The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments with the Biostimulant
2.2. Evaluation of Productivity and Pomological Analysis
2.3. Proximate Composition
2.3.1. Sugar Content
2.3.2. Protein Content
2.3.3. Lipid Content and Fatty Acid Profile
2.3.4. Moisture, Ashes, and Mineral Content
2.4. Ascorbic Acid Content
2.5. Total Tocopherol Content
2.6. Total Carotenoid Content
2.7. Lycopene Content
2.8. Total Polyphenol Content
2.9. Evaluation of Antioxidant Properties
2.9.1. ABTS assay
2.9.2. DPPH assay
2.9.3. FRAP assay
2.10. Statistycal Analysis
3. Results and Discussion
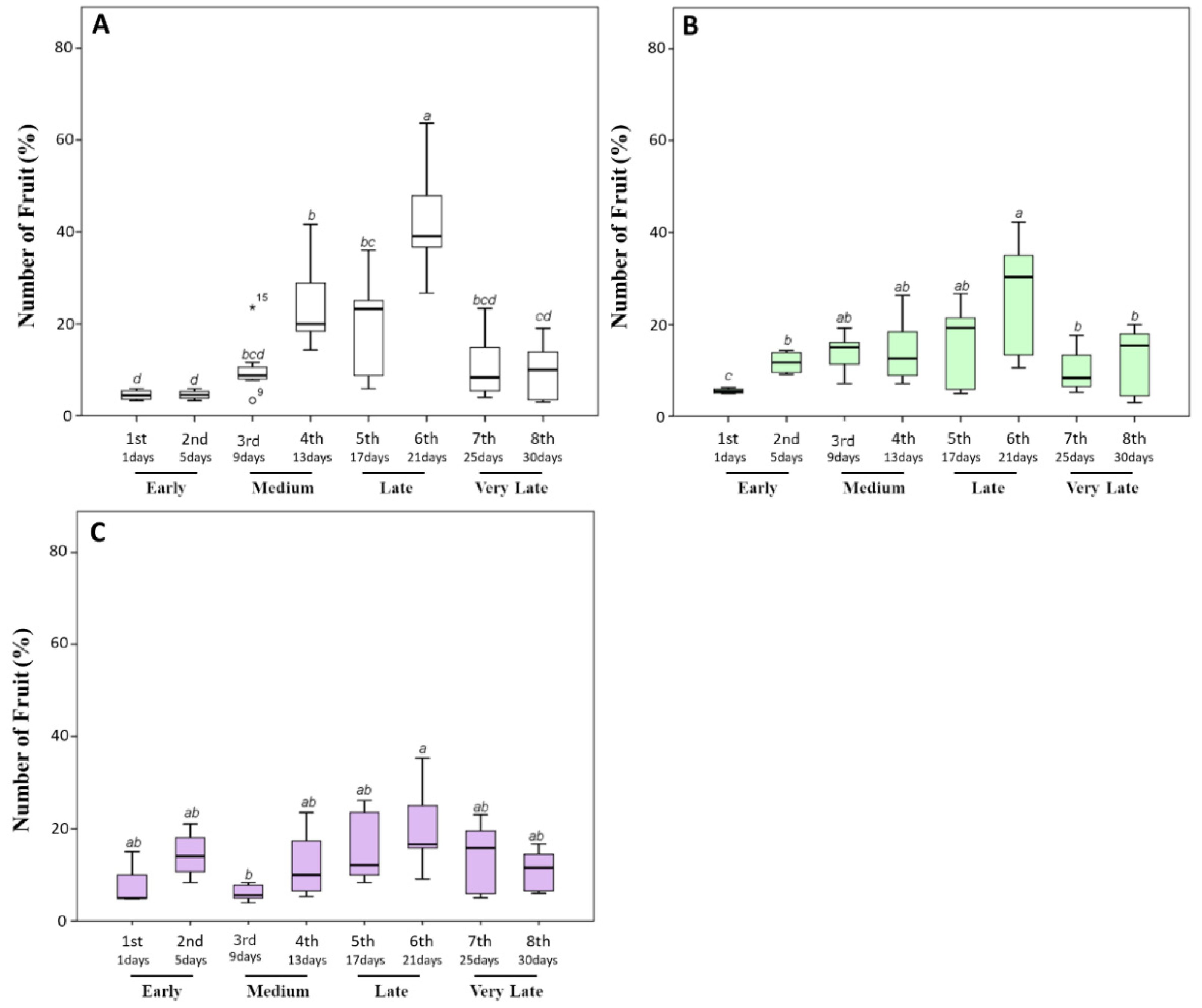
3.1. Biostimulant Treatments Reduced the Ripening Time and Increased Yield in Early Harvest Stages
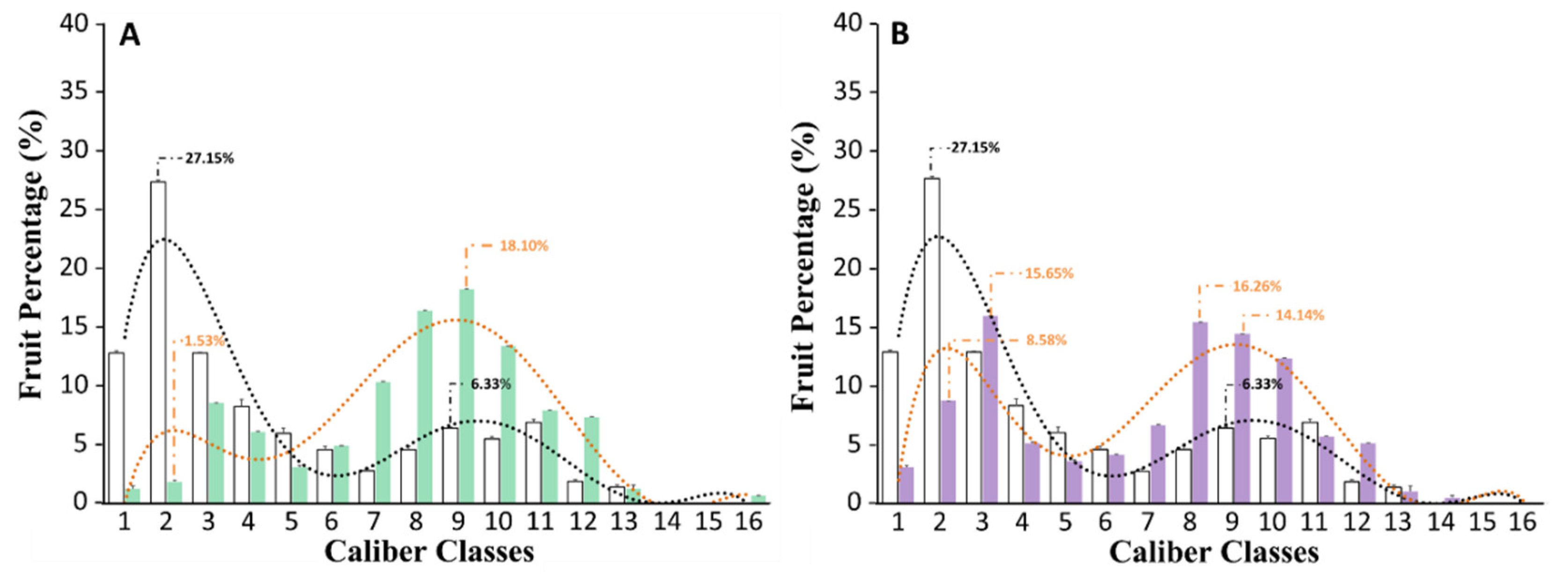
3.2. Biostimulant Treatments Increased the Size of Tomato Fruits
3.3. Biostimulant Treatments Affected the Proximate Composition of Tomato Fruits
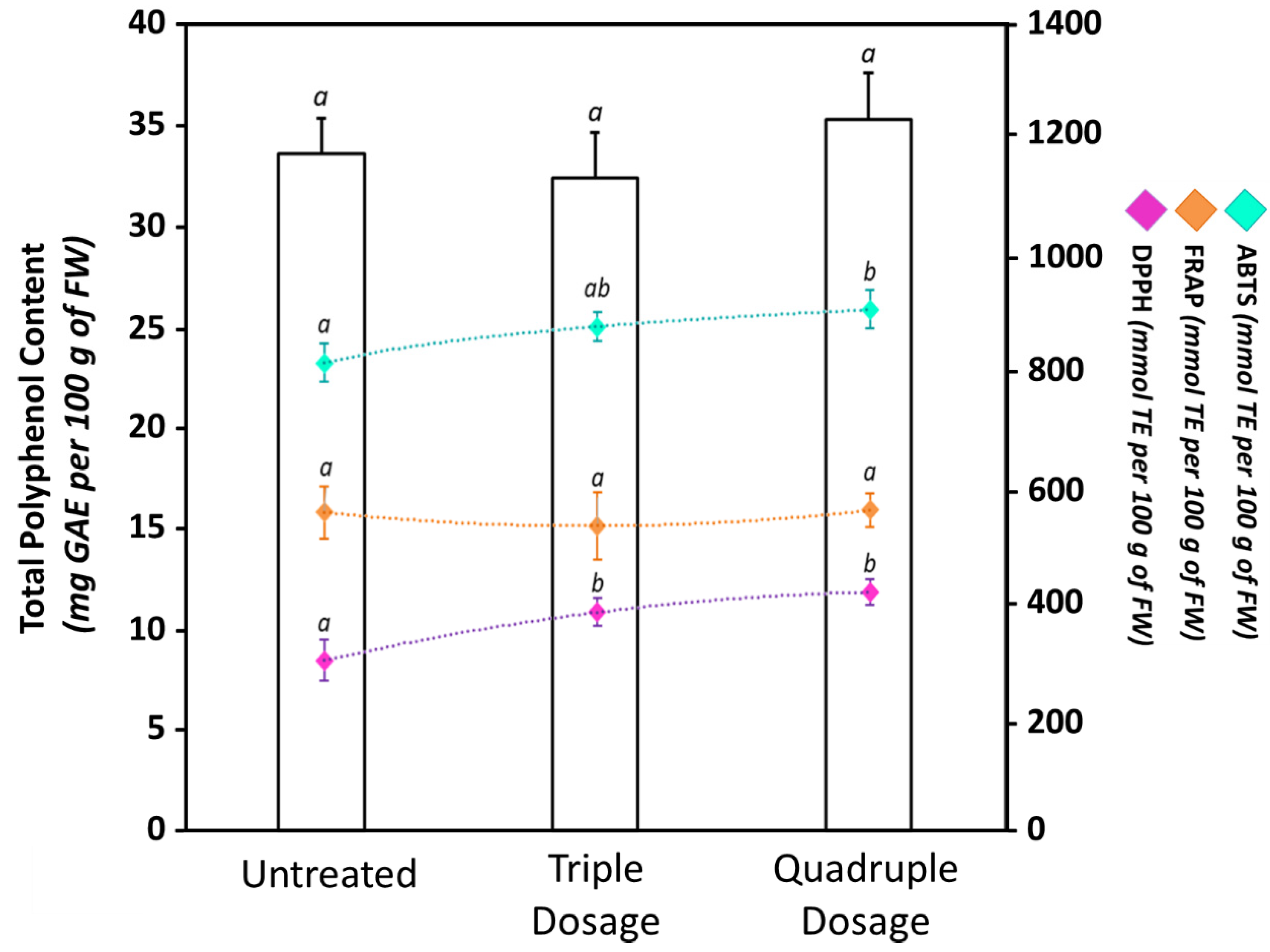
3.4. Biostimulant Treatments Did Not Affect the Phytochemical Composition of Tomato Fruits, but Increased Their Antioxidant Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tripathi, A.D.; Mishra, R.; Maurya, K.K.; Singh, R.B.; Wilson, D.W. Estimates for world population and global food availability for global health. In The Role of Functional Food Security in Global Health; Elsevier: Amsterdam, The Netherlands, 2019; pp. 3–24. [Google Scholar]
- Carvalho, F.P. Agriculture, pesticides, food security and food safety. Environ. Sci. Policy 2006, 9, 685–692. [Google Scholar] [CrossRef]
- Szentpétery, Z.; Hegedűs, Z.; Jolánkai, M. Impact of agrochemicals on yield quality and pesticide residues of winter wheat varieties. Cereal Res. Commun. 2005, 33, 635–640. [Google Scholar] [CrossRef]
- Ramankutty, N.; Mehrabi, Z.; Waha, K.; Jarvis, L.; Kremen, C.; Herrero, M.; Rieseberg, L.H. Trends in global agricultural land use: Implications for environmental health and food security. Annu. Rev. Plant Biol. 2018, 69, 789–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Joshi, N.; Kumar, M. Agrochemicals and human well-being: A review in context of Indian agriculture. Int. J. Cosmet. Sci. 2020, 8, 1539–1543. [Google Scholar] [CrossRef] [Green Version]
- Elahi, E.; Weijun, C.; Zhang, H.; Nazeer, M. Agricultural intensification and damages to human health in relation to agrochemicals: Application of artificial intelligence. Land use policy 2019, 83, 461–474. [Google Scholar] [CrossRef]
- Gentile, C.; Di Gregorio, E.; Di Stefano, V.; Mannino, G.; Perrone, A.; Avellone, G.; Sortino, G.; Inglese, P.; Farina, V. Food quality and nutraceutical value of nine cultivars of mango (Mangifera indica L.) fruits grown in Mediterranean subtropical environment. Food Chem. 2019, 277, 471–479. [Google Scholar] [CrossRef]
- Farina, V.; Tinebra, I.; Perrone, A.; Sortino, G.; Palazzolo, E.; Mannino, G.; Gentile, C. Physicochemical, Nutraceutical and Sensory Traits of Six Papaya (Carica papaya L.) Cultivars Grown in Greenhouse Conditions in the Mediterranean Climate. Agronomy 2020, 10, 501. [Google Scholar] [CrossRef] [Green Version]
- Annunziata, A.; Vecchio, R. Organic farming and sustainability in food choices: An analysis of consumer preference in Southern Italy. Agric. Agric. Sci. Procedia 2016, 8, 193–200. [Google Scholar] [CrossRef]
- Sahota, A.; Haumann, B.; Givens, H.; Baldwin, C.J. Ecolabeling and consumer interest in sustainable products. Sustain. Food Ind. 2009, 159–181. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Matteazzi, A.; Andreotti, C. Foliar Applications of Biostimulants Promote Growth, Yield and Fruit Quality of Strawberry Plants Grown under Nutrient Limitation. Agronomy 2019, 9, 483. [Google Scholar] [CrossRef] [Green Version]
- Graziani, G.; Ritieni, A.; Cirillo, A.; Cice, D.; Di Vaio, C. Effects of Biostimulants on Annurca Fruit Quality and Potential Nutraceutical Compounds at Harvest and during Storage. Plants 2020, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, A.; Lops, F.; Disciglio, G.; Lopriore, G. Effects of plant biostimulants on fruit set, growth, yield and fruit quality attributes of ‘Orange rubis®’apricot (Prunus armeniaca L.) cultivar in two consecutive years. Sci. Hortic. 2018, 239, 26–34. [Google Scholar] [CrossRef]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The Use of a Plant-Based Biostimulant Improves Plant Performances and Fruit Quality in Tomato Plants Grown at Elevated Temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef] [Green Version]
- Abubakar, A.R.; Ashraf, N.; Ashraf, M. Effect of plant biostimulants on fruit cracking and quality attributes of pomegranate cv. Kandhari kabuli. Sci. Res. Essays 2013, 8, 2171–2175. [Google Scholar]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871. [Google Scholar] [CrossRef] [Green Version]
- Bargiacchi, E.; Miele, S.; Romani, A.; Campo, M. Biostimulant activity of hydrolyzable tannins from sweet chestnut (Castanea sativa Mill.). Acta Hortic. 2012, 1009, 111–116. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Agliassa, C.; Acquadro, A.; Contartese, V.; Garabello, C.; Bertea, C.M. Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants 2020, 9, 1308. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Grange, E.; Mannino, G.; Van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance During Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Mannino, G.; Di Stefano, V.; Lauria, A.; Pitonzo, R.; Gentile, C. Vaccinium macrocarpon (Cranberry)-Based Dietary Supplements: Variation in Mass Uniformity, Proanthocyanidin Dosage and Anthocyanin Profile Demonstrates Quality Control Standard Needed. Nutrients 2020, 12, 992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannino, G.; Nerva, L.; Gritli, T.; Novero, M.; Fiorilli, V.; Bacem, M.; Bertea, C.M.; Lumini, E.; Chitarra, W.; Balestrini, R. Effects of Different Microbial Inocula on Tomato Tolerance to Water Deficit. Agronomy 2020, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Xia, H.; Lin, L.; Wang, J.; Yuan, L.; Li, K.; Zhang, J.; Lv, X.; Liang, D. SUNRED, a natural extract-based biostimulant, application stimulates anthocyanin production in the skins of grapes. Sci. Rep. 2019, 9, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrajón-Catalán, E.; Álvarez-Martínez, F.J.; Borrás, F.; Pérez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic analysis of the effects of a commercial complex biostimulant on pepper crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Kumar, R.; Trivedi, K.; Anand, K.G.V.; Ghosh, A. Science behind biostimulant action of seaweed extract on growth and crop yield: Insights into transcriptional changes in roots of maize treated with Kappaphycus alvarezii seaweed extract under soil moisture stressed conditions. J. Appl. Phycol. 2020, 32, 599–613. [Google Scholar] [CrossRef]
- Contartese, V.; Garabello, C.; Occhipinti, A.; Barbero, F.; Bertea, C.M. Effects of a new biostimulant on gene expression and metabolic responses of tomato plants. In Proceedings of the II World Congress on the Use of Biostimulants in Agriculture 1148, Florence, Italy, 16–19 November 2015; pp. 35–42. [Google Scholar]
- Bradstreet, R.B. Kjeldahl method for organic nitrogen. Anal. Chem. 1954, 26, 185–187. [Google Scholar] [CrossRef]
- Jung, S.; Rickert, D.A.; Deak, N.A.; Aldin, E.D.; Recknor, J.; Johnson, L.A.; Murphy, P.A. Comparison of Kjeldahl and Dumas methods for determining protein contents of soybean products. J. Am. Oil Chem. Soc. 2003, 80, 1169. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Maffei, M.E. Chemical partitioning and DNA fingerprinting of some pistachio (Pistacia vera L.) varieties of different geographical origin. Phytochemistry 2019, 160, 40–47. [Google Scholar] [CrossRef]
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical Characterization and DNA Fingerprinting of Griffonia simplicifolia Baill. Molecules 2019, 24, 1032. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, K.L.; Kaur, G. Spectrophotometric determination of L-ascorbic acid in vegetables and fruits. Analyst 1981, 106, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Parimelazhagan, T. Pharmacological Assays of Plant-Based Natural Products; Springer: Berlin/Heidelberg, Germany, 2015; Volume 71, ISBN 3319268112. [Google Scholar]
- Rayhan, M.; Mumtaz, B.; Motalab, M.; Zubair, M.A.; Haque, M.Z.; Saha, B.K. Extraction and Quantification of Lycopene, β-Carotene and Total Phenolic Contents from Papaya (Carica papaya) and Formulation of Lycopene Enriched Fruit Drinks. Am. J. Food Nutr. 2019, 7, 55–63. [Google Scholar]
- Vigliante, I.; Mannino, G.; Maffei, M.E. OxiCyan®, a phytocomplex of bilberry (Vaccinium myrtillus) and spirulina (Spirulina platensis), exerts both direct antioxidant activity and modulation of ARE/Nrf2 pathway in HepG2 cells. J. Funct. Foods 2019, 61, 103508. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I.; Montalba, R. Technological approaches to sustainable agriculture at a crossroads: An agroecological perspective. Sustainability 2017, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Mannino, G.; Gentile, C.; Porcu, A.; Agliassa, C.; Caradonna, F.; Bertea, C.M. Chemical Profile and Biological Activity of Cherimoya (Annona cherimola Mill.) and Atemoya (Annona atemoya) Leaves. Molecules 2020, 25, 2612. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Biostimulants in Agriculture. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Huther, C.M.; Martinazzo, E.G.; Schock, A.A.; Rombaldi, C.V.; Bacarin, M.A. Production components in transformed and untransformed ‘Micro-Tom’tomato plants. Rev. Ciência Agronômica 2018, 49, 85–92. [Google Scholar]
- Martí, E.; Gisbert, C.; Bishop, G.J.; Dixon, M.S.; García-Martínez, J.L. Genetic and physiological characterization of tomato cv. Micro-Tom. J. Exp. Bot. 2006, 57, 2037–2047. [Google Scholar] [CrossRef] [Green Version]
- Gratão, P.L.; Monteiro, C.C.; Antunes, A.M.; Peres, L.E.P.; Azevedo, R.A. Acquired tolerance of tomato (Lycopersicon esculentum cv. Micro-Tom) plants to cadmium-induced stress. Ann. Appl. Biol. 2008, 153, 321–333. [Google Scholar] [CrossRef]
- Rothan, C.; Just, D.; Fernandez, L.; Atienza, I.; Ballias, P.; Lemaire-Chamley, M. Culture of the tomato Micro-Tom cultivar in greenhouse. In Plant Signal Transduction; Springer: Berlin/Heidelberg, Germany, 2016; pp. 57–64. [Google Scholar]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. Hort. Sci. 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Ali, N.; Farrell, A.; Ramsubhag, A.; Jayaraman, J. The effect of Ascophyllum nodosum extract on the growth, yield and fruit quality of tomato grown under tropical conditions. J. Appl. Phycol. 2016, 28, 1353–1362. [Google Scholar] [CrossRef]
- Cerdán, M.; Sánchez-Sánchez, A.; Oliver, M.; Juárez, M.; Sánchez-Andreu, J.J. Effect of foliar and root applications of amino acids on iron uptake by tomato plants. In Proceedings of the IV Balkan Symposium on Vegetables and Potatoes 830, Plovdiv, Bulgaria, 9–12 September 2008; pp. 481–488. [Google Scholar]
- Guardiola, J.L.; García-Luis, A. Increasing fruit size in Citrus. Thinning and stimulation of fruit growth. Plant Growth Regul. 2000, 31, 121–132. [Google Scholar] [CrossRef]
- Kavipriya, R.; Dhanalakshmi, P.K.; Jayashree, S.; Thangaraju, N. Seaweed extract as a biostimulant for legume crop, green gram. J. Ecobiotechnol. 2011, 3, 16–19. [Google Scholar]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and nutritional quality of Vesuvian Piennolo tomato PDO as affected by farming system and biostimulant application. Agronomy 2019, 9, 505. [Google Scholar] [CrossRef] [Green Version]
- Benítez, J.J.; Castillo, P.M.; Del Río, J.C.; León-Camacho, M.; Domínguez, E.; Heredia, A.; Guzmán-Puyol, S.; Athanassiou, A.; Heredia-Guerrero, J.A. Valorization of tomato processing by-products: Fatty acid extraction and production of bio-based materials. Materials 2018, 11, 2211. [Google Scholar]
- Simopoulos, A.P. Omega-6/omega-3 essential fatty acid ratio and chronic diseases. Food Rev. Int. 2004, 20, 77–90. [Google Scholar] [CrossRef]
- Rattan, R.K.; Patel, K.P.; Manjaiah, K.M.; Datta, S.P. Micronutrients in soil, plant, animal and human health. J. Indian Soc. Soil Sci. 2009, 57, 546–558. [Google Scholar]
- Bhat, B.A.; Islam, S.T.; Ali, A.; Sheikh, B.A.; Tariq, L.; Islam, S.U.; Dar, T.U.H. Role of Micronutrients in Secondary Metabolism of Plants. In Plant Micronutrients; Springer: Berlin/Heidelberg, Germany, 2020; pp. 311–329. [Google Scholar]
- Römheld, V.; Marschner, H. Function of micronutrients in plants. Micronutr. Agric. 1991, 4, 297–328. [Google Scholar]
- Gomez, P.; Ferrer, M.Á.; Fernández-Trujillo, J.P.; Calderon, A.; Artes, F.; Egea-Cortines, M.; Weiss, J. Structural changes, chemical composition and antioxidant activity of cherry tomato fruits (cv. Micro-Tom) stored under optimal and chilling conditions. J. Sci. Food Agric. 2009, 89, 1543–1551. [Google Scholar] [CrossRef]
- Gupta, U.; Gupta, S. Role of vitamins in human health and nutrition: Sources and morbidity. Curr. Nutr. Food Sci. 2015, 11, 105–115. [Google Scholar] [CrossRef]
- Chudow, M.; Adams, B. ABC’s of Vitamin Supplementation in Critical Illness. J. Pharm. Pract. 2020, 0897190020958234. [Google Scholar] [CrossRef] [PubMed]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; Di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Kocira, A.; Świeca, M.; Kocira, S.; Złotek, U.; Jakubczyk, A. Enhancement of yield, nutritional and nutraceutical properties of two common bean cultivars following the application of seaweed extract (Ecklonia maxima). Saudi J. Biol. Sci. 2018, 25, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Kocira, A.; Kocira, S.; Świeca, M.; Złotek, U.; Jakubczyk, A.; Kapela, K. Effect of foliar application of a nitrophenolate–based biostimulant on the yield and quality of two bean cultivars. Sci. Hortic. 2017, 214, 76–82. [Google Scholar] [CrossRef]
- El-Garhy, H.A.S.; Abdel-Rahman, F.A.; Shams, A.S.; Osman, G.H.; Moustafa, M. Comparative Analyses of Four Chemicals Used to Control Black Mold Disease in Tomato and Its Effects on Defense Signaling Pathways, Productivity and Quality Traits. Plants 2020, 9, 808. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, A.; Kunicki, E.; Jezdinsky, A.; Kalisz, A.; Sekara, A. The effect of biostimulants on the quality parameters of tomato grown for the processing industry. Eff. Biostimulants Qual. Parameters Tomato Grown Process. Ind. 2015, 203–217. [Google Scholar]
- Sut, S.; Ferrarese, I.; Shrestha, S.S.; Kumar, G.; Slaviero, A.; Sello, S.; Altissimo, A.; Pagni, L.; Gattesco, F.; Dall’Acqua, S. Comparison of Biostimulant Treatments in Acmella oleracea Cultivation for Alkylamides Production. Plants 2020, 9, 818. [Google Scholar] [CrossRef]
- Mannino, G.; Perrone, A.; Campobenedetto, C.; Schittone, A.; Margherita Bertea, C.; Gentile, C. Phytochemical profile and antioxidative properties of Plinia trunciflora fruits: A new source of nutraceuticals. Food Chem. 2020, 307. [Google Scholar] [CrossRef]
- Polo, J.; Mata, P. Evaluation of a biostimulant (Pepton) based in enzymatic hydrolyzed animal protein in comparison to seaweed extracts on root development, vegetative growth, flowering, and yield of gold cherry tomatoes grown under low stress ambient field conditions. Front. Plant Sci. 2018, 8, 2261. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Untreated | Triple Dosage | Quadruple Dosage | |
|---|---|---|---|
| Nutritional Content(g per 100 g of FW) | |||
| Protein content | 14.569 ± 0.201 b | 15.038 ± 0.138 a | 14.712 ± 0.194 ab |
| Fat content | 0.528 ± 0.016 b | 0.631 ± 0.018 a | 0.531 ± 0.016 b |
| Sugar content | 2.285 ± 0.034 b | 2.411 ± 0.078 a | 2.364 ± 0.051 ab |
| TSS (°Brix) | 6.832 ± 0.321 a | 7.001 ± 0.175 a | 7.122 ± 0.489 a |
| Moisture content | 90.438 ± 0.154 a | 90.252 ± 0.143 a | 89.088 ± 0.015 b |
| Ashes | 0.761 ± 0.021 a | 0.783 ± 0.012 a | 0.758 ± 0.009 a |
| Available energy (kcal) | 72.168 ± 1.084 c | 75.475 ± 1.026 a | 73.083 ± 1.124 b |
| Mineral Content(mg per 100 g of FW) | |||
| K | 541.671 ± 0.772 ab | 549.724 ± 4.953 a | 529.188 ± 9.972 b |
| Na | 10.874 ± 0.283 c | 13.535 ± 0.471 a | 12.799 ± 0.271 b |
| Ca | 27.653 ± 0.469 a | 23.612 ± 0.425 b | 23.248 ± 0.139 b |
| Mg | 20.574 ± 0.053 a | 21.027 ± 0.375 a | 20.977 ± 0.424 a |
| P | 113.791 ± 2.546 b | 126.579 ± 2.487 a | 126.637 ± 0.996 a |
| Cl | 2.753 ± 0.033 b | 2.394 ± 0.080 c | 2.985 ± 0.047 a |
| Fe | 0.505 ± 0.022 b | 0.577 ± 0.014 a | 0.563 ± 0.02 a |
| Cu | 0.096 ± 0.003 b | 0.117 ± 0.008 a | 0.111 ± 0.007 a |
| Zn | 0.089 ± 0.005 c | 0.111 ± 0.006 a | 0.103 ± 0.002 b |
| Mn | 0.145 ± 0.002 b | 0.163 ± 0.004 a | 0.165 ± 0.006 a |
| Si | 0.506 ± 0.013 b | 0.625 ± 0.021 a | 0.386 ± 0.003 c |
| B | 0.046 ± 0.005 a | 0.052 ± 0.003 a | 0.049 ± 0.004 a |
| Mo | 0.013 ± 0.001 b | 0.016 ± 0.002 a | 0.014 ± 0.002 ab |
| Bioactive Compound Content(mg per 100 g of FW) | |||
| Lycopene | 7.182 ± 0.173 a | 6.972 ± 0.312 a | 7.013 ± 0.392 a |
| Carotenoids | 13.524 ± 1.235 a | 14.091 ± 0.924 a | 13.892 ± 0.783 a |
| Tocopherols | 0.398 ± 0.057 b | 0.424 ± 0.036 ab | 0.445 ± 0.074 a |
| Ascorbic acid | 15.925 ± 1.267 a | 14.180 ± 2.202 a | 14.781 ± 1.892 a |
| Polyphenols | 35.652 ± 1.723 a | 33.420 ± 2.279 a | 35.318 ± 2.323 a |
| Untreated | Triple Dosage | Quadruple Dosage | |
|---|---|---|---|
| C16:3ω3 | 0.202 ± 0.026 b | 0.866 ± 0.009 a | 0.236 ± 0.007 b |
| C16:1ω7 | 0.431 ± 0.044 b | 0.655 ± 0.102 a | 0.404 ± 0.015 b |
| C16:1ω10 | 0.089 ± 0.017 a | 0.155 ± 0.012 a | 0.079 ± 0.054 a |
| C16:0 | 19.734 ± 0.935 a | 19.604 ± 0.150 a | 19.997 ± 1.925 a |
| C18:2ω6 | 48.984 ± 2.988 a | 37.964 ± 0.013 b | 49.220 ± 3.007 a |
| C18:1ω9 | 24.441 ± 1.262 a | 27.154 ± 2.273 a | 23.848 ± 0.359 a |
| C18:1ω9 | 1.410 ± 0.122 b | 2.352 ± 0.039 a | 1.332 ± 0.045 b |
| C18:0 | 4.128 ± 0.181 a | 4.060 ± 0.045 a | 4.516 ± 0.336 a |
| SFA | 23.863 ± 1.117 b | 29.664 ± 0.195 a | 24.514 ± 2.261 b |
| UFA | 75.559 ± 4.459 a | 69.149 ± 2.449 a | 75.122 ± 3.490 a |
| MUFA | 26.372 ± 1.445 b | 30.318 ± 2.427 a | 25.664 ± 0.475 b |
| PUFA | 49.187 ± 3.014 a | 38.831 ± 0.022 b | 49.457 ± 3.015 a |
| SFA/UFA | 0.315 ± 0.013 b | 0.428 ± 0.014 a | 0.326 ± 0.009 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. https://doi.org/10.3390/biom10121662
Mannino G, Campobenedetto C, Vigliante I, Contartese V, Gentile C, Bertea CM. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules. 2020; 10(12):1662. https://doi.org/10.3390/biom10121662
Chicago/Turabian StyleMannino, Giuseppe, Cristina Campobenedetto, Ivano Vigliante, Valeria Contartese, Carla Gentile, and Cinzia M. Bertea. 2020. "The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality" Biomolecules 10, no. 12: 1662. https://doi.org/10.3390/biom10121662