Nephroprotective Effect of Pleurotus ostreatus and Agaricus bisporus Extracts and Carvedilol on Ethylene Glycol-Induced Urolithiasis: Roles of NF-κB, p53, Bcl-2, Bax and Bak

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Mushrooms
2.3. Preliminary Phytochemical Screening of Mushrooms
2.4. Preparation of Mushroom Aqueous Extracts
2.5. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis of Aqueous extracts
2.6. Preparation of Carvedilol Dose
2.7. Induction of Hyperoxaluria
2.8. Animal Grouping
2.9. Blood and Tissue Sampling
2.10. Biochemical Analysis
2.11. Determination of TNF-α and IL-1β Levels by ELISA
2.12. Histological Investigations
2.13. Immunohistochemical Investigations
2.14. Imaging and Semi-Quantitative Analysis of NF-κB, p53 and Bcl-2 Expressions
2.15. Western Blot Analysis
2.16. Statistical Analysis
3. Results
3.1. Phytochemical Screening
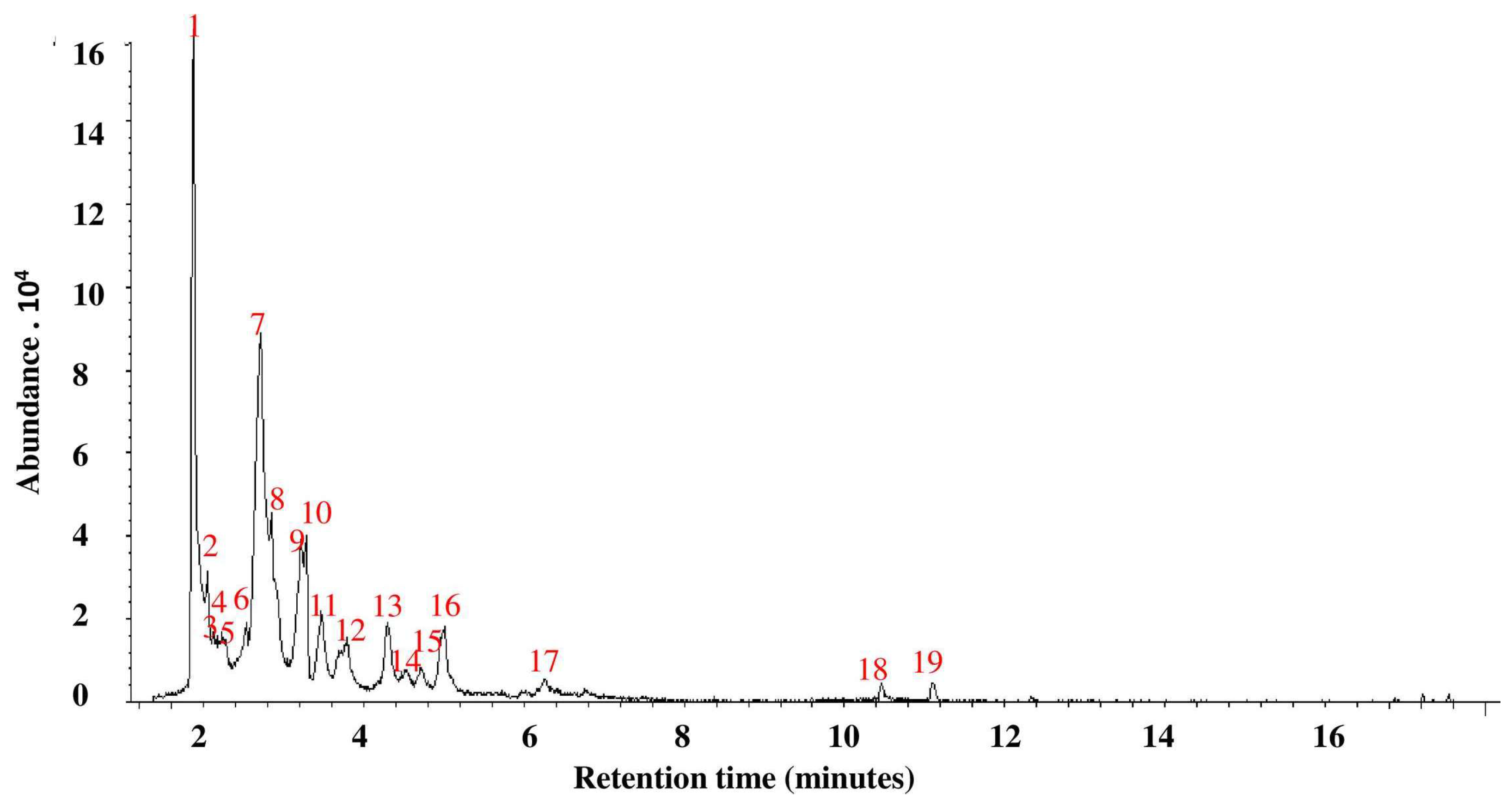
3.2. GC-MS Analysis of P. ostreatus Aqueous Extract
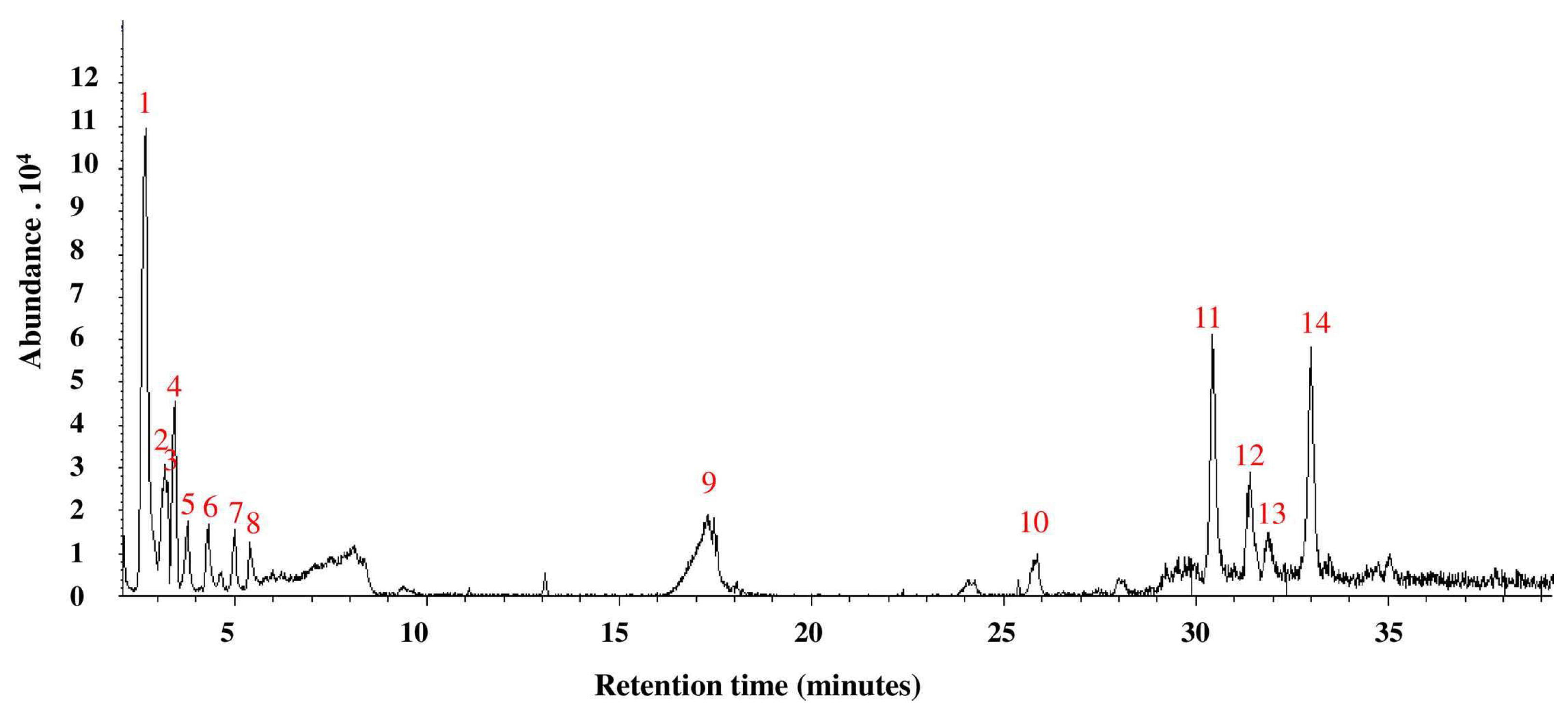
3.3. GC-MS Analysis of A. bisporus Aqueous Extract
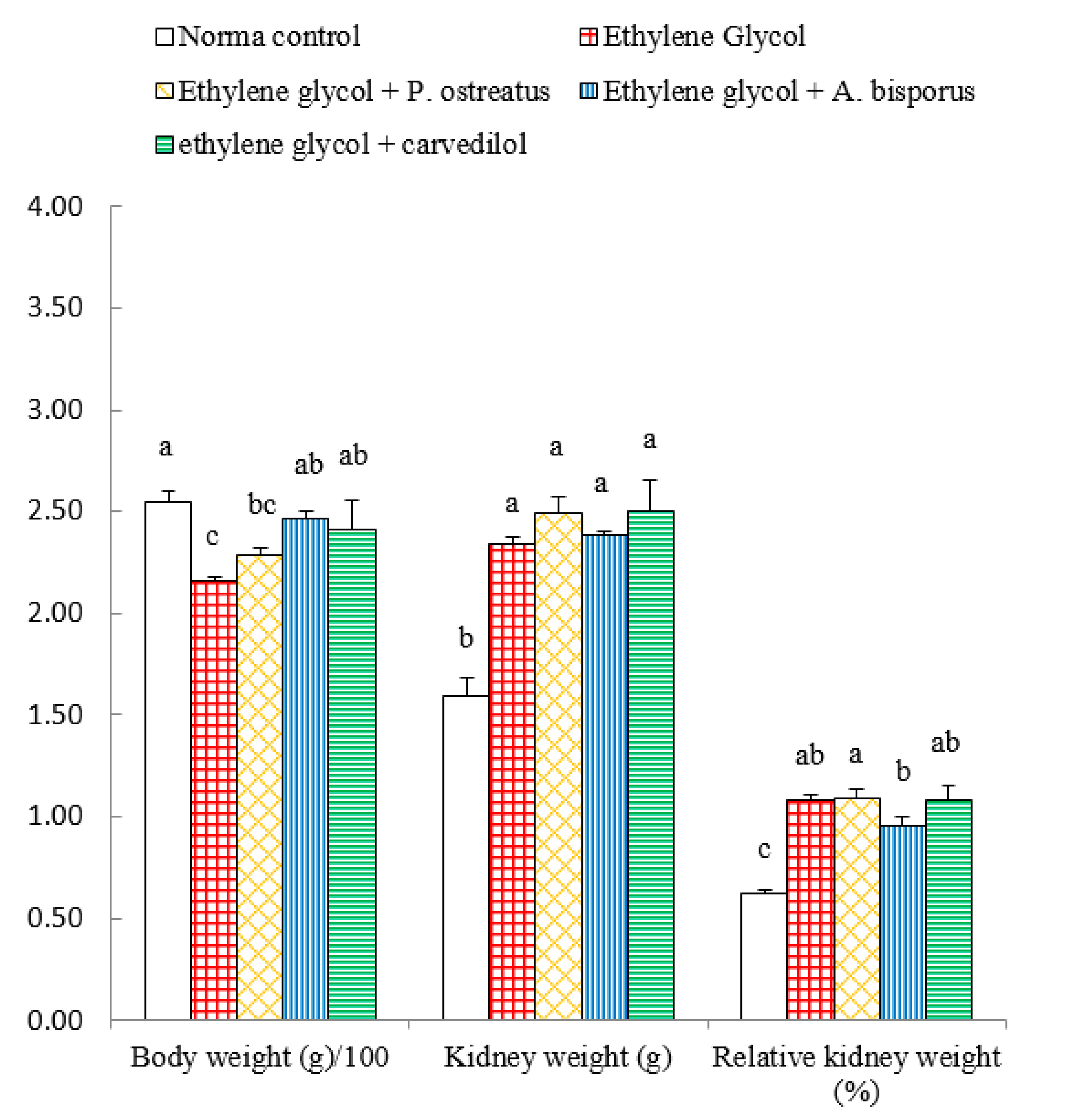
3.4. Effect on Body Weight, Kidney Weight and Relative Kidney Weight
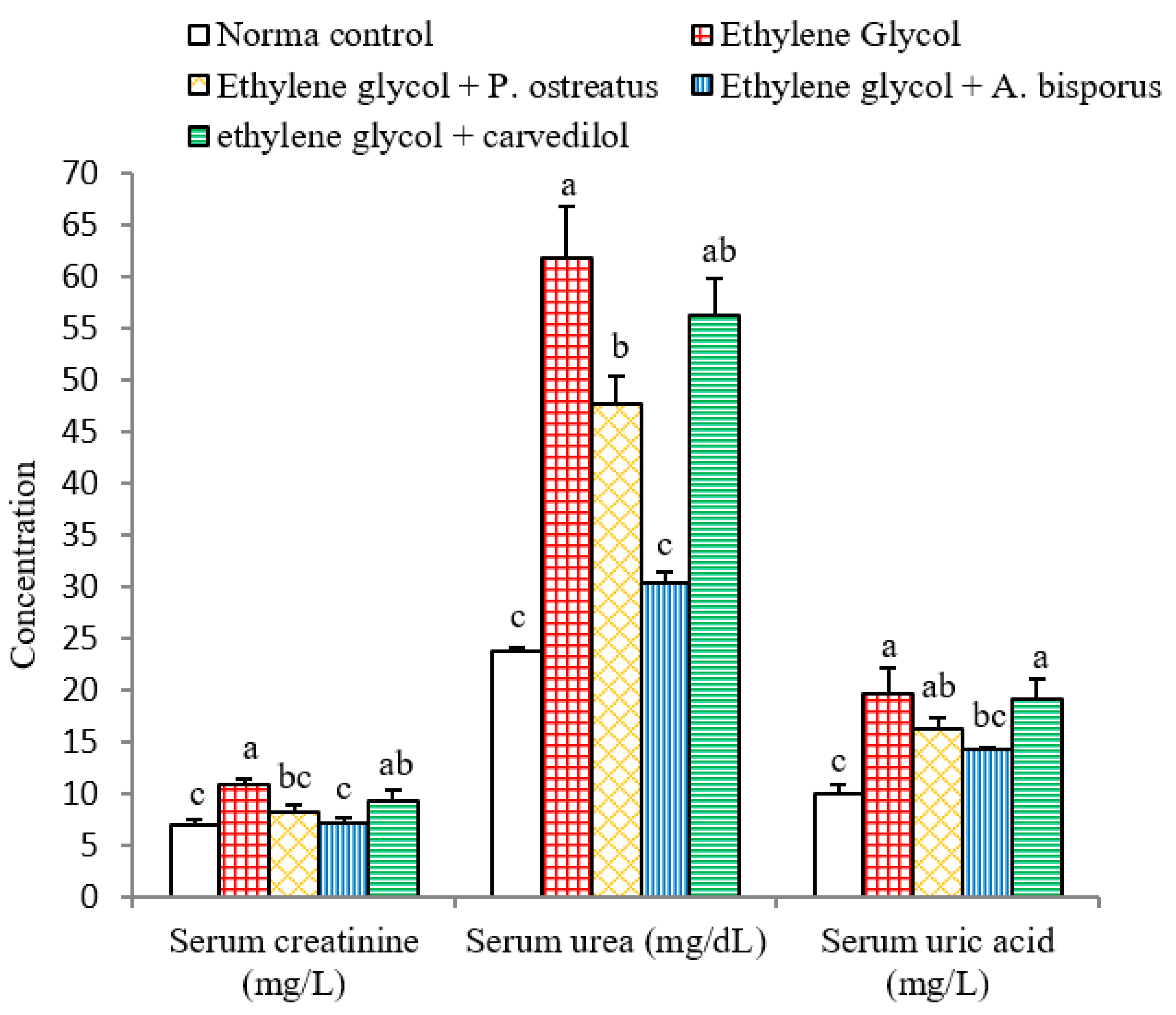
3.5. Effect on Serum Creatinine, Urea and Uric Acid Levels
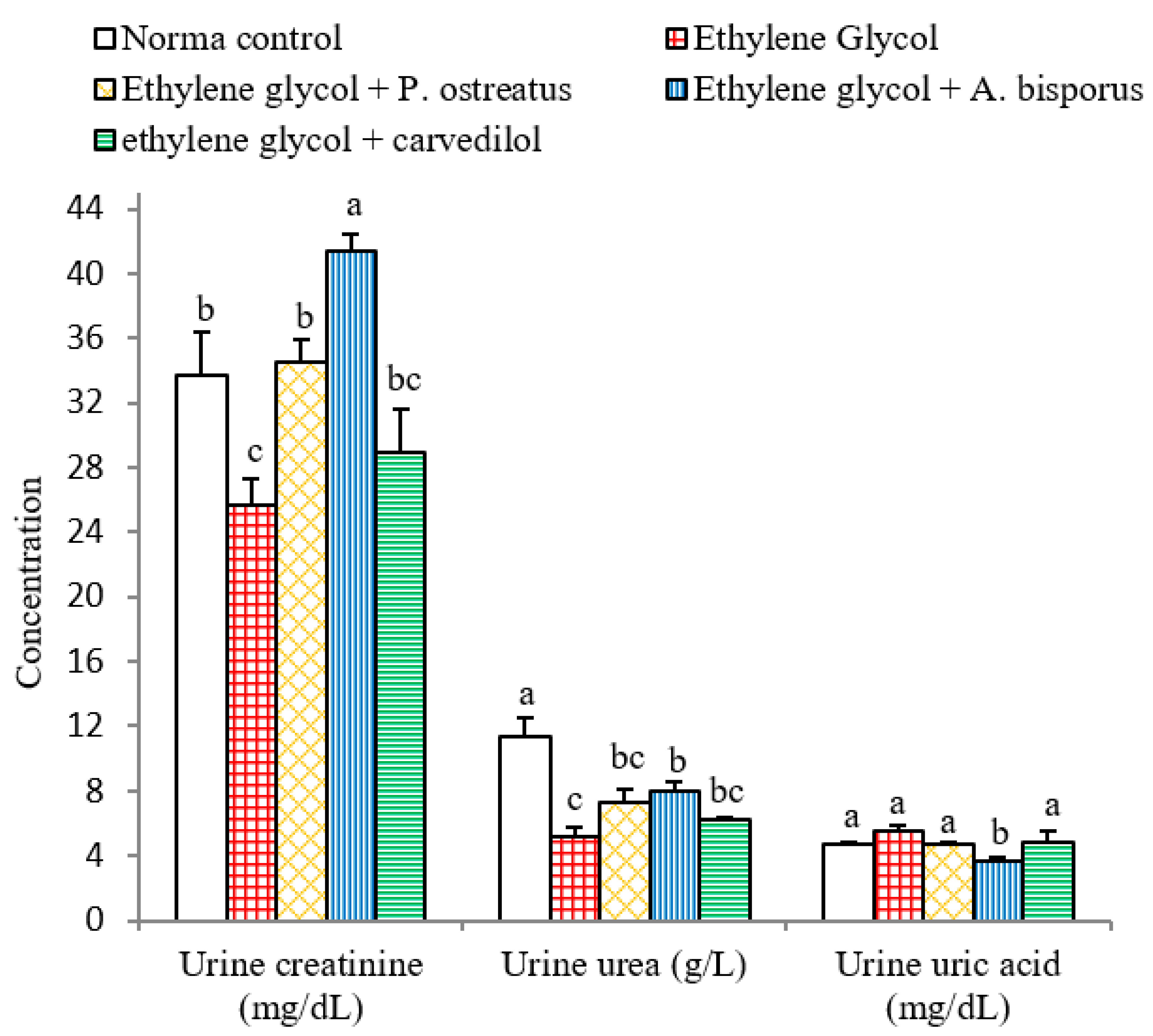
3.6. Effect on Urine Creatinine, Urea and Uric Acid Levels
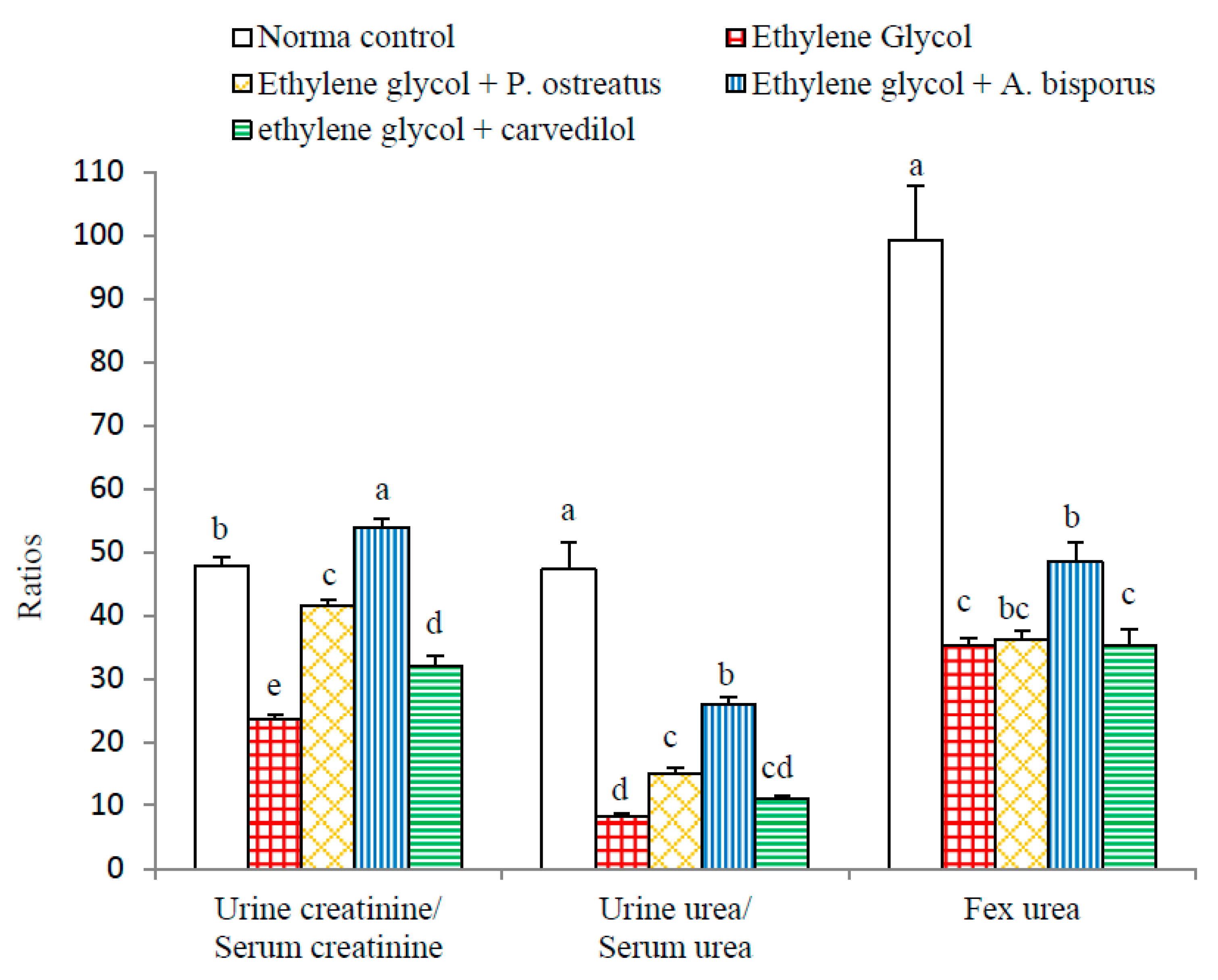
3.7. Effect on Various Ratios Related to Kidney Functions:
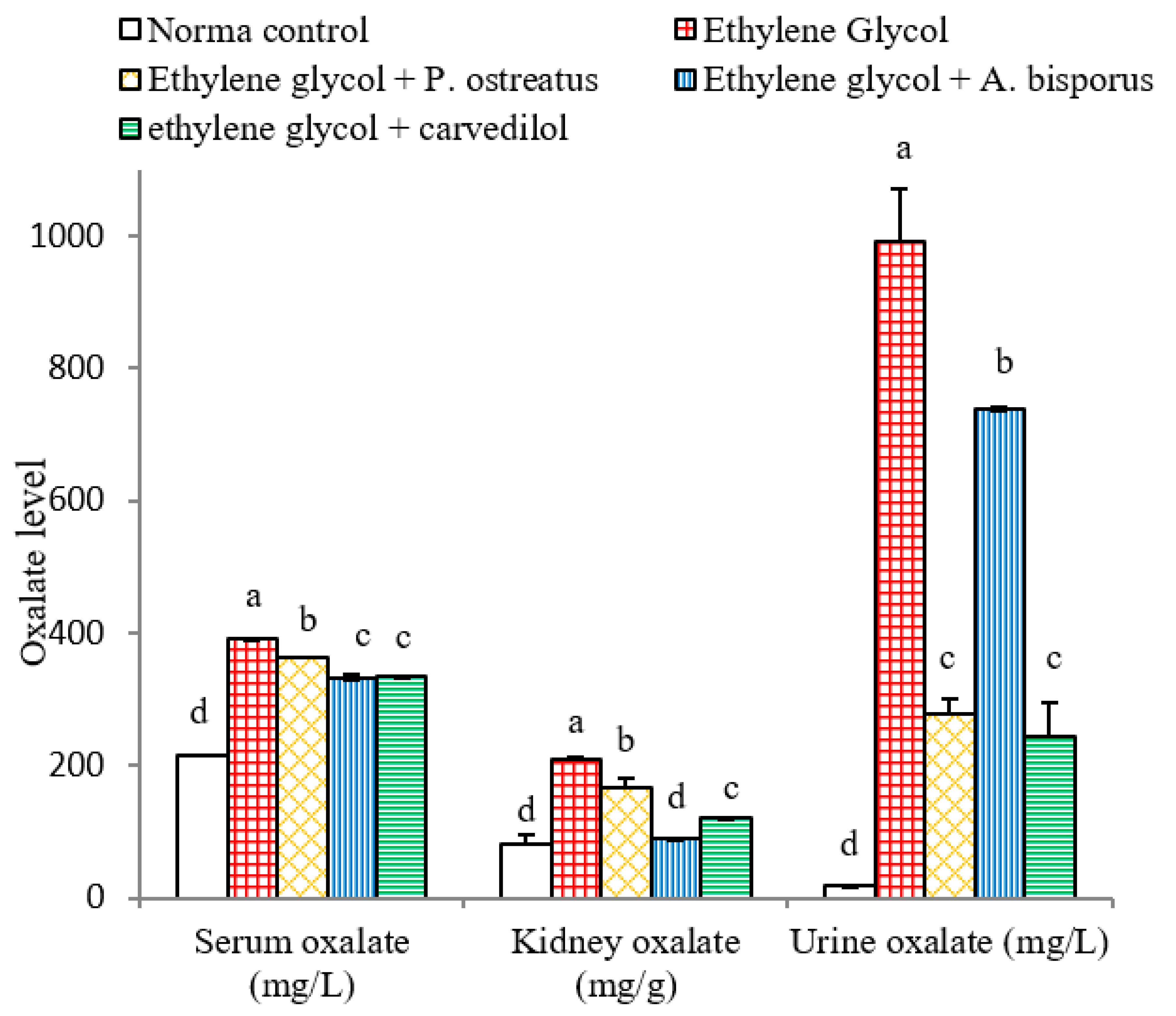
3.8. Effect on Serum, Urine and kidney Oxalate Levels
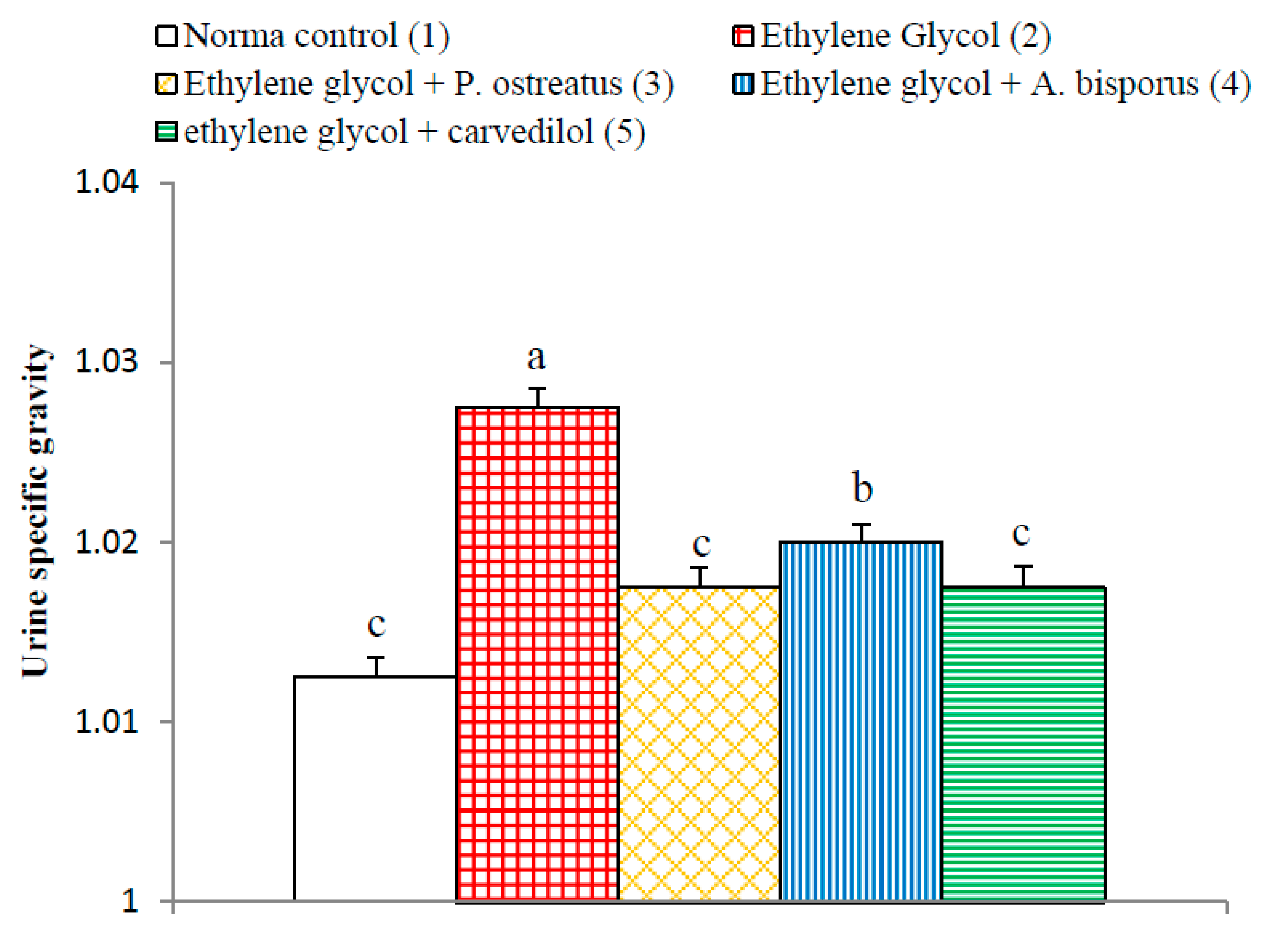
3.9. Effect on Urine Specific Gravity
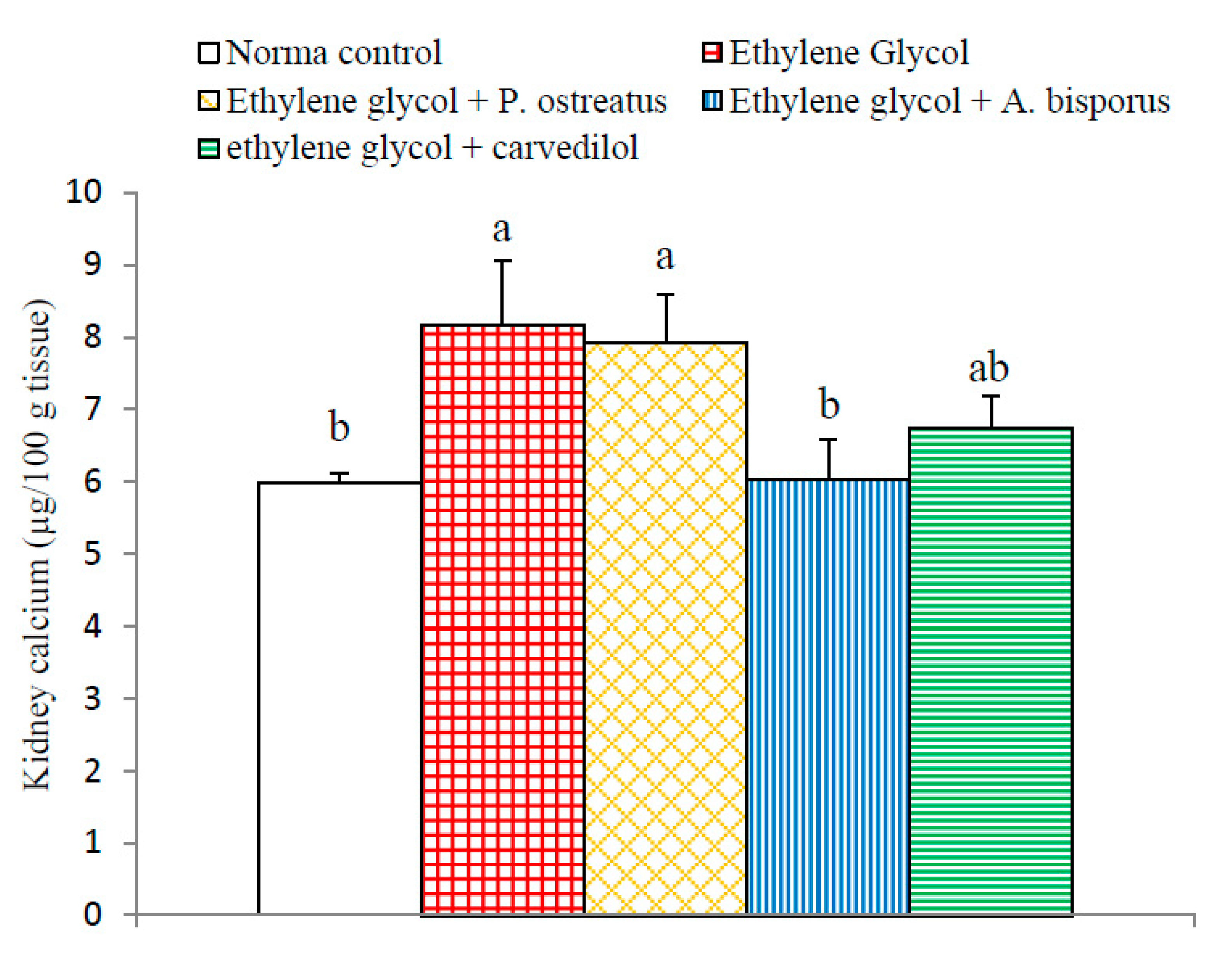
3.10. Effect on Kidney Calcium Level
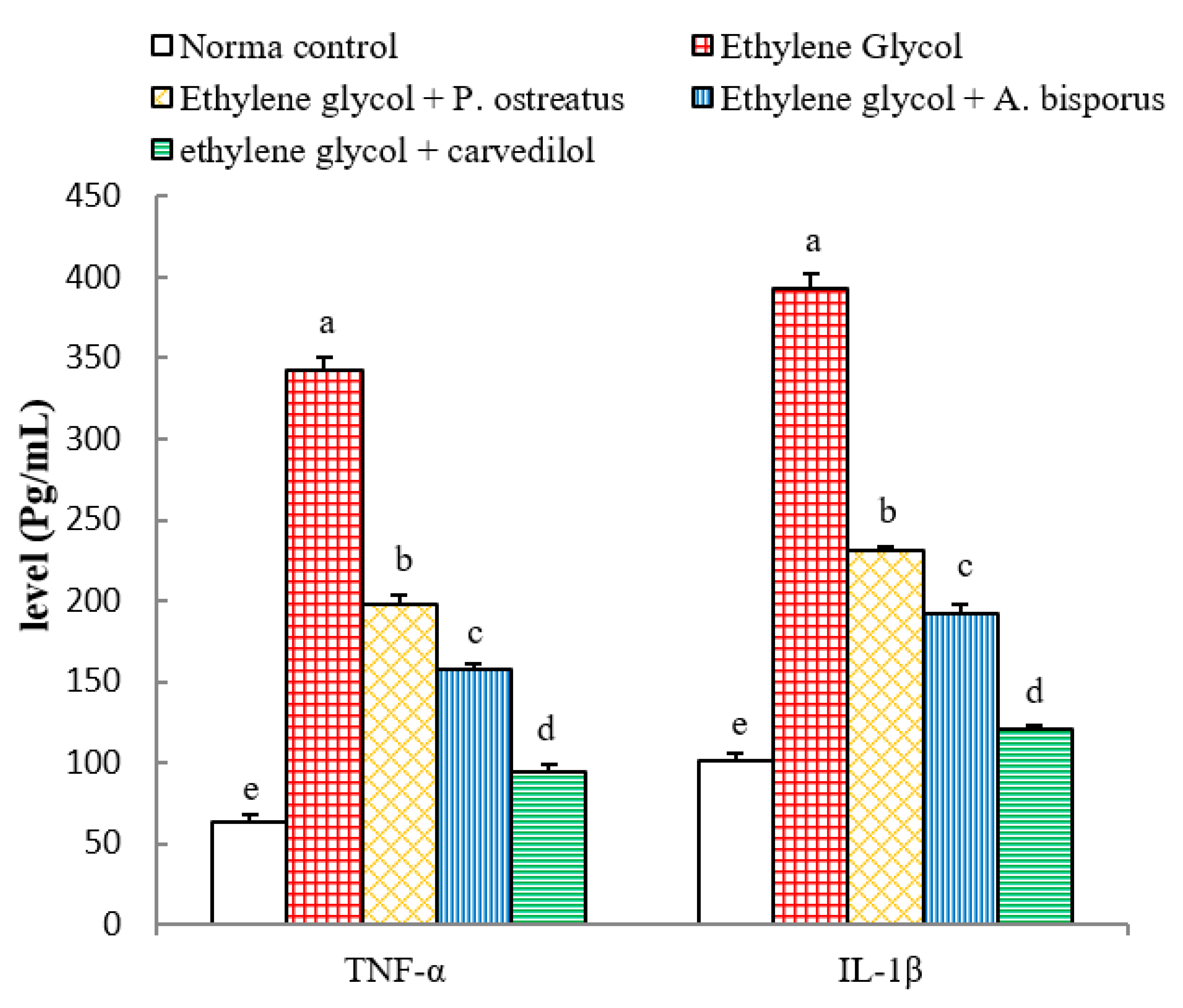
3.11. Effects on Serum TNF-α and IL-1β Levels
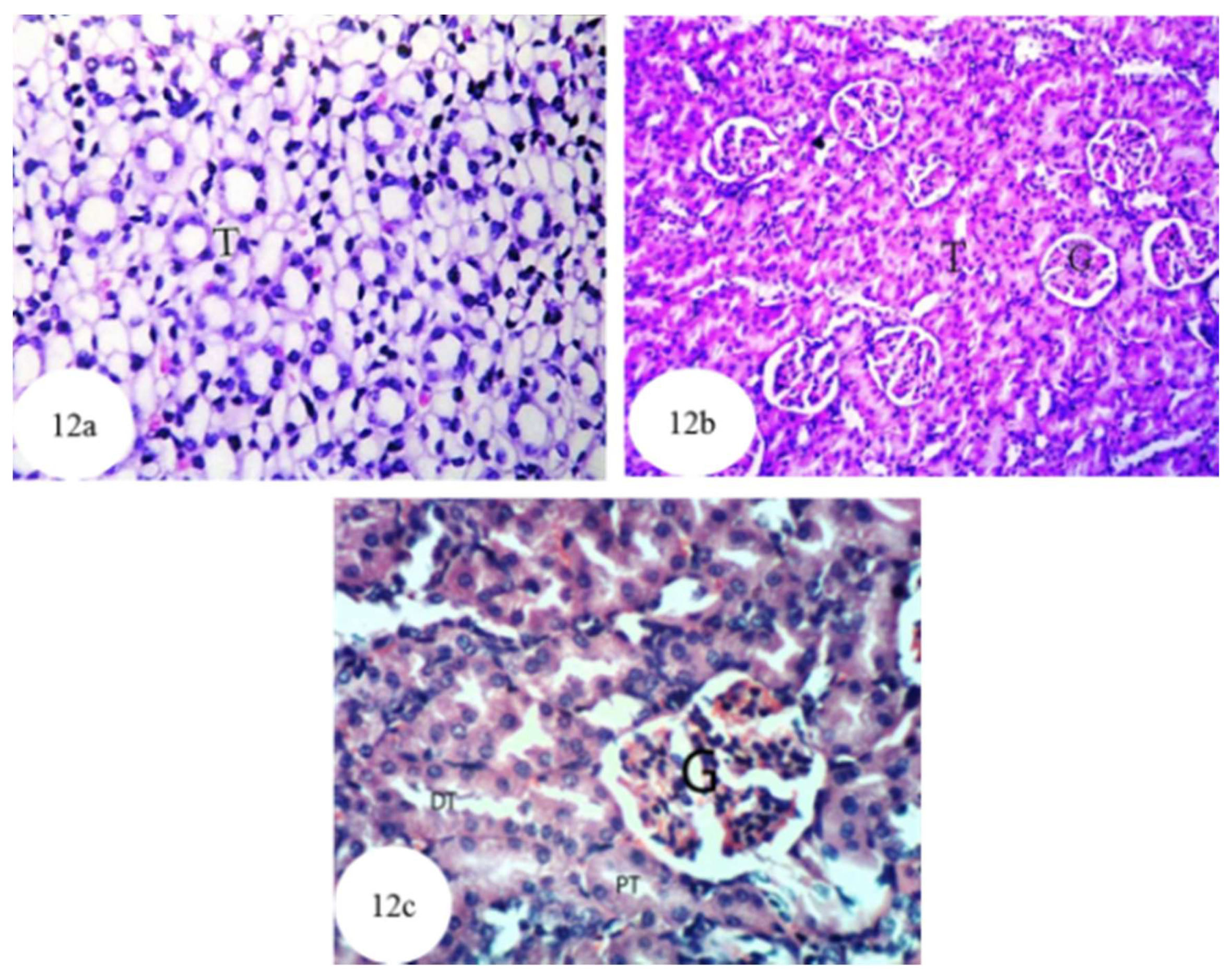
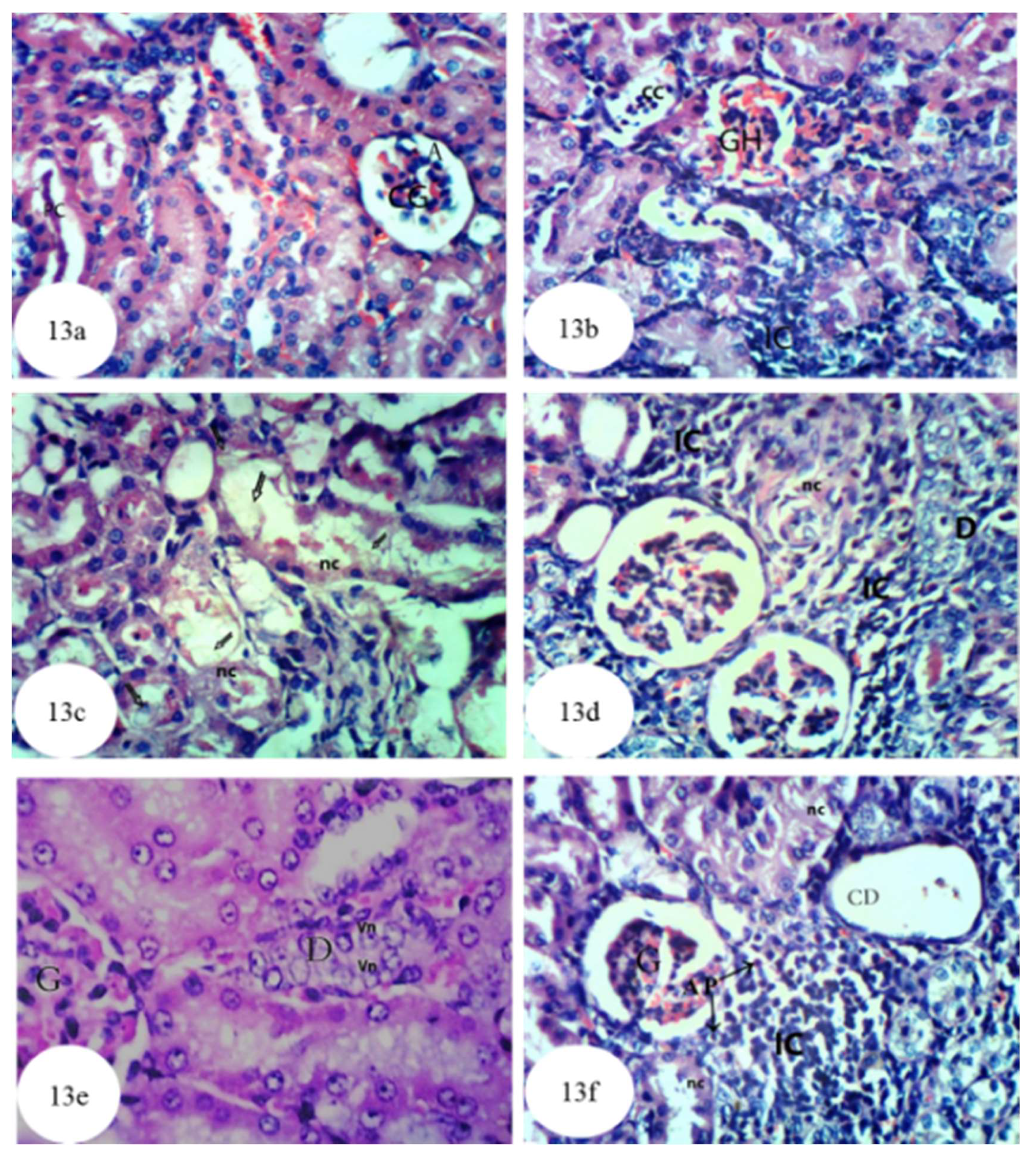
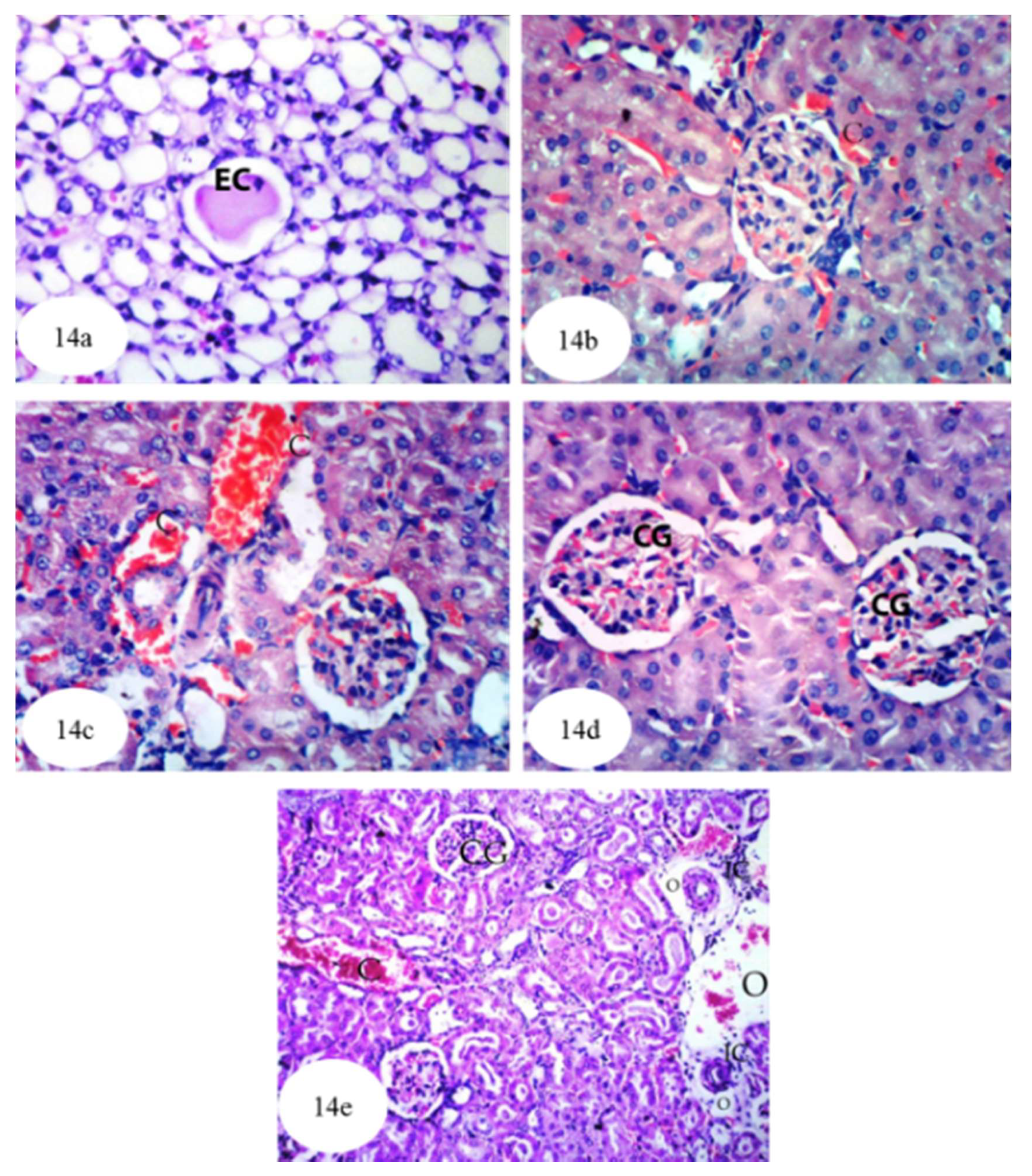
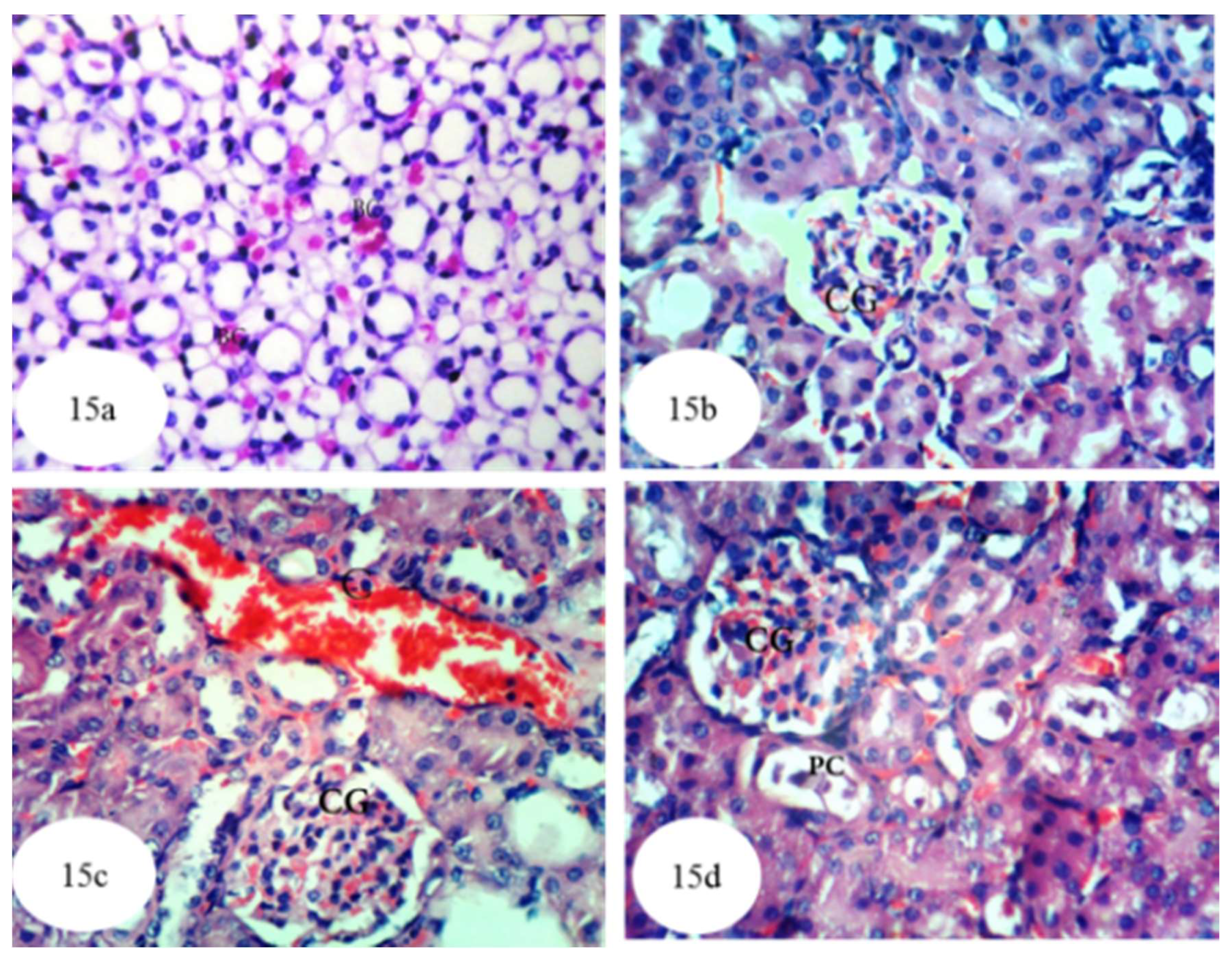
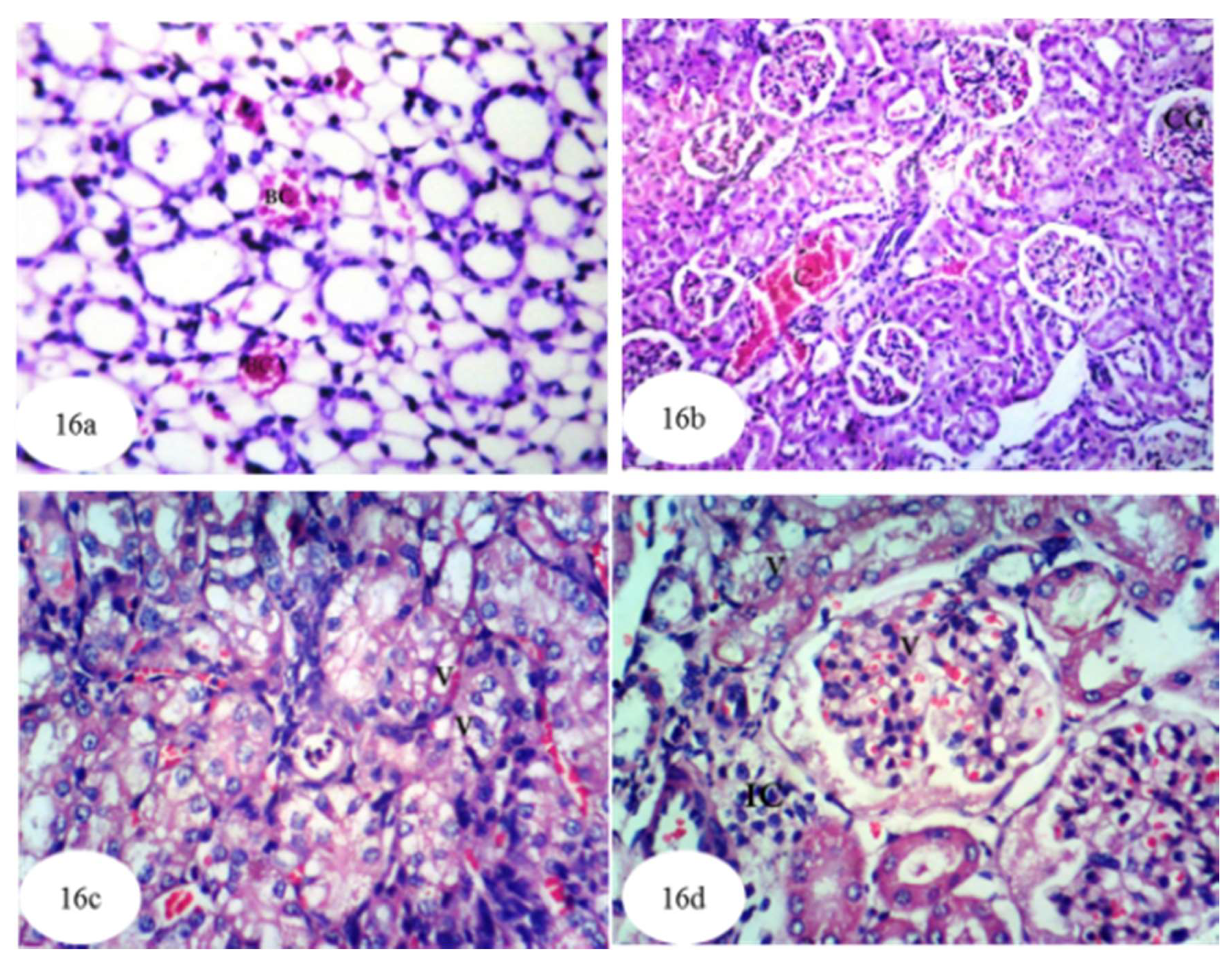
3.12. Histological Effects
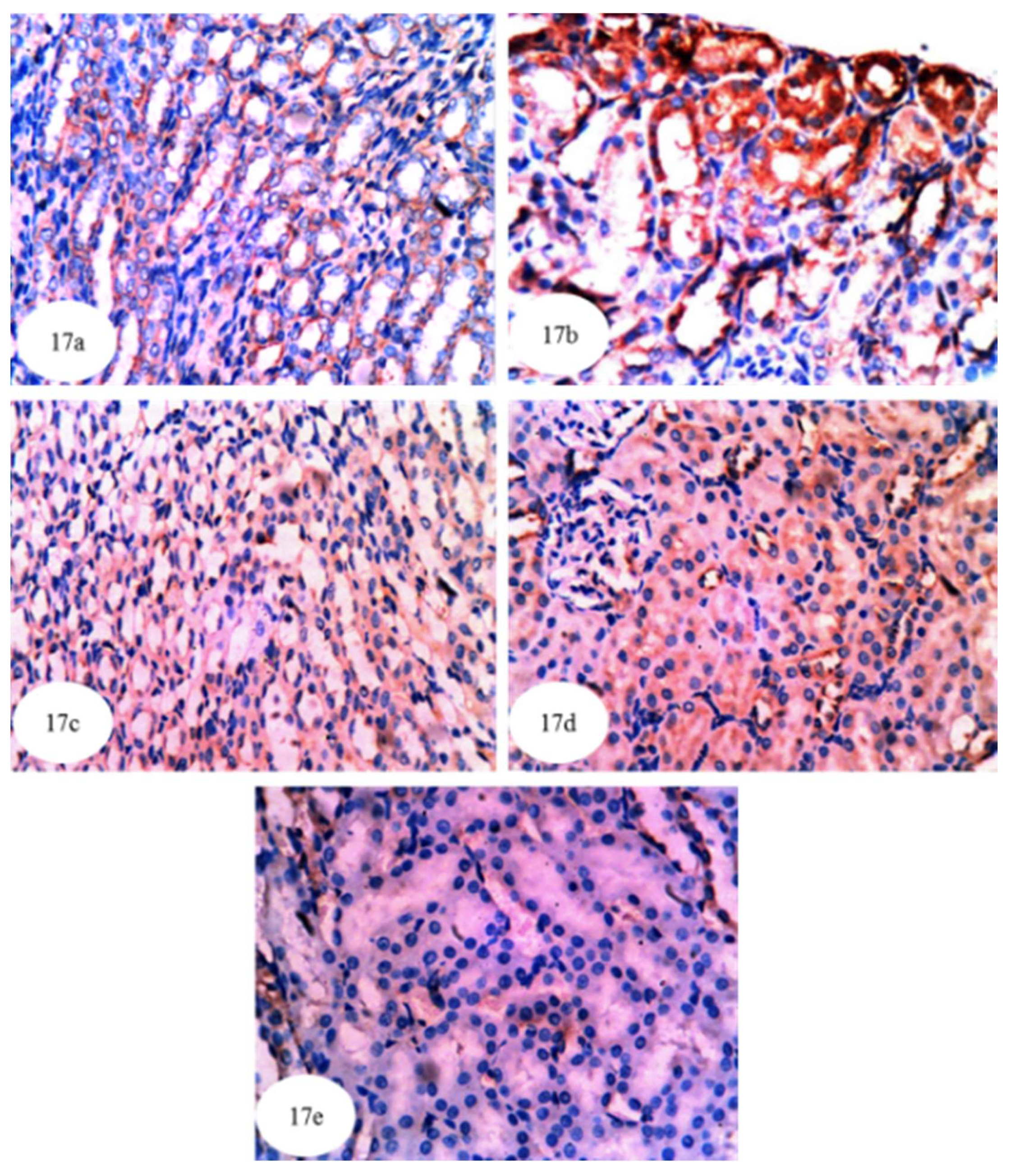
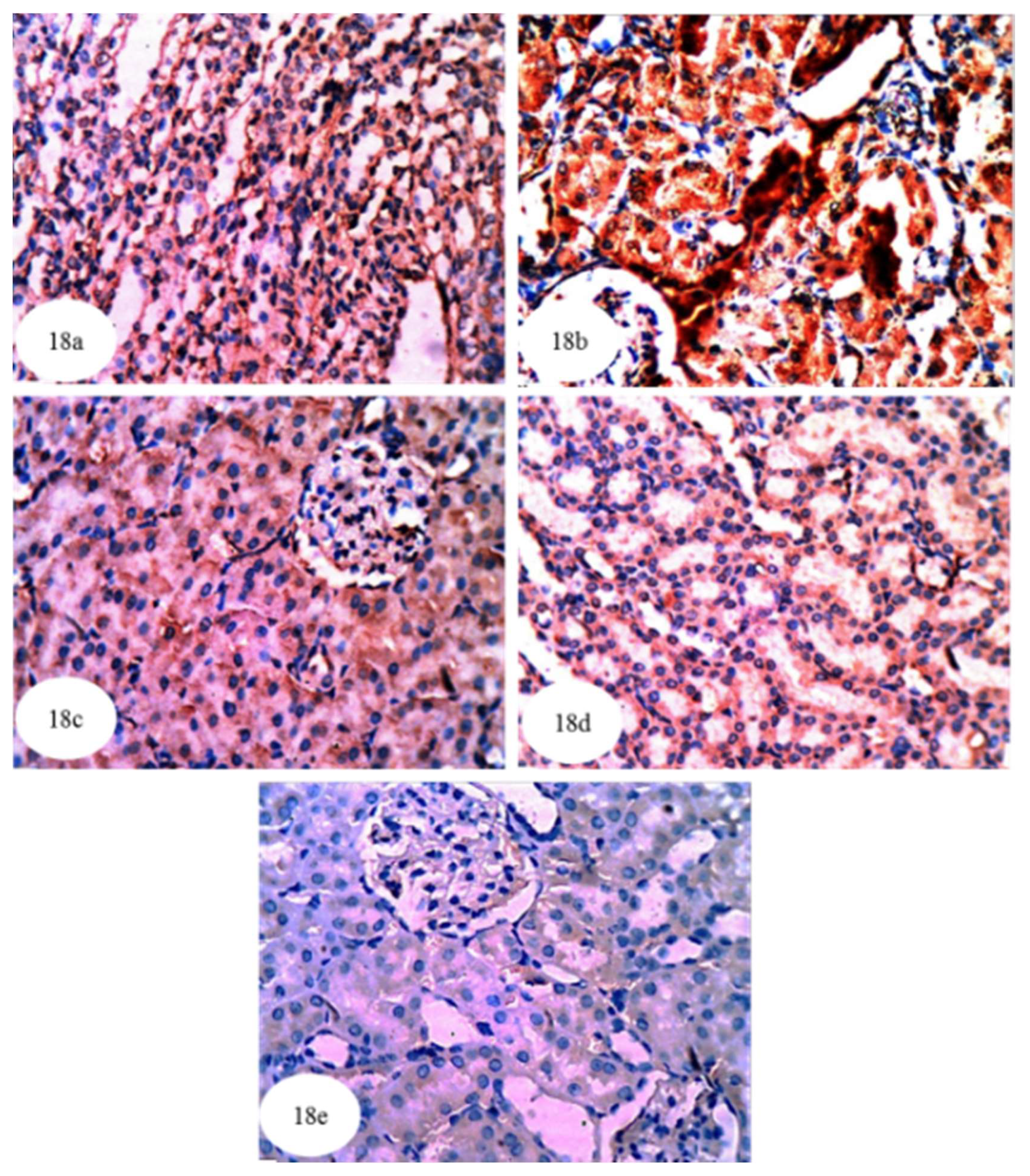
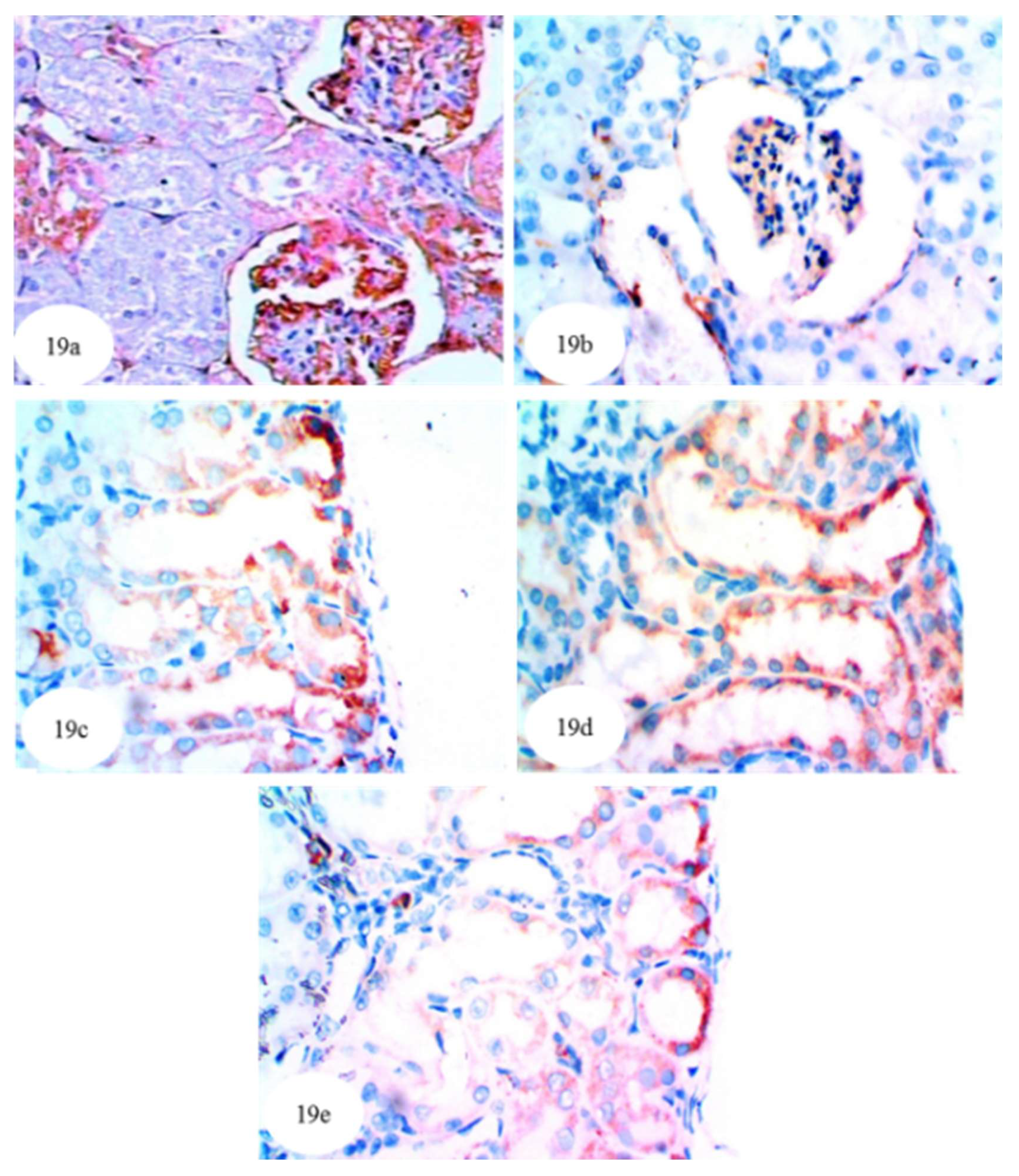
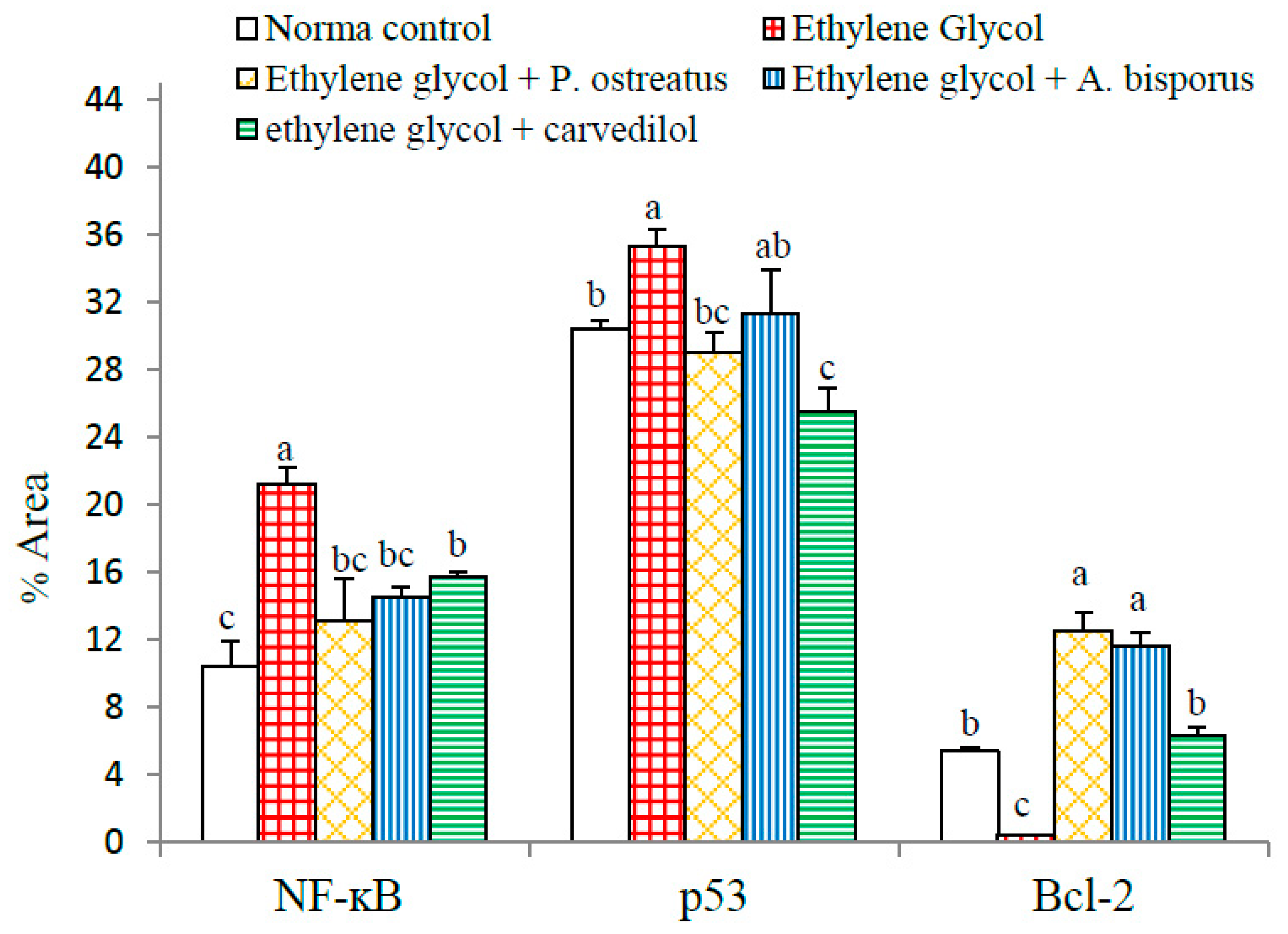
3.13. Effect on Immunohistochemically Detected Kidney NF-κB, p53 and Bcl-2
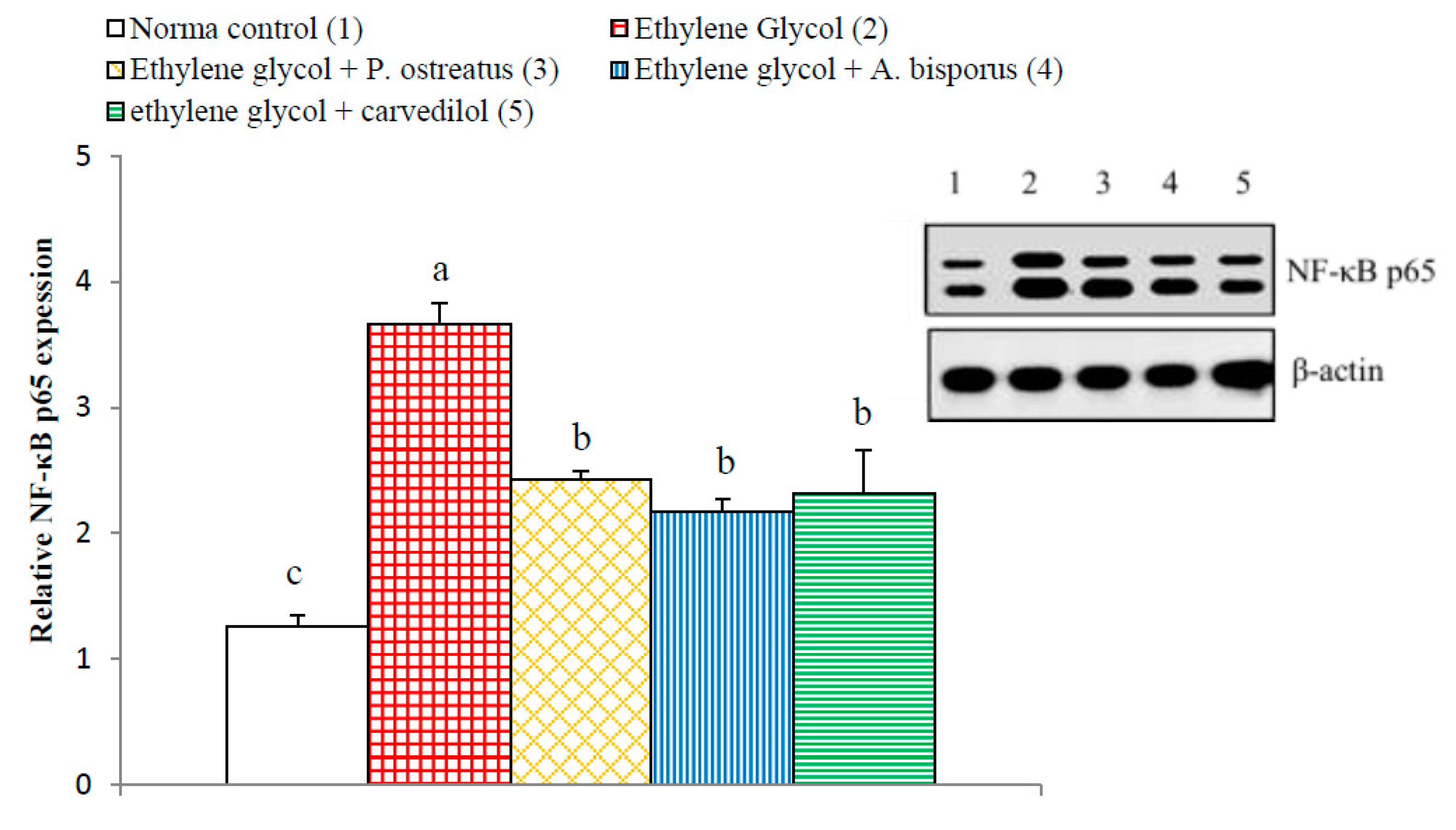
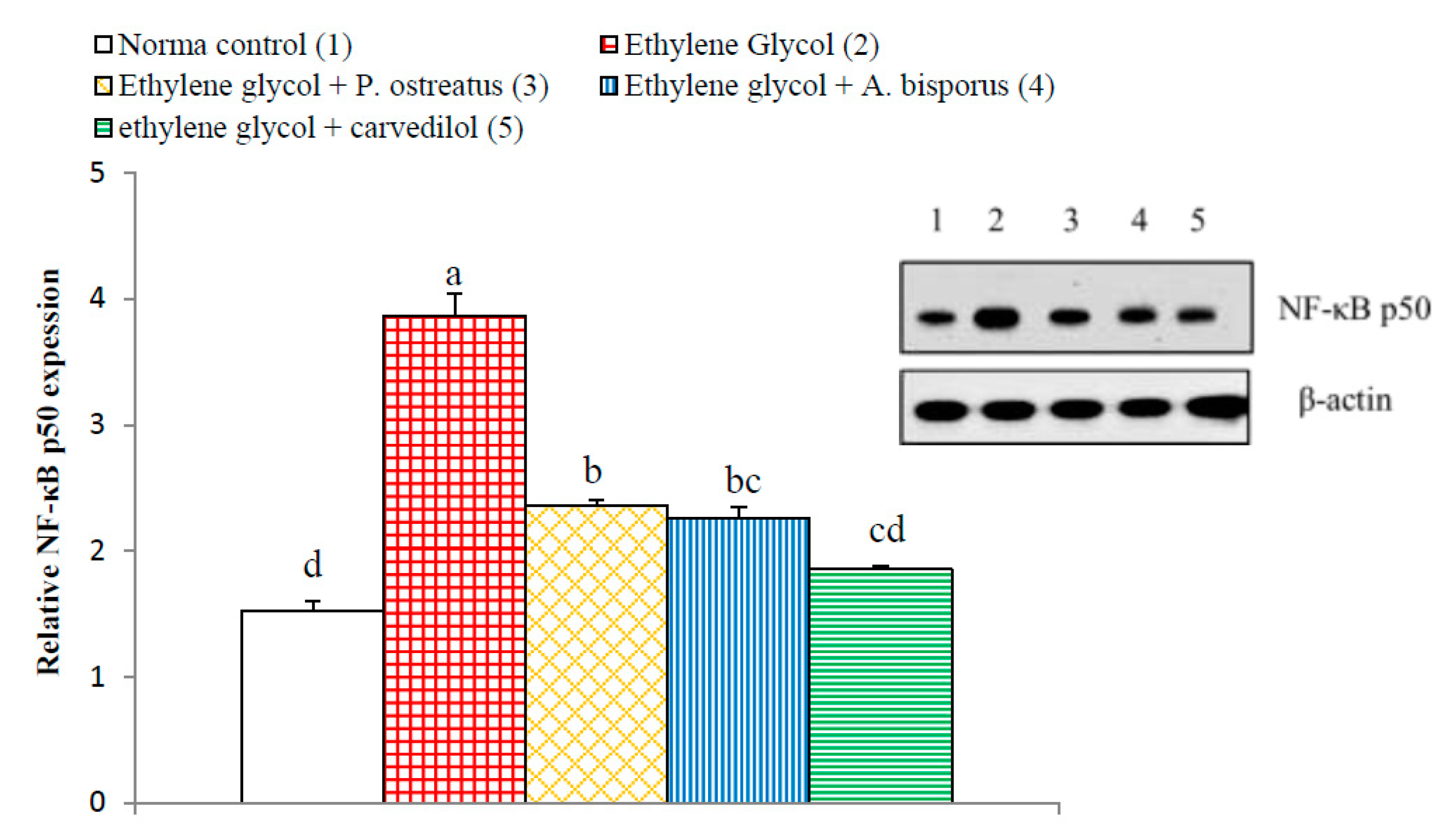
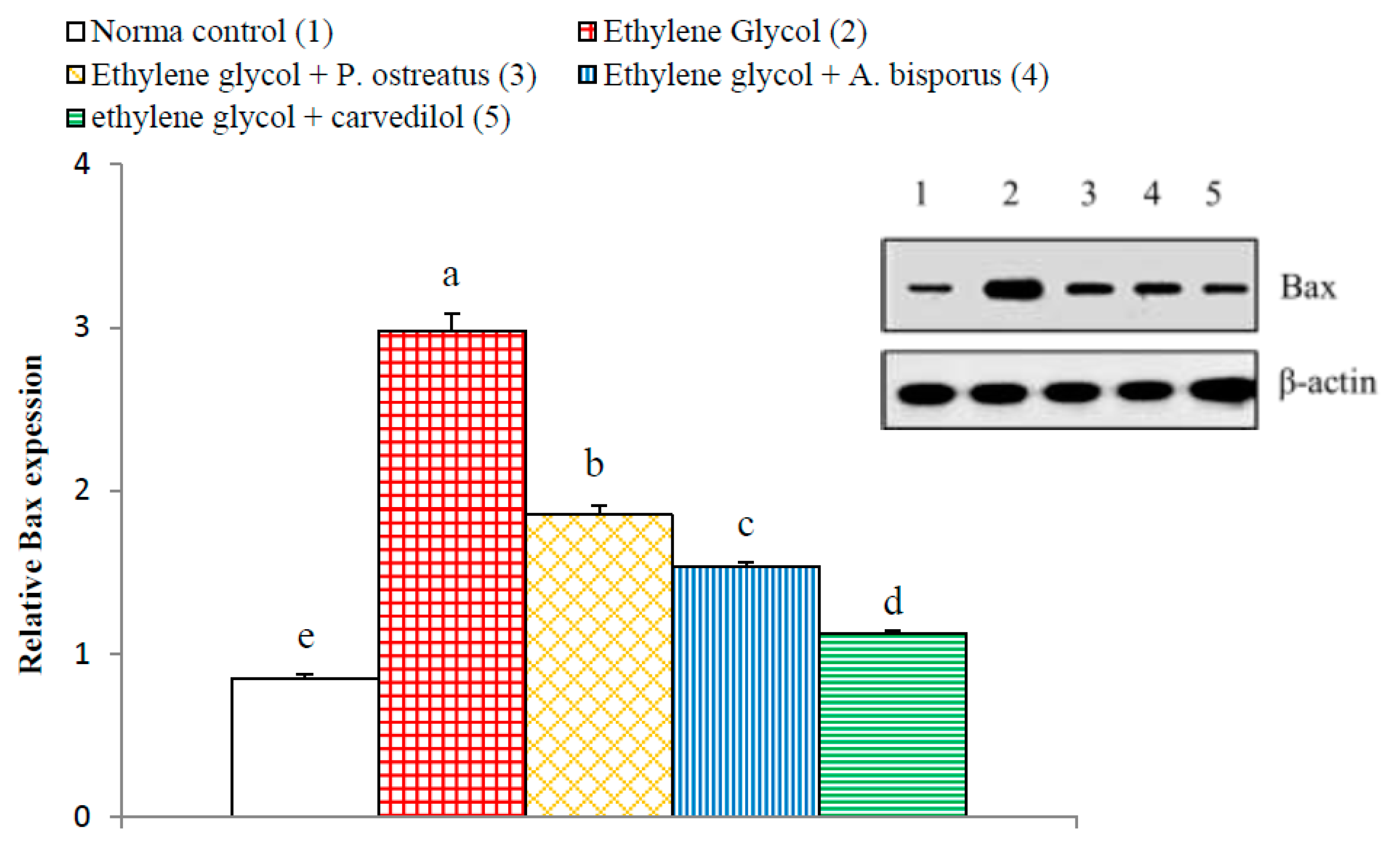
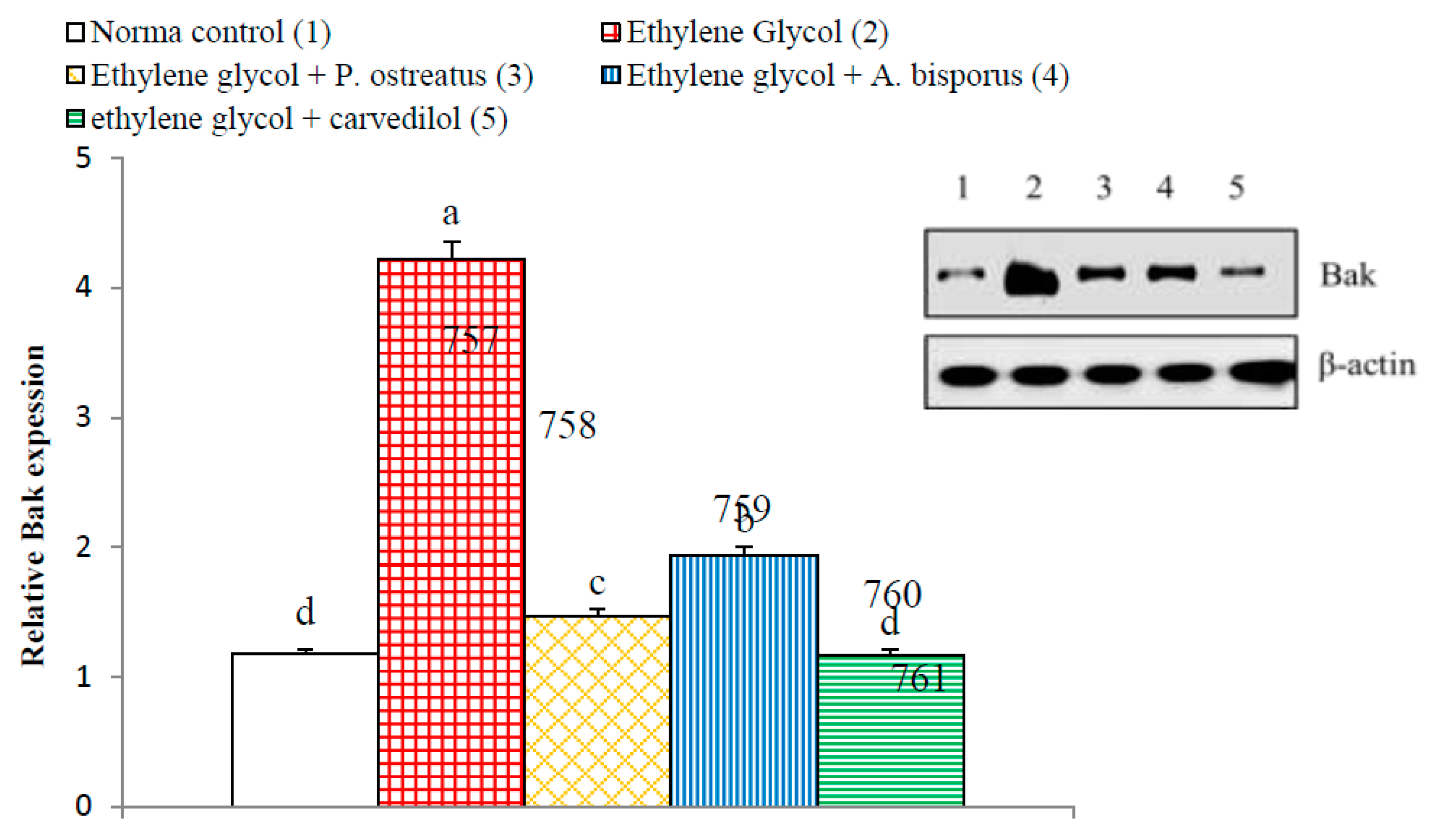
3.14. Effect on Kidney p65, p50, Bax and Bak Detected by Western Blot
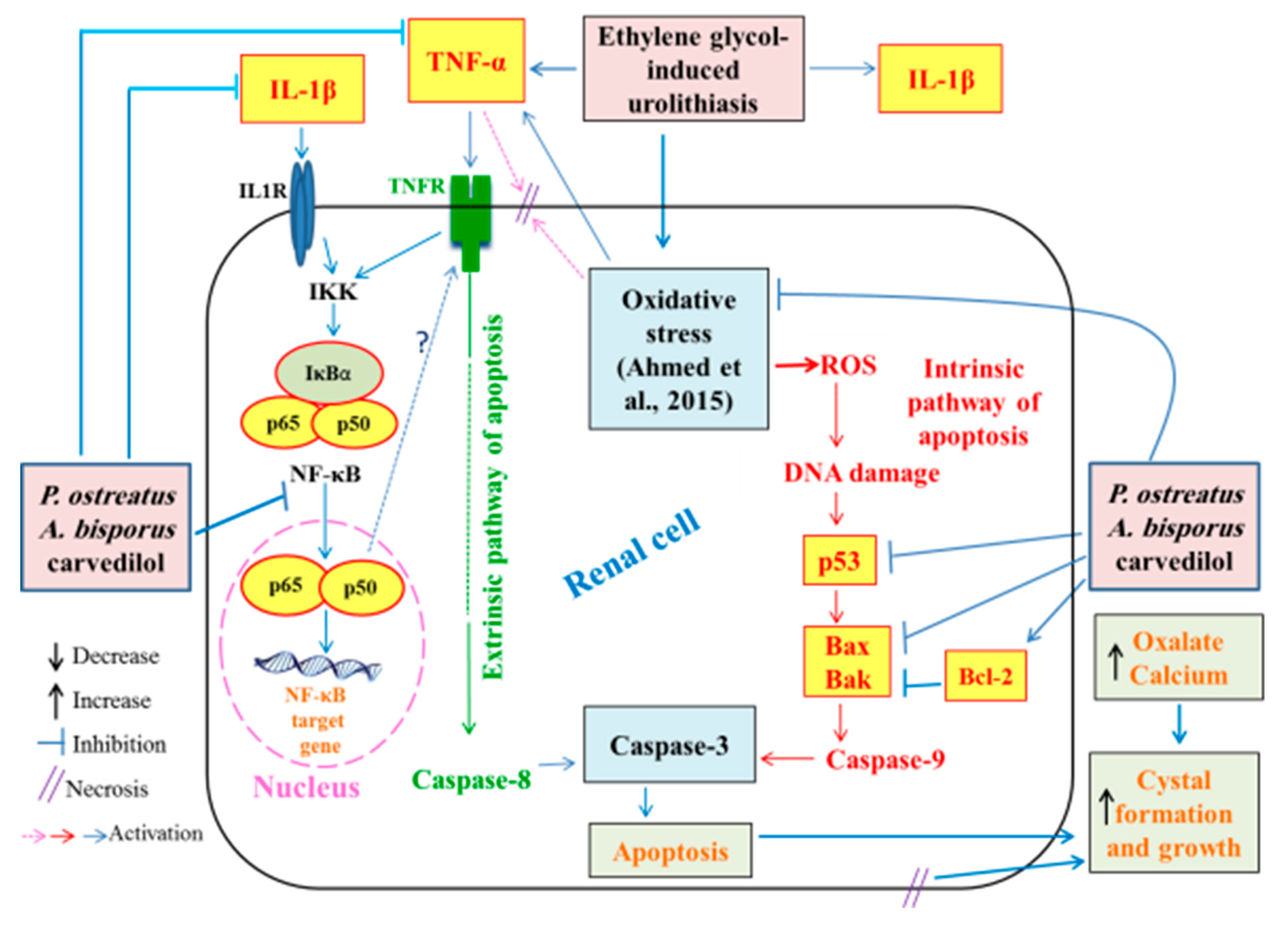
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ilhan, M.; Ergene, B.; Süntar, I.; Özbilgin, S.; Çitoğlu, G.S.; Demirel, M.A.; Keleş, H.; Altun, L.; Akkol, E.K. Preclinical evaluation of antiurolithiatic activity of Viburnum opulus L. on sodium oxalate-induced urolithiasis rat model. Evid. Based Complement Altern. Med. 2014, 2014, 578103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rad, A.K.; Hajzadeh, M.; Rajaei, Z.; Sadeghian, M.; Hashemi, N.; Keshavarzi, Z. Preventive effect of Cynodon dactylon against ethylene glycol-induced nephrolithiasis in male rats. Avicenna J. Phytomed. 2011, 1, 14–23. [Google Scholar]
- Laikangbam, R.; Damayanti Devi, M. Inhibition of calcium oxalate crystal deposition on kidneys of urolithiatic rats by Hibiscus sabdariffa L. extract. Urol. Res. 2012, 40, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Atmani, F.; Slimani, Y.; Mimouni, M.; Aziz, M.; Hacht, B.; Ziyyat, A. Effect of aqueous extract from Herniaria hirsuta L. on experimentally nephrolithiasic rats. J. Ethnopharmacol. 2004, 95, 87–93. [Google Scholar] [CrossRef]
- Alelign, T.; Petros, B. Kidney stone disease: An update on current concepts. Adv. Urol. 2018, 2018, 3068365. [Google Scholar] [CrossRef]
- Peck, A.B.; Canales, B.K.; Nguyen, C.Q. Oxalate-degrading microorganisms or oxalate-degrading enzymes: Which is the future therapy for enzymatic dissolution of calcium-oxalate uroliths in recurrent stone disease? Urolithiasis 2016, 44, 45–50. [Google Scholar] [CrossRef]
- Danpure, C.J. Primary hyperoxaluria. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Childs, B., Kinzler, K.W., Vogelstein, B., Eds.; McGraw-Hill: New York, NY, USA, 2001; Volume 2, pp. 3323–3367. [Google Scholar]
- Hoppe, B. An update on primary hyperoxaluria. Nat. Rev. Nephrol. 2012, 8, 467–475. [Google Scholar] [CrossRef]
- Massey, L.K.; Roman-Smith, H.; Sutton, R.A. Effect of dietary oxalate and calcium on urinary oxalate and risk of formation of calcium oxalate kidney stones. J. Am. Diet Assoc. 1993, 93, 901–906. [Google Scholar] [CrossRef]
- Holmes, R.P.; Goodman, H.O.; Assimos, D.G. Contribution of dietary oxalate to urinary oxalate excretion. Kidney Int. 2001, 59, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.R.; Glenton, P.A.; Byer, K.J. Dietary oxalate and calcium oxalate nephrolithiasis. J. Urol. 2007, 178, 2191–2196. [Google Scholar] [CrossRef]
- Dahiya, T.; Pundir, C.S. In vivo oxalate degradation by liposome encapsulated oxalate oxidase in rat model of hyperoxaluria. Indian J. Med. Res. 2013, 137, 136–1341. [Google Scholar] [PubMed]
- Kifayatullah, M.; Rahim, H.; Ullah Jan, N.; Abbas, S.; Khan, M.S.; Ikram, M. Anti-urolithiatic effect of Pericampylus glaucus against ethylene glycol induced urolithiasis in male Sprague Dawley rats. Sains Malays. 2019, 48, 1075–1081. [Google Scholar] [CrossRef]
- Tiselius, H.G. Epidemiology and medical management of stone disease. Br. J. Urol. Int. 2003, 91, 758–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.R. Reactive oxygen species, inflammation and calcium oxalate nephrolithiasis. Transl. Androl. Urol. 2014, 3, 256–276. [Google Scholar]
- Khan, S.R.; Hackett, R.L. Retention of calcium oxalate crystals in renal tubules. Scanning Microsc. 1991, 5, 707–712. [Google Scholar]
- Khan, S.R. Calcium oxalate crystal interaction with renal tubular epithelium, mechanism of crystal adhesion and its impact on stone development. Urol. Res. 1995, 23, 71–79. [Google Scholar] [CrossRef]
- Dal Moro, F.; Mancini, M.; Tavolini, I.M.; De Marco, V.; Bassi, P. Cellular and molecular gateways to urolithiasis: A new insight. Urol. Int. 2005, 74, 193–197. [Google Scholar] [CrossRef]
- Williams, H.E.; Wandzilak, T.R. Oxalate synthesis, transport and hyperoxaluric syndromes. J. Urol. 1989, 141, 742–749. [Google Scholar] [CrossRef]
- Kuhad, A.; Chopra, K. Attenuation of diabetic nephropathy by tocotrienol; involvement of NF-κB signaling pathway. Life Sci. 2009, 84, 296–301. [Google Scholar] [CrossRef]
- Grande, M.T.; Pérez-Barriocanal, F.; López-Novoa, J.M. Role of inflammation in túbulo-interstitial damage associated to obstructive nephropathy. J. Inflamm. 2010, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Horuz, R.; Göktaş, C.; Çetinel, C.A.; Akça, O.; Aydın, H.; Ekici, I.D.; Albayrak, S.; Sarıca, K. Role of TNF-associated cytokines in renal tubular cell apoptosis induced by hyperoxaluria. Urolithiasis 2013, 41, 197–203. [Google Scholar] [CrossRef]
- Vousden, K.H.; Lane, D.P. P53 in health and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 275–283. [Google Scholar] [CrossRef]
- Jang, J.H.; Surh, Y.J. Potentiation of cellular antioxidant capacity by Bcl-2: Implications for its antiapoptotic function. Biochem. Pharmacol. 2003, 66, 1371–1379. [Google Scholar] [CrossRef]
- Shafi, H.; Moazzami, B.; Pourghasem, M.; Kasaeian, A. An overview of treatment options for urinary stones. Casp. J. Intern. Med. 2016, 7, 1–6. [Google Scholar]
- Bashir, S.; Gilani, A.H. Antiurolithiatic effect Bergenia ligulata rhizome: An explanation of the underlying mechanisms. J. Ethnopharmacol. 2009, 122, 106–116. [Google Scholar] [CrossRef]
- Divakar, K.; Pawar, A.T.; Chandrasekhar, S.B.; Dighe, S.B.; Divakar, G. Protective effect of the hydro-alcoholic extract of Rubia cordifolia roots against ethylene glycol induced urolithiasis in rats. Food Chem. Toxicol. 2010, 48, 1013–1018. [Google Scholar] [CrossRef]
- Ahmed, O.; Ahmed, M.; Ragab, M. Antiurolithic and antioxidant influence of Pleurotus ostreatus and Agaricus bisporus aqueous extracts and carvedilol in male rats. Am. J. Biochem. 2015, 5, 99–112. [Google Scholar]
- Nirumand, M.C.; Hajialyani, M.; Rahimi, R.; Farzaei, M.H.; Zingue, S. Dietary Plants for the Prevention and Management of Kidney Stones: Preclinical and Clinical Evidence and Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 765. [Google Scholar] [CrossRef] [Green Version]
- Lindequist, U.; Niedermeyer, T.H.J.; Jülich, W.D. The pharmacological potential of mushrooms. Evid. Based Complement Altern. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Wang, X. Selection of a culture medium for reducing costs and intracellular polysaccharide production by Agaricus blazei AB2003. Food Technol. Biotechnol. 2005, 47, 210–214. [Google Scholar]
- Gordon, M.; Bihari, B.; Goosby, E.; Gorter, R.; Greco, M.; Guralnik, M.; Mimura, T.; Rudinicki, V.; Wong, R.; Kaneko, Y. A placebo controlled trial of the immune modulator, lentinan, in HIV-positive patients: A phase I/II trial. J. Med. 1998, 29, 305–330. [Google Scholar]
- Wasser, S.P.; Weis, A.L. Therapeutic effects of substances occurring in higher Basidiomycetes mushrooms: A modern perspective. Crit. Rev. Immunol. 1999, 19, 65–96. [Google Scholar]
- Lakhanpal, T.N.; Rana, M. Medicinal and nutraceutical genetic resources of mushrooms. Plant Gene. Resour. Charact. Util. 2005, 3, 288–303. [Google Scholar] [CrossRef]
- Katya, K.; Yun, Y.H.; Park, G.; Lee, J.Y.; Yoo, G.; Bai, S.C. Evaluation of the efficacy of fermented by-product of mushroom, Pleurotus ostreatus, as a fish meal replacer in juvenile amur catfish, Silurus asotus: Effects on growth, serological characteristics and immune responses. Asian Australas. J. Anim. Sci. 2014, 27, 1478–1486. [Google Scholar] [CrossRef]
- Singh, V.; Vyas, D.; Pandey, R.; Sheikh, I.M. Pleurotus ostreatus produces antioxidant and antiarthritic activity in Wistar albino rats. World J. Pharm. Pharma. Sci. 2015, 4, 1230–1246. [Google Scholar]
- Chen, S.; Oh, S.R.; Phung, S.; Hur, G.; Ye, J.J.; Kwok, S.L.; Shrode, G.E.; Belury, M.; Adams, L.S.; Williams, D. Anti-aromatase activity of phytochemicals in white button mushrooms (Agaricus bisporus). Cancer Res. 2006, 66, 12026–12034. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.C.; Jeong, Y.T.; Yang, B.K.; Islam, R.; Koyyalamudi, S.R.; Pang, G.; Cho, K.Y.; Song, C.H. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetic and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Niksic, M.; Jakovljevic, D.; Helsper, J.P.F.G.; van Griensven, L.J.L.D. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phellinus linteus. Food Chem. 2011, 129, 1667–1675. [Google Scholar] [CrossRef]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martinez, J.A.; Garcia-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated RAW 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Feuerstein, G.Z.; Ruffolo, R.R., Jr. Carvedilol, a novel vasodilating beta blocker with the potential for cardiovascular organ protection. Eur. Heart J. 1996, 17 (Suppl. B), 24–29. [Google Scholar] [CrossRef] [Green Version]
- de Araújo Júnior, R.F.; Souza, T.O.; de Medeiros, C.A.; de Souza, L.B.; de Freitas, M.L.; de Lucena, H.F.; do Socorro Costa Feitosa Alves, M.; de Araújo, A.A. Carvedilol decrease IL-1β and TNF-ά, inhibits MMP-2, MMP-9, COX-2, and RANKL expression, and up-regulates OPG in a rat model of periodontitis. PLoS ONE 2013, 8, e66391. [Google Scholar] [CrossRef]
- Singh, D.; Chander, V.; Chopra, K. Carvedilol and trimetazidine attenuates ferric nitrilotriacetate-induced oxidative renal injury in rats. Toxicology 2003, 191, 143–151. [Google Scholar] [CrossRef]
- Hayashi, T.; De Velasco, M.A.; Saitou, Y.; Nose, K.; Nishioka, T.; Ishii, T.; Uemura, H. Carvedilol protects tubular epithelial cells from ischemia-reperfusion injury by inhibiting oxidative stress. Int. J. Urol. 2010, 17, 989–995. [Google Scholar] [CrossRef]
- Rodrigues, M.A.C.; da Silva Faria, M.C.; dos Santos, N.A.; Gobe, G.C.; dos Santos, A.C. Carvedilol efficiently protects kidneys without affecting the antitumor efficacy of cisplatin in mice. Chem. Biol. Interact. 2013, 206, 90–99. [Google Scholar] [CrossRef]
- Claus, F.D. Pharmacognosy, 4th ed.; Henry Kumpton: London, UK, 1961. [Google Scholar]
- El Bohi, K.M.; Hashimoto, Y.; Muzandu, K.; Ikenaka, Y.; Ibrahim, Z.S.; Kazusaka, A.; Fujita, S.; Ishizuka, M. Protective effect of Pleurotus cornucopiae mushroom extract on carbon tetrachloride-induced hepatotoxicity. Jpn. J. Vet. Res. 2009, 57, 109–118. [Google Scholar]
- Yamac, M.; Kanbak, G.; Zeytinoglu, M.; Senturk, H.; Bayramoglu, G.; Dokumacioglu, A.; van Griensven, L.J.D. Pancreas protective effect of button mushroom Agaricus bisporus (J. E. Lange) imbach (Agaricomycetidae) extract on rats with streptozotocin-induced diabetes. Int. J. Med. Mushroom 2010, 12, 379–389. [Google Scholar] [CrossRef]
- Arozal, W.; Watanabe, K.; Veeraveedu, P.T.; Ma, M.; Thandavarayan, R.A.; Sukumaran, V.; Suzuki, K.; Kodama, M.; Aizawa, Y. Protective effect of carvedilol on daunorubicin-induced cardiotoxicity and nephrotoxicity in rats. Toxicology 2010, 274, 18–26. [Google Scholar] [CrossRef]
- Ghaeni, F.A.; Amin, B.; Hariri, A.T.; Meybodi, N.T.; Hosseinzadeh, H. Antilithiatic effects of crocin on ethylene glycol-induced lithiasis in rats. Urolithiasis 2014, 42, 549–558. [Google Scholar] [CrossRef]
- Tsuji, H.; Shimizu, N.; Nozawa, M.; Umekawa, T.; Yoshimura, K.; De Velasco, M.A.; Uemura, H.; Khan, S.R. Osteopontin knockdown in the kidneys of hyperoxaluric rats leads to reduction in renal calcium oxalate crystal deposition. Urolithiasis 2014, 42, 195–202. [Google Scholar]
- Randhawa, R.; Bhardwaj, R.; Kaur, T. Amelioration of hyperoxaluria-induced kidney dysfunction by chemical chaperone 4-phenylbutyric acid. Urolithiasis 2019, 47, 171–179. [Google Scholar] [CrossRef]
- Khan, S.R. Animal models of kidney stone formation: An analysis. World J. Urol. 1997, 15, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.S.; Ma, M.C.; Chen, J.; Chen, C.F. Changes in the oxidant–antioxidant balance in the kidney of rats with nephrolithiasis induced by ethylene glycol. J. Urol. 2002, 167, 2584–2593. [Google Scholar] [CrossRef]
- Murray, R.L. Creatinine. In Clinical Chemistry; Kaplan, A., Pesce, A.J., Eds.; CV Mosby Co.: St Louis, MO, USA, 1984; pp. 1247–1253. [Google Scholar]
- Tabacco, A.; Meiattini, F.; Moda, E.; Tarli, P. Simplified enzymic/colorimetric serum urea nitrogen determination. Clin. Chem. 1979, 25, 336–337. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980, 26, 227–231. [Google Scholar] [CrossRef]
- Young, D.S. Effects of Drugs on Clinical Laboratory Tests, 5th ed.; American Association for Clinical Chemistry Press: Washington, DC, USA, 2000. [Google Scholar]
- Gindler, E.M.; King, J.D. Rapid colorimetric determination of calcium in biologic fluids with methylthymol blue. Am. J. Clin. Pathol. 1972, 58, 376–382. [Google Scholar] [CrossRef]
- Bagshaw, S.M.; Bennett, M.; Devarajan, P.; Bellomo, R. Urine biochemistry in septic and non-septic acute kidney injury: A prospective observational study. J. Crit. Care 2013, 28, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Banchroft, J.D.; Stevens, A.; Turner, D.R. Theory and Practice of Histological Techniques, 4th ed.; Churchill Livingston: New York, NY, USA; London, UK; San Francisco, CA, USA; Tokyo, Japan, 1996. [Google Scholar]
- Gao, H.; Zhou, Y.W. Inhibitory effect of picroside II on hepatocyte apoptosis. Acta Pharmacol. Sin. 2005, 26, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Roa, M.; Blane, K.; Zonneberg, M. One-Way Analysis of Variance. PC-STAT, Version IA (C) Copyright; The University of Georgia: Athens, Georgia, 1985. [Google Scholar]
- Rathee, D.; Rathee, P.; Rathee, S.; Rathee, D. Phytochemical screening and antimicrobial activity of Picrorrhiza kurroa, an Indian traditional plant used to treat chronic diarrhea. Arab. J. Chem. 2016, 9, S1307–S1313. [Google Scholar] [CrossRef] [Green Version]
- Al-Marzoqi, A.H.; Hadi, M.Y.; Hameed, I.H. Determination of metabolites products by Cassia angustifolia and evaluate antimicobial activity. J. Pharmacogn. Phytother. 2016, 8, 25–48. [Google Scholar]
- Dardiry, M.H.O.; Mohamed, A.A.A.; Abdelrady, E. Effect of lead (Pb) on phytochemical variability of Jatropha curcas (L.): A versatile perennial of Euphorbiaceae family. J. Biol. Stud. 2018, 1, 133–145. [Google Scholar]
- Jin, H.U.; Lee, J.H.; Kim, D.H.; Kim, K.; Lee, G.W.; Choi, S.J.; Chang, P.; Paik, H. Antioxidative and nitric oxide scavenging activity of branched-chain amino acids. Food Sci. Biotechnol. 2015, 24, 1555–1558. [Google Scholar] [CrossRef]
- Cecotti, R.; Carpana, E.; Bergomi, P.; Tava, A. Volatile constituents of Trifolium pratense spp. nivale quantified at different growth stages, and evaluation of their antimicrobial activity. Nat. Prod. Commun. 2013, 8, 1625–1628. [Google Scholar] [CrossRef] [Green Version]
- Semwal, P.; Painuli, S. Antioxidant, antimicrobial, and GC-MS profiling of Saussurea obvallata (Brahma Kamal) from Uttarakhand Himalaya. Semwal Painuli Clin. Phytosci. 2019, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Moutevelis-Minakakis, P.; Papavassilopoulou, E.; Michas, G.; Georgikopoulou, K.; Ragoussi, M.; Neophytou, N.; Zoumpoulakis, P.; Mavromoustakos, T.; Hadjipavlou-Litina, D. Synthesis, in silico docking experiments of new 2-pyrrolidinone derivatives and study of their anti-inflammatory activity. Bioorg. Med. Chem. 2011, 19, 2888–2902. [Google Scholar] [CrossRef]
- Thangam, R.; Suresh, V.; Rajkumar, M.; Vincent, J.D.; Gunasekaran, P.; Anbazhagan, C.; Kaveri, K.; Kannan, S. Antioxidant and in vitro anticancer effect of 2-pyrrolidinone rich fraction of Brassica oleracea var. capitata through induction of apoptosis in human cancer cells. Phytother. Res. 2013, 27, 1664–1670. [Google Scholar] [CrossRef]
- Jadeja, R.N.; Powell, F.L.; Martin, P.M. Repurposing fumaric acid esters to treat conditions of oxidative stress and inflammation: A promising emerging approach with broad potential. In Drug Repurposing; Farid, A.B., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- Kontogiorgis, C.A.; Hadjipavlou-Litina, D.J.; Schulz, E. Antioxidant activity of DLω-phenyl-amino acid octyl esters with anti-inflammatory activity: Correlation of the structure with lipophilicity. Arzneimittelforschung 2001, 51, 485–488. [Google Scholar]
- Lin-Holderer, J.; Li, L.; Gruneberg, D.; Marti, H.H.; Kunze, R. Fumaric acid esters promote neuronal survival upon ischemic stress through activation of the Nrf2 but not HIF-1 signaling pathway. Neuropharmacology 2016, 105, 228–240. [Google Scholar] [CrossRef]
- Schulz, E.; Kobow, M.; Sprung, W.D. Analgesic and anti-inflammatory activity of DL-2-(alkoxyphenyl)glycine octyl esters. Agents Actions 1991, 32, 65–66. [Google Scholar] [CrossRef]
- Junnila, M.; Rahko, T.; Sukura, A.; Lindberg, L.A. Reduction of carbon tetrachloride-induced hepatotoxic effects by oral administration of betaine in male Han-Wistar rats: A morphometric histological study. Vet. Pathol. 2000, 37, 231–238. [Google Scholar] [CrossRef]
- Yokogawa, K.; Watanabe, M.; Takeshita, H.; Nomura, M.; Mano, Y.; Miyamoto, K. Serum aminotransferase activity as a predictor of clearance of drugs metabolized by CYP isoforms in rats with acute hepatic failure induced by carbon tetrachloride. Int. J. Pharma. 2004, 269, 479–489. [Google Scholar] [CrossRef]
- Ezeuko, V.C.; Nwokocha, C.R.; Mounmbegna, P.E.; Nriagu, C.C. Effects of Zingiber officinale on liver function of mercuric chloride-induced hepatotoxicity in adult Wistar rats. Electron. J. Biomed. 2007, 3, 40–45. [Google Scholar]
- de Water, R.; Noordermeer, C.; van der Kwast, T.H.; Nizze, H.; Boevé, E.R.; Kok, D.J.; Schröder, F.H. Calcium oxalate nephrolithiasis: Effect of renal crystal deposition on the cellular composition of the renal interstitium. Am. J. Kidney Dis. 1999, 33, 761–771. [Google Scholar] [CrossRef]
- Lulat, S.I.; Yadav, Y.C.; Balaraman, R.; Maheshwari, R. Antiurolithiatic effect of lithocare against ethylene glycol-induced urolithiasis in Wistar rats. J. Pharmacol. 2016, 48, 78–82. [Google Scholar]
- Wang, J.; Zhang, Q.; Jin, W.; Niu, X.; Zhang, H. Effects and mechanism of low molecular weight fucoidan in mitigating the peroxidative and renal damage induced by adenine. Carbohydr. Polym. 2011, 84, 417–423. [Google Scholar] [CrossRef]
- Saeidi, J.; Bozorgi, H.; Zendehdel, A.; Mehrzad, J. Therapeutic effects of aqueous extracts of Petroselinum sativum on ethylene glycol-induced kidney calculi in rats. Urol. J. 2012, 9, 361–366. [Google Scholar]
- Aggarwal, D.; Kaushal, R.; Kaur, T.; Bijarnia, R.K.; Puri, S.; Singla, S.K. The most potent antilithiatic agent ameliorating renal dysfunction and oxidative stress from Bergenia ligulata rhizome. J. Ethnopharmacol. 2014, 158, 85–93. [Google Scholar] [CrossRef]
- Noorafshan, A.; Karbalay-Doust, S.; Karimi, F. Diosmin reduces calcium oxalate deposition and tissue degeneration in nephrolithiasis in rats: A stereological study. Korean J. Urol. 2013, 54, 252–257. [Google Scholar] [CrossRef]
- Heather, M.; David, W.; Andrew, J. Macrophages and the kidney. Hypertension 2004, 13, 285–290. [Google Scholar]
- Sirag, H.M. Biochemical and Hematological Studies for the Protective effect of oyster mushroom (Pleurotus ostreatus) against glycerol-induced acute renal failure in rats. J. Biol. Sci. 2009, 9, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.J.; Moon, M.E.; Park, H.S.; Kim, H.W.; Im, S.Y.; Lee, J.H.; Kim, Y.H. Effects of chitosan oligosaccharide (COS) on the glycerol-induced acute renal failure in vitro and in vivo. Food Chem. Toxicol. 2008, 46, 710–716. [Google Scholar] [CrossRef]
- Shah, J.G.; Patel, B.G.; Patel, S.B.; Patel, R.K. Antiurolithiatic and antioxidant activity of Hordeum vulgare seeds on ethylene glycol-induced urolithiasis in rats. Indian J. Pharmacol. 2012, 44, 672–677. [Google Scholar] [CrossRef]
- Ahmed, A.; Wadud, A.; Jahan, N.; Bilal, A.; Hajera, S. Efficacy of Adiantum capillus veneris Linn in chemically induced urolithiasis in rats. J. Ethnopharmacol. 2013, 146, 411–416. [Google Scholar] [CrossRef]
- Bijarnia, R.K.; Bachtler, M.; Chandak, P.G.; van Goor, H.; Pasch, A. Sodium thiosulfate ameliorates oxidative stress and preserves renal function in hyperoxaluric rats. PLoS ONE 2015, 10, e0124881. [Google Scholar] [CrossRef]
- Adeneye, A.A.; Benebo, A.S. Protective effect of the aqueous leaf and seed extract of Phyllanthus amarus on gentamicin and acetaminophen-induced nephrotoxic rats. J. Ethnopharmacol. 2008, 118, 318–323. [Google Scholar] [CrossRef]
- Kim, H.Y.; Yokozawa, T.; Nakagawa, T.; Sasaki, S. Protective effect of gamma-aminobutyric acid against glycerol-induced acute renal failure in rats. Food Chem. Toxicol. 2004, 42, 2009–2014. [Google Scholar] [CrossRef]
- He, L.; Shen, P.; Fu, Q.; Li, J.; Dan, M.; Wang, X.; Jia, W. Nephro-protective effect of Kangqianling decoction on chronic renal failure rats. J. Ethnopharmacol. 2009, 122, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Ghodasara, J.; Pawar, A.; Deshmukh, C.; Kuchekar, B. Inhibitory effect of rutin and curcumin on experimentally-induced calcium oxalate urolithiasis in rats. Pharmacogn. Res. 2010, 2, 388–392. [Google Scholar]
- Siener, R. Impact of dietary habits on stone incidence. Urol. Res. 2006, 34, 131–133. [Google Scholar] [CrossRef]
- Selvam, R.; Kalaiselvi, P.; Govindaraj, A.; Murugan, V.M.; Satish Kumar, A.S. Effect of A. lanata leaf extract and Vediuppu chunnam on the urinary risk factors of calcium oxalate urolithiasis during experimental hyperoxaluria. Pharmacol. Res. 2001, 43, 89–93. [Google Scholar] [CrossRef]
- Soundararajan, P.; Mahesh, R.; Ramesh, T.; Begum, V.H. Effect of Aerva lanata on calcium oxalate urolithiasis in rats. Indian J. Exp. Biol. 2006, 44, 981–986. [Google Scholar]
- Naguib, Y.M.; Azmy, R.M.; Samaka, R.M.; Salem, M.F. Pleurotus ostreatus opposes mitochondrial dysfunction and oxidative stress in acetaminophen-induced hepato-renal injury. BMC Complement. Altern. Med. 2014, 14, 494. [Google Scholar] [CrossRef]
- Rajasekar, R.; Jayaprakash, K. Protective effect of Agaricus bisporus (Button mushroom) on cisplatin induced nephrotoxicity in albino rats. World J. Pharmacol. Sci. 2016, 4, 366–372. [Google Scholar]
- Hart, P.D.; Bakris, G.L. Should β-blockers be used to control hypertension in people with chronic kidney disease? Semin. Nephrol. 2007, 27, 555–564. [Google Scholar] [CrossRef]
- Carvalho Rodrigues, M.A.; Rodrigues, J.L.; Martins, N.M.; Barbosa, F.; Curti, C.; Santos, N.A.G.; Santos, A.C. Carvedilol protects against cisplatin-induced oxidative stress, redox state unbalance and apoptosis in rat kidney mitochondria. Chem. Biol. Interact. 2011, 189, 45–51. [Google Scholar] [CrossRef]
- Gadge, N.B.; Jalalpure, S.S. Curative treatment with extracts of Bombax ceiba fruit reduces risk of calcium oxalate urolithiasis in rats. Pharma. Biol. 2012, 50, 310–317. [Google Scholar] [CrossRef]
- Patel, P.K.; Patel, M.A.; Vyas, B.A.; Shah, D.R.; Gandhi, T.R. Antiurolithiatic activity of saponin rich fraction from the fruits of Solanum xanthocarpum Schrad. & Wendl. (Solanaceae) against ethylene glycol induced urolithiasis in rats. J. Ethnopharmacol. 2012, 144, 160–170. [Google Scholar]
- Bodakhe, K.S.; Namdeo, K.P.; Patra, K.C.; Machwal, L.; Pareta, S.K. A polyherbal formulation attenuates hyperoxaluria-induced oxidative stress and prevents subsequent deposition of calcium oxalate crystals and renal cell injury in rat kidneys. Chin. J. Nat. Med. 2013, 11, 466–471. [Google Scholar] [CrossRef]
- Saha, S.; Shrivastav, P.S.; Verma, R.J. Antioxidative mechanism involved in the preventive efficacy of Bergenia ciliata rhizomes against experimental nephrolithiasis in rats. Pharm. Biol. 2014, 52, 712–722. [Google Scholar] [CrossRef]
- Karadi, R.V.; Gadge, N.B.; Alagawadi, K.R.; Savadi, R.V. Effect of Moringa oleifera Lam. root-wood on ethylene glycol induced urolithiasis in rats. J. Ethnopharmacol. 2006, 105, 306–311. [Google Scholar] [CrossRef]
- Freitas, A.M.; Schor, N.; Boim, M.A. The effect of Phyllanthus niruri on urinary inhibitors of calcium oxalate crystallization and other factors associated with renal stone formation. Br. J. Urol. Int. 2002, 89, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Thamilselvan, S.; Khan, S.R.; Menon, M. Oxalate and calcium oxalate mediated free radical toxicity in renal epithelial cells: Effect of antioxidants. Urol. Res. 2003, 31, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Scheid, C.R.; Cao, L.C.; Honeyman, T.; Jonassen, J.A. How elevated oxalate can promote kidney stone disease: Changes at the surface and in the cytosol of renal cells that promote crystal adherence and growth. Front. Biosci. 2004, 9, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandhare, A.D.; Patil, M.V.K.; Bodhankar, S.L. l-Arginine attenuates the ethylene glycol induced urolithiasis in ininephrectomized hypertensive rats: Role of KIM-1, NGAL, and NOs. Ren. Fail. 2015, 37, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Cruzan, G.; Corley, R.A.; Hard, G.C.; Mertens, J.J.; McMartin, K.E.; Snellings, W.M.; Gingell, R.; Deyo, J.A. Subchronic toxicity of ethylene glycol in Wistar and F-344 rats related to metabolism and clearance of metabolites. Toxicol. Sci. 2004, 81, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Bashir, S.; Khan, S.R.; Gilani, A.H. Antiurolithic activity of Origanum vulgare is mediated through multiple pathways. BMC Complement. Altern. Med. 2011, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.R.; Glenton, P.A.; Byer, K.J. Modeling of hyperoxaluric calcium oxalate nephrolithiasis: Experimental induction of hyperoxaluria by hydroxy-L-proline. Kidney Int. 2006, 70, 914–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, T.; Bijarnia, R.K.; Singla, S.K.; Tandon, C. In vivo efficacy of Trachyspermum ammi anticalcifying protein in urolithiatic rat model. J. Ethnopharmacol. 2009, 126, 459–462. [Google Scholar] [CrossRef]
- Pawar, A.T.; Gaikwad, G.D.; Metkari, K.S.; Tijore, K.A.; Ghodasara, J.V.; Kuchekar, B.S. Effect of Terminalia chebula fruit extract on ethylene glycol induced urolithiasis in rats. Biomed. Aging Pathol. 2012, 2, 99–103. [Google Scholar] [CrossRef]
- Makasana, A.; Ranpariya, V.; Desai, D.; Mendpara, J.; Parekh, V. Evaluation for the anti-urolithiatic activity of Launaea procumbens against ethylene glycol-induced renal calculi in rats. Toxicol. Rep. 2014, 1, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liang, Q.; Sun, Y.; Diao, L.; Qin, Z.; Wang, W.; Lu, J.; Fu, S.; Ma, B.; Yue, Z. Potential mechanisms responsible for the antinephrolithic effects of an aqueous extract of Fructus aurantii. Evid. Based Complement. Altern. Med. 2015, 2015, 491409. [Google Scholar] [CrossRef] [Green Version]
- Bashir, S.; Gilani, A.H. Antiurolithic effect of berberine is mediated through multiple pathways. Eur. J. Pharmacol. 2011, 651, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Corley, R.A.; Wilson, D.M.; Hard, G.C.; Stebbins, K.E.; Bartels, M.J.; Soelberg, J.J.; Dryzga, M.D.; Gingell, R.; McMartin, K.E.; Snellings, W.M. Dosimetry considerations in the enhanced sensitivity of male Wistar rats to chronic ethylene glycol-induced nephrotoxicity. Toxicol. Appl. Pharmacol. 2008, 228, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Hackett, R.L.; Shevock, P.N.; Khan, S.R. Alteration in MDCK and LLC-PK1 cells exposed to oxalate and calcium oxalate monohydrate crystals. Scanning Microsc. 1995, 9, 587–596. [Google Scholar] [PubMed]
- Scheid, C.R.; Koul, H.K.; Kennington, L.; Hill, W.A.; Luber-Narod, J.; Jonassen, J.; Honeyman, T.; Menon, M. Oxalate-induced damage to renal tubular cells. Scanning Microsc. 1995, 9, 1097–1105. [Google Scholar]
- Gilmore, T.D. Introduction to NF-κB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [Green Version]
- Sanz, A.B.; Sanchez-Niño, M.D.; Ramos, A.M.; Moreno, J.A.; Santamaria, B.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. NF-κB in renal inflammation. J. Am. Soc. Nephrol. 2010, 21, 1254–1262. [Google Scholar] [CrossRef] [Green Version]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Brasier, A.R. The NF-κB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar] [CrossRef]
- Schreck, R.; Peter Rieber, P.; Baeuerle, P.A. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-κB transcription factor and HIV-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Jost, P.J.; Ruland, J. Aberrant NF-κB signaling in lymphoma: Mechanisms, consequences, and therapeutic implications. Blood 2007, 109, 2700–2707. [Google Scholar] [CrossRef] [Green Version]
- Garufi, A.; Ubertini, V.; Mancini, F.; D’Orazi, V.; Baldari, S.; Moretti, F.; Bossi, G.; D’Orazi1, G. The beneficial effect of Zinc (II) on low-dose chemotherapeutic sensitivity involves p53 activation in wild-type p53-carrying colorectal cancer cells. J. Exp. Clin. Cancer Res. 2015, 34, 87. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.M.; Fahim, H.I.; Ahmed, H.Y.; Al-Muzafar, H.M.; Ahmed, R.R.; Amin, K.A.; El-Nahass, E.; Abdelazeem, W.H. The preventive effects and the mechanisms of action of navel orange peel hydroethanolic extract, naringin, and naringenin in N-acetyl-p-aminophenol-induced liver Injury in Wistar rats. Oxid. Med. Cell. Longev. 2019, 2019, 2745352. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.M.; Ali, T.M.; Abdel Gaid, M.A.; Elberry, A.A. Effects of enalapril and paricalcitol treatment on diabetic nephropathy and renal expressions of TNF-α, p53, caspase-3 and Bcl-2 in STZ-induced diabetic rats. PLoS ONE 2019, in press. [Google Scholar]
- Borghetti, G.; Yamaguchi, A.A.; Aikawa, J.; Yamazaki, R.K.; Pereira de Brito, G.A.; Luiz Claudio Fernandes, L.C. Fish oil administration mediates apoptosis of Walker 256 tumor cells by modulation of p53, Bcl-2, caspase-7 and caspase-3 protein expression. Lipids Health Dis. 2015, 14, 94. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, K.; Dashzeveg, N.; Yoshida, K. Tumor suppressor p53 induces miR-1915 processing to inhibit Bcl-2 in the apoptotic response to DNA damage. FEBS J. 2014, 281, 2937–2944. [Google Scholar] [CrossRef]
- Barkett, M.; Gilmore, T.D. Control of apoptosis by Rel/NF-κB transcription factors. Oncogene 1999, 18, 6910–6924. [Google Scholar] [CrossRef] [Green Version]
- Ashikawa, K.; Shishodia, S.; Fokt, I.; Priebe, W.; Aggarwal, B.B. Evidence that activation of nuclear factor-kappaB is essential for the cytotoxic effects of doxorubicin and its analogues. Biochem. Pharmacol. 2004, 67, 353–364. [Google Scholar] [CrossRef]
- Campbell, K.J.; Rocha, S.; Perkins, N.D. Active repression of antiapoptotic gene expression by RelA(p65) NF-kappa B. Mol. Cell. 2004, 13, 853–865. [Google Scholar] [CrossRef]
- Ho, W.C.; Dickson, K.M.; Barker, P.A. Nuclear factor-kappaB induced by doxorubicin is deficient in phosphorylation and acetylation and represses nuclear factor-kappaB-dependent transcription in cancer cells. Cancer Res. 2005, 65, 4273–4281. [Google Scholar] [CrossRef] [Green Version]
- Bednarski, B.K.; Baldwin, A.S., Jr.; Kim, H.J. Addressing reported pro-apoptotic functions of NF-kB:targeted inhibition of canonical NF-kB enhances the apoptotic effects of doxorubicin. PLoS ONE 2009, 4, e6992. [Google Scholar] [CrossRef] [Green Version]
- Parrondo, R.; de las Pozas, A.; Reiner, T.; Rai, P.; Perez-Stable, C. NF-κB activation activation enhances cell death by antimitotic drugs in human prostate cancer cells. Mol. Cancer 2010, 9, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmet, N.M.; Ender, O. Effect of urinary stone disease and its treatment on renal function. World J. Nephrol. 2015, 4, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Wang, W.; Peck, A.B.; Khan, S.R. Activation of the NLRP3 inflammasome in association with calcium oxalate crystal induced reactive oxygen species in kidneys. J. Urol. 2015, 193, 1684–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, C.; Sarikaya, S.; Basak, K.; Cetinel, C.A.; Narter, F.; Eryildirim, B.; Saglam, E.; Sarica, K. Limitation of apoptotic changes and crystal deposition by Tutukon following hyperoxaluria-induced tubular cell injury in rat model. Urolithisasis 2015, 43, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gao, B.; Wang, Y.; Liu, Z.; Yasui, T.; Liu, P.; Liu, J.; Emmanuel, N.; Zhu, Q.; Xiao, C. Renal tubular epithelial cell injury, apoptosis and inflammation are involved in melamine-related kidney stone formation. Urol. Res. 2012, 40, 717–723. [Google Scholar] [CrossRef]
- Hu, H.; Chen, W.; Ding, J.; Jia, M.; Yin, J.; Guo, Z. Fasudil prevents calcium oxalate crystal deposit and renal fibrogenesis in glyoxylate-induced nephrolithic mice. Exp. Mol. Pathol. 2015, 98, 277–285. [Google Scholar] [CrossRef]
- Hsieh, N.; Shih, C.H.; Chen, H.Y. Effects of Tamm-Horsfall protein on the protection of MCDK cells from oxalate induced free radical injury. Urol. Res. 2003, 31, 10–16. [Google Scholar] [CrossRef]
- Jedinak, A.; Dudhgaonkar, S.; Wu, Q.; Simon, J.; Sliva, D. Anti-inflammatory activity of edible oyster mushroom is mediated through the inhibition of NF-κB and AP-1 signaling. Nutr. J. 2011, 2011, 10–52. [Google Scholar] [CrossRef] [Green Version]
- Amstad, P.A.; Liu, H.; Ichimiya, M.; Berezesky, I.K.; Trump, B.F.; Buhimschi, I.A.; Gutierrez, P.L. BCL-2 is involved in preventing oxidant-induced cell death and in decreasing oxygen radical production. Redox Rep. 2001, 6, 351–362. [Google Scholar] [CrossRef]
- Volman, J.J.; Helsper, J.P.; Wei, S.; Baars, J.J.; van Griensven, L.J.; Sonnenberg, A.S.; Mensink, R.P.; Plat, J. Effects of mushroom-derived beta-glucan-rich polysaccharide extracts on nitric oxide production by bone marrow-derived macrophages and nuclear factor-kappaB transactivation in Caco-2 reporter cells: Can effects be explained by structure? Mol. Nut. Food Res. 2010, 54, 268–276. [Google Scholar] [CrossRef]
- Arab, H.H.; El-Sawalhi, M.M. Carvedilol alleviates adjuvant-induced arthritis and subcutaneous air pouch edema: Modulation of oxidative stress and inflammatory mediators. Toxicol. Appl. Pharmacol. 2013, 268, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Yasar, A.; Erdemir, F.; Parlaktas, B.S.; Atilgan, D.; Koseoglu, R.D.; Saylan, O.; Firat, F. The effect of carvedilol on serum and tissue oxidative stress parameters in partial ureteral obstruction induced rat model. Kaohsiung J. Med. Sci. 2013, 29, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Shioji, K.; Kihara, Y.; Takenaka, H.; Onozawa, Y.; Kishimoto, C. Cardioprotective effects of carvedilol on acute autoimmune myocarditis: Anti-inflammatory effects associated with antioxidant property. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H83–H90. [Google Scholar] [CrossRef]
- Sahu, B.D.; Koneru, M.; Bijargi, S.R.; Kota, A.; Sistla, R. Chromium-induced nephrotoxicity and ameliorative effect of carvedilol in rats: Involvement of oxidative stress, apoptosis and inflammation. Chem. Biol. Interact. 2014, 223, 69–79. [Google Scholar] [CrossRef] [PubMed]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushroom | P. ostreatus | A. bisporus | |
|---|---|---|---|
| Test | |||
| 1—Carbohydrates and/or glycosides (Molish test) | +ve | +ve | |
2—Alkaloids and/or basic nitrogenous substances
| |||
| +ve | +ve | ||
| +ve | +ve | ||
| +ve | +ve | ||
3—Flavonoides
| |||
| +ve | +ve | ||
| −ve | −ve | ||
| 4—Resins test | +ve | +ve | |
5—Tannins
| |||
| −ve | −ve | ||
| +ve | +ve | ||
6—Unsaturated sterols and/or triterpenes
| |||
| +ve | +ve | ||
| +ve | +ve | ||
| Number | Retention Time (Minutes) | Compound or Group (From Central Library Search Report) | Area % (≥0.5%) |
|---|---|---|---|
| 1 | 1.87 | (S)-(+)-Isoleucinol | 23.0 |
| 2 | 2.05 | Diethylhydroxylamine | 3.6 |
| 3 | 2.13 | 3-(dimethylamino)-propanenitrile | 1.4 |
| 4 | 2.17 | 1-Methylcyclopropanemethanol | 1.0 |
| 5 | 2.23 | N-Butyl-formamide | 3.5 |
| 6 | 2.54 | 2-(ethylthio) tetrahydro 2H-Pyran | 4.0 |
| 7 | 2.72 | N-Hydroxy-N-methyl methenamine | 24.2 |
| 8 | 2.85 | 2-Ethyl-2-butenal | 9.4 |
| 9 | 3.22 | 4-Methyl- 2,4,6-cycloheptatrien-1-one | 6.4 |
| 10 | 3.29 | 2-Phenylethanal | 4.2 |
| 11 | 3.47 | 2-Pyrrolidinone | 4.4 |
| 12 | 3.79 | 3-Mercapto-1-propanol | 4.0 |
| 13 | 4.30 | N,N,N′-Trimethyl-1,3-propanediamine | 3.4 |
| 14 | 4.54 | 2,5-Dihydro-3-methyl-furan | 1.2 |
| 15 | 4.72 | 2,3-Dimethyl-pentanal | 1.2 |
| 16 | 5.01 | N,N-Diethyl-N′, N′-dimethyl-1,2-ethanediamine | 3.6 |
| 17 | 6.26 | 4-Amino-1,2,5-Oxadiazole-3-carbonitrile | 0.5 |
| 18 | 10.46 | Thiomorpholine | 0.5 |
| 19 | 11.11 | 2,3′-Dipyridyl | 0.5 |
| Number | Retention Time | Compound or Group (From Central Library Search Report) | Area % (≥0.5%) |
|---|---|---|---|
| 1 | 2.69 | N-Methoxymethanamine | 30.6 |
| 2 | 3.19 | Phenyl oxirane | 5.8 |
| 3 | 3.27 | 2-Phenylethanal | 2.6 |
| 4 | 3.44 | 2-Pyrrolidinone | 7.9 |
| 5 | 3.78 | 2-Propenylidene cyclobutene | 3.0 |
| 6 | 4.31 | 1-Methoxy-3-methyl- benzene | 2.4 |
| 7 | 5.01 | 3-Pentanone | 2.1 |
| 8 | 5.40 | N-[4-Aminobutyl]aziridine | 1.7 |
| 9 | 17.33 | N-Methoxy-methanamine | 4.2 |
| 10 | 25.88 | 2,4,6,7,8,8a-Hexahydro-3,8-dimethyl-4-(1-methylethylidene)-(8S-cis)-5(1H)-azulenone | 3.2 |
| 11 | 30.45 | 6-Ethyl-2,3-dihydro-2,7-dimethyl-5-oxo-5H-oxazolo[3,2-a]pyridine-8-carbonitrile | 13.7 |
| 12 | 31.41 | Oxime, (5. alpha.) androstan-3-one | 6.5 |
| 13 | 31.92 | 6-Amino-2-Phenazinol | 2.1 |
| 14 | 32.99 | Fumaric acid, 2-heptyl octyl ester | 13.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, O.M.; Ebaid, H.; El-Nahass, E.-S.; Ragab, M.; Alhazza, I.M. Nephroprotective Effect of Pleurotus ostreatus and Agaricus bisporus Extracts and Carvedilol on Ethylene Glycol-Induced Urolithiasis: Roles of NF-κB, p53, Bcl-2, Bax and Bak. Biomolecules 2020, 10, 1317. https://doi.org/10.3390/biom10091317
Ahmed OM, Ebaid H, El-Nahass E-S, Ragab M, Alhazza IM. Nephroprotective Effect of Pleurotus ostreatus and Agaricus bisporus Extracts and Carvedilol on Ethylene Glycol-Induced Urolithiasis: Roles of NF-κB, p53, Bcl-2, Bax and Bak. Biomolecules. 2020; 10(9):1317. https://doi.org/10.3390/biom10091317
Chicago/Turabian StyleAhmed, Osama M., Hossam Ebaid, El-Shaymaa El-Nahass, Mahmoud Ragab, and Ibrahim M. Alhazza. 2020. "Nephroprotective Effect of Pleurotus ostreatus and Agaricus bisporus Extracts and Carvedilol on Ethylene Glycol-Induced Urolithiasis: Roles of NF-κB, p53, Bcl-2, Bax and Bak" Biomolecules 10, no. 9: 1317. https://doi.org/10.3390/biom10091317