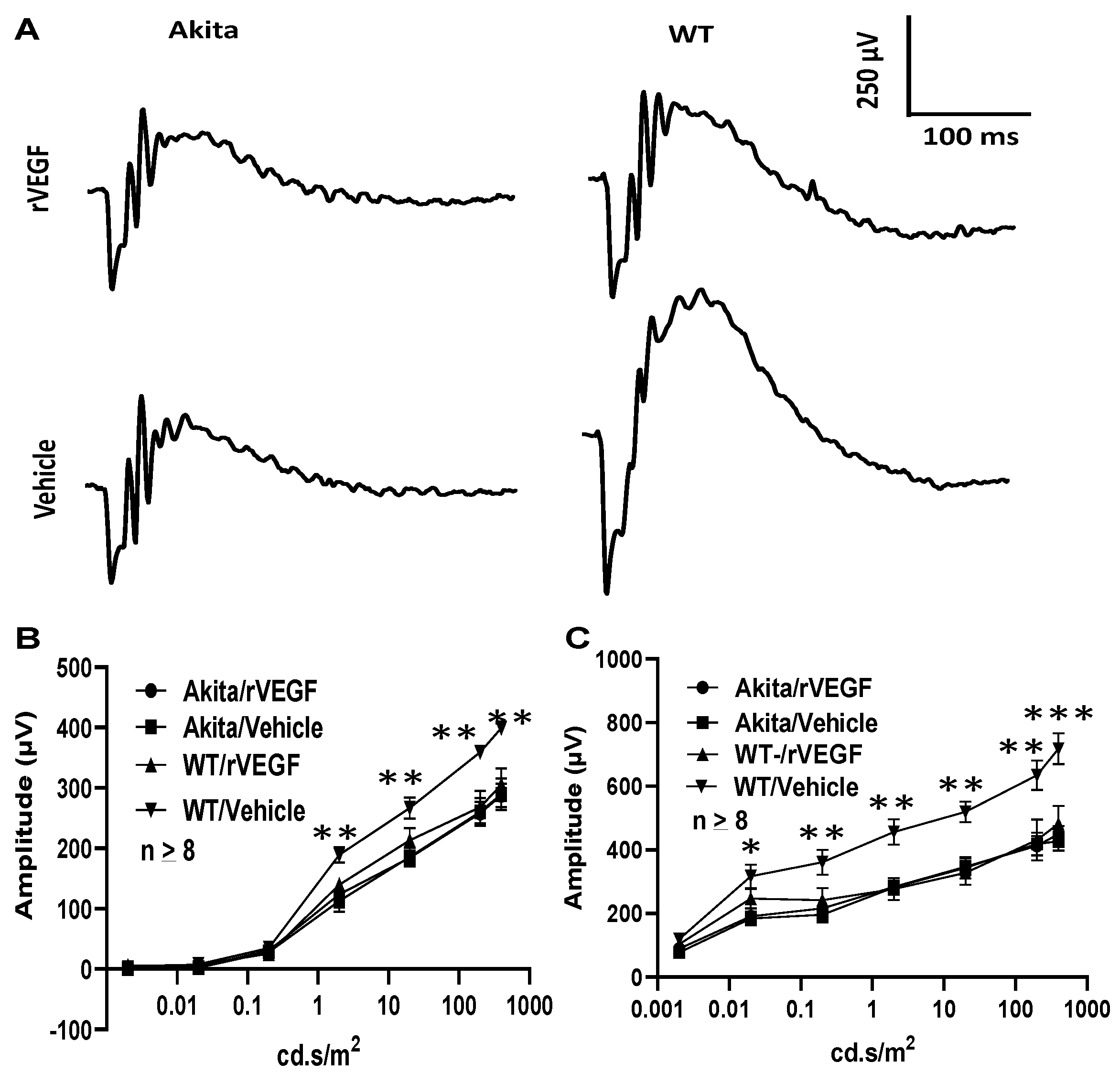
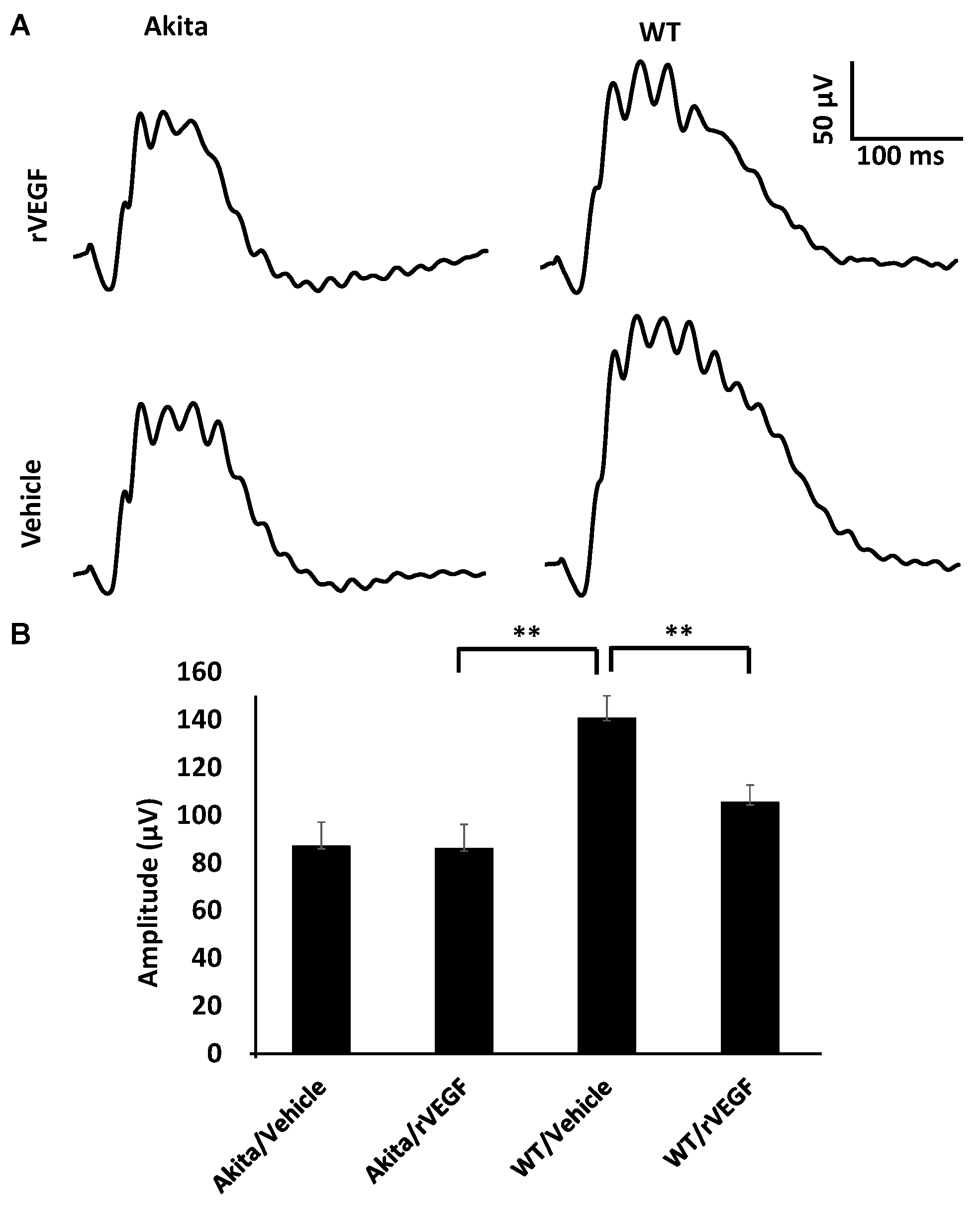
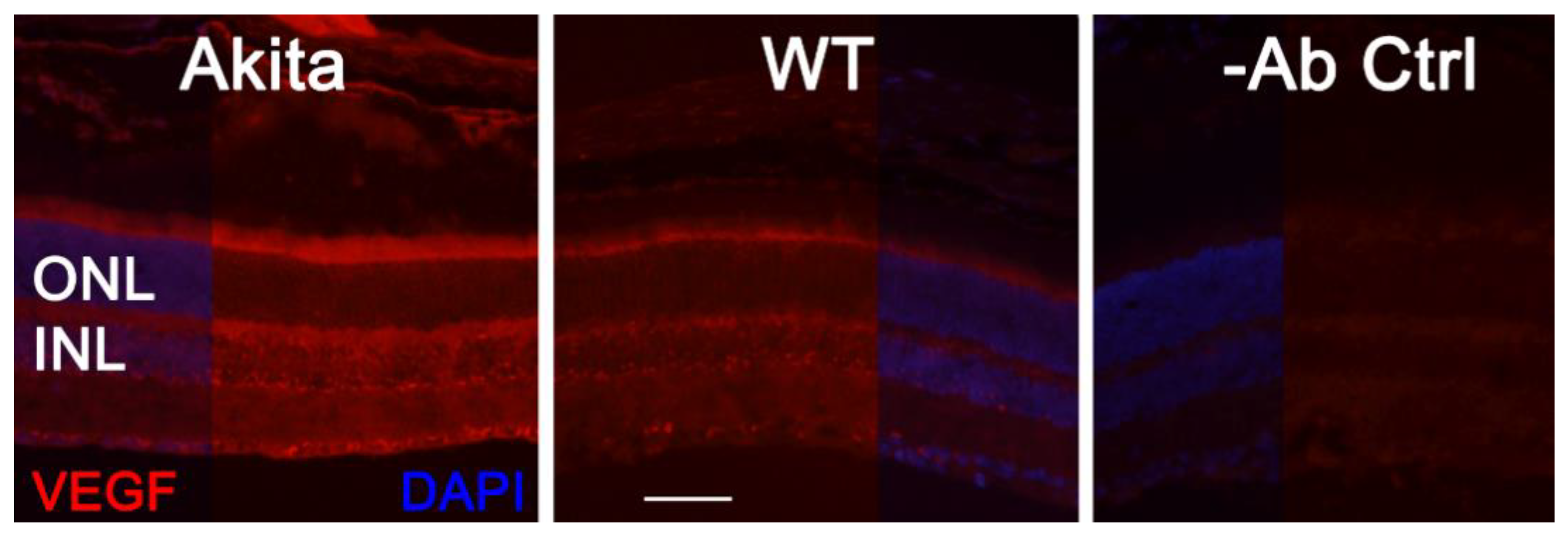
3.2. Effect of VEGF on Phtoreceptor Function in Mouse Model of DR
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, P.M.; Roon, P.; Van Ells, T.K.; Ganapathy, V.; Smith, S.B. Death of retinal neurons in streptozotocin-induced diabetic mice. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3330–3336. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.W.; Antonetti, D.A.; Barber, A.J.; LaNoue, K.F.; Nakamura, M. New insights into the pathophysiology of diabetic retinopathy: Potential cell-specific therapeutic targets. Diabetes Technol. Ther. 2000, 2, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, H.; Hirose, H.; Miyake, Y. S-cone pathway sensitivity in diabetes measured with threshold versus intensity curves on flashed backgrounds. Investig. Ophthalmol. Vis. Sci. 1996, 37, 680–684. [Google Scholar]
- Aspinall, P.A. Rod-cone interaction: Some indirect evidence. Acta Ophthalmol. 1977, 55, 294–302. [Google Scholar] [CrossRef]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juen, S.; Kieselbach, G.F. Electrophysiological changes in juvenile diabetics without retinopathy. Arch. Ophthalmol. 1990, 108, 372–375. [Google Scholar] [CrossRef]
- Fu, S.; Dong, S.; Zhu, M.; Sherry, D.M.; Wang, C.; You, Z.; Haigh, J.J.; Le, Y.Z. Müller glia are a major cellular source of survival signals for retinal neurons in diabetes. Diabetes 2015, 64, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Ma, J.X.; Guo, J.; Wang, J.; Zhu, M.; Chen, Y.; Le, Y.Z. Muller cell-derived VEGF is a significant contributor to retinal neovascularization. J. Pathol. 2009, 219, 446–454. [Google Scholar] [CrossRef]
- Wang, J.; Xu, X.; Elliott, M.H.; Zhu, M.; Le, Y.Z. Muller cell-derived VEGF is essential for diabetes-induced retinal inflammation and vascular leakage. Diabetes 2010, 59, 2297–2305. [Google Scholar] [CrossRef] [Green Version]
- Aiello, L.P.; Avery, R.L.; Arrigg, P.G.; Keyt, B.A.; Jampel, H.D.; Shah, S.T.; Pasquale, L.R.; Thieme, H.; Iwamoto, M.A.; Park, J.E.; et al. Vascular endothelial growth factor in ocular fluid of patients with diabetic retinopathy and other retinal disorders. N. Engl. J. Med. 1994, 331, 1480–1487. [Google Scholar] [CrossRef]
- Alon, T.; Hemo, I.; Itin, A.; Pe’er, J.; Stone, J.; Keshet, E. Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity. Nat. Med. 1995, 1, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhu, M.; Le, Y.Z. Functions of Muller cell-derived vascular endothelial growth factor in diabetic retinopathy. World J. Diabetes 2015, 6, 726–733. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, D.P.; Hintz, T.M.; Scharfman, H.E. Modulation of vascular endothelial growth factor (VEGF) expression in motor neurons and its electrophysiological effects. Brain Res. Bull. 2008, 76, 36–44. [Google Scholar] [CrossRef] [Green Version]
- McCloskey, D.P.; Croll, S.D.; Scharfman, H.E. Depression of synaptic transmission by vascular endothelial growth factor in adult rat hippocampus and evidence for increased efficacy after chronic seizures. J. Neurosci. 2005, 25, 8889–8897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.W.; Choi, M.; Kim, Y.S.; Park, H.; Lee, H.R.; Yun, C.O.; Kim, E.J.; Choi, J.S.; Kim, S.; Rhim, H.; et al. Vascular endothelial growth factor (VEGF) signaling regulates hippocampal neurons by elevation of intracellular calcium and activation of calcium/calmodulin protein kinase II and mammalian target of rapamycin. Cell Signal. 2008, 20, 714–725. [Google Scholar] [CrossRef]
- Lin, J.; Li, G.; Den, X.; Xu, C.; Liu, S.; Gao, Y.; Liu, H.; Zhang, J.; Li, X.; Liang, S. VEGF and its receptor-2 involved in neuropathic pain transmission mediated by P2X(2)(/)(3) receptor of primary sensory neurons. Brain Res. Bull. 2010, 83, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Bouzioukh, F.; Daoudal, G.; Falk, J.; Debanne, D.; Rougon, G.; Castellani, V. Semaphorin3A regulates synaptic function of differentiated hippocampal neurons. Eur. J. Neurosci. 2006, 23, 2247–2254. [Google Scholar] [CrossRef]
- Malechka, V.V.; Moiseyev, G.; Takahashi, Y.; Shin, Y.; Ma, J.X. Impaired Rhodopsin Generation in the Rat Model of Diabetic Retinopathy. Am. J. Pathol. 2017, 187, 2222–2231. [Google Scholar] [CrossRef] [Green Version]
- Shih, S.C.; Ju, M.; Liu, N.; Smith, L.E. Selective stimulation of VEGFR-1 prevents oxygen-induced retinal vascular degeneration in retinopathy of prematurity. J. Clin. Investig. 2003, 112, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Saint-Geniez, M.; Maharaj, A.S.; Walshe, T.E.; Tucker, B.A.; Sekiyama, E.; Kurihara, T.; Darland, D.C.; Young, M.J.; D’Amore, P.A. Endogenous VEGF is required for visual function: Evidence for a survival role on muller cells and photoreceptors. PLoS ONE 2008, 3, e3554. [Google Scholar] [CrossRef] [Green Version]
- Nishijima, K.; Ng, Y.S.; Zhong, L.; Bradley, J.; Schubert, W.; Jo, N.; Akita, J.; Samuelsson, S.J.; Robinson, G.S.; Adamis, A.P.; et al. Vascular endothelial growth factor-A is a survival factor for retinal neurons and a critical neuroprotectant during the adaptive response to ischemic injury. Am. J. Pathol. 2007, 171, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.Z. VEGF production and signaling in Müller glia are critical to modulating vascular function and neuronal integrity in diabetic retinopathy and hypoxic retinal vascular diseases. Vis. Res. 2017, 139, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.Z.; Xu, B.; Chucair-Elliott, A.J.; Zhang, H.; Zhu, M. VEGF mediates retinal Müller cell viability and neuroprotection through BDNF in diabetes. Biomolecules 2021, 11, 712. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.J.; Antonetti, D.A.; Kern, T.S.; Reiter, C.E.; Soans, R.S.; Krady, J.K.; Levison, S.W.; Gardner, T.W.; Bronson, S.K. The Ins2Akita mouse as a model of early retinal complications in diabetes. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2210–2218. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Liu, Y.; Zhu, M.; Xu, X.; Le, Y.Z. Simplified system to investigate alteration of retinal neurons in diabetes. Adv. Exp. Med. Biol. 2014, 801, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Anderson, R.E.; Agbaga, M.P.; Rucker, E.B., 3rd; Le, Y.Z. Loss of BCL-XL in rod photoreceptors: Increased susceptibility to bright light stress. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5583–5589. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.; Ash, J.D.; Al-Ubaidi, M.R.; Chen, Y.; Ma, J.; Anderson, R.E. Targeted expression of Cre recombinase to cone photoreceptors in transgenic mice. Mol. Vis. 2004, 10, 1011–1018. [Google Scholar] [PubMed]
- Le, Y.; Zheng, L.; Zheng, W.; Agbaga, M.; Zhu, M.; Ash, J.D.; Anderson, R.E. Mouse opsin promoter controlled expression of Cre recombinase in transgenic mice. Mol. Vis. 2006, 12, 389–398. [Google Scholar]
- Xu, H.Z.; Le, Y.Z. Significance of outer blood-retina barrier breakdown in diabetes and ischemia. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2160–2164. [Google Scholar] [CrossRef]
- Ohno-Matsui, K.; Hirose, A.; Yamamoto, S.; Saikia, J.; Okamoto, N.; Gehlbach, P.; Duh, E.J.; Hackett, S.; Chang, M.; Bok, D.; et al. Inducible expression of vascular endothelial growth factor in adult mice causes severe proliferative retinopathy and retinal detachment. Am. J. Pathol. 2002, 160, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Wen, R.; Song, Y.; Kjellstrom, S.; Tanikawa, A.; Liu, Y.; Li, Y.; Zhao, L.; Bush, R.A.; Laties, A.M.; Sieving, P.A. Regulation of rod phototransduction machinery by ciliary neurotrophic factor. J. Neurosci. 2006, 26, 13523–13530. [Google Scholar] [CrossRef] [Green Version]
- McGill, T.J.; Prusky, G.T.; Douglas, R.M.; Yasumura, D.; Matthes, M.T.; Nune, G.; Donohue-Rolfe, K.; Yang, H.; Niculescu, D.; Hauswirth, W.W.; et al. Intraocular CNTF reduces vision in normal rats in a dose-dependent manner. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5756–5766. [Google Scholar] [CrossRef]
- Wen, R.; Tao, W.; Li, Y.; Sieving, P.A. CNTF and retina. Prog. Retin. Eye Res. 2012, 31, 136–151. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shao, Y.; Sasore, T.; Moiseyev, G.; Zhou, K.; Ma, X.; Du, Y.; Ma, J.X. Interphotoreceptor Retinol-Binding Protein Ameliorates Diabetes-Induced Retinal Dysfunction and Neurodegeneration Through Rhodopsin. Diabetes 2021, 70, 788–799. [Google Scholar] [CrossRef]
- Kung, F.; Wang, W.; Tran, T.S.; Townes-Anderson, E. Sema3A Reduces Sprouting of Adult Rod Photoreceptors In Vitro. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4318–4331. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Zhu, M.; Li, D.; Wu, Q.; Le, Y.-Z. VEGF as a Direct Functional Regulator of Photoreceptors and Contributing Factor to Diabetes-Induced Alteration of Photoreceptor Function. Biomolecules 2021, 11, 988. https://doi.org/10.3390/biom11070988
Hu J, Zhu M, Li D, Wu Q, Le Y-Z. VEGF as a Direct Functional Regulator of Photoreceptors and Contributing Factor to Diabetes-Induced Alteration of Photoreceptor Function. Biomolecules. 2021; 11(7):988. https://doi.org/10.3390/biom11070988
Chicago/Turabian StyleHu, Jianyan, Meili Zhu, Dai Li, Qiang Wu, and Yun-Zheng Le. 2021. "VEGF as a Direct Functional Regulator of Photoreceptors and Contributing Factor to Diabetes-Induced Alteration of Photoreceptor Function" Biomolecules 11, no. 7: 988. https://doi.org/10.3390/biom11070988