Biological Properties of a Novel Multifunctional Host Defense Peptide from the Skin Secretion of the Chaco Tree Frog, Boana raniceps

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Collection of the Specimens and Skin Secretion Harvesting
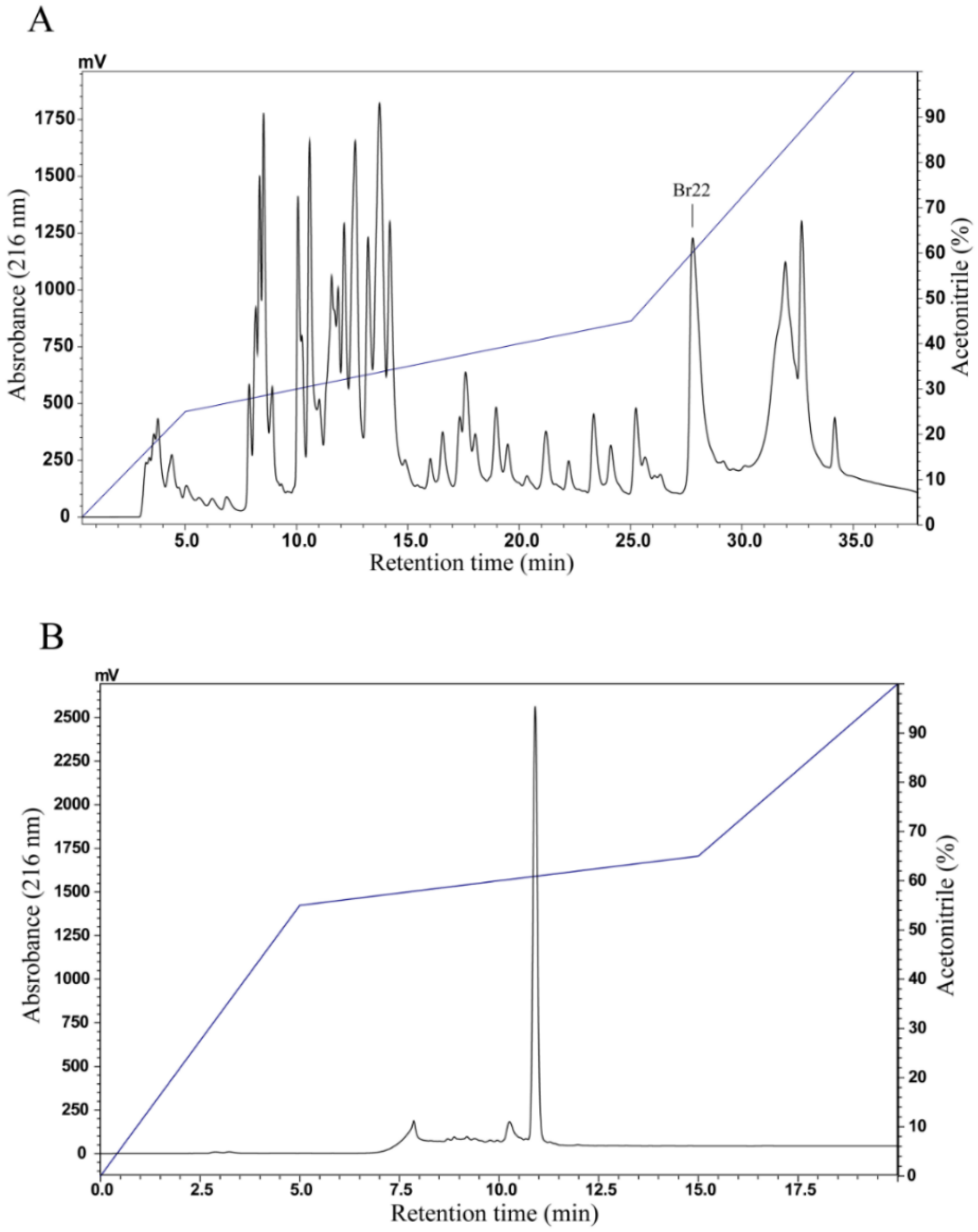
2.2. Peptide Purification
2.3. Peptide Quantification
2.4. Structural Analysis
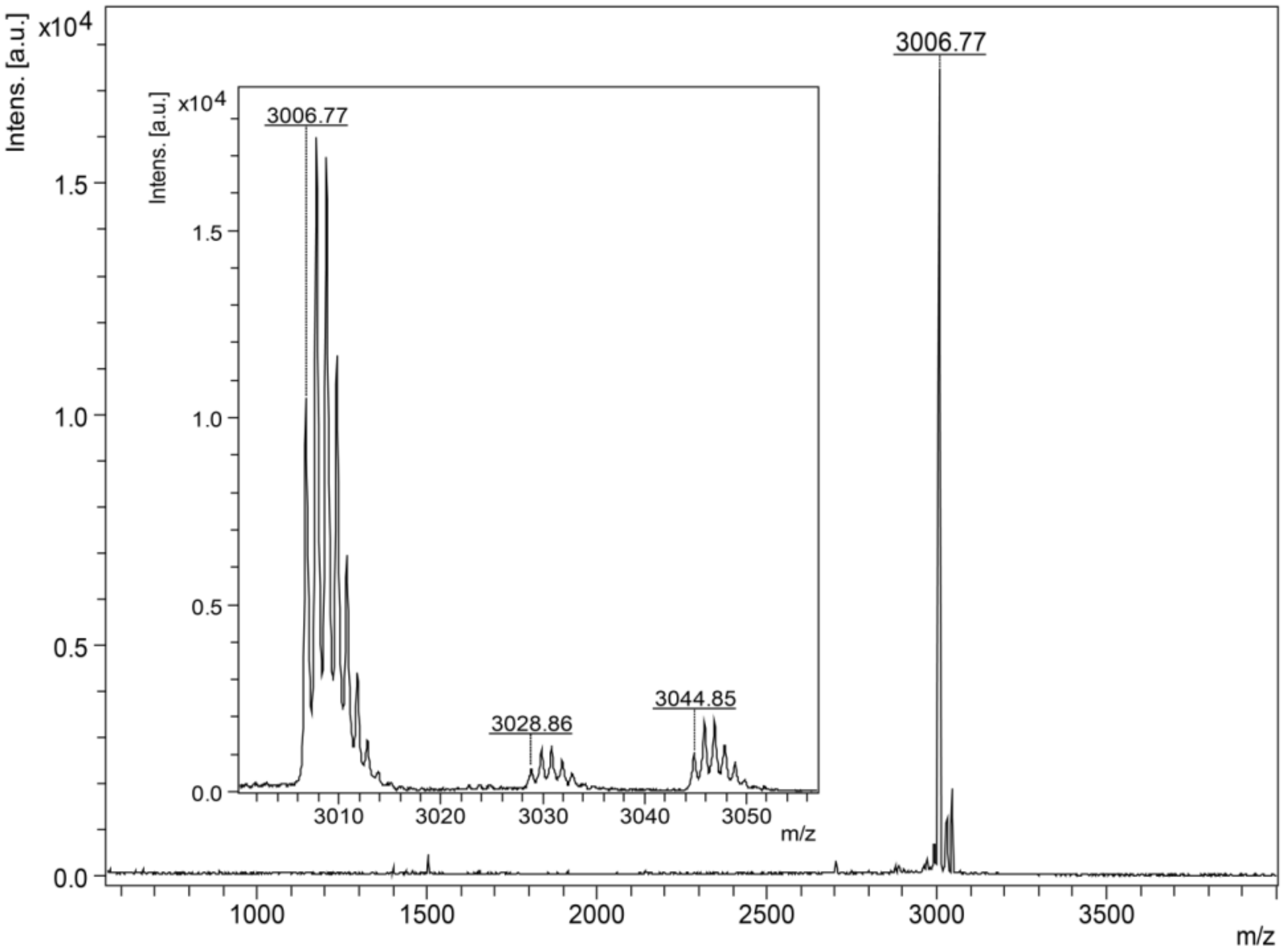
2.4.1. Mass Spectrometry Analysis and Edman Degradation
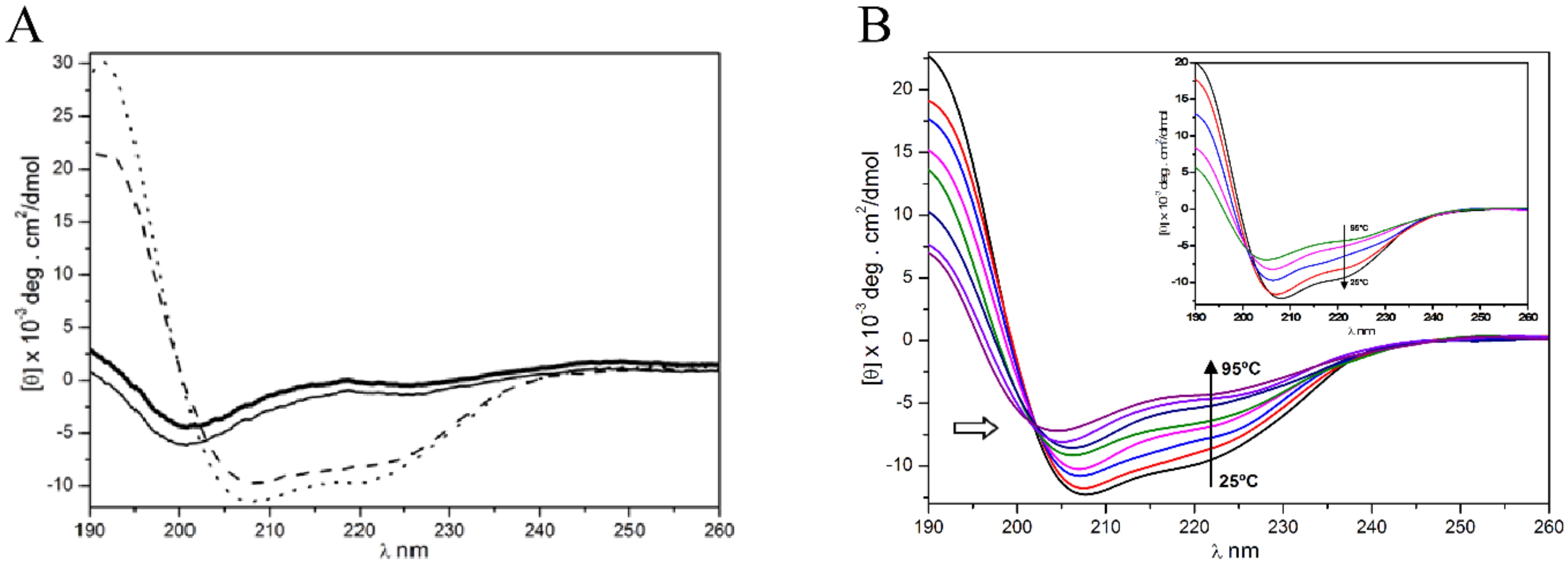
2.4.2. Circular Dichroism Analysis
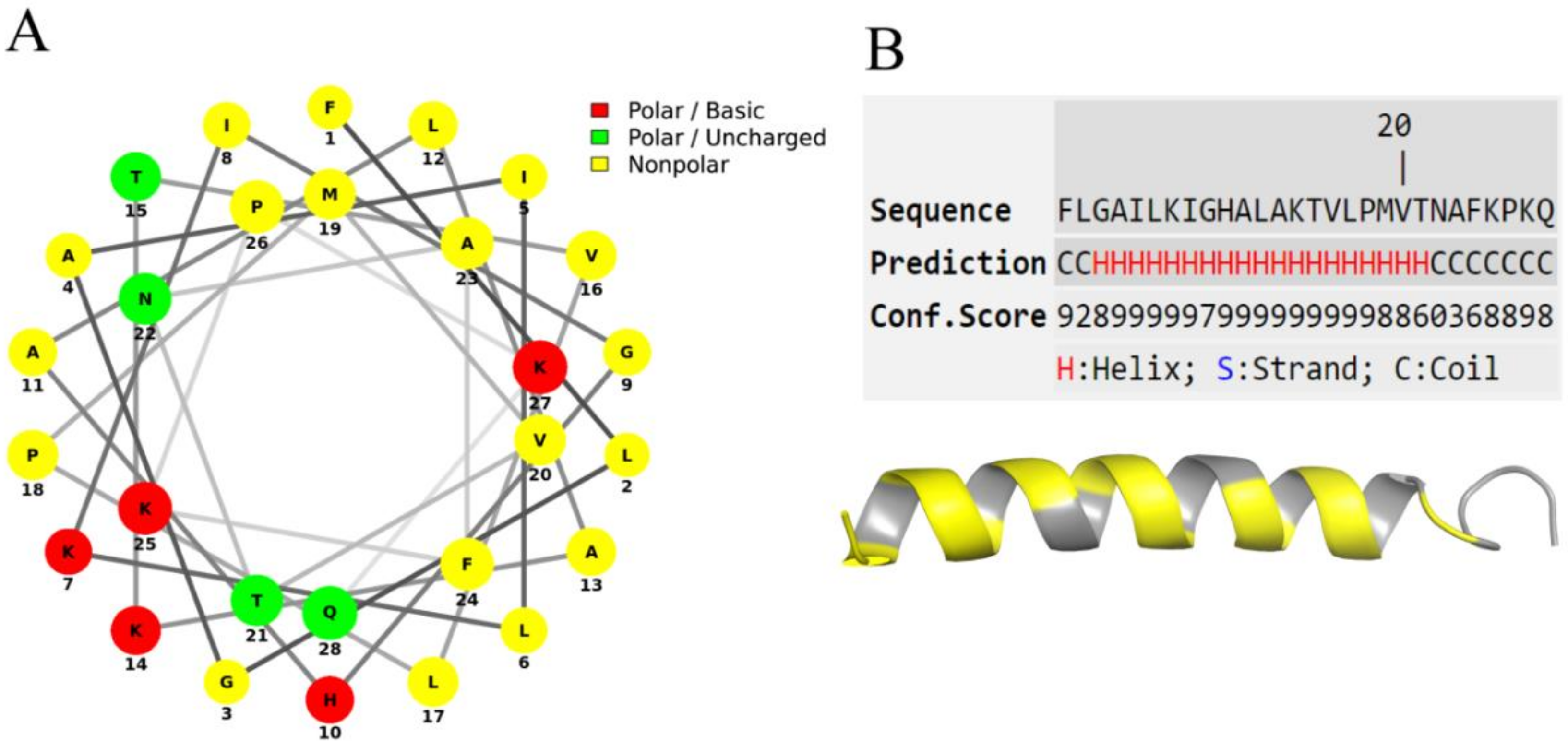
2.4.3. Bioinformatics Analysis
2.5. Antimicrobial Assays
2.6. Anti-Epimastigote Activity Against Trypanosoma cruzi
2.7. Hemolytic Assay
2.8. Cytotoxicity Assay
2.9. Antiviral Assays
2.9.1. Chikungunya Virus Assay
2.9.2. Dengue Serotype 4 Virus Assay (DENV4)
2.9.3. Yellow Fever Virus Assay
2.10. Neutrophil Phagocytosis and Oxidative Burst
3. Results and Discussion
3.1. Purification Procedure and Mass Spectrometry Analysis
3.2. Sequence Analysis
3.3. Secondary Structure Prediction and Circular Dichroism Analysis
3.4. Antimicrobial Activity
3.5. Antitrypanosomal Activity
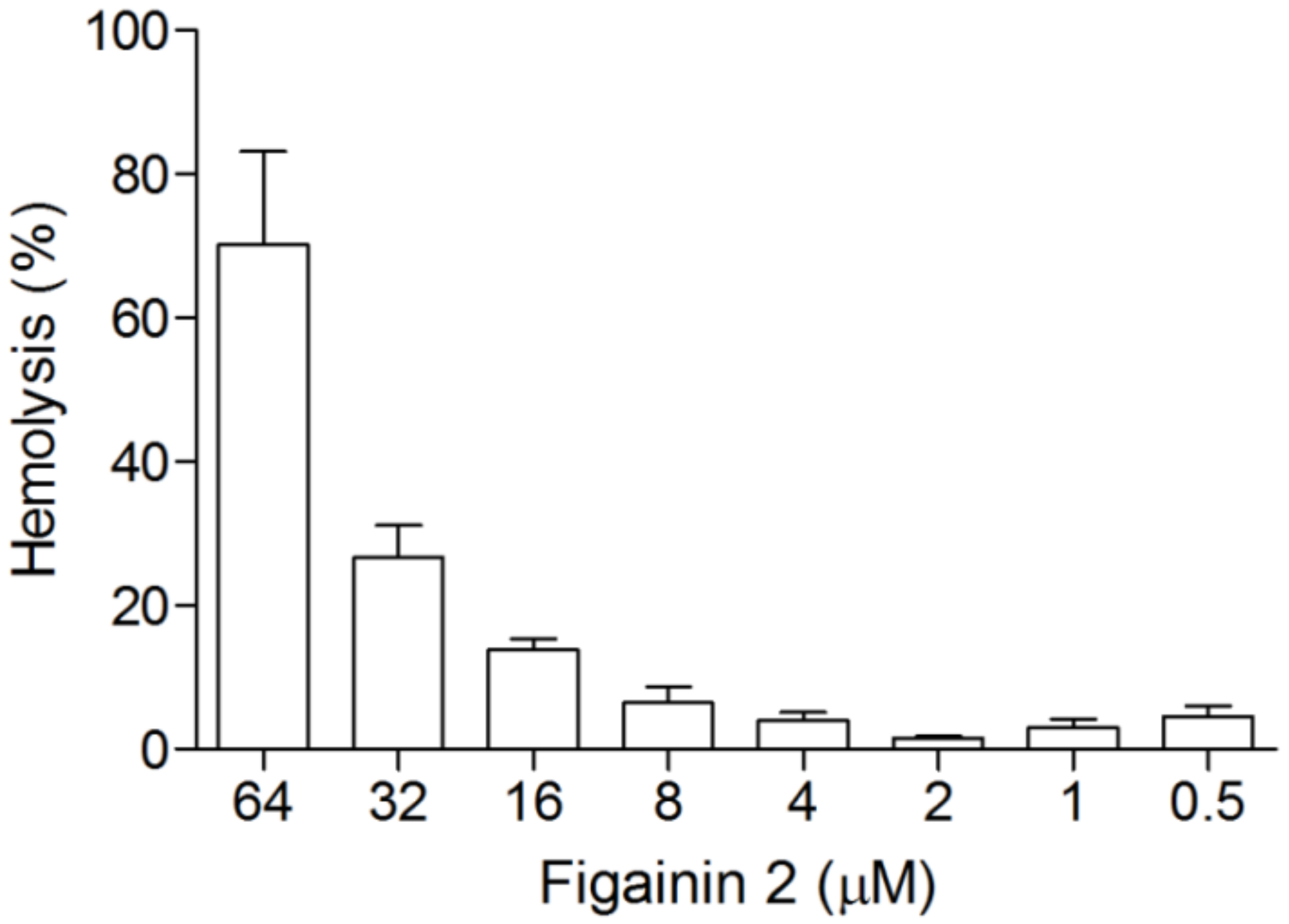
3.6. Hemolytic Activity
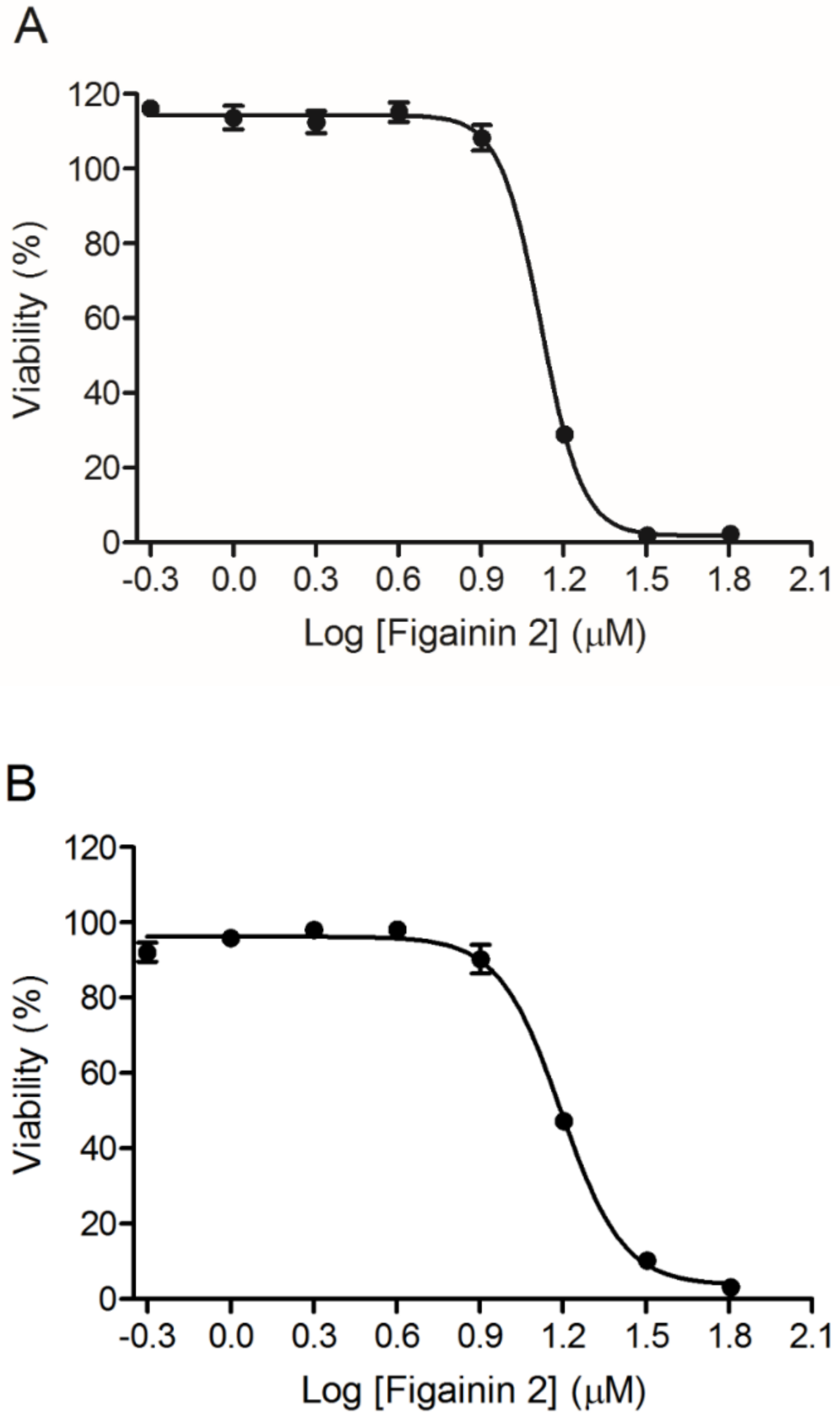
3.7. Antiproliferative Activity
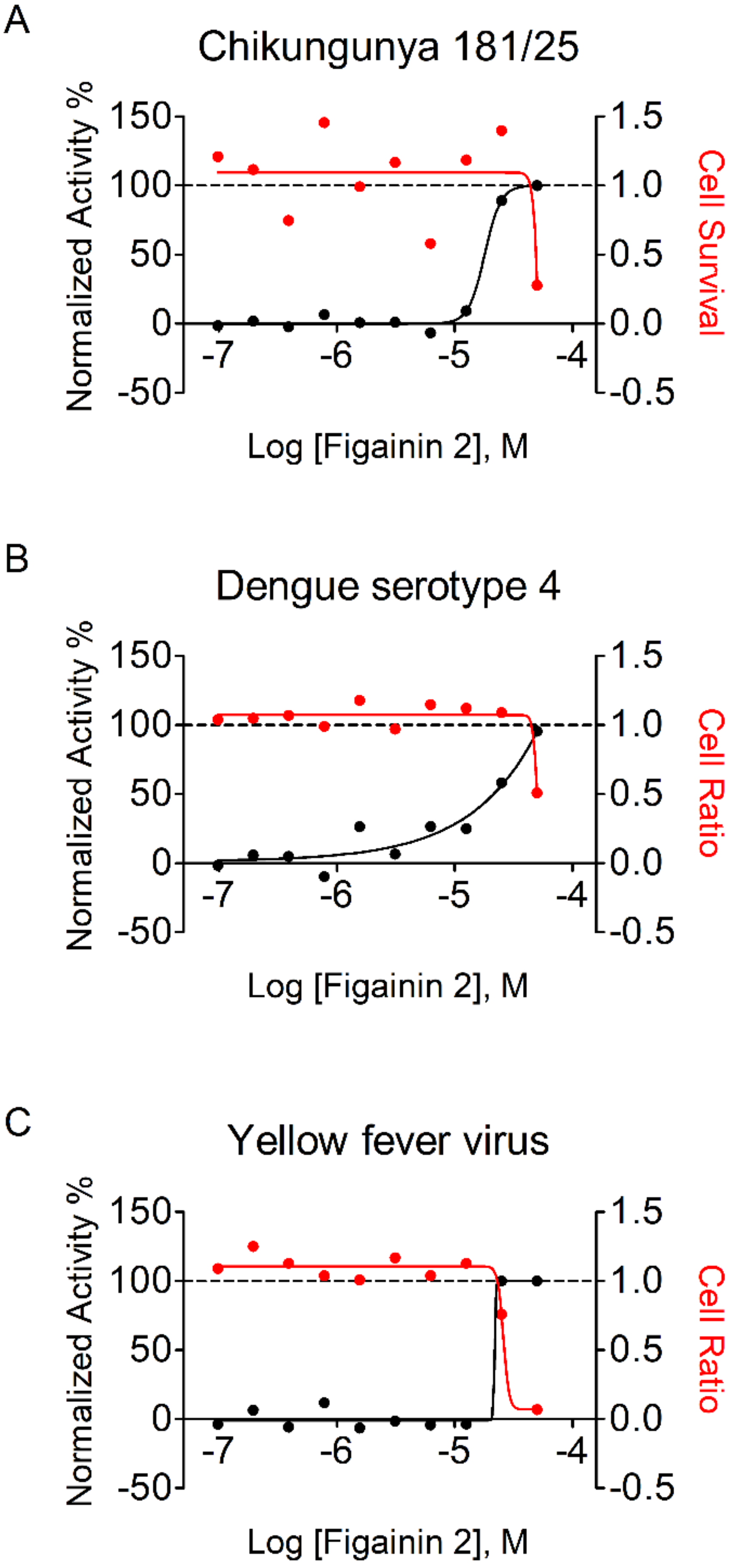
3.8. Antiviral Properties
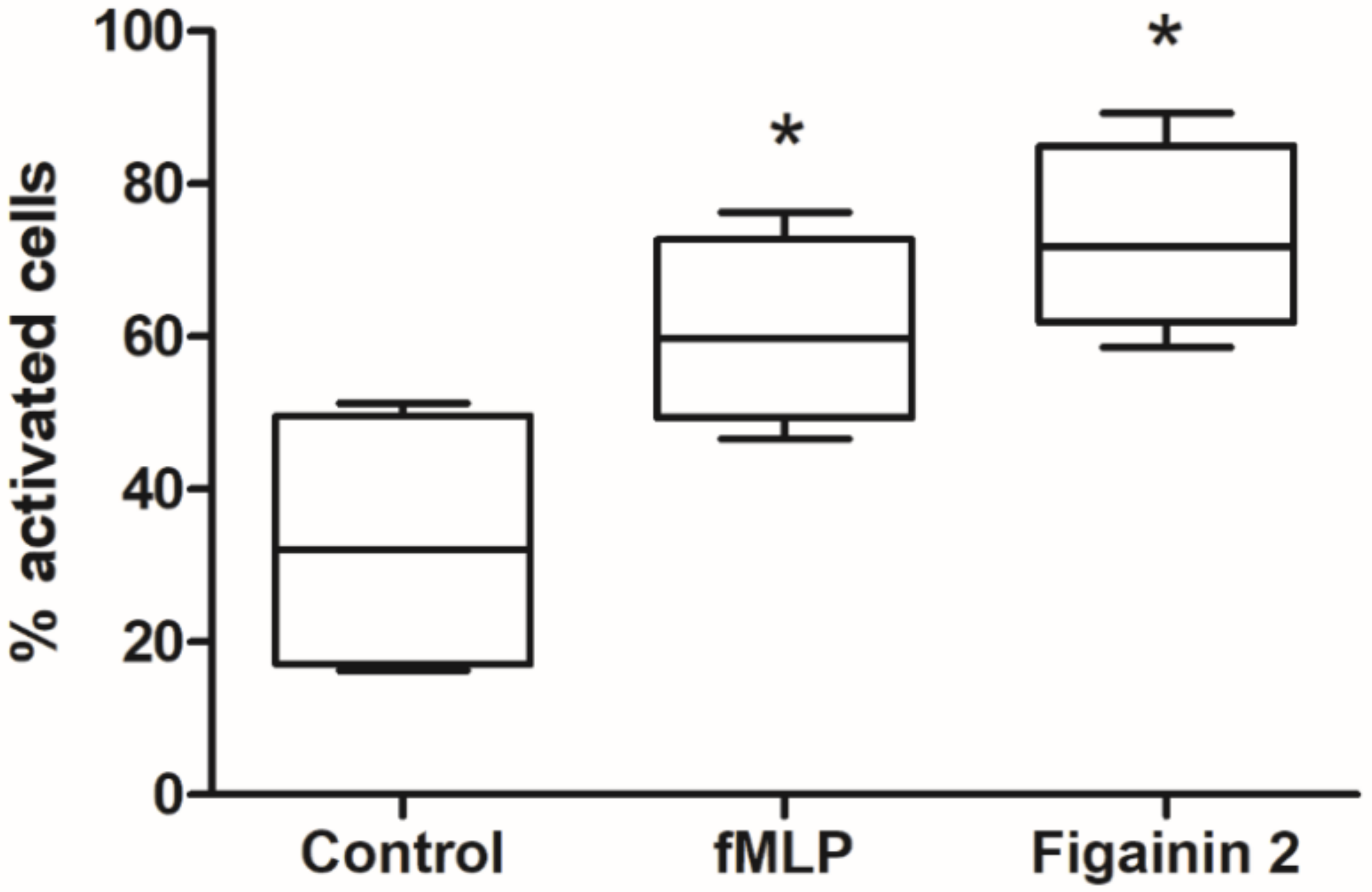
3.9. Modulatory Activity on the Innate Immune System
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hede, K. Antibiotic resistance: An infectious arms race. Nature 2014, 509, S2–S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heure, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. N. Microbes N. Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laxminarayan, R.; Van Boeckel, T.; Frost, I.; Kariuki, S.; Khan, E.A.; Limmathurotsakul, D.; Larsson, D.G.J.; Levy-Hara, G.; Mendelson, M.; Outterson, K.; et al. The Lancet Infectious Diseases Commission on antimicrobial resistance: 6 years later. Lancet Infect. Dis. 2020, 1–10. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance—The need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- Laxminarayan, R.; Matsoso, P.; Pant, S.; Brower, C.; Røttingen, J.-A.; Klugman, K.; Davies, S. Access to effective antimicrobials: A worldwide challenge. Lancet 2016, 387, 168–175. [Google Scholar] [CrossRef]
- Spellberg, B.; Bartlett, J.G.; Gilbert, D.N. The future of antibiotics and resistance. N. Engl. J. Med. 2013, 368, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.-H. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn, I.; Feudi, C.; Wang, Y.; Kaspar, H.; Feßler, A.T.; Lübke-Becker, A.; Michael, G.B.; Shen, J.; Schwarz, S. Identification of novel variants of the colistin resistance gene mcr-3 in Aeromonas spp. from the national resistance monitoring programme GERM-Vet and from diagnostic submissions. J. Antimicrob. Chemother. 2018, 73, 1217–1221. [Google Scholar] [CrossRef]
- Hayashi, W.; Togashi, M.; Taniguchi, Y.; Koide, S.; Nagano, Y.; Nagano, N. First report of colistin resistance in OXA-181 carbapenemase-producing Klebsiella pneumoniae ST3130 in Japan. J. Glob. Antimicrob. Resist. 2018, 12, 179–180. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid. Res. 2012, 51, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Why and how are peptide–lipid interactions utilized for self-defense? Magainins and tachyplesins as archetypes. Biochim. Biophys. Acta Biomembr. 1999, 1462, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.B.; Costa, F.J.; Pires, O.R.; Fontes, W.; Castro, M.S. The amazing world of peptide engineering: The example of antimicrobial peptides from frogs and their analogues. Protein Pept. Lett. 2016, 23, 722–737. [Google Scholar] [CrossRef]
- Huang, Y.; Feng, Q.; Yan, Q.; Hao, X.; Chen, Y. Alpha-helical cationic anticancer peptides: A promising candidate for novel anticancer drugs. Mini Rev. Med. Chem. 2015, 15, 73–81. [Google Scholar] [CrossRef]
- Shartouny, J.R.; Jacob, J. Mining the tree of life: Host defense peptides as antiviral therapeutics. Semin. Cell. Dev. Biol. 2019, 88, 147–155. [Google Scholar] [CrossRef]
- Aitken, A.; Learmonth, M.P. Protein determination by UV absorption. In The Protein Protocols Handbook, 2nd ed.; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2002; pp. 3–6. [Google Scholar]
- Greenfield, N.J.; Fasman, G.D. Computed circular dichroism spectra for the evaluation of protein conformation. Biochemistry 1969, 8, 4108–4116. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bjellqvist, B.; Hughes, G.J.; Pasquali, C.; Paquet, N.; Ravier, F.; Sanchez, J.C.; Frutiger, S.; Hochstrasser, D. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis 1993, 14, 1023–1031. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Mol, A.R.; Castro, M.S.; Fontes, W. NetWheels: A web application to create high quality peptide helical wheel and net projections. BioRxiv 2018, 416347. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2015, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libério, M.S.; Joanitti, G.A.; Azevedo, R.B.; Cilli, E.M.; Zanotta, L.C.; Nascimento, A.C.; Sousa, M.V.; Júnior, O.R.P.; Fontes, W.; Castro, M.S. Anti-proliferative and cytotoxic activity of pentadactylin isolated from Leptodactylus labyrinthicus on melanoma cells. Amino Acids 2011, 40, 51–59. [Google Scholar] [CrossRef]
- Pascoalino, B.S.; Courtemanche, G.; Cordeiro, M.T.; Gil, L.H.V.G.; Freitas-Junior, L. Zika antiviral chemotherapy: Identification of drugs and promising starting points for drug discovery from an FDA-approved library. F1000Res 2016, 5, 2523. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, E.A.; Campos, P.F.; Andrade, A.C.; Bloch, C. Response of Hypsiboas raniceps to Abiotic and Biotic Stresses: Gene Expression and MALDI-Mass Spectrometry Imaging Analysis of Skin Peptides. Unpublished. 2017. Available online: https://www.ncbi.nlm.nih.gov/protein/AVG44203?report=genbank&log$=protalign&blast_rank=1&RID=A824TTER014 (accessed on 16 September 2019).
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Farias, S.E.; Kline, K.G.; Klepacki, J.; Wu, C.C. Quantitative improvements in peptide recovery at elevated chromatographic temperatures from microcapillary liquid chromatography−mass spectrometry analyses of brain using selected reaction monitoring. Anal. Chem. 2010, 82, 3435–3440. [Google Scholar] [CrossRef] [Green Version]
- Wang, G. Prediction and design of antimicrobial peptides: Methods and applications to genomes and proteomes. In Antimicrobial peptides: Discovery, Design and Novel Therapeutic Strategies, 2nd ed.; Wang, G., Ed.; Cabi: Wallingford, UK, 2017; p. 288. [Google Scholar]
- Phoenix, D.A.; Dennison, S.R.; Harris, F. Cationic antimicrobial peptides. In Antimicrobial Peptides; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013. [Google Scholar]
- Johnson, W.C., Jr. Secondary structure of proteins through circular dichroism spectroscopy. Annu. Rev. Biophys. Biophys. Chem. 1988, 17, 145–166. [Google Scholar] [CrossRef]
- Gans, P.J.; Lyu, P.C.; Manning, M.C.; Woody, R.W.; Kallenbach, N.R. The helix–coil transition in heterogeneous peptides with specific side-chain interactions: Theory and comparison with CD spectral data. Biopolymers 1991, 31, 1605–1614. [Google Scholar] [CrossRef]
- Holtzer, M.E.; Holtzer, A. Alpha-helix to random coil transitions: Determination of peptide concentration from the CD at the isodichroic point. Biopolymers 1992, 32, 1675–1677. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Ferreira, T.C.G.; Cilli, E.M.; Crusca, E., Jr.; Mendes-Giannini, M.J.S.; Sebben, A.; Ricart, C.A.O.; Sousa, M.V.; Fontes, W. Hylin a1, the first cytolytic peptide isolated from the arboreal South American frog Hypsiboas albopunctatus (“spotted treefrog”). Peptides 2009, 30, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Nacif-Marçal, L.; Pereira, G.R.; Abranches, M.V.; Costa, N.C.S.; Cardoso, S.A.; Honda, E.R.; de Paula, S.O.; Feio, R.N.; Oliveira, L.L. Identification and characterization of an antimicrobial peptide of Hypsiboas semilineatus (Spix, 1824) (Amphibia, Hylidae). Toxicon 2015, 99, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, B.S.; Melo, J.A.; Leite, J.R.S.; Silva, L.P.; Prates, M.V.; Vinecky, F.; Barbosa, E.A.; Verly, R.M.; Mehta, A.; Nicoli, J.R. Post-secretory events alter the peptide content of the skin secretion of Hypsiboas raniceps. Biochem. Biophys. Res. Commun. 2008, 377, 1057–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural determinants of host defense peptides for antimicrobial activity and target cell selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, e24122. [Google Scholar] [CrossRef]
- Neelay, O.P.; Peterson, C.A.; Snavely, M.E.; Brown, T.C.; TecleMariam, A.F.; Campbell, J.A.; Blake, A.M.; Schneider, S.C.; Cremeens, M.E. Antimicrobial peptides interact with peptidoglycan. J. Mol. Struct. 2017, 1146, 329–336. [Google Scholar] [CrossRef]
- Pinto, E.G.; Pimenta, D.C.; Antoniazzi, M.M.; Jared, C.; Tempone, A.G. Antimicrobial peptides isolated from Phyllomedusa nordestina (Amphibia) alter the permeability of plasma membrane of Leishmania and Trypanosoma cruzi. Exp. Parasitol. 2013, 135, 655–660. [Google Scholar] [CrossRef]
- Krugliak, M.; Feder, R.; Zolotarev, V.Y.; Gaidukov, L.; Dagan, A.; Ginsburg, H.; Mor, A. Antimalarial activities of dermaseptin S4 derivatives. Antimicrob. Agents Chemother. 2000, 44, 2442–2451. [Google Scholar] [CrossRef] [Green Version]
- Raja, Z.; André, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef]
- Asthana, N.; Yadav, S.P.; Ghosh, J.K. Dissection of antibacterial and toxic activity of melittin a leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J. Biol. Chem. 2004, 279, 55042–55050. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Wang, C.; Guo, X.; Li, W.; Wang, Y.; He, Q. Broad activity against porcine bacterial pathogens displayed by two insect antimicrobial peptides moricin and cecropin B. Mol. Cells 2013, 35, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef] [PubMed]
- VanCompernolle, S.E.; Taylor, R.J.; Oswald-Richter, K.; Jiang, J.; Youree, B.E.; Bowie, J.H.; Tyler, M.J.; Conlon, J.M.; Wade, D.; Aiken, C. Antimicrobial peptides from amphibian skin potently inhibit human immunodeficiency virus infection and transfer of virus from dendritic cells to T cells. J. Virol. 2005, 79, 11598–11606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holthausen, D.J.; Lee, S.H.; Kumar, V.T.; Bouvier, N.M.; Krammer, F.; Ellebedy, A.H.; Wrammert, J.; Lowen, A.C.; George, S.; Pillai, M.R. An amphibian host defense peptide is virucidal for human H1 hemagglutinin-bearing influenza viruses. Immunity 2017, 46, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinstraesser, L.; Kraneburg, U.; Jacobsen, F.; Al-Benna, S. Host defense peptides and their antimicrobial-immunomodulatory duality. Immunobiology 2011, 216, 322–333. [Google Scholar] [CrossRef]
- Zheng, Y.; Niyonsaba, F.; Ushio, H.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Cathelicidin LL-37 induces the generation of reactive oxygen species and release of human α-defensins from neutrophils. Br. J. Dermatol. 2007, 157, 1124–1131. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Someya, A.; Hirata, M.; Ogawa, H.; Nagaoka, I. Evaluation of the effects of peptide antibiotics human β-defensins-1/-2 and LL-37 on histamine release and prostaglandin D2 production from mast cells. Eur. J. Immunol. 2001, 31, 1066–1075. [Google Scholar] [CrossRef]
- Chen, Q.; Wade, D.; Kurosaka, K.; Wang, Z.Y.; Oppenheim, J.J.; Yang, D. Temporin A and related frog antimicrobial peptides use formyl peptide receptor-like 1 as a receptor to chemoattract phagocytes. J. Immunol. 2004, 173, 2652–2659. [Google Scholar] [CrossRef] [Green Version]
- Auvynet, C.; ElAmri, C.; Lacombe, C.; Bruston, F.; Bourdais, J.; Nicolas, P.; Rosenstein, Y. Structural requirements for antimicrobial versus chemoattractant activities for dermaseptin S9. FEBS J. 2008, 275, 4134–4151. [Google Scholar] [CrossRef] [PubMed]
- Popov, C.S.F.C.; Magalhães, B.S.; Goodfellow, B.J.; Bocca, A.L.; Pereira, D.M.; Andrade, P.B.; Valentão, P.; Pereira, P.J.B.; Rodrigues, J.E.; de Holanda Veloso, P.H.; et al. Host-defense peptides AC12, DK16 and RC11 with immunomodulatory activity isolated from Hypsiboas raniceps skin secretion. Peptides 2019, 113, 11–21. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Mass Calc. (Da) | Mass Obs. (Da) | Net Charge | Hydrophobic Ratio (%) | GRAVY |
|---|---|---|---|---|---|
| Figainin 2 | 3005.76 | 3005.77 | +4 | 53 | 0.575 |
| Water | TFE 10% | TFE 30% | TFE 50% | |
|---|---|---|---|---|
| Figainin 2 | 9.10 | 20.10 | 62.08 | 69.80 |
| 25 °C | 35 °C | 45 °C | 55 °C | 65 °C | 75 °C | 85 °C | 95 °C | |
|---|---|---|---|---|---|---|---|---|
| Figainin 2 | 69.80 | 64.50 | 59.70 | 56.10 | 54.20 | 48.03 | 43.80 | 40.27 |
| Microorganisms | Figainin 2 (µM) |
|---|---|
| Gram-negative bacteria (MIC) | |
| E. coli (ATCC 25922) | 8 |
| P. aeruginosa (ATCC 27853) | 32 |
| K. pneumoniae (ATCC 13883) | 8 |
| K. pneumoniae carbapanemase (KPC) MR | 16 |
| Gram-positive bacteria (MIC) | |
| S. aureus (ATCC 25923) | 8 |
| E. faecalis (ATCC 29212) | 8 |
| S. epidermidis (ATCC 12228) | 4 |
| E. casseliflavus (ATCC 700327) | 4 |
| Yeast (MIC) | |
| C. albicans (ATCC 90028) | NA |
| C. parapsilosis (ATCC 22019) | NA |
| Trypanosoma epimastigotes (IC50) T. cruzi | 6.32 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, C.J.C.; Magalhães, A.C.M.; Prías-Márquez, C.A.; Falico, D.A.; dos Santos Júnior, A.C.M.; Lima, B.D.; Ricart, C.A.O.; de Pilger, D.R.B.; Bonotto, R.M.; Moraes, C.B.; et al. Biological Properties of a Novel Multifunctional Host Defense Peptide from the Skin Secretion of the Chaco Tree Frog, Boana raniceps. Biomolecules 2020, 10, 790. https://doi.org/10.3390/biom10050790
Santana CJC, Magalhães ACM, Prías-Márquez CA, Falico DA, dos Santos Júnior ACM, Lima BD, Ricart CAO, de Pilger DRB, Bonotto RM, Moraes CB, et al. Biological Properties of a Novel Multifunctional Host Defense Peptide from the Skin Secretion of the Chaco Tree Frog, Boana raniceps. Biomolecules. 2020; 10(5):790. https://doi.org/10.3390/biom10050790
Chicago/Turabian StyleSantana, Carlos José Correia, Ana Carolina Martins Magalhães, César Augusto Prías-Márquez, Diego A. Falico, Agenor C. M. dos Santos Júnior, Beatriz D. Lima, Carlos André Ornelas Ricart, Denise Regina Bairros de Pilger, Rafaela Milan Bonotto, Carolina Borsoi Moraes, and et al. 2020. "Biological Properties of a Novel Multifunctional Host Defense Peptide from the Skin Secretion of the Chaco Tree Frog, Boana raniceps" Biomolecules 10, no. 5: 790. https://doi.org/10.3390/biom10050790