An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. MSC Isolation and Growth
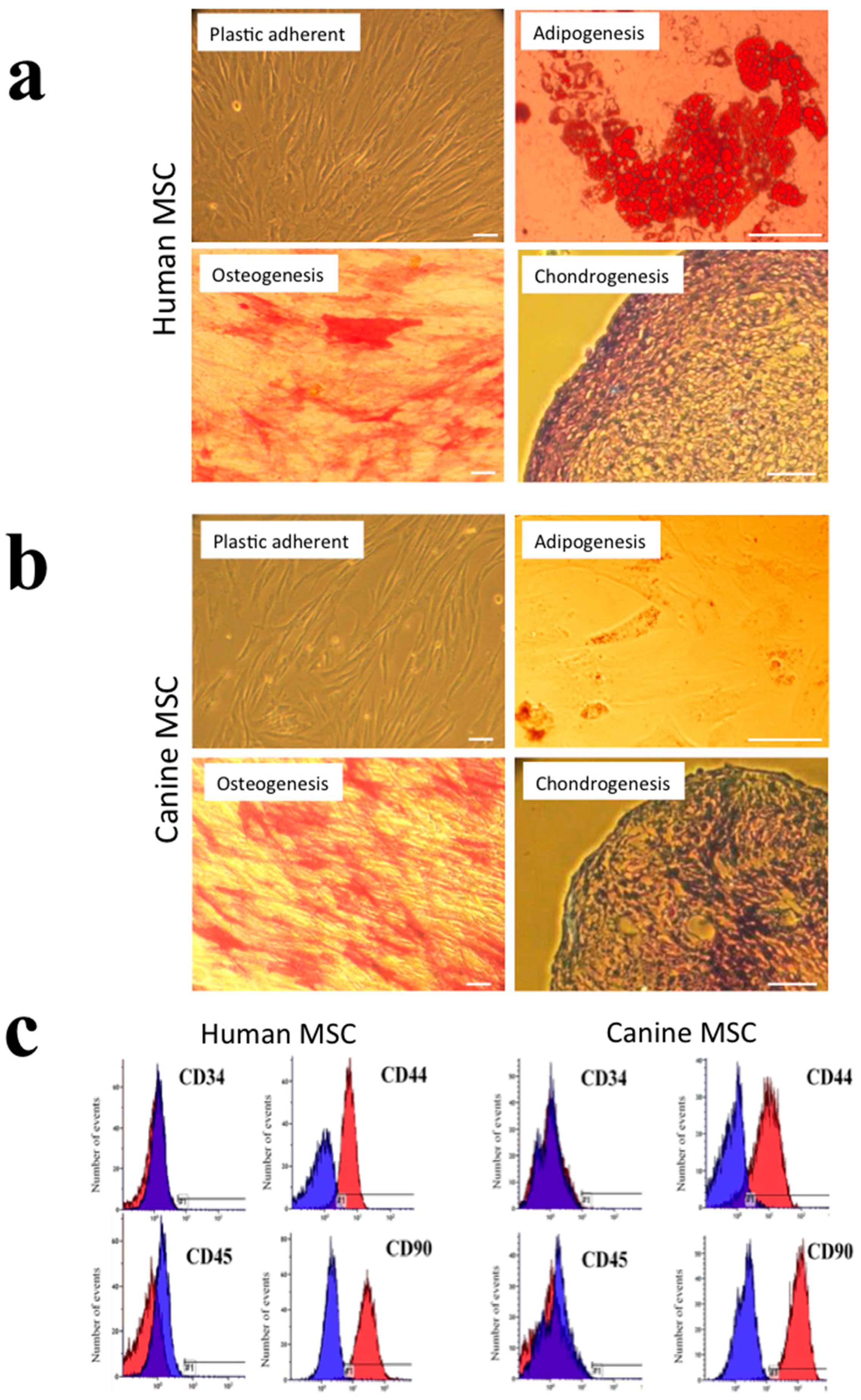
2.2. Phenotyping Human and Canine MSCs
2.2.1. Adipogenic Differentiation
2.2.2. Osteogenic Differentiation
2.2.3. Chondrogenic Differentiation
2.2.4. CD Immunoprofiling
2.3. Preparation of Conditioned Medium from Human and Canine MSC Cultures
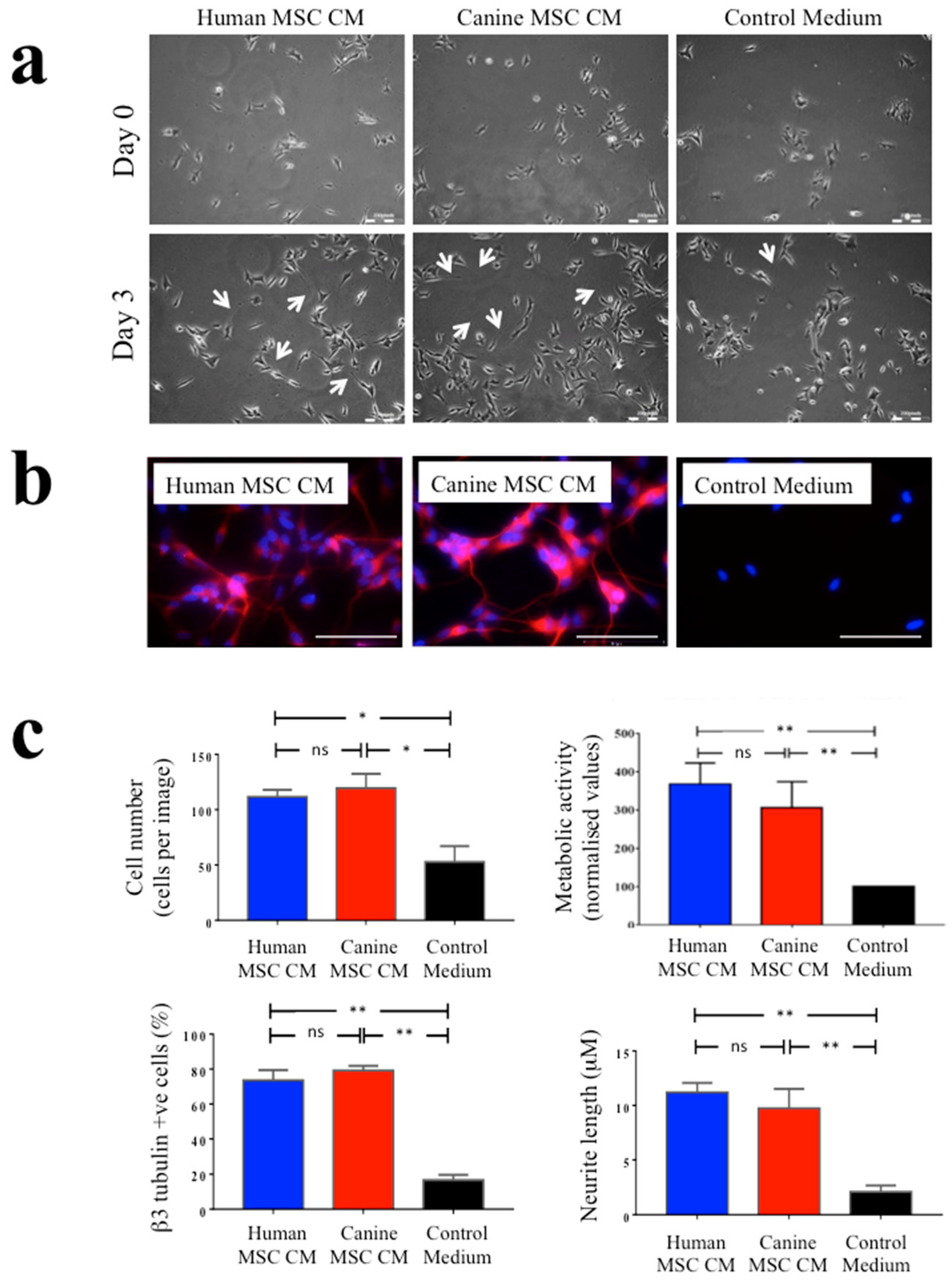
2.4. Assessment of the Neurotrophic Effects of Human and Canine MSC CM
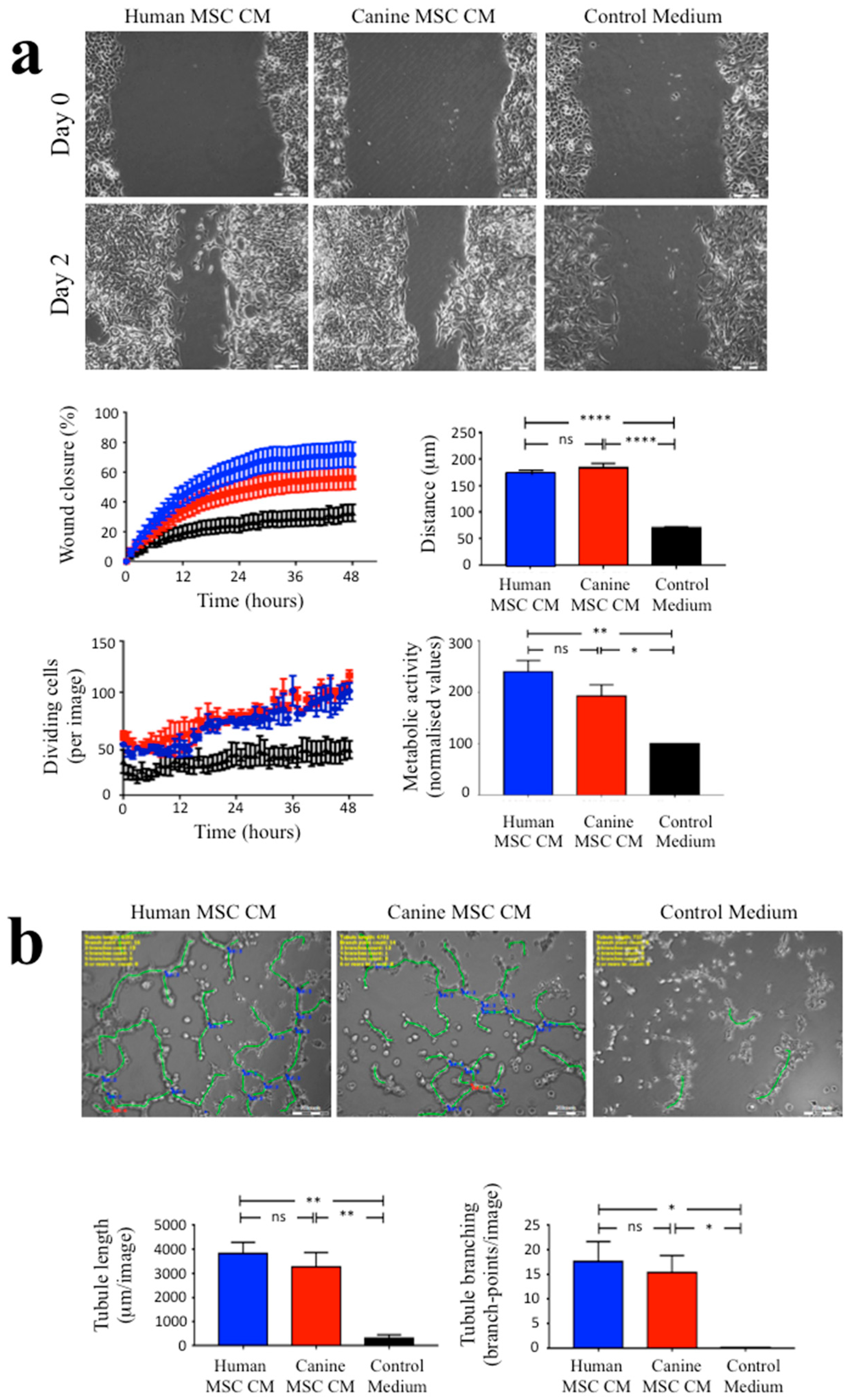
2.5. Assessment of the Angiogenic Effects of Human and Canine MSC CM
2.6. Assessment of Viable Cell Proliferation Using MTT Assays
2.7. Statistical Analysis
3. Results
3.1. Phenotypic Characterisation of Human and Canine MSC Cultures
3.2. The Neurotrophic Effects of Human and Canine MSC CM on SH-SY5Y Neuronal Cells
3.3. The Angiogenic Effects of Human and Canine MSC CM on EA.hy926 Endothelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kwon, B.K.; Streijger, F.; Hill, C.E.; Anderson, A.J.; Bacon, M.; Beattie, M.S.; Blesch, A.; Bradbury, E.J.; Brown, A.; Bresnahan, J.C.; et al. Large animal and primate models of spinal cord injury for the testing of novel therapies. Exp. Neurol. 2015, 269, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Nardone, R.; Florea, C.; Höller, Y.; Brigo, F.; Versace, V.; Lochner, P.; Golaszewski, S.; Trinka, E. Rodent, large animal and non-human primate models of spinal cord injury. Zoology 2017, 123, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Cibelli, J.; Emborg, M.E.; Prockop, D.J.; Roberts, M.; Schatten, G.; Rao, M.; Harding, J.; Mirochnitchenko, O. Strategies for improving animal models for regenerative medicine. Cell Stem Cell. 2013, 12, 271–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif-Alhoseini, M.; Khormali, M.; Rezaei, M.; Safdarian, M.; Hajighadery, A.; Khalatbari, M.M.; Safdarian, M.; Meknatkhah, S.; Rezvan, M.; Chalangari, M.; et al. Animal models of spinal cord injury: A systematic review. Spinal Cord 2017, 55, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Filipp, M.E.; Travis, B.J.; Henry, S.S.; Idzikowski, E.C.; Magnuson, S.A.; Loh, M.Y.F.; Hellebrand, D.J.; Hanna, A.S. Differences in neuroplasticity after spinal cord injury in varying animal models and humans. Neural Regen Res. 2019, 14, 7–19. [Google Scholar] [PubMed]
- Kwon, B.K.; Hillyer, J.; Tetzlaff, W. Translational research in spinal cord injury: A survey of opinion from the, S.C.I community. J. Neurotrauma 2010, 27, 21–33. [Google Scholar] [CrossRef]
- Filli, L.; Schwab, M.E. The rocky road to translation in spinal cord repair. Ann. Neurol. 2012, 72, 491–501. [Google Scholar] [CrossRef]
- Jeffery, N.D.; Smith, P.M.; Lakatos, A.; Ibanez, C.; Ito, D.; Franklin, R.J. Clinical canine spinal cord injury provides an opportunity to examine the issues in translating laboratory techniques into practical therapy. Spinal Cord 2006, 44, 584–593. [Google Scholar] [CrossRef]
- McMahill, B.G.; Borjesson, D.L.; Sieber-Blum, M.; Nolta, J.A.; Sturges, B.K. Stem cells in canine spinal cord injury-promise for regenerative therapy in a large animal model of human disease. Stem Cell Rev. Rep. 2015, 11, 180–193. [Google Scholar] [CrossRef]
- Moore, S.A.; Granger, N.; Olby, N.J.; Spitzbarth, I.; Jeffery, N.D.; Tipold, A.; Nout-Lomas, Y.S.; da Costa, R.C.; Stein, V.M.; Noble-Haeusslein, L.J.; et al. Targeting Translational Successes through CANSORT-SCI: Using Pet Dogs To Identify Effective Treatments for Spinal Cord Injury. J. Neurotrauma 2017, 34, 2007–2018. [Google Scholar] [CrossRef]
- Moore, S.A.; Zidan, N.; Spitzbarth, I.; Nout-Lomas, Y.S.; Granger, N.; da Costa, R.C.; Lebine, J.M.; Jeffrey, N.D.; Stein, V.M.; Tipold, A.; et al. Development of an International Canine Spinal Cord Injury observational registry: A collaborative data-sharing network to optimize translational studies of SCI. Spinal Cord 2018, 56, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Levine, G.J.; Porter, B.F.; Topp, K.; Noble-Haeusslein, L.J. Naturally occurring disk herniation in dogs: An opportunity for pre-clinical spinal cord injury research. J. Neurotrauma. 2011, 28, 675–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, P.; Spitzbarth, I.; Haist, V.; Stein, V.M.; Tipold, A.; Puff, C.; Beineke, A.; Baumgärtner, W. Spatio-temporal development of axonopathy in canine intervertebral disc disease as a translational large animal model for nonexperimental spinal cord injury. Brain Pathol. 2013, 23, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.T.; El Masri, W.; Osman, A.; Chowdhury, J.; Johnson, W.E. Concise review: Bone marrow for the treatment of spinal cord injury: Mechanisms and clinical applications. Stem Cells. 2011, 29, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cofano, F.; Boido, M.; Monticelli, M.; Zenga, F.; Ducati, A.; Vercelli, A.; Garbossa, D. Mesenchymal Stem Cells for Spinal Cord Injury: Current Options, Limitations, and Future of Cell Therapy. Int. J. Mol. Sci. 2019, 20, 2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Yang, X. The Efficacy and Safety of Mesenchymal Stem Cell Transplantation for Spinal Cord Injury Patients: A Meta-Analysis and Systematic Review. Cell Transplant. 2019, 28, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Sousa, N.; Salgado, A.J. Mesenchymal stem cells secretome: A new paradigm for central nervous system regeneration? Cell. Mol. Life Sci. 2013, 70, 3871–3882. [Google Scholar] [CrossRef]
- Nakajima, H.; Uchida, K.; Guerrero, A.R.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.T.; Johnson, W.E.B.; et al. Transplantation of mesenchymal stem cells promotes an alternative pathway of macrophage activation and functional recovery after spinal cord injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.T.; Uchida, K.; Bara, J.J.; Roberts, S.; El Masri, W.; Johnson, W.E. Spinal motor neurite outgrowth over glial scar inhibitors is enhanced by coculture with bone marrow stromal cells. Spine J. 2014, 14, 1722–1733. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.T.; Masri, W.E.; Osman, A.; Roberts, S.; Trivedi, J.; Ashton, B.A.; Johnson, W.E.B. The cell culture expansion of bone marrow stromal cells from humans with spinal cord injury: Implications for future cell transplantation therapy. Spinal Cord 2008, 46, 811–817. [Google Scholar] [CrossRef]
- Wright, K.T.; Griffiths, G.J.; Johnson, W.E. A comparison of high-content screening versus manual analysis to assay the effects of mesenchymal stem cell-conditioned medium on neurite outgrowth in vitro. J. Biomol. Screen. 2010, 15, 576–582. [Google Scholar] [CrossRef]
- Walter, M.N.; Kohli, N.; Khan, N.; Major, T.; Fuller, H.; Wright, K.T.; Kuiper, J.-H.; Johnson, W.E.B. Human mesenchymal stem cells stimulate EaHy926 endothelial cell migration: Combined proteomic and in vitro analysis of the influence of donor-donor variability. J. Stem Cells Regen. Med. 2015, 11, 18–24. [Google Scholar] [PubMed]
- Al Delfi, I.R.; Sheard, J.J.; Wood, C.R.; Vernallis, A.; Innes, J.F.; Myint, P.; Johnson, W.E.B. Canine mesenchymal stem cells are neurotrophic and angiogenic: An in vitro assessment of their paracrine activity. Vet. J. 2016, 217, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Granger, N.; Blamires, H.; Franklin, R.J.; Jeffery, N.D. Autologous olfactory mucosal cell transplants in clinical spinal cord injury: A randomized double-blinded trial in a canine translational model. Brain 2012, 135, 3227–3237. [Google Scholar] [CrossRef] [Green Version]
- Penha, E.M.; Meira, C.S.; Guimarães, E.T.; Mendonça, M.V.P.; Gravely, F.A.; Bahia Pinheiro, C.M.; Bahia Pinheiro, T.M.; Barrouin-Melo, S.M.; Ribeiro-Dos-Santos, R.; Botelho Pereira Soares, M. Use of Autologous Mesenchymal Stem Cells Derived from Bone Marrow for the Treatment of Naturally Injured Spinal Cord in Dogs. Stem Cells Int. 2014, 2014, 437521. [Google Scholar] [CrossRef]
- Mendonca, M.V.P.; Larocca, T.F.; de Freitas Souza, B.S.; Villarreal, C.F.; Silva, L.F.M.; Matos, A.C.; Novaes, M.A.; Pinheiro Bahia, C.M.; de Oliveira Melo Martinez, A.C.; Martins Kaneto, C.; et al. Safety and neurological assessments after autologous transplantation of bone marrow mesenchymal stem cells in subjects with chronic spinal cord injury. Stem Cell Res. Ther. 2014, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Russell, K.A.; Chow, N.H.C.; Dukoff, D.; Gibson, T.W.G.; LaMarre, J.; Betts, D.H.; Koch, T.G. Characterization and Immunomodulatory Effects of Canine Adipose Tissue- and Bone Marrow-Derived Mesenchymal Stromal Cells. PLoS ONE 2016, 11, e0167442. [Google Scholar] [CrossRef] [Green Version]
- Watt, S.M.; Gullo, F.; van der Garde, M.; Markeson, D.; Camicia, R.; Khoo, C.P.; Zwaginga, J.J. The angiogenic properties of mesenchymal stem/stromal cells and their therapeutic potential. Br. Med. Bull. 2013, 108, 25–53. [Google Scholar] [CrossRef]
- Teshima, T.; Matsuoka, A.; Shiba, M.; Dairaku, K.; Matsumoto, H.; Suzuki, R.; Koyama, H. Comparison of Properties of Stem Cells Isolated from Adipose Tissue and Lipomas in Dogs. Stem Cells Int. 2019, 2019, 1609876. [Google Scholar] [CrossRef] [Green Version]
- Humenik, F.; Cizkova, D.; Cikos, S.; Luptakova, L.; Madari, A.; Mudronova, D.; Kuricova, M.; Farbakova, J.; Spirkova, A.; Petrovova, E.; et al. Canine Bone Marrow-derived Mesenchymal Stem Cells: Genomics, Proteomics and Functional Analyses of Paracrine Factors. Mol. Cell Proteom. 2019, 18, 1824–1835. [Google Scholar] [CrossRef]
- Jung, D.I.; Ha, J.; Kang, B.T.; Kim, J.W.; Quan, F.S.; Lee, J.H.; Woo, E.-J.; Park, H.-M. A comparison of autologous and allogenic bone marrow-derived mesenchymal stem cell transplantation in canine spinal cord injury. J. Neurol. Sci. 2009, 285, 67–77. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, Y.; Rhew, D.; Kuk, M.; Kim, M.; Kim, W.H.; Kweon, O.K. Effect of the combination of mesenchymal stromal cells and chondroitinase ABC on chronic spinal cord injury. Cytotherapy 2015, 17, 1374–1383. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, S.H.; Kim, W.H.; Kweon, O.K. Transplantation of adipose derived mesenchymal stem cells for acute thoracolumbar disc disease with no deep pain perception in dogs. J. Vet. Sci. 2016, 17, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalhão, C.C.M.; Ramos, I.P.; Hochman-Mendez, C.; Brunswick, T.H.K.; Souza, S.A.L.; Gutfilen, B.; Dos Santos Goldenberg, R.C.; Coelho-Sampaio, T. Safety of Allogeneic Canine Adipose Tissue-Derived Mesenchymal Stem Cell Intraspinal Transplantation in Dogs with Chronic Spinal Cord Injury. Stem Cells Int. 2017, 2017, 3053759. [Google Scholar] [CrossRef] [PubMed]
- Vikartovska, Z.; Kuricova, M.; Farbakova, J.; Liptak, T.; Mudronova, D.; Humenik, F.; Madari, A.; Maloveska, M.; Sykova, E.; Cizkova, D. Stem Cell Conditioned Medium Treatment for Canine Spinal Cord Injury: Pilot Feasibility Study. Int. J. Mol. Sci. 2020, 21, 5129. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delfi, I.R.T.A.; Wood, C.R.; Johnson, L.D.V.; Snow, M.D.; Innes, J.F.; Myint, P.; Johnson, W.E.B. An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury. Biomolecules 2020, 10, 1301. https://doi.org/10.3390/biom10091301
Delfi IRTA, Wood CR, Johnson LDV, Snow MD, Innes JF, Myint P, Johnson WEB. An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury. Biomolecules. 2020; 10(9):1301. https://doi.org/10.3390/biom10091301
Chicago/Turabian StyleDelfi, Ibtesam R. T. Al, Chelsea R. Wood, Louis D. V. Johnson, Martyn D. Snow, John F. Innes, Peter Myint, and William E. B. Johnson. 2020. "An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury" Biomolecules 10, no. 9: 1301. https://doi.org/10.3390/biom10091301