N-Terminal Segment of TvCyP2 Cyclophilin from Trichomonas vaginalis Is Involved in Self-Association, Membrane Interaction, and Subcellular Localization

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Plasmids
2.2. Preparations of Recombinant Proteins and Peptides
2.3. SEC-MALS Experiment
2.4. Crystallization Screening and X-ray Data Collection
2.5. Refinement and Structure Determination
2.6. NMR Spectroscopy
2.7. CD Spectroscopy
2.8. Immunofluorescence Assay (IFA)
2.9. Immunoprecipitation (IP)
2.10. Western Blot Analysis
3. Results
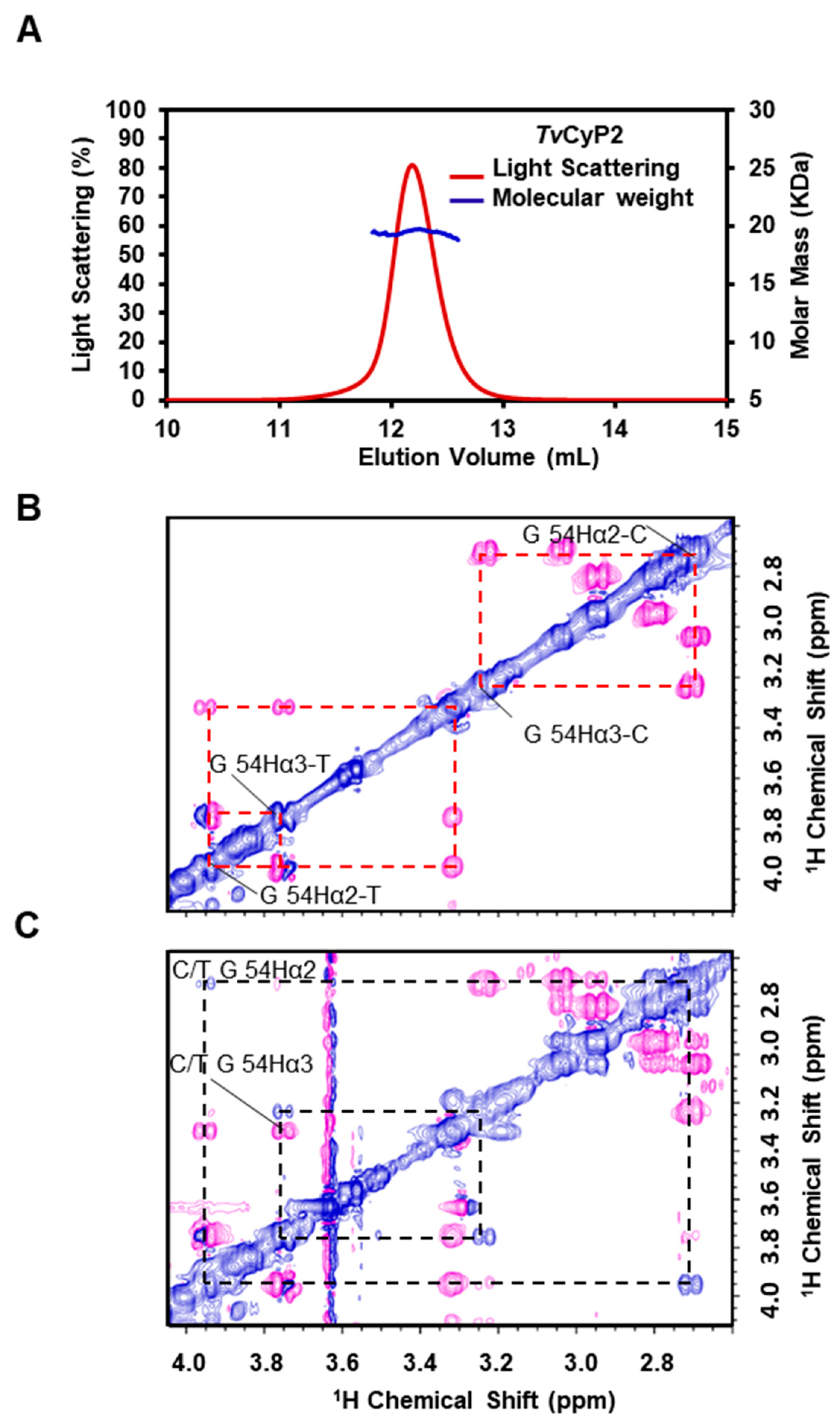
3.1. TvCyP2 Forming a Monomer and Possessing PPIase Activity
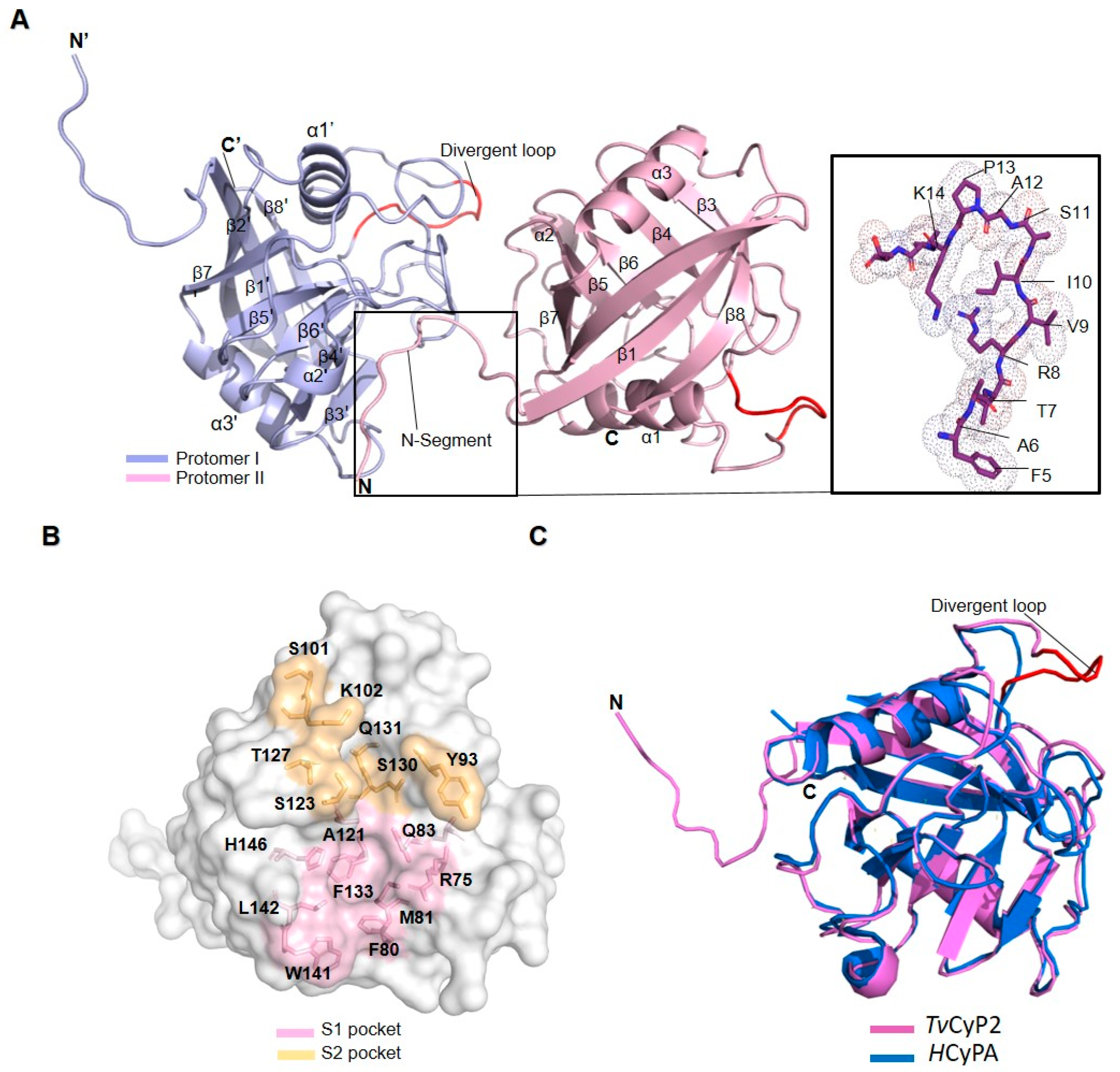
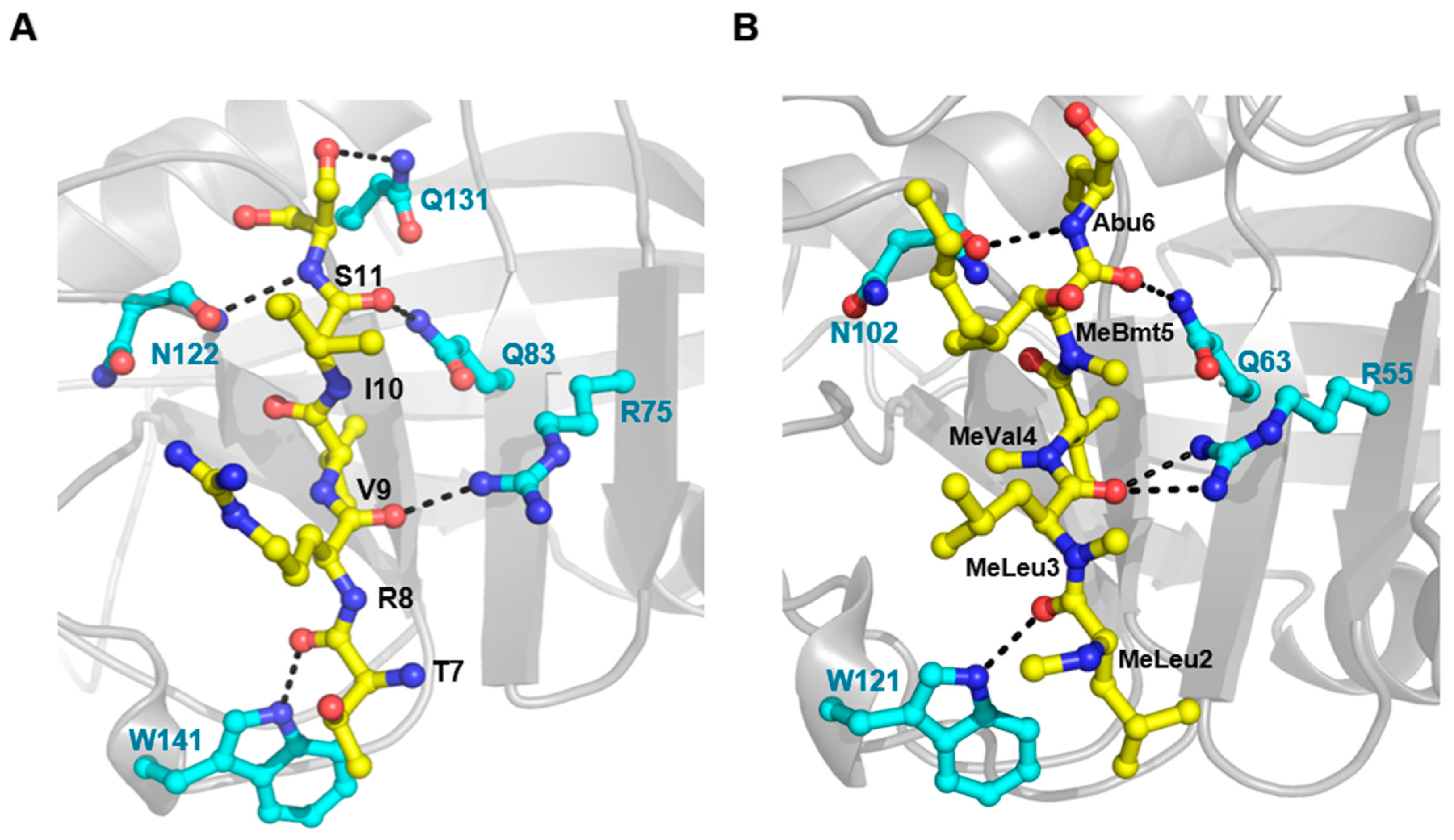
3.2. X-ray Crystal and NMR Solution Structures of TvCyP2
3.3. Self-Association of the N-Terminal Segment also Observed in Solution
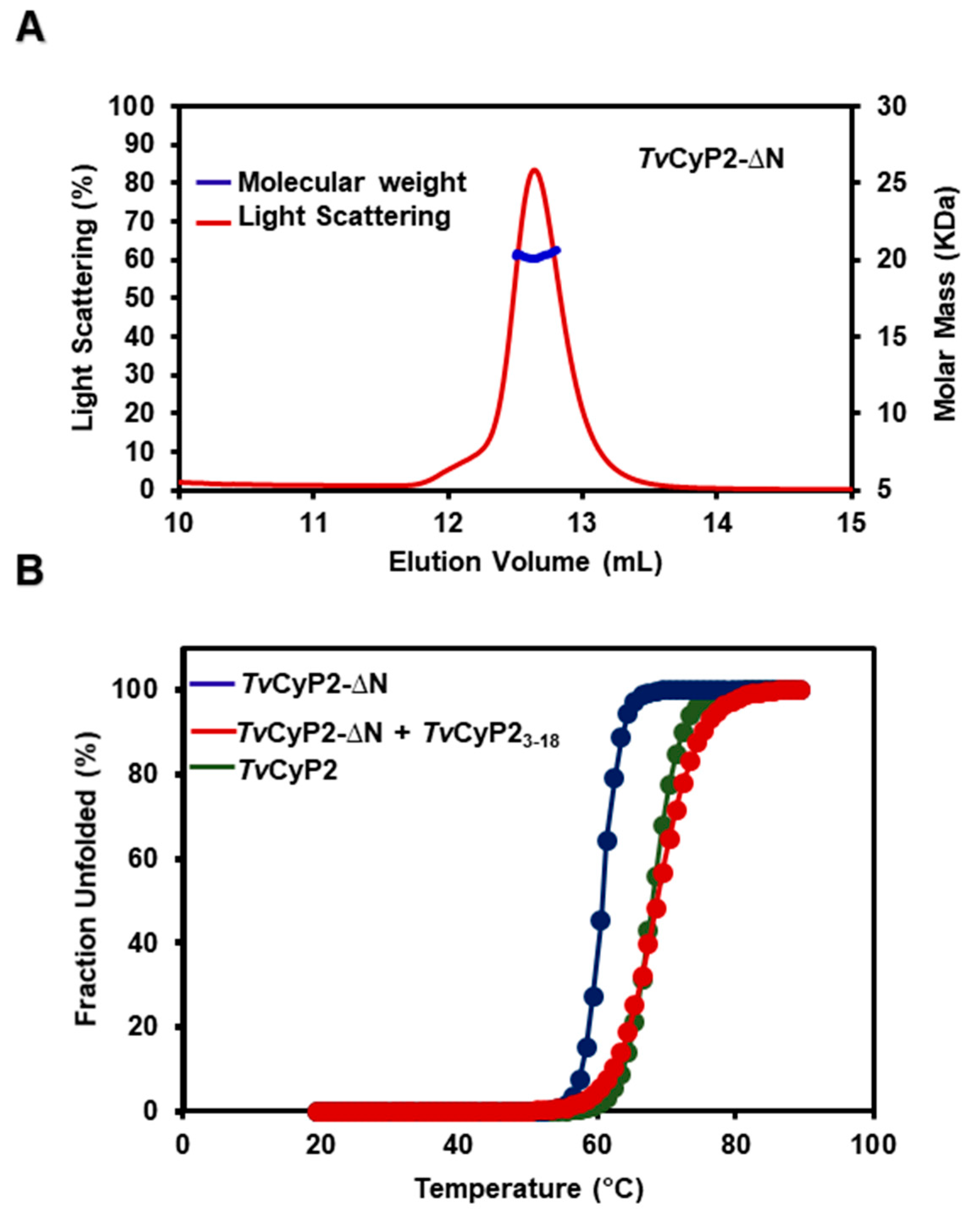
3.4. Thermostability and Structure of TvCyP2 without N-Terminal Segment (TvCyP2-∆N)
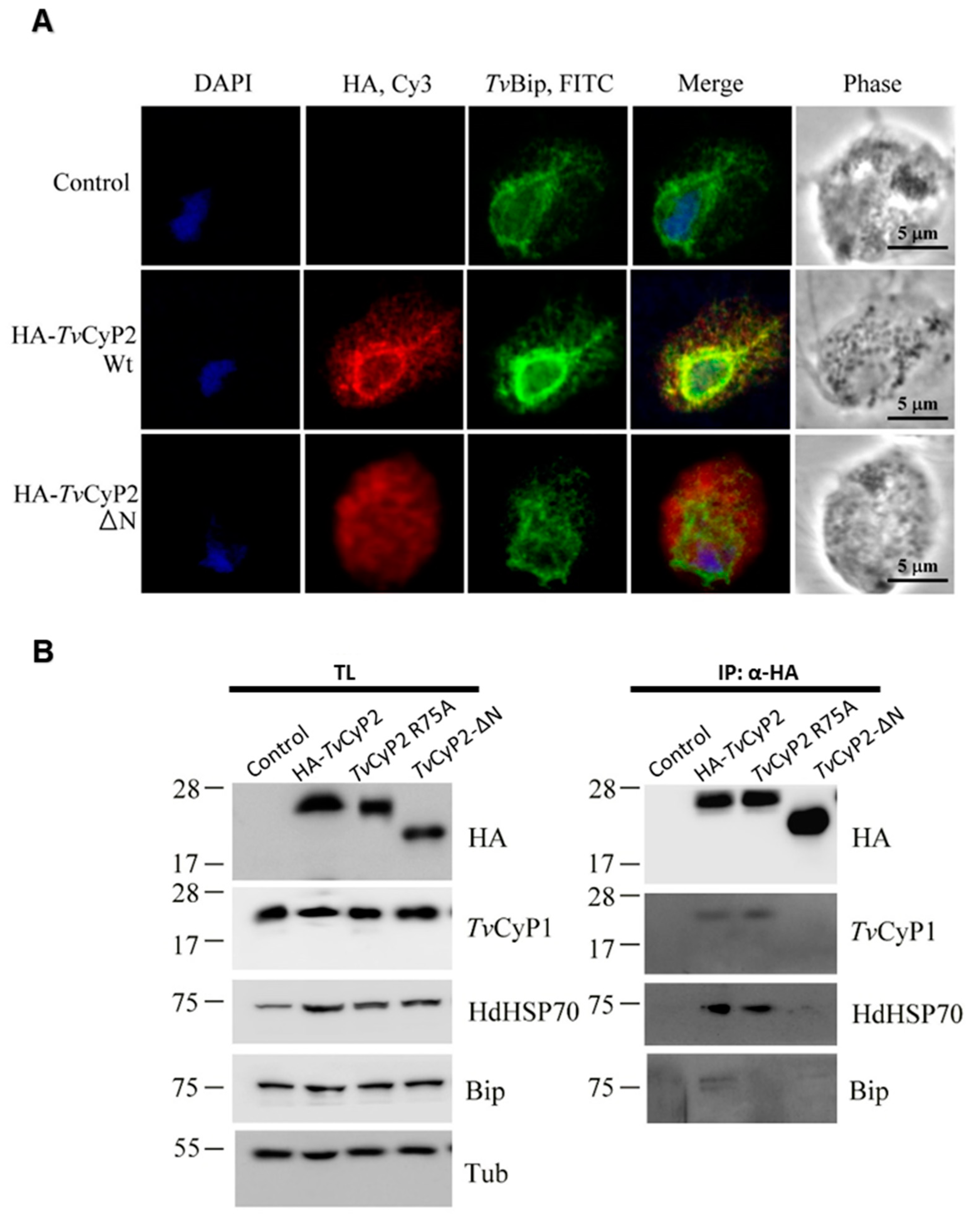
3.5. N-Terminal Segment Affects the Subcellular Localization and Formation of Differential Complexes in Cellular Processes
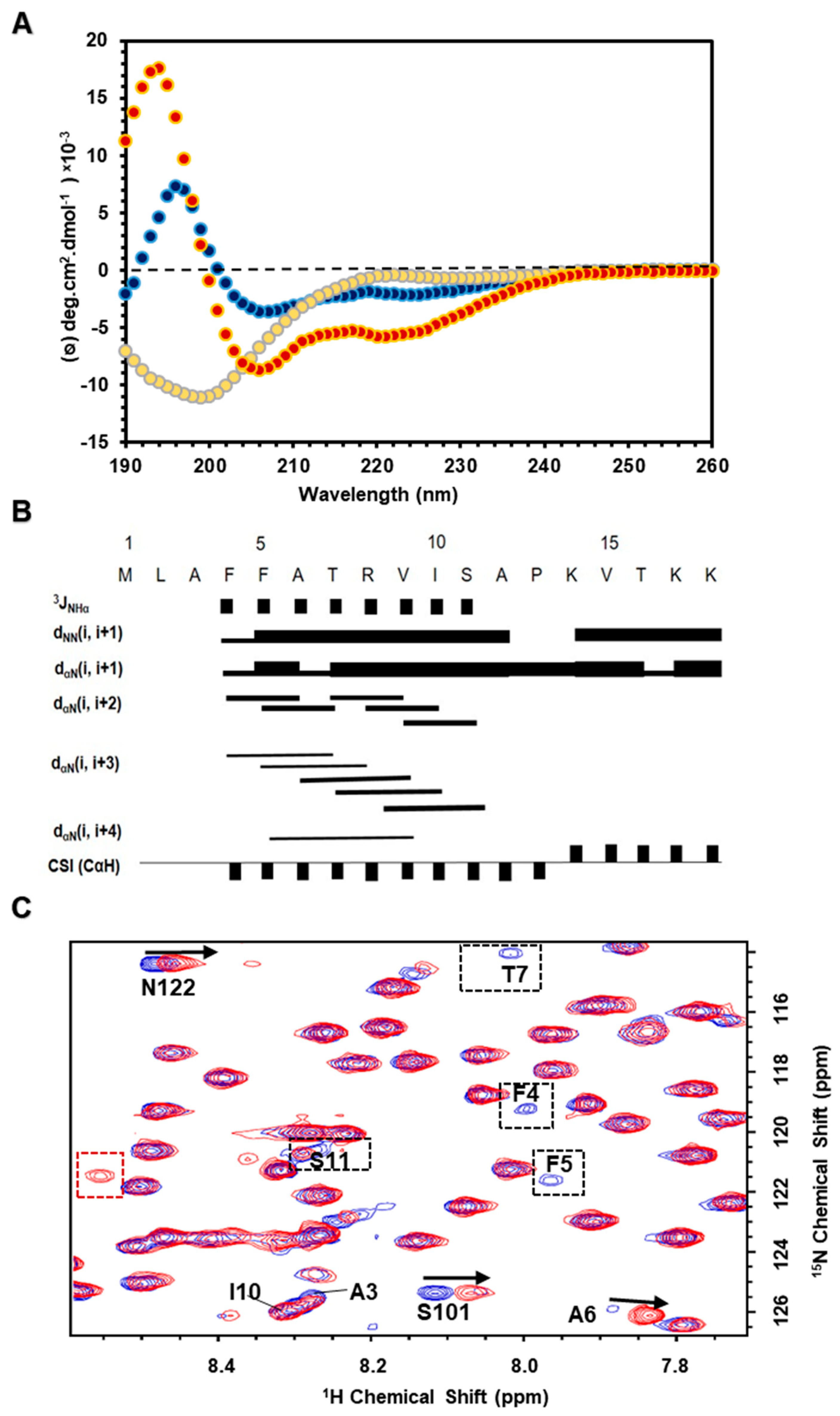
3.6. N-Terminal Segment in Membrane Mimicking Environments Forming an α-Helical Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, P.; Heitman, J. The cyclophilins. Genome Biol. 2005, 6, 226. [Google Scholar] [CrossRef] [Green Version]
- Andreotti, A.H. Native state proline isomerization: An intrinsic molecular switch. Biochemistry 2003, 42, 9515–9524. [Google Scholar] [CrossRef]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: A key player for human disease. Cell Death Dis. 2013, 4, e888. [Google Scholar] [CrossRef] [Green Version]
- Dawar, F.U.; Xiong, Y.; Khattak, M.N.K.; Li, J.; Lin, L.; Mei, J. Potential role of cyclophilin A in regulating cytokine secretion. J. Leukoc. Biol. 2017, 102, 989–992. [Google Scholar] [CrossRef]
- Galat, A. Peptidylprolyl cis/trans isomerases (immunophilins): Biological diversity-targets-functions. Curr. Top. Med. Chem. 2003, 3, 1315–1347. [Google Scholar] [CrossRef]
- Singh, K.; Winter, M.; Zouhar, M.; Rysanek, P. Cyclophilins: Less studied proteins with critical roles in pathogenesis. Phytopathology 2018, 108, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Ryffel, B.; Woerly, G.; Greiner, B.; Haendler, B.; Mihatsch, M.J.; Foxwell, B.M. Distribution of the cyclosporine binding protein cyclophilin in human tissues. Immunology 1991, 72, 399–404. [Google Scholar] [PubMed]
- Sherry, B.; Yarlett, N.; Strupp, A.; Cerami, A. Identification of cyclophilin as a proinflammatory secretory product of lipopolysaccharide-activated macrophages. Proc. Natl. Acad. Sci. USA 1992, 89, 3511–3515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikol, V.; Kallen, J.; Pflugl, G.; Walkinshaw, M.D. X-ray structure of a monomeric cyclophilin A-cyclosporin A crystal complex at 2.1 A resolution. J. Mol. Biol. 1993, 234, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.R.; Hall, D.R.; Berriman, M.; Nunes, J.A.; Leonard, G.A.; Fairlamb, A.H.; Hunter, W.N. The three-dimensional structure of a plasmodium falciparum cyclophilin in complex with the potent anti-malarial cyclosporin A. J. Mol. Biol. 2000, 298, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Huai, Q.; Kim, H.Y.; Liu, Y.; Zhao, Y.; Mondragon, A.; Liu, J.O.; Ke, H. Crystal structure of calcineurin-cyclophilin-cyclosporin shows common but distinct recognition of immunophilin-drug complexes. Proc. Natl. Acad. Sci. USA 2002, 99, 12037–12042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Ferreira, P.A.; Orry, A. From drosophila to humans: Reflections on the roles of the prolyl isomerases and chaperones, cyclophilins, in cell function and disease. J. Neurogenet. 2012, 26, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.; Luo, C.; Hogan, P.G. Transcription factors of the NFAT family: Regulation and function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef]
- Chen, Z.; Mi, L.; Xu, J.; Yu, J.; Wang, X.; Jiang, J.; Xing, J.; Shang, P.; Qian, A.; Li, Y.; et al. Function of HAb18G/CD147 in invasion of host cells by severe acute respiratory syndrome coronavirus. J. Infect. Dis. 2005, 191, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajo-Lozoya, J.; Ma-Lauer, Y.; Malesevic, M.; Theuerkorn, M.; Kahlert, V.; Prell, E.; von Brunn, B.; Muth, D.; Baumert, T.F.; Drosten, C.; et al. Human coronavirus NL63 replication is cyclophilin A-dependent and inhibited by non-immunosuppressive cyclosporine A-derivatives including alisporivir. Virus Res. 2014, 184, 44–53. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sato, Y.; Sasaki, T. Suppression of coronavirus replication by cyclophilin inhibitors. Viruses 2013, 5, 1250–1260. [Google Scholar] [CrossRef] [Green Version]
- Chopard, C.; Tong, P.B.V.; Toth, P.; Schatz, M.; Yezid, H.; Debaisieux, S.; Mettling, C.; Gross, A.; Pugniere, M.; Tu, A.; et al. Cyclophilin A enables specific HIV-1 Tat palmitoylation and accumulation in uninfected cells. Nat. Commun. 2018, 9, 2251. [Google Scholar] [CrossRef]
- Liu, C.; Perilla, J.R.; Ning, J.; Lu, M.; Hou, G.; Ramalho, R.; Himes, B.A.; Zhao, G.; Bedwell, G.J.; Byeon, I.J.; et al. Cyclophilin A stabilizes the HIV-1 capsid through a novel non-canonical binding site. Nat. Commun. 2016, 7, 10714. [Google Scholar] [CrossRef] [Green Version]
- Watashi, K.; Shimotohno, K. Cyclophilin and viruses: Cyclophilin as a cofactor for viral infection and possible anti-viral target. Drug Target Insights 2007, 2, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Johns Hopkins University. Coronavirus Resource Center. Available online: https://coronavirus.jhu.edu/ (accessed on 30 July 2020).
- Mitteregger, D.; Aberle, S.W.; Makristathis, A.; Walochnik, J.; Brozek, W.; Marberger, M.; Kramer, G. High detection rate of Trichomonas vaginalis in benign hyperplastic prostatic tissue. Med. Microbiol. Immunol. 2012, 201, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.C.; Mthiyane, T.N.; Baisley, K.; McHunu, S.L.; Ferguson, J.B.; Smit, T.; Crucitti, T.; Gareta, D.; Dlamini, S.; Mutevedzi, T.; et al. Prevalence of sexually transmitted infections among young people in South Africa: A nested survey in a health and demographic surveillance site. PLoS Med. 2018, 15, e1002512. [Google Scholar] [CrossRef] [PubMed]
- Upcroft, P.; Upcroft, J.A. Drug targets and mechanisms of resistance in the anaerobic protozoa. Clin. Microbiol. Rev. 2001, 14, 150–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, I.; Muwonge, R.; Mittal, S.; Banerjee, D.; Kundu, P.; Mandal, R.; Biswas, J.; Basu, P. Association between high risk human papillomavirus infection and co-infection with candida spp. and Trichomonas vaginalis in women with cervical premalignant and malignant lesions. J. Clin. Virol. 2017, 87, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Gimenes, F.; Souza, R.P.; Bento, J.C.; Teixeira, J.J.; Maria-Engler, S.S.; Bonini, M.G.; Consolaro, M.E. Male infertility: A public health issue caused by sexually transmitted pathogens. Nat. Rev. Urol. 2014, 11, 672–687. [Google Scholar] [CrossRef] [PubMed]
- McClelland, R.S.; Sangare, L.; Hassan, W.M.; Lavreys, L.; Mandaliya, K.; Kiarie, J.; Ndinya-Achola, J.; Jaoko, W.; Baeten, J.M. Infection with Trichomonas vaginalis increases the risk of HIV-1 acquisition. J. Infect. Dis. 2007, 195, 698–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, J.R.; Judson, G.; Alderete, J.F.; Mundodi, V.; Kucknoor, A.S.; Giovannucci, E.L.; Platz, E.A.; Sutcliffe, S.; Fall, K.; Kurth, T.; et al. Prospective study of Trichomonas vaginalis infection and prostate cancer incidence and mortality: Physicians’ health study. J. Natl. Cancer Inst. 2009, 101, 1406–1411. [Google Scholar] [CrossRef]
- Fichorova, R.N. Impact of T. vaginalis infection on innate immune responses and reproductive outcome. J. Reprod. Immunol. 2009, 83, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, O.; Rigo, G.V.; Macedo, A.J.; Tasca, T. Trichomonas vaginalis clinical isolates: Cytoadherence and adherence to polystyrene, intrauterine device, and vaginal ring. Parasitol. Res. 2017, 116, 3275–3284. [Google Scholar] [CrossRef]
- Hrdý, I.; Tachezy, J.; Müller, M. Metabolism of trichomonad hydrogenosomes. In Hydrogenosomes and Mitosomes: Mitochondria of Anaerobic Eukaryotes; Tachezy, J., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 127–158. [Google Scholar]
- Hsu, H.M.; Ong, S.J.; Lee, M.C.; Tai, J.H. Transcriptional regulation of an iron-inducible gene by differential and alternate promoter entries of multiple Myb proteins in the protozoan parasite Trichomonas vaginalis. Eukaryot. Cell 2009, 8, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.J.; Hsu, H.M.; Liu, H.W.; Chu, C.H.; Tai, J.H. Multifarious transcriptional regulation of adhesion protein gene ap65-1 by a novel Myb1 protein in the protozoan parasite Trichomonas vaginalis. Eukaryot. Cell 2006, 5, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, S.J.; Hsu, H.M.; Liu, H.W.; Chu, C.H.; Tai, J.H. Activation of multifarious transcription of an adhesion protein ap65-1 gene by a novel Myb2 protein in the protozoan parasite Trichomonas vaginalis. J. Biol. Chem. 2007, 282, 6716–6725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, T.; Lou, Y.C.; Chou, C.C.; Wei, S.Y.; Sadotra, S.; Cho, C.C.; Lin, M.H.; Tai, J.H.; Hsu, C.H.; Chen, C. Structural basis of interaction between dimeric cyclophilin 1 and Myb1 transcription factor in Trichomonas vaginalis. Sci. Rep. 2018, 8, 5410. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.M.; Huang, Y.H.; Aryal, S.; Liu, H.W.; Chen, C.; Chen, S.H.; Chu, C.H.; Tai, J.H. Endomembrane protein trafficking regulated by a TvCyP2 cyclophilin in the protozoan parasite, Trichomonas vaginalis. Sci. Rep. 2020, 10, 1275. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.Y.; Lou, Y.C.; Tsai, J.Y.; Ho, M.R.; Chou, C.C.; Rajasekaran, M.; Hsu, H.M.; Tai, J.H.; Hsiao, C.D.; Chen, C. Structure of the Trichomonas vaginalis Myb3 DNA-binding domain bound to a promoter sequence reveals a unique C-terminal beta-hairpin conformation. Nucleic Acids Res. 2012, 40, 449–460. [Google Scholar] [CrossRef]
- McPherson, A. Current approaches to macromolecular crystallization. Eur. J. Biochem. 1990, 189, 1–23. [Google Scholar] [CrossRef]
- Moreno, A. Advanced methods of protein crystallization. Methods Mol. Biol. 2017, 1607, 51–76. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. [20] Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- Lawrence, M.C. The application of the molecular replacement method to the de novo determination of protein structure. Q. Rev. Biophys. 1991, 24, 399–424. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66 Pt 4, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Zwart, P.H.; Afonine, P.V.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; McKee, E.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; et al. Automated structure solution with the PHENIX suite. Methods Mol. Biol. 2008, 426, 419–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66 Pt 2, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., III; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, T.; Lou, Y.C.; Aryal, S.; Tai, J.H.; Chen, C. (1)H, (13)C and (15)N resonance assignments and secondary structures of cyclophilin 2 from Trichomonas vaginalis. Biomol. NMR Assign. 2018, 12, 27–30. [Google Scholar] [CrossRef]
- Johnson, B.A.; Blevins, R.A. NMR view: A computer program for the visualization and analysis of NMR data. J. Biomol. NMR 1994, 4, 603–614. [Google Scholar] [CrossRef]
- Hsu, H.M.; Chu, C.H.; Wang, Y.T.; Lee, Y.; Wei, S.Y.; Liu, H.W.; Ong, S.J.; Chen, C.; Tai, J.H. Regulation of nuclear translocation of the Myb1 transcription factor by TvCyclophilin 1 in the protozoan parasite Trichomonas vaginalis. J. Biol. Chem. 2014, 289, 19120–19136. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.H.; Huang, Y.H.; Liu, H.W.; Hsu, H.M.; Tai, J.H. Membrane localization of a Myb3 transcription factor regulated by a TvCyP1 cyclophilin in the parasitic protozoan Trichomonas vaginalis. FEBS J. 2018, 285, 929–946. [Google Scholar] [CrossRef] [Green Version]
- Wexler-Cohen, Y.; Stevens, G.C.; Barnoy, E.; van der Bliek, A.M.; Johnson, P.J. A dynamin-related protein contributes to Trichomonas vaginalis hydrogenosomal fission. FASEB J. 2014, 28, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.; Page, A.P.; Kontopidis, G.; Husi, H.; Walkinshaw, M.D. The X-ray structure of a divergent cyclophilin from the nematode parasite Brugia malayi. FEBS Lett. 1998, 425, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.L.; Walker, J.R.; Ouyang, H.; MacKenzie, F.; Butler-Cole, C.; Newman, E.M.; Eisenmesser, E.Z.; Dhe-Paganon, S. The crystal structure of human WD40 repeat-containing peptidylprolyl isomerase (PPWD1). FEBS J. 2008, 275, 2283–2295. [Google Scholar] [CrossRef] [PubMed]
- Reidt, U.; Wahl, M.C.; Fasshauer, D.; Horowitz, D.S.; Luhrmann, R.; Ficner, R. Crystal structure of a complex between human spliceosomal cyclophilin H and a U4/U6 snRNP-60K peptide. J. Mol. Biol. 2003, 331, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Rajiv, C.; Davis, T.L. Structural and functional insights into human nuclear cyclophilins. Biomolecules 2018, 8, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallen, J.; Mikol, V.; Taylor, P.; Walkinshaw, M.D. X-ray structures and analysis of 11 cyclosporin derivatives complexed with cyclophilin A. J. Mol. Biol. 1998, 283, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Ryffel, B.; Woerly, G.; Murray, M.; Eugster, H.P.; Car, B. Binding of active cyclosporins to cyclophilin A and B, complex formation with calcineurin A. Biochem. Biophys. Res. Commun. 1993, 194, 1074–1083. [Google Scholar] [CrossRef]
- Furst, M.; Zhou, Y.; Merfort, J.; Muller, M. Involvement of PpiD in sec-dependent protein translocation. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 273–280. [Google Scholar] [CrossRef]
- Mentel, M.; Zimorski, V.; Haferkamp, P.; Martin, W.; Henze, K. Protein import into hydrogenosomes of Trichomonas vaginalis involves both N-terminal and internal targeting signals: A case study of thioredoxin reductases. Eukaryot. Cell 2008, 7, 1750–1757. [Google Scholar] [CrossRef] [Green Version]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Hrdý, I.; Müller, M. Primary structure and eubacterial relationships of the pyruvate:ferredoxin oxidoreductase of the amitochondriate eukaryote Trichomonas vaginalis. J. Mol. Evol. 1995, 41, 388–396. [Google Scholar] [CrossRef]
- Bradley, P.J.; Lahti, C.J.; Plümper, E.; Johnson, P.J. Targeting and translocation of proteins into the hydrogenosome of the protist Trichomonas: Similarities with mitochondrial protein import. EMBO J. 1997, 16, 3484–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | ||||
| Crystal | TvCyP2_apo1 | TvCyP2_apo2 | TvCyP2_apo3 | TvCyP2_apo4 |
| Space group | P212121 | P212121 | P212121 | P212121 |
| Cell dimensions | ||||
| a, b, c (Å) | 50.8, 56.1, 59.8 | 53.8, 54.2, 59.8 | 54.3, 54.6, 59.9 | 52.0, 54.5, 59.6 |
| α,β,γ (°) | 90.0, 90.0, 90.0 | 90.0, 90.0, 90.0 | 90.0, 90.0, 90.0 | 90.0, 90.0, 90.0 |
| Resolution (Å) | 28.09–1.89 (1.95–1.89) | 27.13–2.35 (2.44–2.35) | 27.15–1.85 (1.92–1.85) | 26.17–2.56 (2.65–2.56) |
| Rmergea,b | 0.039 (0.135) | 0.070 (0.327) | 0.055 (0.350) | 0.077 (0.405) |
| I/σ (I)a | 33.8 (10.6) | 16.8 (2.8) | 23.9 (3.1) | 18.5 (3.2) |
| Completeness (%) a | 98.2 (90.9) | 91.4 (91.8) | 99.7 (97.8) | 99.9 (100.0) |
| Redundancy a | 4.7 (4.7) | 4.5 (3.5) | 4.6 (4.3) | 6.6 (5.8) |
| CC1/2 a,c | 0.993 (0.979) | 0.970 (0.919) | 0.981 (0.924) | 0.978 (0.918) |
| CC * | 0.998 (0.995) | 0.992 (0.979) | 0.995 (0.980) | 0.994 (0.978) |
| Refinement | ||||
| Resolution (Å) | 28.09–1.89 | 27.13–2.35 | 27.15–1.85 | 26.17–2.56 |
| No. of reflections | 14,047 | 6685 | 14941 | 5340 |
| Rwork/Rfreed | 0.1394/0.1785 | 0.1551/0.2254 | 0.1463/0.1821 | 0.1677/0.2308 |
| No. of atoms | ||||
| Protein | 1350 | 1363 | 1374 | 1366 |
| Ligand/ion | 42 | 61 | ||
| Water | 196 | 26 | 106 | 22 |
| B-factor | ||||
| Protein | 18.91 | 32.78 | 21.67 | 32.85 |
| Ligand/ion | 63.93 | 64.08 | ||
| Water | 32.60 | 35.07 | 35.41 | 31.99 |
| R.m.s. deviation | ||||
| Bond lengths (Å) | 0.007 | 0.008 | 0.007 | 0.008 |
| Bond angles (°) | 1.19 | 1.28 | 1.26 | 1.25 |
| Ramachandran plot e | ||||
| Favoured (%) | 96.0 | 94.4 | 96.6 | 95.5 |
| Allowed (%) | 3.9 | 5.0 | 3.3 | 4.4 |
| Outliers (%) | 0 | 0.5 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aryal, S.; Hsu, H.-M.; Lou, Y.-C.; Chu, C.-H.; Tai, J.-H.; Hsu, C.-H.; Chen, C. N-Terminal Segment of TvCyP2 Cyclophilin from Trichomonas vaginalis Is Involved in Self-Association, Membrane Interaction, and Subcellular Localization. Biomolecules 2020, 10, 1239. https://doi.org/10.3390/biom10091239
Aryal S, Hsu H-M, Lou Y-C, Chu C-H, Tai J-H, Hsu C-H, Chen C. N-Terminal Segment of TvCyP2 Cyclophilin from Trichomonas vaginalis Is Involved in Self-Association, Membrane Interaction, and Subcellular Localization. Biomolecules. 2020; 10(9):1239. https://doi.org/10.3390/biom10091239
Chicago/Turabian StyleAryal, Sarita, Hong-Ming Hsu, Yuan-Chao Lou, Chien-Hsin Chu, Jung-Hsiang Tai, Chun-Hua Hsu, and Chinpan Chen. 2020. "N-Terminal Segment of TvCyP2 Cyclophilin from Trichomonas vaginalis Is Involved in Self-Association, Membrane Interaction, and Subcellular Localization" Biomolecules 10, no. 9: 1239. https://doi.org/10.3390/biom10091239