The Impact of Phytases on the Release of Bioactive Inositols, the Profile of Inositol Phosphates, and the Release of Selected Minerals in the Technology of Buckwheat Beer Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzymes
2.2.1. Amylase
2.2.2. Phytases and Acid Phosphatase
2.3. Preparation of Laboratory Worts
Mashing
2.4. Inositol Analysis
2.5. Phytate Analysis
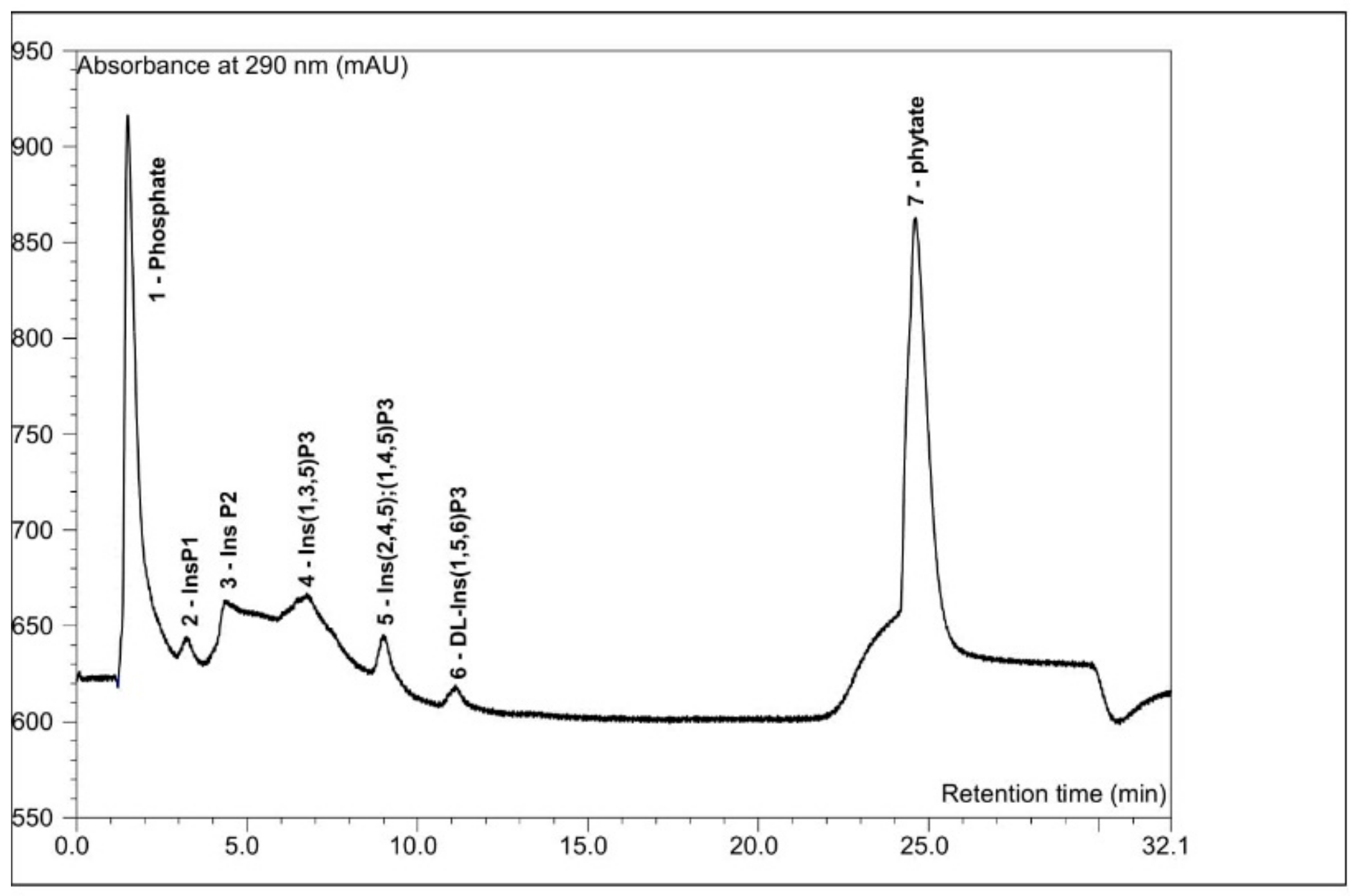
2.6. Semi-Qualitative Analysis of the Inositol Phosphate Profile
2.6.1. Purification of the Inositol Phosphates Using Ion-Exchange Chromatography
2.6.2. HPLC Analysis
2.7. Determination of Metal Ions
2.8. Statistical Analysis
3. Results and Discussion
3.1. Release of Bioactive Inositols
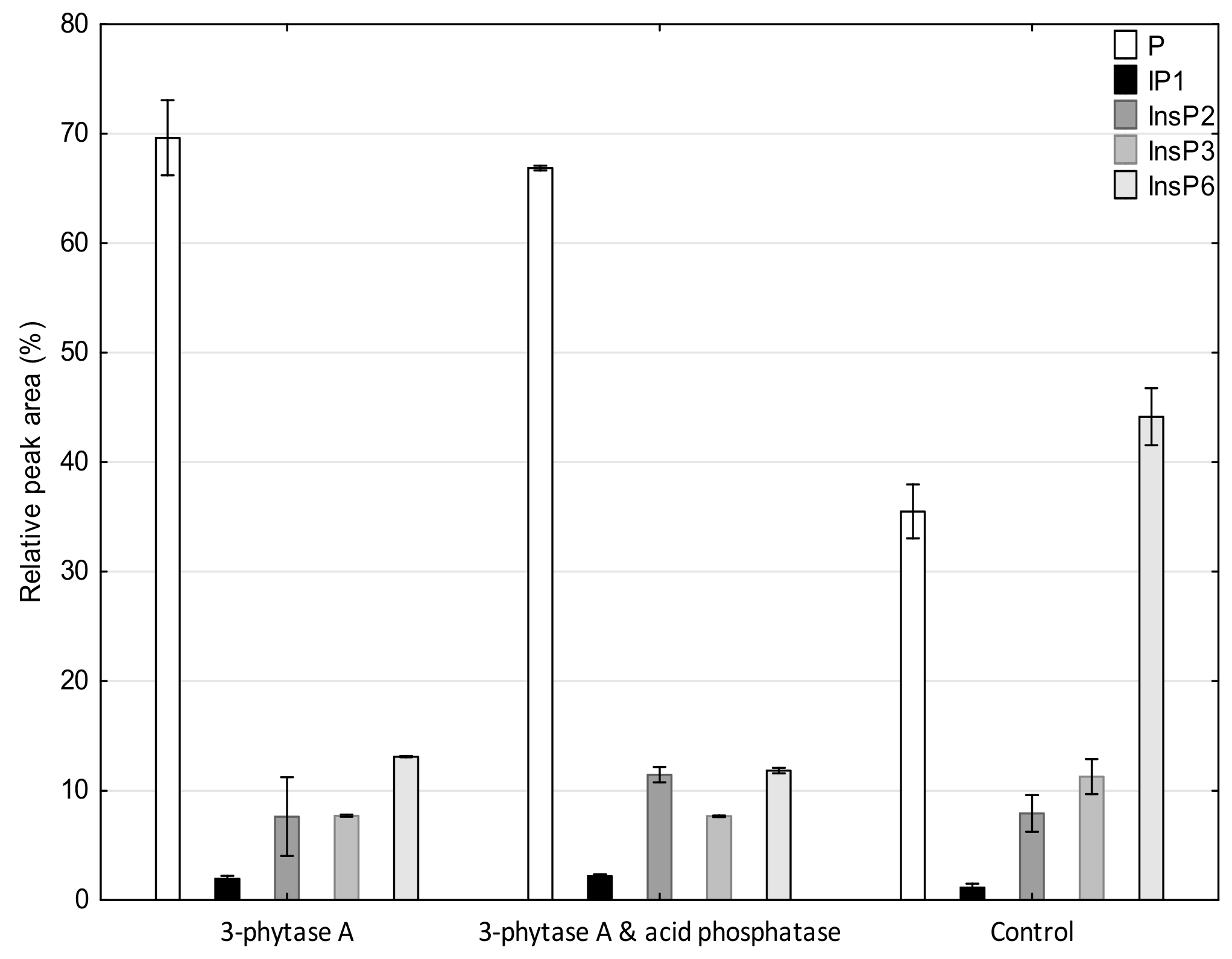
3.2. Phytate and Inositol Phosphates Profile
3.3. Mineral Availability
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Starzynska-Janiszewska, A.; Dulinski, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged Tempe-Type Fermentation in Order to Improve Bioactive Potential and Nutritional Parameters of Quinoa Seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Rubio-Flores, M.; Serna-Saldivar, S.O. Technological and Engineering Trends for Production of Gluten-Free Beers. Food Eng. Rev. 2016, 8, 468–482. [Google Scholar] [CrossRef]
- Tang, C.; Gong, Q.; Sun, X. Functional Properties of Buckwheat (Fagopyrum esculentum Moench) Seed Protein Isolate: Effects of Limited Enzymatic Hydrolysis with Trypsin. Eur. J. Plant Sci. Biotechnol. 2008, 4, 110–116. [Google Scholar]
- Gimenez-Bastida, J.A.; Piskula, M.; Zielinski, H. Recent Advances in Development of Gluten-Free Buckwheat Products. Trends Food Sci. Technol. 2015, 44, 58–65. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Piskula, M.K.; Zieliñski, H. Recent Advances in Processing and Development of Buckwheat Derived Bakery and Non-Bakery Products—A Review. Polish J. Food Nutr. Sci. 2015, 65, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Nepali, B.; Bhandari, D.; Shrestha, J. Mineral Nutrient Content of Buckwheat (Fagopyrum Esculentum Moench) for Nutritional Security in Nepal. Malaysian J. Sustain. Agric. 2019, 3, 1–4. [Google Scholar] [CrossRef]
- Phiarais, B.P.N.; Mauch, A.; Schehl, B.D.; Zarnkow, M.; Gastl, M.; Herrmann, M.; Zannini, E.; Arendt, E.K. Processing of a Top Fermented Beer Brewed from 100% Buckwheat Malt with Sensory and Analytical Characterisation. J. Inst. Brew. 2010, 116, 265–274. [Google Scholar] [CrossRef]
- Duliński, R.; Starzyńska-Janiszewska, A.; Byczyński, Ł.; Błaszczyk, U. Myo-Inositol Phosphates Profile of Buckwheat and Quinoa Seeds: Effects of Hydrothermal Processing and Solid-State Fermentation with Rhizopus Oligosporus. Int. J. Food Prop. 2017, 20, 2088–2095. [Google Scholar] [CrossRef] [Green Version]
- Edney, M.J.; Rossnagel, B.G.; Raboy, V. Effect of Low-Phytate Barley on Malt Quality, Including Mineral Loss, during Fermentation. J. Am. Soc. Brew. Chem. 2007, 65, 81–85. [Google Scholar] [CrossRef]
- Balk, J.; Connorton, J.M.; Wan, Y.; Lovegrove, A.; Moore, K.L.; Uauy, C.; Sharp, P.A. Improving Wheat as a Source of Iron and Zinc for Global Nutrition. Nutrition 2019, 44, 53–59. [Google Scholar] [CrossRef]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on Environment and Human Nutrition. A Challenge for Molecular Breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frontela, C.; Ros, G.; Martínez, C. Phytic Acid Content and “In Vitro ” Iron, Calcium and Zinc Bioavailability in Bakery Products: The Effect of Processing. J. Cereal Sci. 2011, 54, 173–179. [Google Scholar] [CrossRef]
- Konietzny, U.; Greiner, R. Molecular and Catalytic Properties of Phytate-Degrading Enzymes (Phytases). Int. J. Food Sci. Technol. 2002, 37, 791–812. [Google Scholar] [CrossRef] [Green Version]
- Avendano, K.A.; Anguiano, M.; Lopez, C.E.; Montanez, L.E.; Sifuentes, L.; Balagurusamy, N. Microbial Enzymes Applications in Food Processing. Agro Food Ind. Hi. Tech. 2016, 27, 63–67. [Google Scholar]
- Azeke, M.A.; Greiner, R.; Jany, K.D. Purification and Characterization of Two Intracellular Phytases from the Tempeh Fungus Rhizopus Oligosporus. J. Food Biochem. 2011, 35, 213–227. [Google Scholar] [CrossRef]
- Haefner, S.; Knietsch, A.; Scholten, E.; Braun, J.; Lohscheidt, M.; Zelder, O. Biotechnological Production and Applications of Phytases. Appl. Microbiol. Biotechnol. 2005, 68, 588–597. [Google Scholar] [CrossRef]
- Mikulski, D.; Kłosowski, G.; Rolbiecka, A. Influence of Phytase and Supportive Enzymes Applied during High Gravity Mash Preparation Onthe Improvement of Technological Indicators Ofthe Alcoholic Fermentation Process. Biomass Bioenergy 2015, 80, 191–202. [Google Scholar] [CrossRef]
- Duliński, R.; Zdaniewicz, M.; Pater, A.; Żyła, K. Impact of Two Commercial Enzymes on the Release of Inositols, Fermentable Sugars, and Peptides in the Technology of Buckwheat Beer. J. Am. Soc. Brew. Chem. 2019, 77, 119–125. [Google Scholar] [CrossRef]
- Valluru, R.; Van den Ende, W. Myo-Inositol and beyond--Emerging Networks under Stress. Plant Sci. 2011, 181, 387–400. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential Role and Therapeutic Interests of Myo-Inositol in Metabolic Diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Dinicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarri, M. Nutritional and Acquired Deficiencies in Inositol Bioavailability. Correlations with Metabolic Disorders. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Garzon, S.; Laganà, A.S.; Monastra, G. Risk of Reduced Intestinal Absorption of Myo-Inositol Caused by D-Chiro-Inositol or by Glucose Transporter Inhibitors. Expert Opin. Drug Metab. Toxicol. 2019, 15, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Tahir, F.; Majid, Z. Inositol Supplementation in the Prevention of Gestational Diabetes Mellitus. Cureus 2019, 11, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Guo, S.; Miao, Z.; Li, Z.; Zhang, H. Myo-Inositol Lowers the Risk of Developing Gestational Diabetic Mellitus in Pregnancies: A Systematic Review and Meta-Analysis of Randomized Controlled Trials with Trial Sequential Analysis. J. Diabetes Complicat. 2018, 32, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Zyla, K.; Mika, M.; Dulinski, R.; Swiatkiewicz, S.; Koreleski, J.; Pustkowiak, H.; Piironen, J. Effects of Inositol, Inositol-Generating Phytase B Applied Alone, and in Combination with 6-Phytase A to Phosphorus-Deficient Diets on Laying Performance, Eggshell Quality, Yolk Cholesterol, and Fatty Acid Deposition in Laying Hens. Poult. Sci. 2012, 91, 1915–1927. [Google Scholar] [CrossRef]
- Qian, J.Y.; Kuhn, M. Evaluation on Gelatinization of Buckwheat Starch: A Comparative Study of Brabender Viscoamylography, Rapid Visco-Analysis, and Differential Scanning Calorimetry. Eur. Food Res. Technol. 1999, 209, 277–280. [Google Scholar] [CrossRef]
- Duliński, R.; Cielecka, E.K.; Pierzchalska, M.; Żyła, K. Phytases Improve Myo-Inositol Bioaccessibility in Rye Bread: A Study Using an in Vitro Method of Digestion and a Caco-2 Cell Culture Model. Food Technol. Biotechnol. 2015, 53, 66–72. [Google Scholar] [CrossRef]
- Duliński, R.; Starzyńska-Janiszewska, A.; Stodolak, B.; Zyla, K. Comparison of High-Performance Ion Chromatography Technique with Microbiological Assay of myo-Inositol in Plant Components of Poultry Feeds. J. Anim. Feed Sci. 2011, 20, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Dionex—Thermo Fisher Scientific Inc. Application Note 65: Analysis of Inositol Phosphates; Dionex—Thermo Fisher Scientific Inc.: Waltham, MA, USA, 2003. [Google Scholar]
- Blaabjerg, K.; Hansen-Møller, J.; Poulsen, H.D. High-Performance Ion Chromatography Method for Separation and Quantification of Inositol Phosphates in Diets and Digesta. J. Chromatogr. B 2010, 878, 347–354. [Google Scholar] [CrossRef]
- Zyła, K.; Grabacka, M.; Pierzchalska, M.; Duliński, R.; Starzyńska-Janiszewska, A. Effect of Inositol and Phytases on Hematological Indices and α-1 Acid Glycoprotein Levels in Laying Hens Fed Phosphorus-Deficient Corn-Soybean Meal-Based Diets. Poult. Sci. 2013, 92, 199–204. [Google Scholar] [CrossRef]
- Qiu, R.; Lu, J. Improved Hydrolase Activity in Barley and Reduced Malting Time by Adding Phytase as an Activator during Malting Steeping. Biotechnol. Lett. 2017, 39, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Kordialik-Bogacka, E.; Bogdan, P.; Ciosek, A. Effects of Quinoa and Amaranth on Zinc, Magnesium and Calcium Content in Beer Wort. Int. J. Food Sci. Technol. 2019, 54, 1706–1712. [Google Scholar] [CrossRef]
- Raes, K.; Knockaert, D.; Struijs, K.; Van Camp, J. Role of Processing on Bioaccessibility of Minerals: Influence of Localization of Minerals and Anti-Nutritional Factors in the Plant. Trends Food Sci. Technol. 2014, 37, 32–41. [Google Scholar] [CrossRef]
- De Nicola, R.; Walker, G.M. Zinc Interactions with Brewing Yeast: Impact on Fermentation Performance. J. Am. Soc. Brew. Chem. 2011, 69, 214–219. [Google Scholar] [CrossRef]
- Walker, G.; Stewart, G. Saccharomyces Cerevisiae in the Production of Fermented Beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Muy-Rangel, D.; Rubio-Carrasco, W.; Contreras-Angulo, L. Differences in Physicochemical, Mineral and Nutraceutical Properties between Regular, Light and Zero Beers. Farmacia 2018, 66, 697–701. [Google Scholar] [CrossRef]
- Lazarte, C.E.; Carlsson, N.G.; Almgren, A.; Sandberg, A.S.; Granfeldt, Y. Phytate, Zinc, Iron and Calcium Content of Common Bolivian Food, and Implications for Mineral Bioavailability. J. Food Compos. Anal. 2015, 39, 111–119. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Enzyme(s) | Mashing | |||||
|---|---|---|---|---|---|---|
| Optimal | Congress | |||||
| MI | % of Total MI | DCI | MI | % of Total MI | DCI | |
| 3-phytase A | 66.74 ± 2.33 a | 13.8 | 211.07 ± 12.14 a | 71.11 ± 6.04 b | 14.9 | 201.38 ± 9.41 a |
| 6-phytase A | 66.78 ± 5.48 a | 13.9 | 188.83 ± 13.71 a | 68.01 ± 1.91 ab | 14.2 | 223.19 ± 31.9 a |
| 3-phytase and acid phosphatase | 92.66 ± 6.37 b | 19.4 | 229.76 ± 10.61 a | 72.85 ± 7.71 b | 15.2 | 211.84 ± 8.09 a |
| Control | 62.08 ± 1.92 a | 13.0 | 210.99 ± 27.85 a | 61.66 ± 2.68 a | 12.9 | 205.35 ± 16.21 a |
| Mashing | Enzyme(s) | InsP6 | % of Reduction | Mg | Zn | Ca |
|---|---|---|---|---|---|---|
| Optimal | 3-phytase A | 0.72 ± 0.29 b | 74 | 124.68 ± 3.94 a | 1.19 ± 0.21 b | 14.08 ± 2.07 a |
| 6-phytase A | 1.24 ± 0.08 c | 55 | 129.37 ± 5.19 a | 1.44 ± 0.11 b | 15.16 ± 1.02 a | |
| 3-phytase and acid phosphatase | 0.55 ± 0.12 a | 80 | 135.26 ± 6.17 a | 1.43 ± 0.15 b | 14.08 ± 0.89 a | |
| Control | 2.741 ± 0.12 d | - | 125.71 ± 10.16 a | 1.02 ± 0.14 a | 16.12 ± 1.41 a | |
| Congress | 3-phytase A | 1.09 ± 0.14 a | 64 | 134.15 ± 13.91 a | 1.25 ± 0.11 b | 14.38 ± 2.21 a |
| 6-phytase A | 1.14 ± 0.12 a | 63 | 148.83 ± 8.20 a | 1.58 ± 0.13 c | 15.69 ± 2.50 a | |
| 3-phytase and acid phosphatase | 0.99 ± 0.07 a | 68 | 144.06 ± 6.78 a | 1.26 ± 0.15 b | 11.45 ± 2.1 a | |
| Control | 3.075 ± 055 b | - | 145.21 ± 4.99 a | 1.12 ± 0.01 a | 18.81 ± 4.29 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duliński, R.; Zdaniewicz, M.; Pater, A.; Poniewska, D.; Żyła, K. The Impact of Phytases on the Release of Bioactive Inositols, the Profile of Inositol Phosphates, and the Release of Selected Minerals in the Technology of Buckwheat Beer Production. Biomolecules 2020, 10, 166. https://doi.org/10.3390/biom10020166
Duliński R, Zdaniewicz M, Pater A, Poniewska D, Żyła K. The Impact of Phytases on the Release of Bioactive Inositols, the Profile of Inositol Phosphates, and the Release of Selected Minerals in the Technology of Buckwheat Beer Production. Biomolecules. 2020; 10(2):166. https://doi.org/10.3390/biom10020166
Chicago/Turabian StyleDuliński, Robert, Marek Zdaniewicz, Aneta Pater, Dagmara Poniewska, and Krzysztof Żyła. 2020. "The Impact of Phytases on the Release of Bioactive Inositols, the Profile of Inositol Phosphates, and the Release of Selected Minerals in the Technology of Buckwheat Beer Production" Biomolecules 10, no. 2: 166. https://doi.org/10.3390/biom10020166