A Synthetic CPP33-Conjugated HOXA9 Active Domain Peptide Inhibits Invasion Ability of Non-Small Lung Cancer Cells


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. DNA Cloning, Transfection, and Peptides
2.3. Cell Invasion Assay
2.4. Quantitative Realtime PCR (qRT-PCR) Analysis
2.5. NF-kB Reporter Gene Assay
2.6. Chromatin Immunoprecipitation (ChIP) Assay
2.7. Cell Cytotoxicity Assay
2.8. Intracellular Transduction of Synthetic Cell-Penetrating Peptide
2.9. In Vivo Metastasis
2.10. Hematoxylin and Eosin Staining and Immunohistochemical Analysis
2.11. Statistical Analyses
3. Results
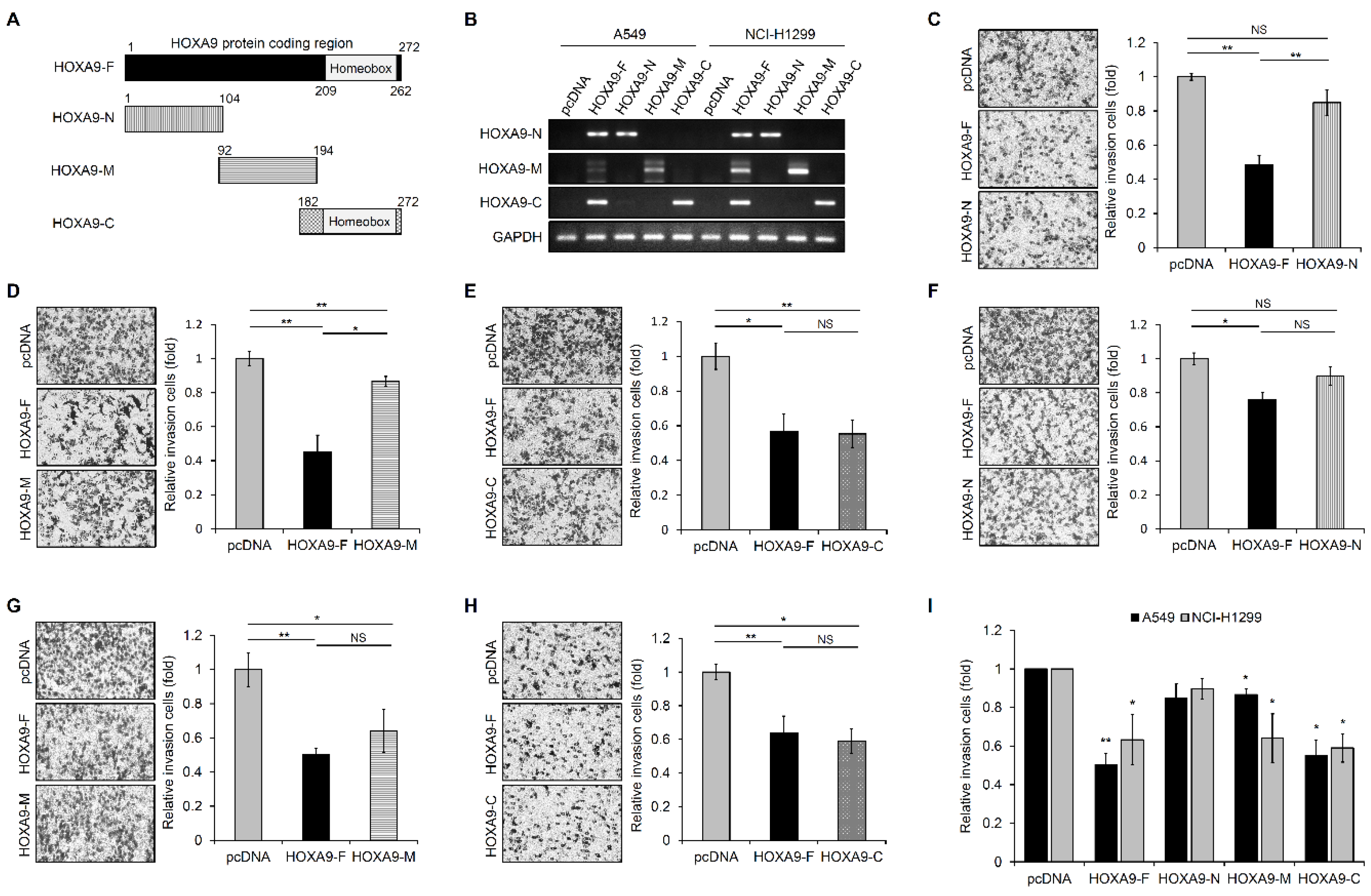
3.1. HOXA9-C Fragment Inhibits Cell Invasion in NSCLC Cells
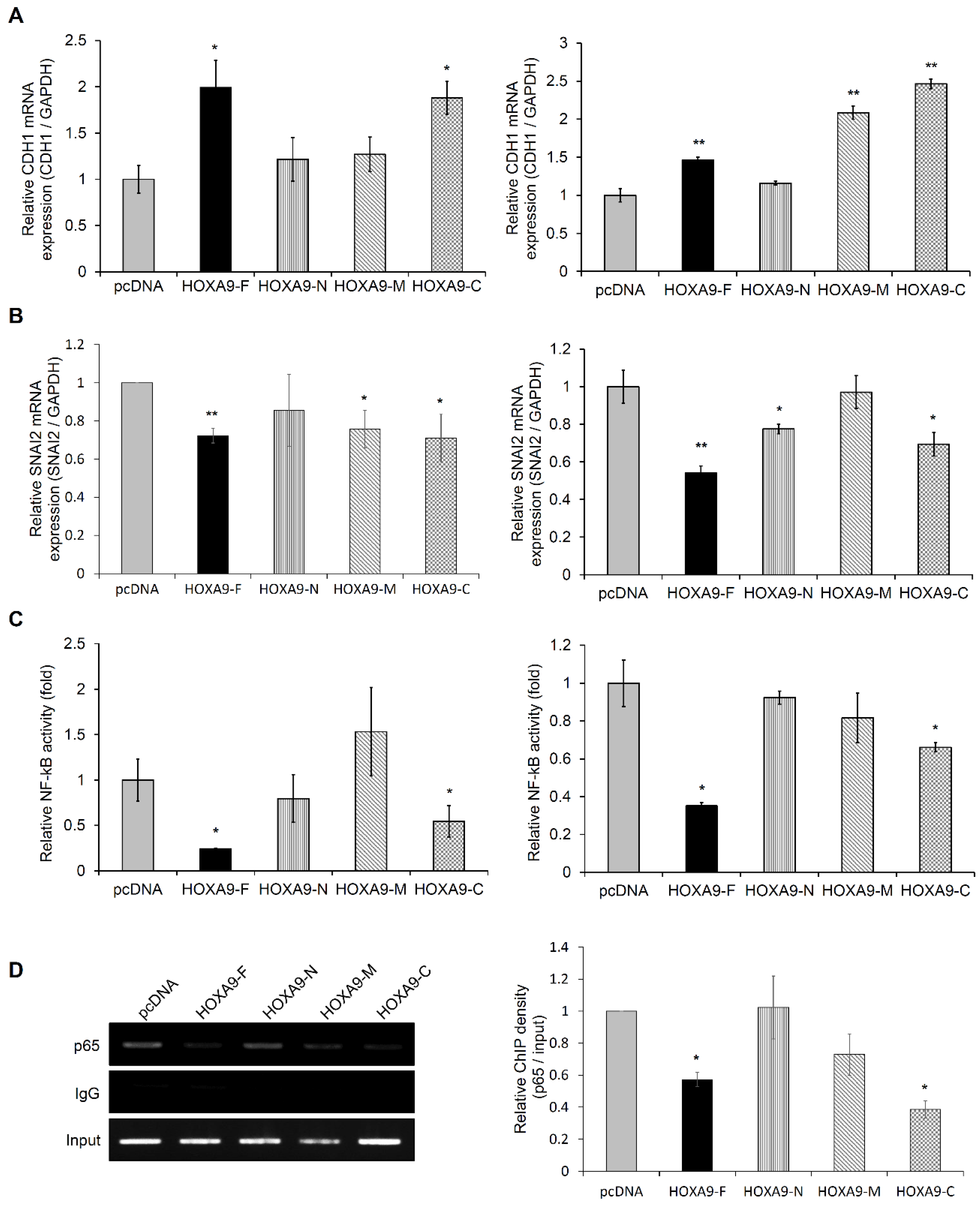
3.2. The HOXA9-C Fragment Regulates Expression of EMT-Related Genes in NSCLC Cells
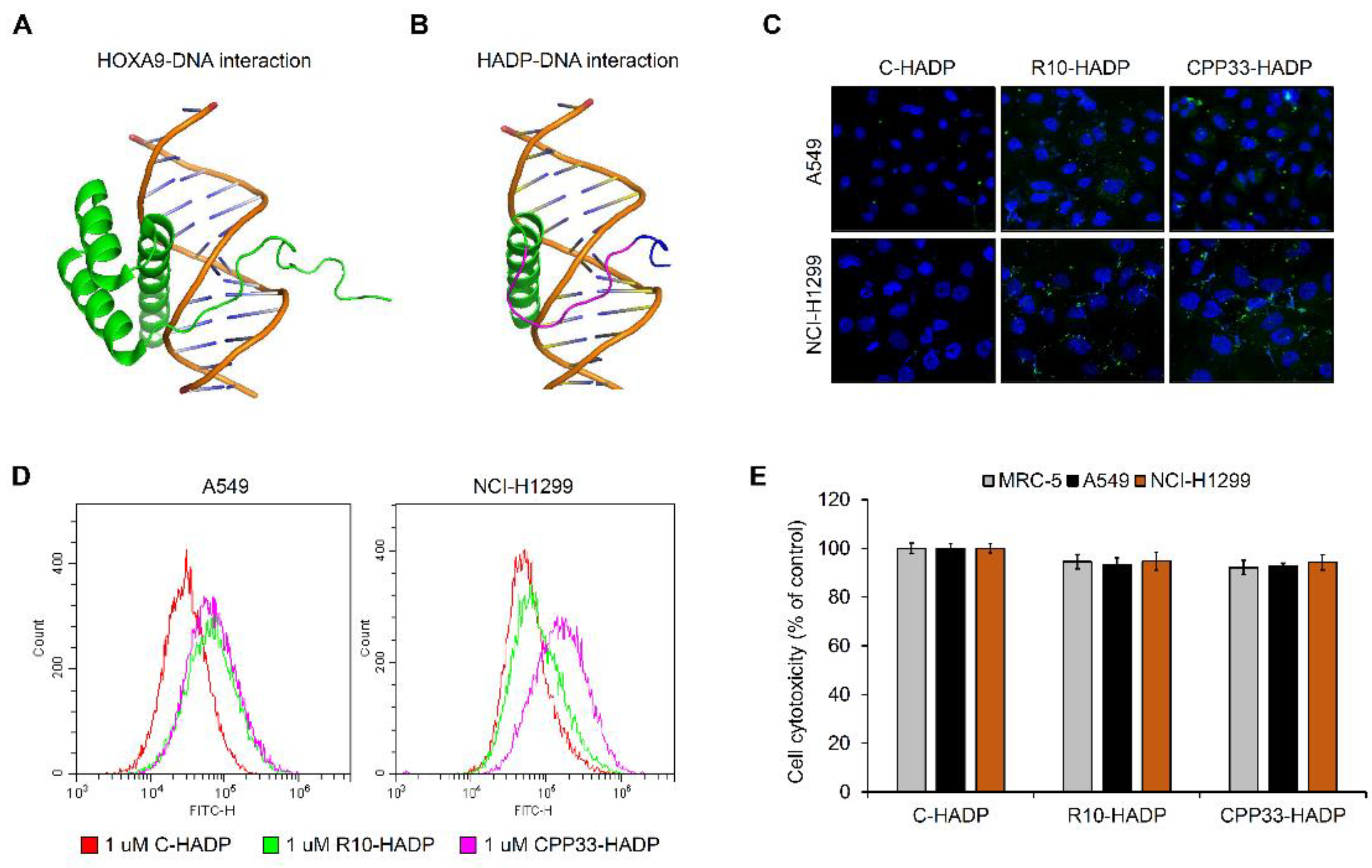
3.3. Intracellular Transduction of Synthetic Cell-Penetrating Peptide HADP (HOXA9 Active Domain Peptide)
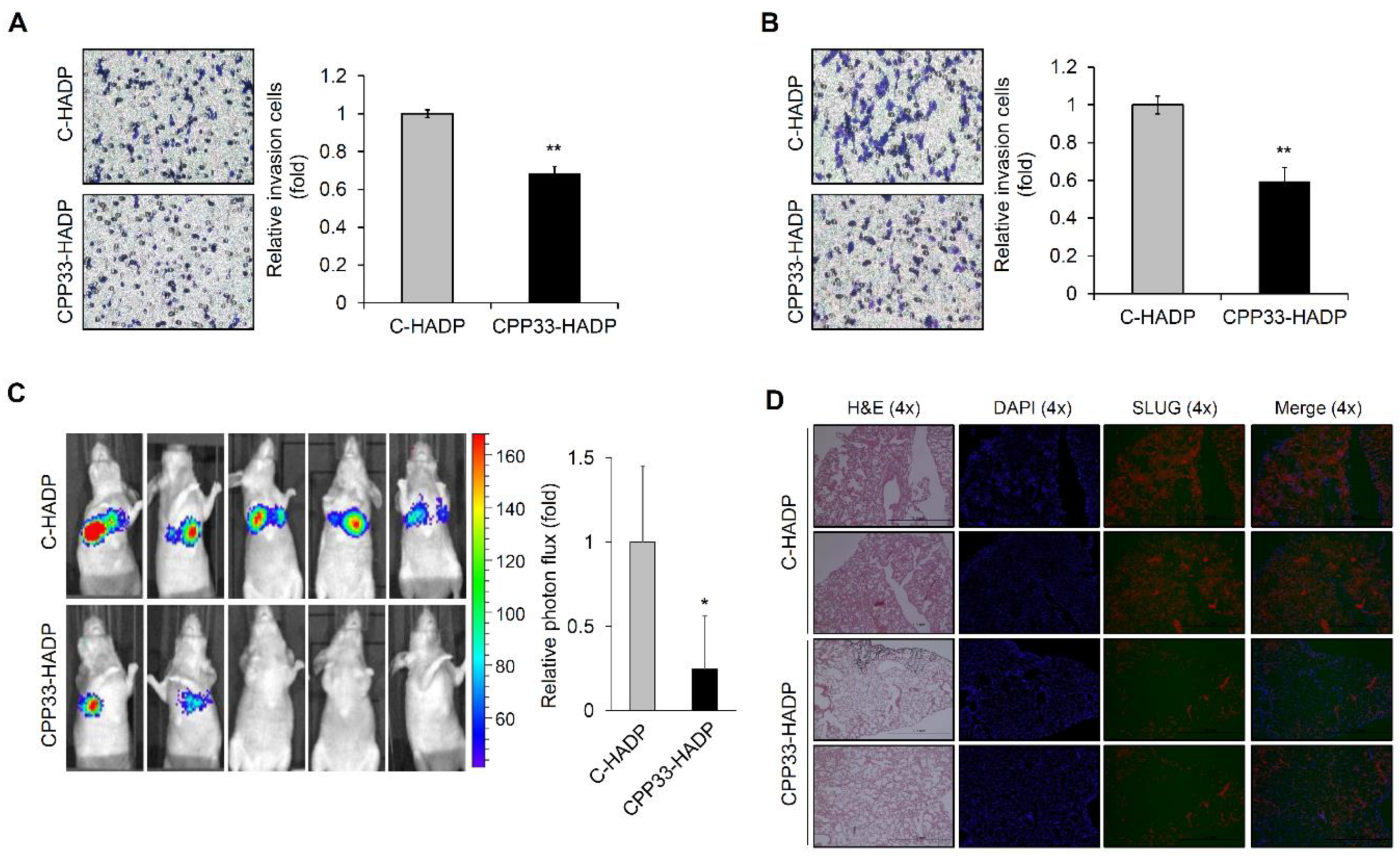
3.4. CPP33-HADP Inhibits Invasion Ability of NSCLC Cells in Both In Vitro and In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J.; Hiromi, Y. Homeotic genes and the homeobox. Annu. Rev. Genet. 1986, 20, 147–173. [Google Scholar] [CrossRef]
- Scott, M.P. Vertebrate homeobox gene nomenclature. Cell 1992, 71, 551–553. [Google Scholar] [CrossRef]
- Lelli, K.M.; Noro, B.; Mann, R.S. Variable motif utilization in homeotic selector (Hox)-cofactor complex formation controls specificity. Proc. Natl. Acad. Sci. USA 2011, 108, 21122–21127. [Google Scholar] [CrossRef] [Green Version]
- Bhatlekar, S.; Fields, J.Z.; Boman, B.M. HOX genes and their role in the development of human cancers. J. Mol. Med. 2014, 92, 811–823. [Google Scholar] [CrossRef]
- Son, J.W.; Jeong, K.J.; Jean, W.S.; Park, S.Y.; Jheon, S.; Cho, H.M.; Park, C.G.; Lee, H.Y.; Kang, J. Genome-wide combination profiling of DNA copy number and methylation for deciphering biomarkers in non-small cell lung cancer patients. Cancer Lett. 2011, 311, 29–37. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, K.U.; Kim, J.E.; Kim, H.H.; Lee, M.K.; Lee, C.H.; Lee, S.Y.; Oh, T.; An, S. Detection of HOXA9 gene methylation in tumor tissues and induced sputum samples from primary lung cancer patients. Clin. Chem. Lab. Med. Cclm/Fescc 2011, 49, 699–704. [Google Scholar] [CrossRef]
- LaRonde-LeBlanc, N.A.; Wolberger, C. Structure of HoxA9 and Pbx1 bound to DNA: Hox hexapeptide and DNA recognition anterior to posterior. Genes Dev. 2003, 17, 2060–2072. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.L.; Lee, D.C.; Sohn, H.A.; Lee, S.Y.; Jeon, H.S.; Lee, J.H.; Park, C.G.; Lee, H.Y.; Yeom, Y.I.; Son, J.W.; et al. Homeobox A9 directly targeted by miR-196b regulates aggressiveness through nuclear Factor-kappa B activity in non-small cell lung cancer cells. Mol. Carcinog. 2016, 55, 1915–1926. [Google Scholar] [CrossRef]
- Baum, B.; Settleman, J.; Quinlan, M.P. Transitions between epithelial and mesenchymal states in development and disease. Semin. Cell Dev. Biol. 2008, 19, 294–308. [Google Scholar] [CrossRef]
- Frankel, A.D.; Bredt, D.S.; Pabo, C.O. Tat protein from human immunodeficiency virus forms a metal-linked dimer. Science 1988, 240, 70–73. [Google Scholar] [CrossRef] [Green Version]
- Green, M.; Loewenstein, P.M. Autonomous functional domains of chemically synthesized human immunodeficiency virus tat trans-activator protein. Cell 1988, 55, 1179–1188. [Google Scholar] [CrossRef]
- Zahid, M.; Robbins, P.D. Cell-type specific penetrating peptides: Therapeutic promises and challenges. Molecules 2015, 20, 13055–13070. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.T.; Zhou, J.; Gore, C.; Zimmern, P. R11, a novel cell-permeable peptide, as an intravesical delivery vehicle. Bju Int. 2011, 108, 1666–1671. [Google Scholar] [CrossRef]
- Kondo, E.; Saito, K.; Tashiro, Y.; Kamide, K.; Uno, S.; Furuya, T.; Mashita, M.; Nakajima, K.; Tsumuraya, T.; Kobayashi, N.; et al. Tumour lineage-homing cell-penetrating peptides as anticancer molecular delivery systems. Nat. Commun. 2012, 3, 951. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, X.; Chen, H.; Bian, Z.; Zhang, G.; Riaz, M.K.; Tyagi, D.; Lin, G.; Zhang, Y.; Wang, J.; et al. Dual-ligand modified liposomes provide effective local targeted delivery of lung-cancer drug by antibody and tumor lineage-homing cell-penetrating peptide. Drug Deliv. 2018, 25, 256–266. [Google Scholar] [CrossRef]
- Chen, R.; Braun, G.B.; Luo, X.; Sugahara, K.N.; Teesalu, T.; Ruoslahti, E. Application of a proapoptotic peptide to intratumorally spreading cancer therapy. Cancer Res. 2013, 73, 1352–1361. [Google Scholar] [CrossRef] [Green Version]
- de Araujo, C.B.; Russo, L.C.; Castro, L.M.; Forti, F.L.; do Monte, E.R.; Rioli, V.; Gozzo, F.C.; Colquhoun, A.; Ferro, E.S. A novel intracellular peptide derived from g1/s cyclin d2 induces cell death. J. Biol. Chem. 2014, 289, 16711–16726. [Google Scholar] [CrossRef] [Green Version]
- Higa, M.; Katagiri, C.; Shimizu-Okabe, C.; Tsumuraya, T.; Sunagawa, M.; Nakamura, M.; Ishiuchi, S.; Takayama, C.; Kondo, E.; Matsushita, M. Identification of a novel cell-penetrating peptide targeting human glioblastoma cell lines as a cancer-homing transporter. Biochem. Biophys. Res. Commun. 2015, 457, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.H.; Hsu, C.Y.; Tsai, C.F.; Chiu, C.C.; Liang, S.S.; Wang, T.N.; Kuo, P.L.; Long, C.Y.; Tsai, E.M. A novel cell-penetrating peptide suppresses breast tumorigenesis by inhibiting beta-catenin/LEF-1 signaling. Sci. Rep. 2016, 6, 19156. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.L.; Koo, H.; Lee, H.Y.; Yeom, Y.I.; Lee, D.C.; Kang, J. Recombinant cell-permeable HOXA9 protein inhibits NSCLC cell migration and invasion. Cell Oncol. 2019, 42, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Cho, K.H.; Lee, S.; Bae, Y.J.; Jeong, K.J.; Rha, S.Y.; Choi, E.J.; Park, J.H.; Kim, J.M.; Lee, J.S.; et al. hTERT mediates norepinephrine-induced Slug expression and ovarian cancer aggressiveness. Oncogene 2015, 34, 3402–3412. [Google Scholar] [CrossRef] [PubMed]
- Janson, G.; Zhang, C.; Prado, M.G.; Paiardini, A. PyMod 2.0: Improvements in protein sequence-structure analysis and homology modeling within PyMOL. Bioinformatics 2017, 33, 444–446. [Google Scholar] [CrossRef] [Green Version]
- Thorsteinsdottir, U.; Mamo, A.; Kroon, E.; Jerome, L.; Bijl, J.; Lawrence, H.J.; Humphries, K.; Sauvageau, G. Overexpression of the myeloid leukemia-associated Hoxa9 gene in bone marrow cells induces stem cell expansion. Blood 2002, 99, 121–129. [Google Scholar] [CrossRef]
- Pojo, M.; Goncalves, C.S.; Xavier-Magalhaes, A.; Oliveira, A.I.; Goncalves, T.; Correia, S.; Rodrigues, A.J.; Costa, S.; Pinto, L.; Pinto, A.A.; et al. A transcriptomic signature mediated by HOXA9 promotes human glioblastoma initiation, aggressiveness and resistance to temozolomide. Oncotarget 2015, 6, 7657–7674. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.Y.; Ladanyi, A.; Lengyel, E.; Naora, H. Expression of the homeobox gene HOXA9 in ovarian cancer induces peritoneal macrophages to acquire an M2 tumor-promoting phenotype. Am. J. Pathol. 2014, 184, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.M.; Mouw, J.K.; Unger, M.A.; Lakins, J.N.; Gbegnon, M.K.; Clemmer, V.B.; Benezra, M.; Licht, J.D.; Boudreau, N.J.; Tsai, K.K.; et al. HOXA9 regulates BRCA1 expression to modulate human breast tumor phenotype. J. Clin. Investig. 2010, 120, 1535–1550. [Google Scholar] [CrossRef]
- Alvarado-Ruiz, L.; Martinez-Silva, M.G.; Torres-Reyes, L.A.; Pina-Sanchez, P.; Ortiz-Lazareno, P.; Bravo-Cuellar, A.; Aguilar-Lemarroy, A.; Jave-Suarez, L.F. HOXA9 is Underexpressed in Cervical Cancer Cells and its Restoration Decreases Proliferation, Migration and Expression of Epithelial-to-Mesenchymal Transition Genes. Asian Pac. J. Cancer Prev. APJCP 2016, 17, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Dong, L.; Qu, X.; Li, K.; Wang, H.; Cheng, L.; He, H.; Wang, L.; Qi, Z.; Ren, H.; et al. Down-expression of homeobox A9 promotes cancer cell invasion in human hepatocellular carcinomas. Int. J. Clin. Exp. Pathol. 2016, 9, 563–575. [Google Scholar]
- Hwang, J.A.; Lee, B.B.; Kim, Y.; Hong, S.H.; Kim, Y.H.; Han, J.; Shim, Y.M.; Yoon, C.Y.; Lee, Y.S.; Kim, D.H. HOXA9 inhibits migration of lung cancer cells and its hypermethylation is associated with recurrence in non-small cell lung cancer. Mol. Carcinog. 2015, 54 (Suppl. 1), E72–E80. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Allison, D.F.; Baranova, N.N.; Wamsley, J.J.; Katz, A.J.; Bekiranov, S.; Jones, D.R.; Mayo, M.W. NF-kappaB regulates mesenchymal transition for the induction of non-small cell lung cancer initiating cells. PLoS ONE 2013, 8, e68597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, P.; Sukumar, S. HOX genes and the NF-kappaB pathway: A convergence of developmental biology, inflammation and cancer biology. Biochim. Biophys. Acta. Rev. Cancer 2020, 1874, 188450. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.; Riley, T.; Liu, P.; Abe, N.; Gomez-Alcala, P.; Dror, I.; Zhou, T.; Rohs, R.; Honig, B.; Bussemaker, H.J.; et al. Cofactor binding evokes latent differences in DNA binding specificity between Hox proteins. Cell 2011, 147, 1270–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dard, A.; Jia, Y.; Reboulet, J.; Bleicher, F.; Lavau, C.; Merabet, S. The human HOXA9 protein uses paralog-specific residues of the homeodomain to interact with TALE-class cofactors. Sci. Rep. 2019, 9, 5664. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Gorman, G.S.; Coward, L.U.; Noker, P.E.; McCormick, D.; Horn, T.L.; Harder, J.B.; Muzzio, M.; Prabhakar, B.; Ganesh, B.; et al. Preclinical pharmacokinetics, metabolism, and toxicity of azurin-p28 (NSC745104) a peptide inhibitor of p53 ubiquitination. Cancer Chemother. Pharmacol. 2011, 68, 513–524. [Google Scholar] [CrossRef]
- Warso, M.A.; Richards, J.M.; Mehta, D.; Christov, K.; Schaeffer, C.; Rae Bressler, L.; Yamada, T.; Majumdar, D.; Kennedy, S.A.; Beattie, C.W.; et al. A first-in-class, first-in-human, phase I trial of p28, a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in patients with advanced solid tumours. Br. J. Cancer 2013, 108, 1061–1070. [Google Scholar] [CrossRef]
- Habault, J.; Poyet, J.L. Recent Advances in Cell Penetrating Peptide-Based Anticancer Therapies. Molecules 2019, 24, 927. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-L.; Koo, H.; Lee, S.-I.; Kang, J.; Han, Y.-H.; Yeom, Y.I.; Lee, D.C. A Synthetic CPP33-Conjugated HOXA9 Active Domain Peptide Inhibits Invasion Ability of Non-Small Lung Cancer Cells. Biomolecules 2020, 10, 1589. https://doi.org/10.3390/biom10111589
Yu S-L, Koo H, Lee S-I, Kang J, Han Y-H, Yeom YI, Lee DC. A Synthetic CPP33-Conjugated HOXA9 Active Domain Peptide Inhibits Invasion Ability of Non-Small Lung Cancer Cells. Biomolecules. 2020; 10(11):1589. https://doi.org/10.3390/biom10111589
Chicago/Turabian StyleYu, Seong-Lan, Han Koo, Se-In Lee, JaeKu Kang, Young-Hyun Han, Young Il Yeom, and Dong Chul Lee. 2020. "A Synthetic CPP33-Conjugated HOXA9 Active Domain Peptide Inhibits Invasion Ability of Non-Small Lung Cancer Cells" Biomolecules 10, no. 11: 1589. https://doi.org/10.3390/biom10111589