
Multi-Stage Transcriptome Analysis Revealed the Growth Mechanism of Feathers and Hair Follicles during Induction Molting by Fasting in the Late Stage of Egg Laying
 ,
,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Animal Experimental Design
2.3. Sample Collection
2.4. Paraffin Tissue Embedding and Section Manufacture and HE Staining
2.5. Feather Composition Determination
2.6. Determination of Skin Follicle Properties
2.7. RNA Extraction and RNA-Seq Analysis
2.7.1. Quality Control of Sequencing Data
2.7.2. Raw Data Quality Control, Comparison with Reference Genome, and DEG Analysis
2.7.3. GO and KEGG
2.7.4. Series Test of Cluster
2.7.5. WGCNA
2.7.6. PPI
2.7.7. Validation of RNA-Seq Data Using Quantitative Real-Time PCR (qRT-PCR)
2.8. Data Processing
3. Results
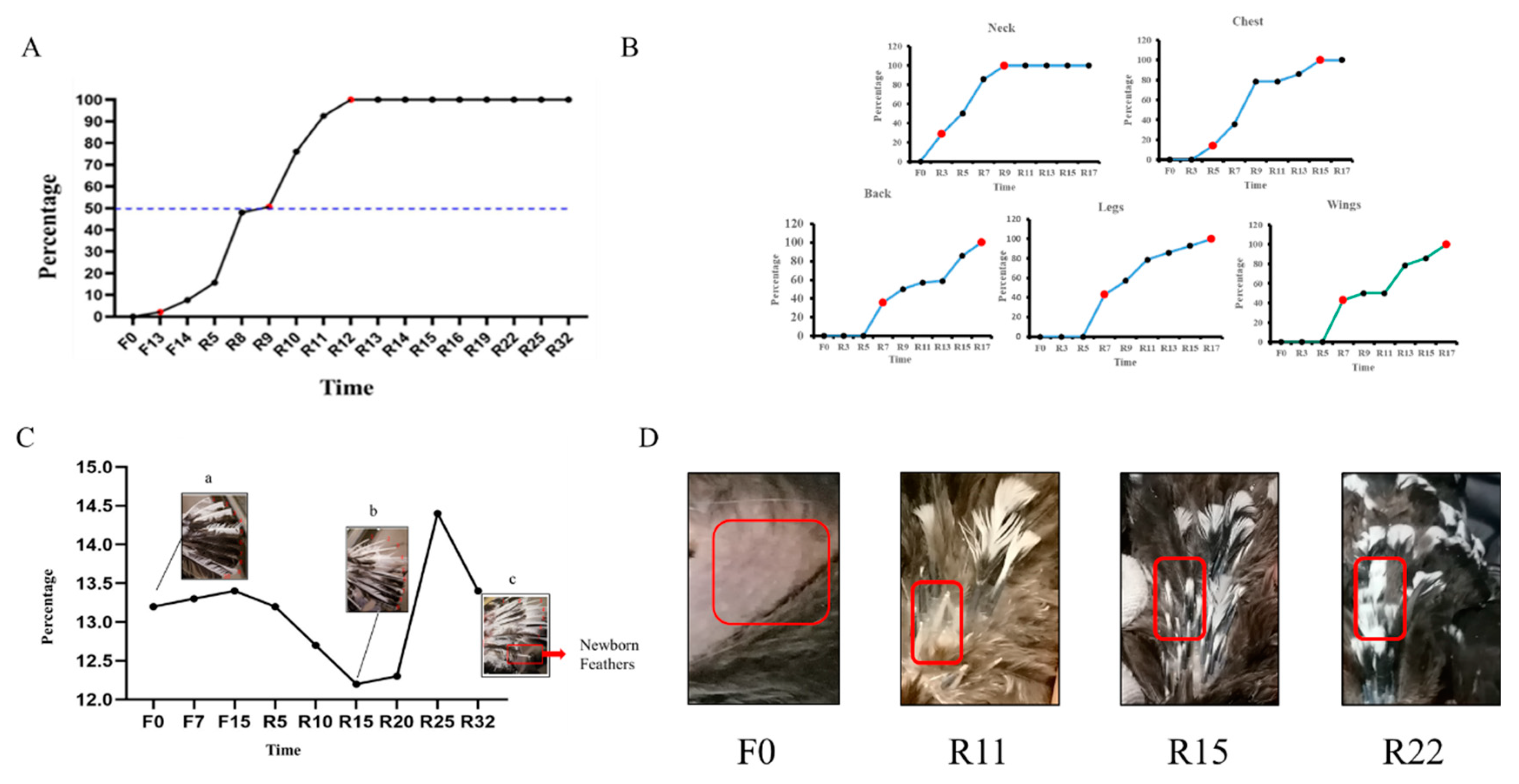
3.1. Changes in the Flock during IM
3.1.1. Chicken Status and Feather Characteristics at Different Periods of IM
3.1.2. Regeneration of Feathers in Different Parts of the Houdan Chicken
3.1.3. The Replacement of the Overall Primary Flight Feathers of the Chicken Flock
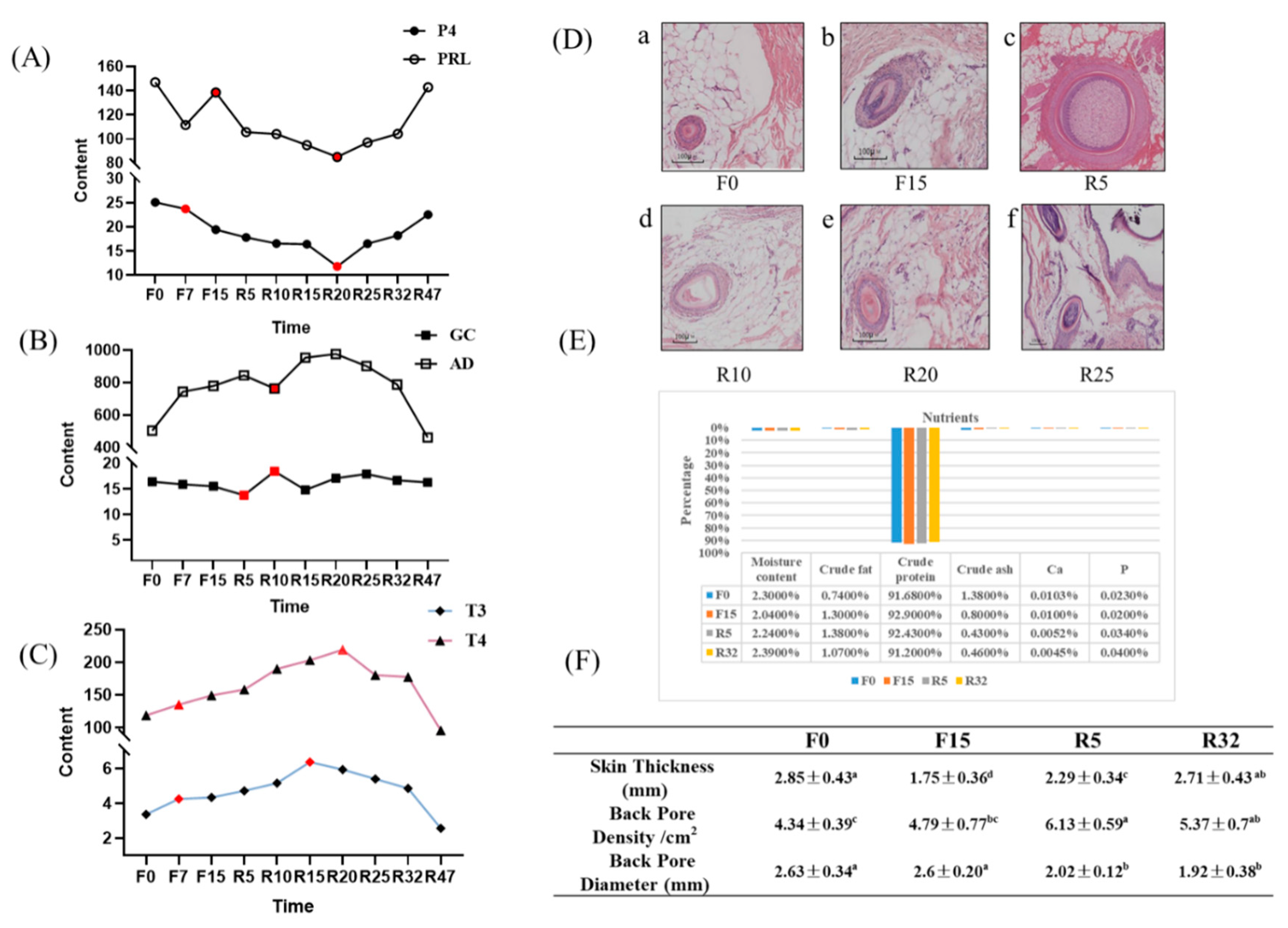
3.2. Serum Hormone Changes and Histological Observation
Changes in Hormone Levels in the Serum
3.3. Histological Observations
3.4. Feather Composition Measurement
3.5. Skin Follicle Properties
3.6. Transcriptome Analysis of Late-Laying Chickens’ Skin
3.6.1. Quality Control of Sequencing Data
3.6.2. Differential Expression Gene (DEG) Analysis
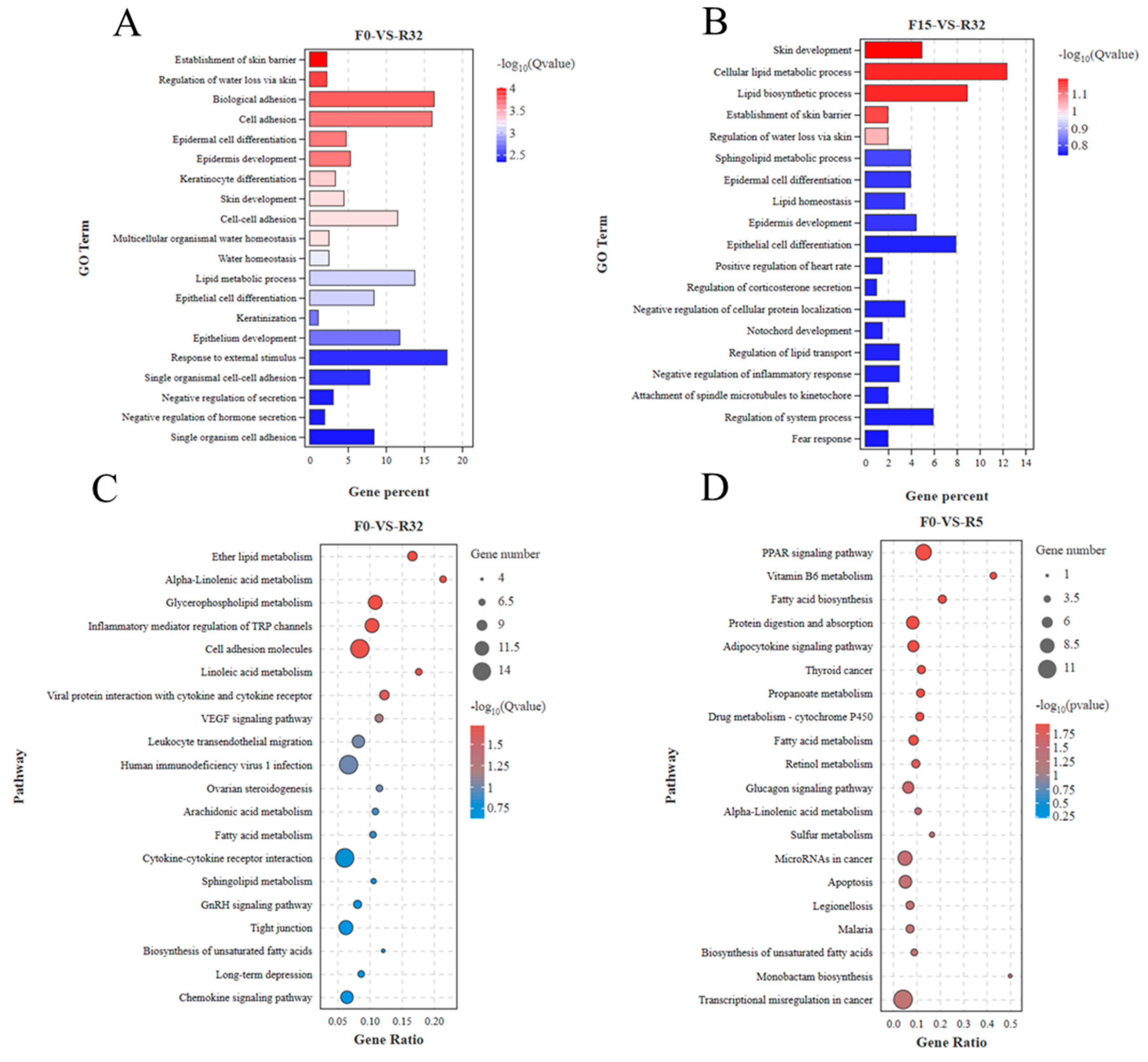
3.6.3. GO and KEGG Analyses
3.6.4. Series Test of Cluster
3.6.5. Protein–Protein Interaction (PPI) Network Analysis
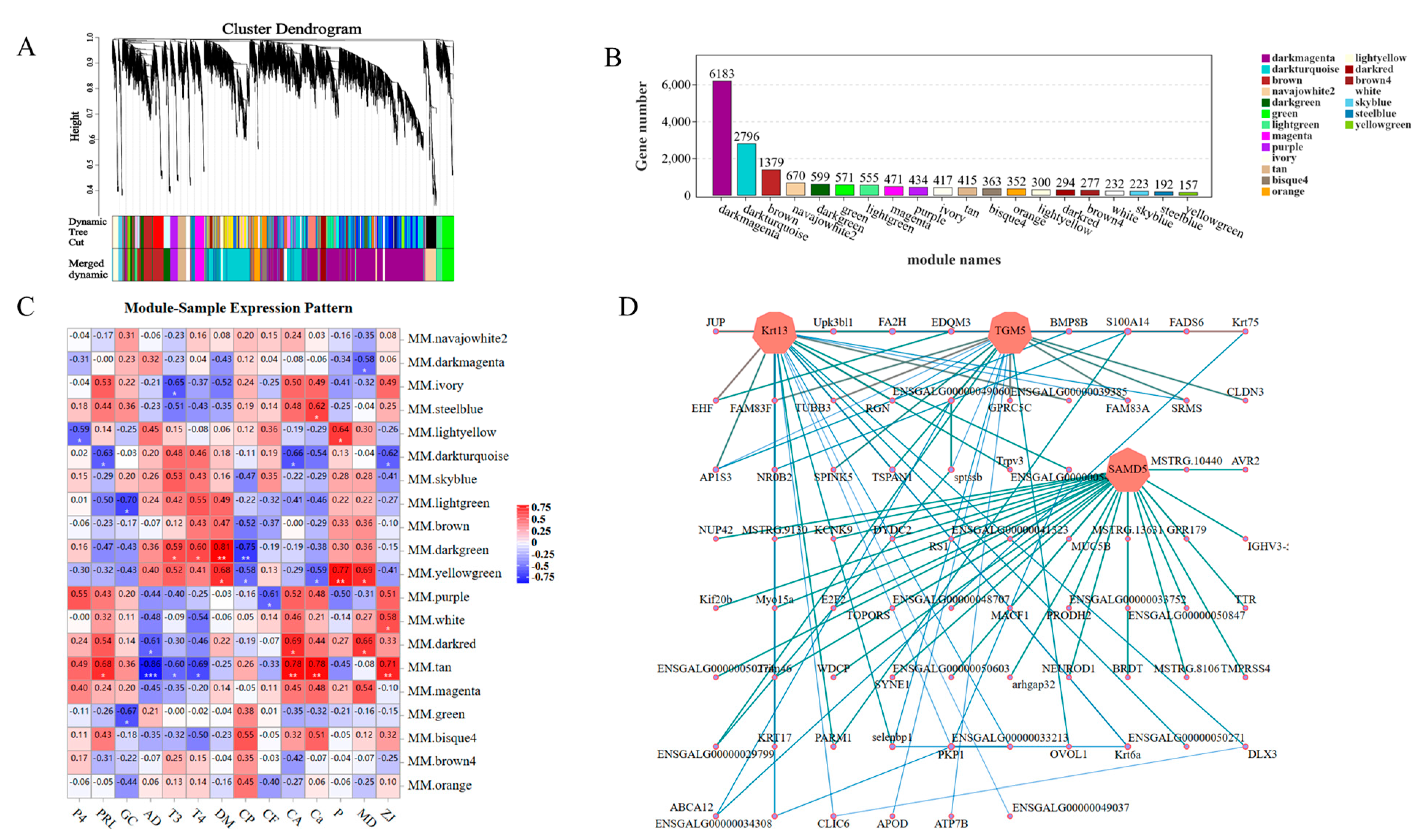
3.6.6. WGCNA
3.6.7. Quantitative Validation of Transcriptome Sequencing Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yousaf, M.; Chaudhry, A.S. History, changing scenarios and future strategies to induce moulting in laying hens. World’s Poult. Sci. J. 2008, 64, 65–75. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, Y.; Wen, J.; Jia, Y.; Wang, L.; Lv, X.; Yang, W.; Qu, C.; Li, H.; Wang, H.; et al. Transcriptomic Analysis of Laying Hens Revealed the Role of Aging-Related Genes during Forced Molting. Genes 2021, 12, 1767. [Google Scholar] [CrossRef]
- Chen, C.F.; Foley, J.; Tang, P.C.; Li, A.; Jiang, T.X.; Wu, P.; Widelitz, R.B.; Chuong, C.M. Development, regeneration, and evolution of feathers. Annu. Rev. Anim. Biosci. 2015, 3, 169–195. [Google Scholar] [CrossRef]
- Xie, W.Y.; Chen, M.J.; Jiang, S.G.; Yan, H.C.; Wang, X.Q.; Gao, C.Q. The Wnt/beta-catenin signaling pathway is involved in regulating feather growth of embryonic chicks. Poult. Sci. 2020, 99, 2315–2323. [Google Scholar] [CrossRef]
- Paster, M.B. Avian Reproductive Endocrinology. Vet. Clin. N. Am. Small Anim. Pract. 1991, 21, 1343–1359. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, V.S.; Yoshimura, Y. Changes in the population of pituitary protein transcription factor-1 nuclei in the anterior pituitary during withdrawal and resumption of feeding in hens. Poult. Sci. 2003, 82, 1637–1640. [Google Scholar] [CrossRef] [PubMed]
- Keeler, C.; Dannies, P.S.; Hodsdon, M.E. The Tertiary Structure and Backbone Dynamics of Human Prolactin. J. Mol. Biol. 2003, 328, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Review: Cornification, morphogenesis and evolution of feathers. Protoplasma 2017, 254, 1259–1281. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Korovin, K.F. Participation of sympathetic-adrenal system catecholamines in regulating the reproduction and molting of the hen. Zhurnal Evoliutsionnoi Biokhimii Fiziol. 1984, 20, 38–42. [Google Scholar]
- Davis, G.S.; Anderson, K.E.; Carroll, A.S. The effects of long-term caging and molt of Single Comb White Leghorn hens on heterophil to lymphocyte ratios, corticosterone and thyroid hormones1. Poult. Sci. 2000, 79, 514–518. [Google Scholar] [CrossRef]
- Sekimoto, K.; Imai, K.; Suzuki, M.; Takikawa, H.; Hoshino, N.; Totsuka, K. Thyroxine-Induced Molting and Gonadal Function of Laying Hens. Poult. Sci. 1987, 66, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Jiang, T.-X.; Lei, M.; Chen, C.-K.; Hsieh Li, S.-M.; Widelitz, R.B.; Chuong, C.-M. Cyclic growth of dermal papilla and regeneration of follicular mesenchymal components during feather cycling. Development 2021, 148, dev198671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, F.; Cao, J.; Liu, X. Skin transcriptome profiles associated with skin color in chickens. PLoS ONE 2015, 10, e0127301. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Wang, H.; Wang, Y.; Bai, X.; Liu, B.; He, J.; Wu, J.; Qi, W.; Zhang, W. Skin transcriptome reveals the dynamic changes in the Wnt pathway during integument morphogenesis of chick embryos. PLoS ONE 2018, 13, e0190933. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.S.; Chen, C.-K.; Fan, W.-L.; Wu, P.; Wu, S.-M.; Chen, J.-J.; Lai, Y.-T.; Mao, C.-T.; Lu, M.-Y.J.; Chen, D.-R.; et al. Transcriptomic analyses of regenerating adult feathers in chicken. BMC Genom. 2015, 16, 756. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Chen, M.J.; Xie, W.Y.; Jiang, S.G.; Wang, X.Q.; Yan, H.C.; Gao, C.Q. Molecular Signaling and Nutritional Regulation in the Context of Poultry Feather Growth and Regeneration. Front. Physiol. 2019, 10, 1609. [Google Scholar] [CrossRef]
- Lendvai, Á.Z.; Tóth, Z.; Mahr, K.; Osváth, G.; Vogel-Kindgen, S.; Gander, B.A. Effects of experimental increase in insulin-like growth factor 1 on feather growth rate, moult intensity and feather quality in a passerine bird. J. Exp. Biol. 2021, 224, jeb242481. [Google Scholar] [CrossRef]
- Fan, Y.; Wu, X.; Li, Y.; Han, H.; Zhang, Y.; Yang, J.; Liu, Y. Effect of polymorphisms in the 5′-flanking sequence of MC1R on feather color in Taihang chickens. Poult. Sci. 2022, 101, 102192. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Agrawal, K.; Verma, P. Chicken feathers: A treasure cove of useful metabolites and value-added products. Environ. Sustain. 2021, 4, 231–243. [Google Scholar] [CrossRef]
- Hussein, A.S. Induced moulting procedures in laying fowl. World’s Poult. Sci. J. 1996, 52, 175–187. [Google Scholar] [CrossRef]
- Tesfaye, T.; Sithole, B.; Ramjugernath, D.; Chunilall, V. Valorisation of chicken feathers: Characterisation of chemical properties. Waste Manag. 2017, 68, 626–635. [Google Scholar] [CrossRef]
- Gupta, S.; Sharma, S.; Aich, A.; Verma, A.K.; Bhuyar, P.; Nadda, A.K.; Mulla, S.I.; Kalia, S. Chicken Feather Waste Hydrolysate as a Potential Biofertilizer for Environmental Sustainability in Organic Agriculture Management. Waste Biomass Valorization 2023, 14, 2783–2799. [Google Scholar] [CrossRef]
- Wang, G.-Y.; Wang, H.-H.; Han, Y.-W.; Xing, T.; Ye, K.-P.; Xu, X.-L.; Zhou, G.-H. Evaluation of the spoilage potential of bacteria isolated from chilled chicken in vitro and in situ. Food Microbiol. 2017, 63, 139–146. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, S.; Chen, L.; Ding, H.; Wu, P.; Zhang, G.; Xie, K.; Dai, G.; Wang, J. UHPLC-MS/MS-Based Nontargeted Metabolomics Analysis Reveals Biomarkers Related to the Freshness of Chilled Chicken. Foods 2020, 9, 1326. [Google Scholar] [CrossRef]
- Wang, K.; Wang, X.; Zhang, L.; Chen, A.; Yang, S.; Xu, Z. Identification of novel biomarkers in chilled and frozen chicken using metabolomics profiling and its application. Food Chem. 2022, 393, 133334. [Google Scholar] [CrossRef]
- Narasimha Rao, D.; Sachindra, N.M. Modified atmosphere and vacuum packaging of meat and poultry products. Food Rev. Int. 2002, 18, 263–293. [Google Scholar] [CrossRef]
- Wang, J.; Sui, J.; Mao, C.; Li, X.; Chen, X.; Liang, C.; Wang, X.; Wang, S.-H.; Jia, C. Identification of Key Pathways and Genes Related to the Development of Hair Follicle Cycle in Cashmere Goats. Genes 2021, 12, 180. [Google Scholar] [CrossRef]
- Wu, C.; Li, J.; Xu, X.; Xu, Q.; Qin, C.; Liu, G.; Wei, C.; Zhang, G.; Tian, K.; Fu, X. Effect of the FA2H Gene on cashmere fineness of Jiangnan cashmere goats based on transcriptome sequencing. BMC Genom. 2022, 23, 527. [Google Scholar] [CrossRef]
- Ji, G.G.; Zhang, M.; Liu, Y.F.; Shan, Y.J.; Tu, Y.J.; Ju, X.J.; Zou, J.M.; Shu, J.T.; Wu, J.F.; Xie, J.F. A gene co-expression network analysis of the candidate genes and molecular pathways associated with feather follicle traits of chicken skin. J. Anim. Breed. Genet. 2021, 138, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Lachner, J.; Ehrlich, F.; Wielscher, M.; Farlik, M.; Hermann, M.; Tschachler, E.; Eckhart, L. Single-cell transcriptomics defines keratinocyte differentiation in avian scutate scales. Sci. Rep. 2022, 12, 126. [Google Scholar] [CrossRef]
- van Roy, F. Beyond E-cadherin: Roles of other cadherin superfamily members in cancer. Nat. Rev. Cancer 2014, 14, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Pouso, A.I.; Silva, F.F.E.; Pérez-Jardón, A.; Chamorro-Petronacci, C.M.; Oliveira-Alves, M.G.; Álvarez-Calderón-Iglesias, Ó.; Caponio, V.C.A.; Pinti, M.; Perrotti, V.; Pérez-Sayáns, M. Overexpression of E-Cadherin Is a Favorable Prognostic Biomarker in Oral Squamous Cell Carcinoma: A Systematic Review and Meta-Analysis. Biology 2023, 12, 239. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Cheng, L.T.; Wang, Z.F.; Wu, Y.Q. Desmoplakin and clinical manifestations of desmoplakin cardiomyopathy. Chin. Med. J. 2021, 134, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Al-Owain, M.; Wakil, S.; Shareef, F.; Al-Fatani, A.; Hamadah, E.; Haider, M.; Al-Hindi, H.; Awaji, A.; Khalifa, O.; Baz, B.; et al. Novel homozygous mutation in DSP causing skin fragility-woolly hair syndrome: Report of a large family and review of the desmoplakin-related phenotypes. Clin. Genet. 2011, 80, 50–58. [Google Scholar] [CrossRef]
- Favre, B.; Begré, N.; Borradori, L. A recessive mutation in the DSP gene linked to cardiomyopathy, skin fragility and hair defects impairs the binding of desmoplakin to epidermal keratins and the muscle-specific intermediate filament desmin. Br. J. Dermatol. 2018, 179, 797–799. [Google Scholar] [CrossRef]
- Szabó, A.; Fébel, H.; Mézes, M.; Horn, P.; Balogh, K.; Romvári, R. Differential utilization of hepatic and myocardial fatty acids during forced molt of laying hens. Poult. Sci. 2005, 84, 106–112. [Google Scholar] [CrossRef]
- Beltran, R.S.; Burns, J.M.; Breed, G.A. Convergence of biannual moulting strategies across birds and mammals. Proc. R. Soc. B Boil. Sci. 2018, 285, 20180318. [Google Scholar] [CrossRef]
- Chen, Z.; Ju, H.; Zhao, T.; Yu, S.; Li, P.; Jia, J.; Li, N.; Jing, X.; Tan, B.; Li, Y. hsa_circ_0092306 Targeting miR-197-3p Promotes Gastric Cancer Development by Regulating PRKCB in MKN-45 Cells. Mol. Ther. Nucleic Acids 2019, 18, 617–626. [Google Scholar] [CrossRef]
- Patergnani, S.; Marchi, S.; Rimessi, A.; Bonora, M.; Giorgi, C.; Mehta, K.D.; Pinton, P. PRKCB/protein kinase C, beta and the mitochondrial axis as key regulators of autophagy. Autophagy 2013, 9, 1367–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, L.; Zhang, Y.; Liu, L.; Huang, Y.; Wen, L.; Yang, C.; Chen, L.; Wang, W.; Zuo, X.; et al. Increased expression of PRKCB mRNA in peripheral blood mononuclear cells from patients with systemic lupus erythematosus. Ann. Hum. Genet. 2018, 82, 200–205. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
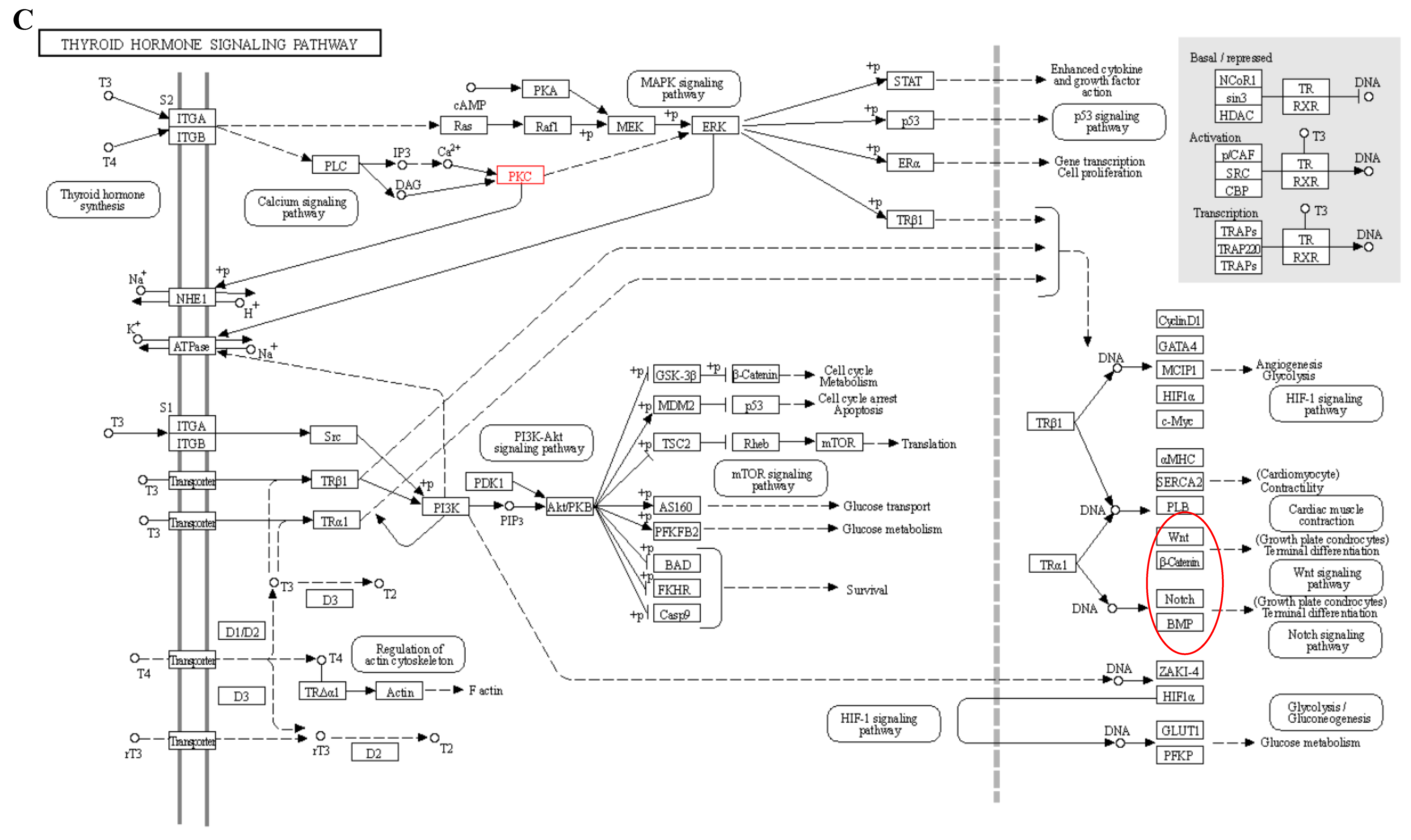{kind=link}
| Test Period | Processing Phase (Days) | Feed | Water | Light (h/d) |
|---|---|---|---|---|
| Fasting Period | 1–3 | No feed | × | 8 |
| 4–15 | No feed | √ | 8 | |
| Recovery Period | 16–30 | Gradually resuming feeding | √ | Increase 0.5 h per day until 16 h |
| Second egg-laying Period | 30–49 | Normal feed | √ | 16 |
| Target Gene | Primer Sequences (5’ to 3’) |
|---|---|
| GAPDH | F: GAACATCATCCCAGCGTCCA |
| R:CGGCAGGTCAGGTCAACAAC | |
| DSP | F: AAAGCAGGCTCTGGAGGCAT |
| R: TTCCAGGCGTTGCTTCAAAC | |
| Krt6a | F: ATGCAGACCCAGATCTCCGA |
| R: GCAGCTCTTCGTACTTGGTT | |
| CDH1 | F: CTGTCTTCGTGCCCCCTATC |
| R: CCCATGCGGTACGTGATCTT | |
| FA2H | F: TCTTCCACATGAAGCCACCC |
| R: TGTCAAAGGGGGACTTGTGG | |
| AOX1 | F:CGTGAATGGGAGAAAGGTGGT |
| R: ACCTCCTCCACAGCCATACT | |
| PKP1 | F: TCATGTCCAACCCACACCTG |
| R: TGCTTTTGGACAGGAGCCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Cai, C.; Liu, X.; Zhang, X.; An, Z.; Zhou, E.; Li, J.; Li, Z.; Li, W.; Sun, G.; et al. Multi-Stage Transcriptome Analysis Revealed the Growth Mechanism of Feathers and Hair Follicles during Induction Molting by Fasting in the Late Stage of Egg Laying. Biology 2023, 12, 1345. https://doi.org/10.3390/biology12101345
Zhang L, Cai C, Liu X, Zhang X, An Z, Zhou E, Li J, Li Z, Li W, Sun G, et al. Multi-Stage Transcriptome Analysis Revealed the Growth Mechanism of Feathers and Hair Follicles during Induction Molting by Fasting in the Late Stage of Egg Laying. Biology. 2023; 12(10):1345. https://doi.org/10.3390/biology12101345
Chicago/Turabian StyleZhang, Lujie, Chunxia Cai, Xinxin Liu, Xiaoran Zhang, Zhiyuan An, Enyou Zhou, Jianzeng Li, Zhuanjian Li, Wenting Li, Guirong Sun, and et al. 2023. "Multi-Stage Transcriptome Analysis Revealed the Growth Mechanism of Feathers and Hair Follicles during Induction Molting by Fasting in the Late Stage of Egg Laying" Biology 12, no. 10: 1345. https://doi.org/10.3390/biology12101345