
Nervous System Development and Neuropeptides Characterization in Embryo and Larva: Insights from a Non-Chordate Deuterostome, the Sea Cucumber Apostichopus japonicus

 , and
, and 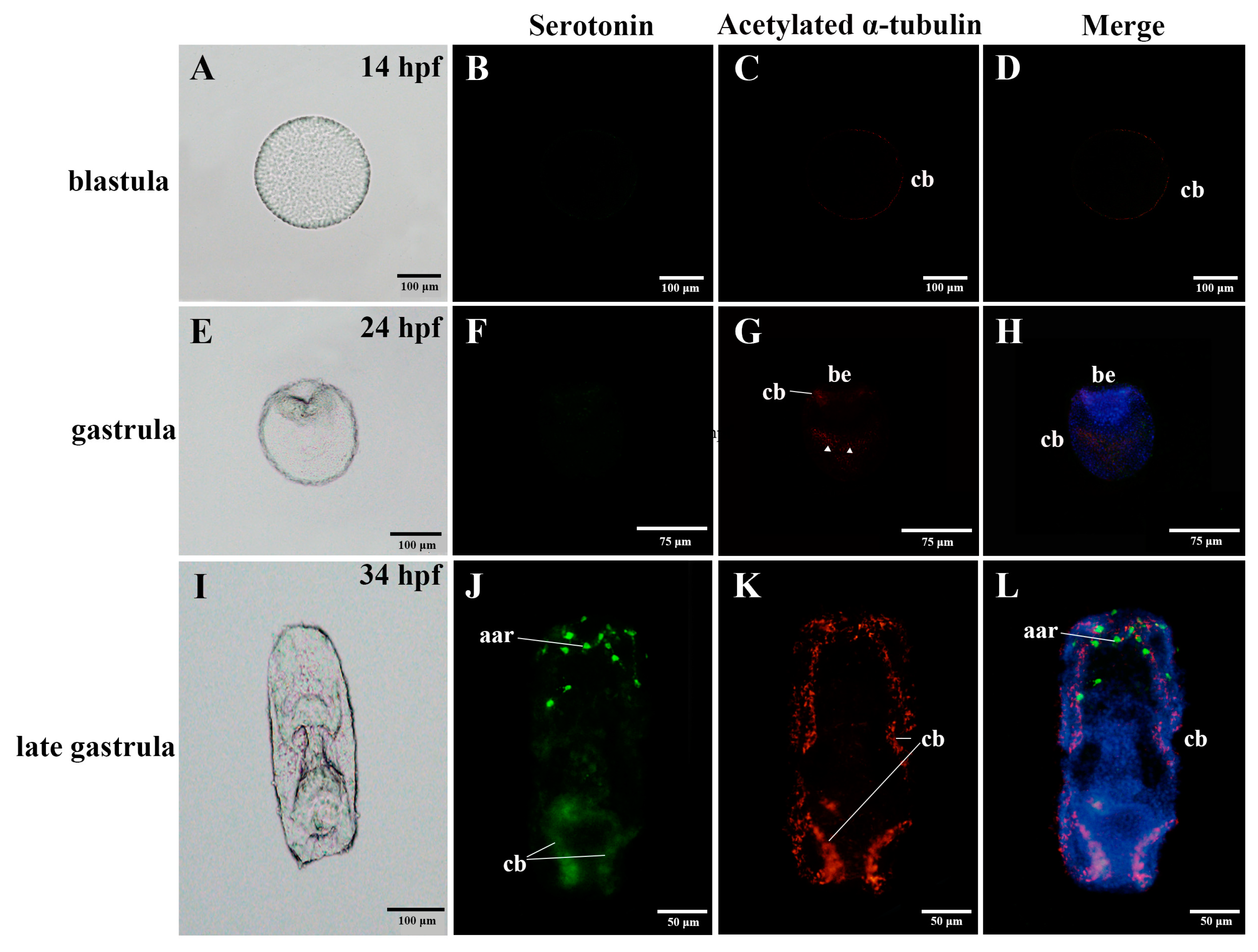{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Embryo and Larval Culture
2.2. Immunostaining
2.3. Identification of Neuropeptide Precursors and Putative Neuropeptides in A. japonicus Embryonic and Larval Stages
2.4. RNA Isolation, cDNA Synthesis and Full-Length Cloning of Putative Neuropeptide Precursor Genes (NPs)
2.5. Quantitative Real-Time PCR (qRT-PCR) in Early Developmental Stages of A. japonicus
3. Results
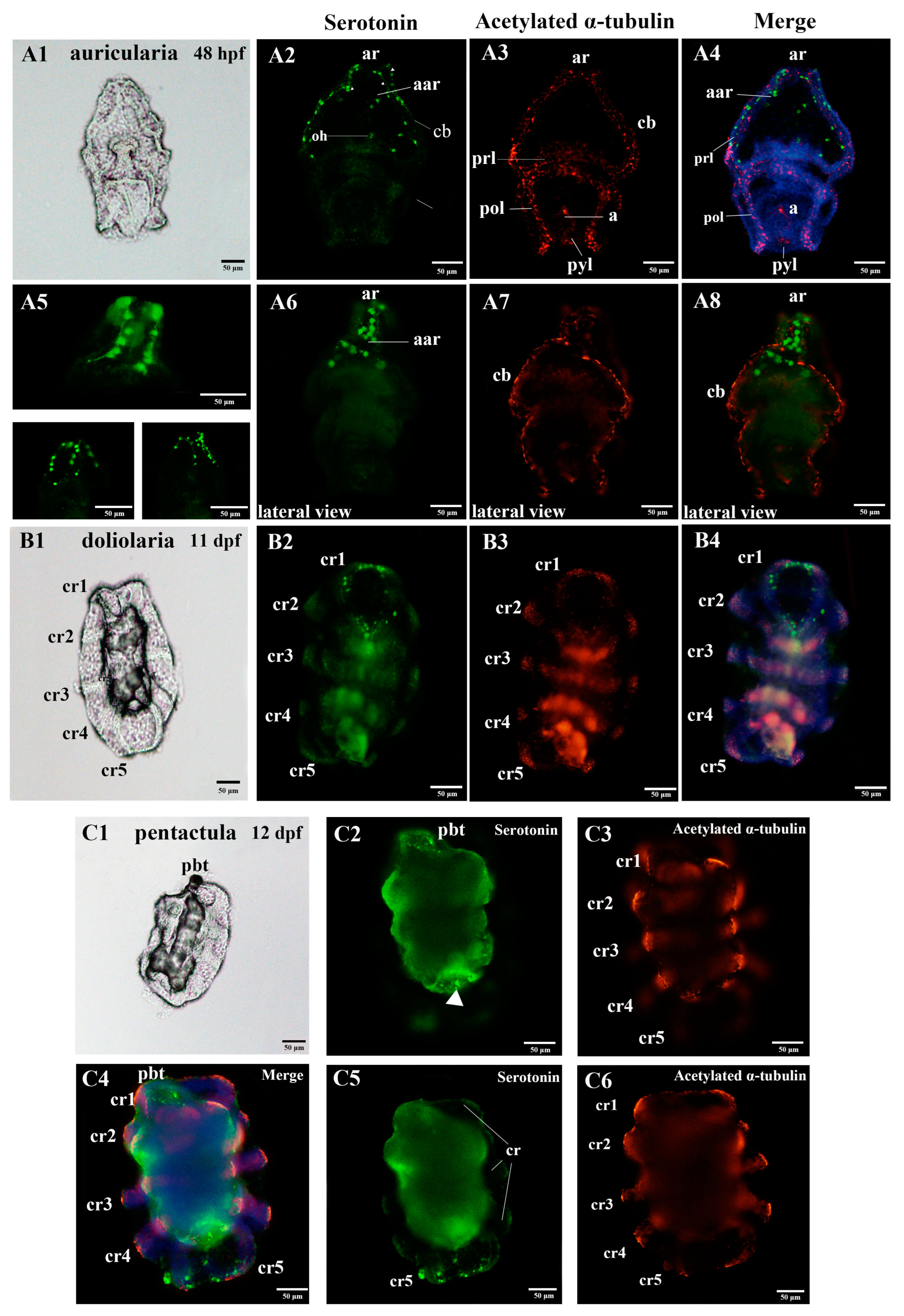
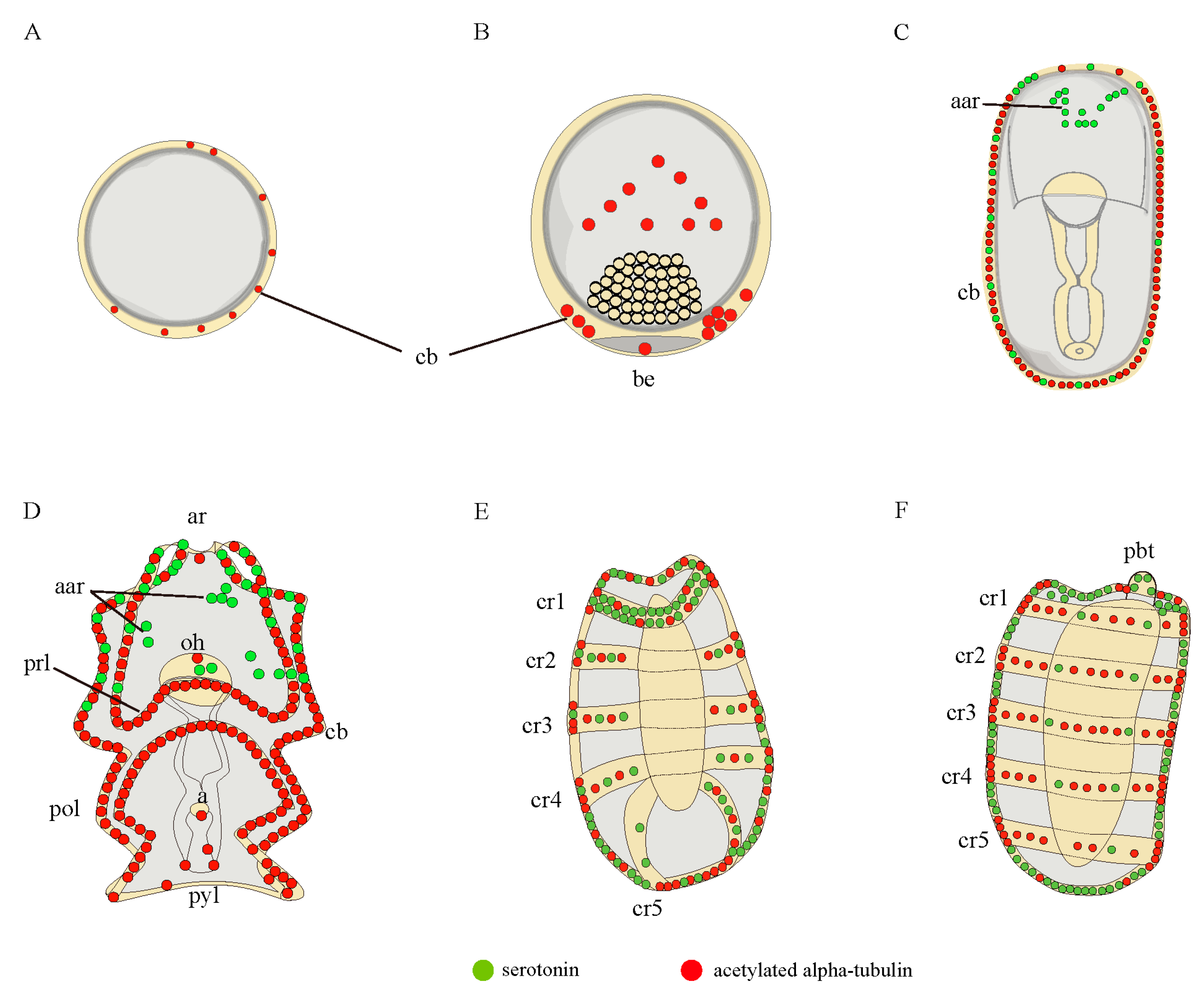
3.1. Nervous System Profile of A. japonicus at Different Developmental Stages
3.2. Identification of Neuropeptide Precursor Transcripts in A. japonicus Embryo and Larvae
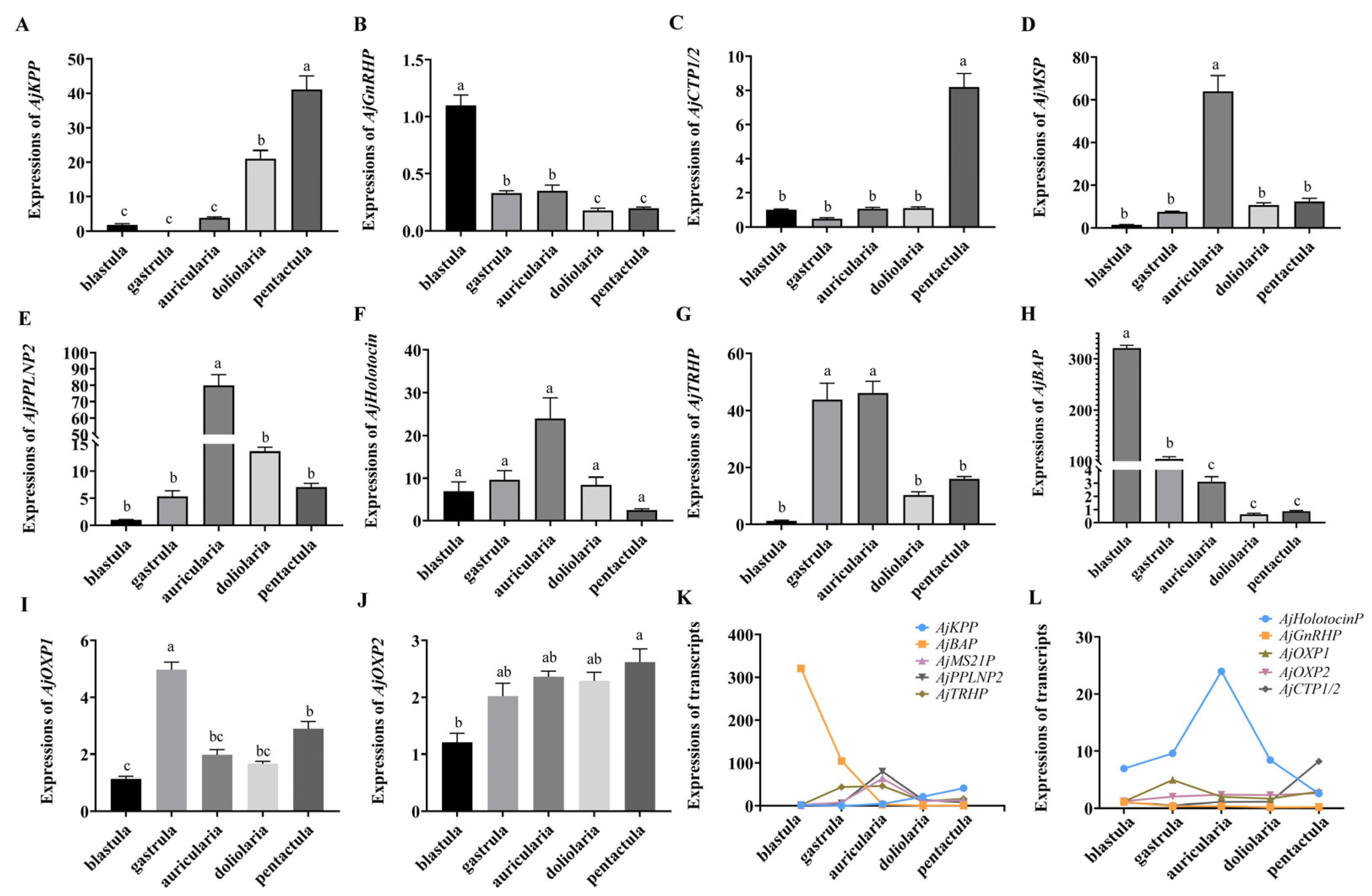
3.3. Quantitative Analysis of Specific Neuropeptide Precursors in A. japonicus Embryos and Larvae
4. Discussion
4.1. Nervous System Complexity at Early Developmental Stages
4.2. Neuropeptides at Early Developmental Stages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arnone, M.I.; Andrikou, C.; Annunziata, R. Echinoderm systems for gene regulatory studies in evolution and development. Curr. Opin. Genet. Dev. 2016, 39, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Nakano, H.; Murabe, N.; Amemiya, S.; Nakajima, Y. Nervous system development of the sea cucumber Stichopus japonicus. Dev. Biol. 2006, 292, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Arraras, J.E.; Rojas-Soto, M.; Jimenez, L.B.; Diaz-Miranda, L. The enteric nervous system of echinoderms: Unexpected complexity revealed by neurochemical analysis. J. Exp. Biol. 2001, 204, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.S.; Zueva, O.R.; Heinzeller, T.; Dolmatov, I.Y. Ultrastructure of the circumoral nerve ring and the radial nerve cords in holothurians (Echinodermata). Zoomorphology 2006, 125, 27–38. [Google Scholar] [CrossRef]
- Hyman, L.H. The Invertebrates; Mc Graw-Hill Publications: New York, NY, USA, 1955. [Google Scholar]
- Diaz-Balzac, C.A.; Abreu-Arbelo, J.E.; Garcia-Arraras, J.E. Neuroanatomy of the tube feet and tentacles in Holothuria glaberrima (Holothuroidea, Echinodermata). Zoomorphology 2010, 129, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Balzac, C.A.; Lazaro-Pena, M.I.; Vazquez-Figueroa, L.D.; Diaz-Balzac, R.J.; Garcia-Arraras, J.E. Holothurian Nervous System Diversity Revealed by Neuroanatomical Analysis. PLoS ONE 2016, 11, e0151129. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Balzac, C.A.; García-Arrarás, J.E. Echinoderm nervous system. Oxf. Res. Encycl. Neurosci. 2018. [Google Scholar] [CrossRef]
- Heinzeller, T.; Welsch, U. The echinoderm nervous system and its phylogenetic interpretation. In Brain Evolution and Cognition; Roth, G., Wullimann, M.F., Eds.; John Wiley and Sons: New York, NY, USA, 2001; pp. 41–75. [Google Scholar]
- Buznikov, G.A.; Peterson, R.E.; Nikitina, L.A.; Bezuglov, V.V.; Lauder, J.M. The pre-nervous serotonergic system of developing sea urchin embryos and larvae: Pharmacologic and immunocytochemical evidence. Neurochem. Res. 2005, 30, 825–837. [Google Scholar] [CrossRef]
- Bishop, C.D.; Burke, R.D. Ontogeny of the holothurian larval nervous system: Evolution of larval forms. Dev. Genes Evol. 2007, 217, 585–592. [Google Scholar] [CrossRef]
- Byrne, M.; Nakajima, Y.; Chee, F.C.; Burke, R.D. Apical organs in echinoderm larvae: Insights into larval evolution in the Ambulacraria. Evol. Dev. 2007, 9, 432–445. [Google Scholar] [CrossRef]
- Carter, H.F.; Thompson, J.R.; Elphick, M.R.; Oliveri, P. The development and neuronal complexity of bipinnaria larvae of the sea star Asterias rubens. Integr. Comp. Biol. 2021, 61, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Sillar, K.T.; Reith, C.A.; McDearmid, J.R. Development and Aminergic Neuromodulation of a Spinal Locomotor Network Controlling Swimming in Xenopus Larvae. Ann. N. Y. Acad. Sci. 1998, 860, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Kaneko, H.; Murray, G.; Burke, R.D. Divergent patterns of neural development in larval echinoids and asteroids. Evol. Dev. 2004, 6, 95–104. [Google Scholar] [CrossRef]
- Hinman, V.F.; Burke, R.D. Embryonic neurogenesis in echinoderms. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e316. [Google Scholar] [CrossRef]
- Nakano, H.; Nakajima, Y.; Amemiya, S. Nervous system development of two crinoid species, the sea lily Metacrinus rotundus and the feather star Oxycomanthus japonicus. Dev. Genes Evol. 2009, 219, 565–576. [Google Scholar] [CrossRef]
- Mercurio, S.; Gattoni, G.; Messinetti, S.; Sugni, M.; Pennati, R. Nervous system characterization during the development of a basal echinoderm, the feather star Antedon mediterranea. J. Comp. Neurol. 2019, 527, 1127–1139. [Google Scholar] [CrossRef] [Green Version]
- Wood, N.J.; Mattiello, T.; Rowe, M.L.; Ward, L.; Perillo, M.; Arnone, M.I.; Elphick, M.R.; Oliveri, P. Neuropeptidergic systems in pluteus larvae of the sea uchin Strongylocentrotus purpuratus: Neurochemical complexity in a "simple" nervous system. Front. Endocrinol. 2018, 9, 628. [Google Scholar] [CrossRef] [Green Version]
- Zueva, O.; Khoury, M.; Heinzeller, T.; Mashanova, D.; Mashanov, V. The complex simplicity of the brittle star nervous system. Front. Zool. 2018, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Formery, L.; Orange, F.; Formery, A.; Yaguchi, S.; Lowe, C.J.; Schubert, M.; Croce, J.C. Neural anatomy of echinoid early juveniles and comparison of nervous system organization in echinoderms. J. Comp. Neurol. 2021, 529, 1135–1156. [Google Scholar] [CrossRef]
- Paganos, P.; Voronov, D.; Musser, J.; Arendt, D.; Arnone, M.I. Single cell RNA sequencing of the Strongylocentrotus purpuratus larva reveals the blueprint of major cell types and nervous system of a non-chordate deuterostome. Elife 2021, 10, e70416. [Google Scholar] [CrossRef]
- Hodin, J.; Heyland, A.; Mercier, A.; Pernet, B.; Cohen, D.L.; Hamel, J.-F.; Allen, J.D.; McAlister, J.S.; Byrne, M.; Cisternas, P.; et al. Culturing echinoderm larvae through metamorphosis. In Echinoderms; Part, A., Kathy, R., Amro, H., Eds.; Academic Press: Amsterdam, The Netherlands, 2019; pp. 125–169. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Zhang, T.; Hamel, J.F.; Mercier, A. Development, settlement, and post-settlement growth. In The Sea Cucumber Apostichopus Japonicus: History, Biology and Aquaculture; Yang, H.S., Hamel, J.F., Eds.; Academic Press: Amsterdam, The Netherlands, 2015; Volume 39, pp. 111–131. [Google Scholar] [CrossRef]
- Burke, R.D.; Brand, D.G.; Bisgrove, B.W. Structure of the nervous system of the auricularia larva Ofparasticopus californicus. Biol. Bull. 1986, 170, 450–460. [Google Scholar] [CrossRef]
- Hökfelt, T.; Broberger, C.; Xu, Z.Q.D.; Sergeyev, V.; Ubink, R.; Diez, M. Neuropeptides—Overview. Neuropharmacology 2000, 39, 1337–1356. [Google Scholar] [CrossRef]
- Watanabe, H.; Fujisawa, T.; Holstein, T.W. Cnidarians and the evolutionary origin of the nervous system. Dev. Growth Differ. 2009, 51, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Elphick, M.R.; Mirabeau, O.; Larhammar, D. Correction: Evolution of neuropeptide signalling systems. J. Exp. Biol. 2018, 221, 19. [Google Scholar] [CrossRef] [Green Version]
- Braubach, O.R.; Dickinson, A.J.; Evans, C.C.; Croll, R.P. Neural control of the velum in larvae of the gastropod, Ilyanassa obsoleta. J. Exp. Biol. 2006, 209, 4676–4689. [Google Scholar] [CrossRef] [Green Version]
- Dyachuk, V.; Odintsova, N. Development of the larval muscle system in the mussel Mytilus trossulus (Mollusca, Bivalvia). Dev. Growth Differ. 2009, 51, 69–79. [Google Scholar] [CrossRef]
- Kiss, T. Diversity and abundance: The basic properties of neuropeptide action in molluscs. Gen. Comp. Endocrinol. 2011, 172, 10–14. [Google Scholar] [CrossRef]
- Veenstra, J.A. Neuropeptide evolution: Neurohormones and neuropeptides predicted from the genomes of Capitella teleta and Helobdella robusta. Gen. Comp. Endocrinol. 2011, 171, 160–175. [Google Scholar] [CrossRef]
- Dickinson, P.S.; Qu, X.; Stanhope, M.E. Neuropeptide modulation of pattern-generating systems in crustaceans: Comparative studies and approaches. Curr. Opin. Neurobiol. 2016, 41, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Kerbl, A.; Conzelmann, M.; Jékely, G.; Worsaae, K. High diversity in neuropeptide immunoreactivity patterns among three closely related species of Dinophilidae (Annelida). J. Comp. Neurol. 2017, 525, 3596–3635. [Google Scholar] [CrossRef] [Green Version]
- Semmens, D.C.; Elphick, M.R. The evolution of neuropeptide signalling: Insights from echinoderms. Brief. Funct. Genom. 2017, 16, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cropper, E.C.; Jing, J.; Vilim, F.S.; Barry, M.A.; Weiss, K.R. Multifaceted expression of peptidergic modulation in the feeding system of Aplysia. ACS Chem. Neurosci. 2018, 9, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, A.; Alexander, J.L.; Swain, M.T.; Webster, S.G.; Wilcockson, D.C. Transcriptomic analysis of crustacean neuropeptide signaling during the moult cycle in the green shore crab, Carcinus maenas. BMC Genom. 2018, 19, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Sun, L.; Wu, J.; Liu, H.; Zheng, L.; Lü, Z.; Chi, C. An FMRFamide neuropeptide in cuttlefish Sepia pharaonis: Identification, characterization, and potential function. Molecules 2020, 25, 1636. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yañez-Guerra, L.A.; Tinoco, A.B.; Escudero Castelán, N.; Egertová, M.; Elphick, M.R. Somatostatin-type and allatostatin-C–type neuropeptides are paralogous and have opposing myoregulatory roles in an echinoderm. Proc. Natl. Acad. Sci. USA 2022, 119, e2113589119. [Google Scholar] [CrossRef]
- Rowe, M.L.; Elphick, M.R. The neuropeptide transcriptome of a model echinoderm, the sea urchin Strongylocentrotus purpuratus. Gen. Comp. Endocrinol. 2012, 179, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Yamano, K.; Fujiwara, A.; Nakamura, A.; Yoshikuni, M. In vitro induction of oocyte maturation in the Japanese sea cucumber Apostichopus japonicus by cubifrin and the developmental ability of the eggs. Fish. Sci. 2013, 79, 823–832. [Google Scholar] [CrossRef]
- Rowe, M.L.; Achhala, S.; Elphick, M.R. Neuropeptides and polypeptide hormones in echinoderms: New insights from analysis of the transcriptome of the sea cucumber Apostichopus japonicus. Gen. Comp. Endocrinol. 2014, 197, 43–55. [Google Scholar] [CrossRef]
- Semmens, D.C.; Mirabeau, O.; Moghul, I.; Pancholi, M.R.; Wurm, Y.; Elphick, M.R. Transcriptomic identification of starfish neuropeptide precursors yields new insights into neuropeptide evolution. Open Biol. 2016, 6, 150224. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.K.; Wang, T.; Suwansa-Ard, S.; Motti, C.A.; Elizur, A.; Zhao, M.; Rowe, M.L.; Hall, M.R.; Elphick, M.R.; Cummins, S.F. The neuropeptidome of the Crown-of-Thorns Starfish, Acanthaster planci. J. Proteom. 2017, 165, 61–68. [Google Scholar] [CrossRef]
- Zandawala, M.; Moghul, I.; Yanez Guerra, L.A.; Delroisse, J.; Abylkassimova, N.; Hugall, A.F.; O’Hara, T.D.; Elphick, M.R. Discovery of novel representatives of bilaterian neuropeptide families and reconstruction of neuropeptide precursor evolution in ophiuroid echinoderms. Open Biol. 2017, 7, 42–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroe, E.B.; Annangudi, S.P.; Wadhams, A.A.; Richmond, T.A.; Yang, N.; Southey, B.R.; Romanova, E.V.; Schoofs, L.; Baggerman, G.; Sweedler, J.V. Exploring the sea urchin neuropeptide landscape by mass spectrometry. J. Am. Soc. Mass Spectrom. 2018, 29, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Suwansa-Ard, S.; Chaiyamoon, A.; Talarovicova, A.; Tinikul, R.; Tinikul, Y.; Poomtong, T.; Elphick, M.R.; Cummins, S.F.; Sobhon, P. Transcriptomic discovery and comparative analysis of neuropeptide precursors in sea cucumbers (Holothuroidea). Peptides 2018, 99, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Talarovicova, A.; Zheng, Y.; Storey, K.B.; Elphick, M.R. Neuropeptide precursors and neuropeptides in the sea cucumber Apostichopus japonicus: A genomic, transcriptomic and proteomic analysis. Sci. Rep. 2019, 9, 8829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chieu, H.D.; Suwansa-Ard, S.; Wang, T.; Elizur, A.; Cummins, S.F. Identification of neuropeptides in the sea cucumber Holothuria leucospilota. Gen. Comp. Endocrinol. 2019, 283, 113229. [Google Scholar] [CrossRef]
- Kato, S.; Tsurumaru, S.; Taga, M.; Yamane, T.; Shibata, Y.; Ohno, K.; Fujiwara, A.; Yamano, K.; Yoshikuni, M. Neuronal peptides induce oocyte maturation and gamete spawning of sea cucumber, Apostichopus japonicus. Dev. Biol. 2009, 326, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Mita, M.; Yoshikuni, M.; Ohno, K.; Shibata, Y.; Paul-Prasanth, B.; Pitchayawasin, S.; Isobe, M.; Nagahama, Y. A relaxin-like peptide purified from radial nerves induces oocyte maturation and ovulation in the starfish, Asterina pectinifera. Proc. Natl. Acad. Sci. USA 2009, 106, 9507–9512. [Google Scholar] [CrossRef] [Green Version]
- Perillo, M.; Arnone, M.I. Characterization of insulin-like peptides (ILPs) in the sea urchin Strongylocentrotus purpuratus: Insights on the evolution of the insulin family. Gen. Comp. Endocr. 2014, 205, 68–79. [Google Scholar] [CrossRef]
- Haraguchi, S.; Ikeda, N.; Abe, M.; Tsutsui, K.; Mita, M. Nucleotide sequence and expression of relaxin-like gonad-stimulating peptide gene in starfish Asterina pectinifera. Gen. Comp. Endocr. 2016, 227, 115–119. [Google Scholar] [CrossRef]
- Lin, M.; Mita, M.; Egertova, M.; Zampronio, C.G.; Jones, A.M.; Elphick, M.R. Cellular localization of relaxin-like gonad-stimulating peptide expression in Asterias rubens: New insights into neurohormonal control of spawning in starfish. J. Comp. Neurol. 2017, 525, 1599–1617. [Google Scholar] [CrossRef]
- Tinoco, A.B.; Semmens, D.C.; Patching, E.C.; Gunner, E.F.; Egertova, M.; Elphick, M.R. Characterization of NGFFYamide signaling in starfish reveals roles in regulation of feeding behavior and locomotory systems. Front. Endocrinol. 2018, 9, 507. [Google Scholar] [CrossRef] [Green Version]
- Yanez-Guerra, L.A.; Delroisse, J.; Barreiro-Iglesias, A.; Slade, S.E.; Scrivens, J.H.; Elphick, M.R. Discovery and functional characterisation of a luqin-type neuropeptide signalling system in a deuterostome. Sci. Rep. 2018, 8, 7220. [Google Scholar] [CrossRef] [PubMed]
- Chieu, H.D.; Turner, L.; Smith, M.K.; Wang, T.; Nocillado, J.; Palma, P.; Suwansa-Ard, S.; Elizur, A.; Cummins, S.F. Aquaculture breeding enhancement: Maturation and spawning in sea cucumbers using a recombinant relaxin-like gonad-stimulating peptide. Front. Genet. 2019, 10, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiyamoon, A.; Tinikul, R.; Nontunha, N.; Chaichotranunt, S.; Poomtong, T.; Sobhon, P.; Tinikul, Y. Characterization of TRH/GnRH-like peptides in the sea cucumber, Holothuria scabra, and their effects on oocyte maturation. Aquaculture 2020, 518, 734814. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, L.; Fan, X.; Guo, X.; Liu, X.; Yang, H. The effect of pedal peptide-type neuropeptide on locomotor behavior and muscle physiology in the sea cucumber Apostichopus japonicus. Front. Physiol. 2020, 11, 559348. [Google Scholar] [CrossRef]
- Wang, T.; Cao, Z.; Shen, Z.; Yang, J.; Chen, X.; Yang, Z.; Xu, K.; Xiang, X.; Yu, Q.; Song, Y. Existence and functions of a kisspeptin neuropeptide signaling system in a non-chordate deuterostome species. Elife 2020, 9, e53370. [Google Scholar] [CrossRef]
- Zhang, Y.; Yanez Guerra, L.A.; Egertova, M.; Zampronio, C.G.; Jones, A.M.; Elphick, M.R. Molecular and functional characterization of somatostatin-type signalling in a deuterostome invertebrate. Open Biol. 2020, 10, 200172. [Google Scholar] [CrossRef]
- Tinoco, A.B.; Barreiro-Iglesias, A.; Guerra, L.A.Y.; Delroisse, J.; Zhang, Y.; Gunner, E.F.; Zampronio, C.G.; Jones, A.M.; Egertová, M.; Elphick, M.R. Ancient role of sulfakinin/cholecystokinin-type signalling in inhibitory regulation of feeding processes revealed in an echinoderm. Elife 2021, 10, e65667. [Google Scholar] [CrossRef]
- Beer, A.J.; Moss, C.; Thorndyke, M. Development of serotonin-like and SALMFamide-like immunoreactivity in the nervous system of the sea urchin Psammechinus miliaris. Biol. Bull. 2001, 200, 268–280. [Google Scholar] [CrossRef]
- Mayorova, T.D.; Tian, S.; Cai, W.; Semmens, D.C.; Odekunle, E.A.; Zandawala, M.; Badi, Y.; Rowe, M.L.; Egertova, M.; Elphick, M.R. Localization of neuropeptide gene expression in larvae of an echinoderm, the starfish Asterias rubens. Front. Neurosci. 2016, 10, 553. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.R.; Paganos, P.; Benvenuto, G.; Arnone, M.I.; Oliveri, P. Post-metamorphic skeletal growth in the sea urchin Paracentrotus lividus and implications for body plan evolution. EvoDevo 2021, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Black, M.M.; Keyser, P. Acetylation of alpha-tubulin in cultured neurons and the induction of alpha-tubulin acetylation in PC12 cells by treatment with nerve growth factor. J. Neurosci. 1987, 7, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Cáceres, A. The expression of acetylated microtubules during axonal and dendritic growth in cerebellar macroneurons which develop in vitro. Dev. Brain Res. 1989, 49, 205–213. [Google Scholar] [CrossRef]
- Gavilán, B.; Perea-Atienza, E.; Martínez, P. Xenacoelomorpha: A case of independent nervous system centralization? Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, S.; Loesel, R.; Purschke, G.; Schmidt-Rhaesa, A.; Scholtz, G.; Stach, T.; Vogt, L.; Wanninger, A.; Brenneis, G.; Döring, C. Invertebrate neurophylogeny: Suggested terms and definitions for a neuroanatomical glossary. Front. Zool. 2010, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Zieger, E.; Candiani, S.; Garbarino, G.; Croce, J.C.; Schubert, M. Roles of retinoic acid signaling in shaping the neuronal architecture of the developing amphioxus nervous system. Mol. Neurobiol. 2018, 55, 5210–5229. [Google Scholar] [CrossRef]
- Boyko, A.V.; Girich, A.S.; Eliseikina, M.G.; Maslennikov, S.I.; Dolmatov, I.Y. Reference assembly and gene expression analysis of Apostichopus japonicus larval development. Sci. Rep. 2019, 9, 1131. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Chen, F.Z.; You, L.J.; Yang, F.; Wang, L.N.; Guo, X.Q.; Gao, F.; Hua, C.; Tan, C.; Fang, L.; Shan, R.Q.; et al. CNGBdb: China National GeneBank DataBase. Hereditas 2020, 42, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Chen, F.; Gao, F.; Li, L.; Liu, K.; You, L.; Hua, C.; Yang, F.; Liu, W.; Peng, C.; et al. CNSA: A data repository for archiving omics data. Database 2020, 2020, baaa055. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, M.; Yin, Y.; Storey, K.B. MiR-200-3p is potentially involved in cell cycle arrest by regulating cyclin a during aestivation in Apostichopus japonicus. Cells 2019, 8, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Chen, M.; Wang, T.; Sun, L.; Xu, D.; Yang, H. Selection of reference genes for qRT-PCR analysis of gene expression in sea cucumber Apostichopus japonicus during aestivation. Chin. J. Oceanol. Limnol. 2014, 32, 1248–1256. [Google Scholar] [CrossRef]
- Nakajima, Y.; Burke, R.D.; Noda, Y. The structure and development of the apical ganglion in the sea urchin pluteus larvae of Strongylocentrotus droebachiensis and Mespilia globulus. Dev. Growth Differ. 1993, 35, 531–538. [Google Scholar] [CrossRef]
- Chee, F.; Byrne, M. Development of the larval serotonergic nervous system in the sea star Patiriella regularis as revealed by confocal imaging. Biol. Bull. 1999, 197, 123–131. [Google Scholar] [CrossRef]
- Hay-Schmidt, A. The evolution of the serotonergic nervous system. Proc. Natl. Acad. Sci. USA 2000, 267, 1071–1079. [Google Scholar] [CrossRef]
- Conzelmann, M.; Offenburger, S.-L.; Asadulina, A.; Keller, T.; Münch, T.A.; Jékely, G. Neuropeptides regulate swimming depth of Platynereis larvae. Proc. Natl. Acad. Sci. USA 2011, 108, E1174–E1183. [Google Scholar] [CrossRef]
- Garner, S.; Zysk, I.; Byrne, G.; Kramer, M.; Moller, D.; Taylor, V.; Burke, R.D. Neurogenesis in sea urchin embryos and the diversity of deuterostome neurogenic mechanisms. Development 2016, 143, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Arikawa, K.; Williams, D.S. Acetylated alpha-tubulin in the connecting cilium of developing rat photoreceptors. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2145–2149. [Google Scholar]
- Hirokawa, T.; Komatsu, M.; Nakajima, Y. Development of the nervous system in the brittle star Amphipholis kochii. Dev. Genes Evol. 2008, 218, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.D. Development of the larval nervous system of the sand dollar, Dendraster excentricus. Cell Tissue Res. 1983, 229, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Lacalli, T.; Gilmour, T.; West, J. Ciliary band innervation in the bipinnaria larva of Pisaster ochraceus. Trans. R. Soc. B Biol. Sci. 1990, 330, 371–390. [Google Scholar] [CrossRef]
- Lacalli, T.C. Ventral neurons in the anterior nerve cord of amphioxus larvae. II. Further data on the pacemaker circuit. J. Morphol. 2003, 257, 212–218. [Google Scholar] [CrossRef]
- Lacalli, T.C. Protochordate body plan and the evolutionary role of larvae: Old controversies resolved? Can. J. Zool. 2005, 83, 216–224. [Google Scholar] [CrossRef]
- Jekely, G.; Melzer, S.; Beets, I.; Kadow, I.C.G.; Koene, J.; Haddad, S.; Holden-Dye, L. The long and the short of it—A perspective on peptidergic regulation of circuits and behaviour. J. Exp. Biol. 2018, 221, jeb166710. [Google Scholar] [CrossRef] [Green Version]
- Odekunle, E.A.; Semmens, D.C.; Martynyuk, N.; Tinoco, A.B.; Garewal, A.K.; Patel, R.R.; Blowes, L.M.; Zandawala, M.; Delroisse, G.; Slade, S.E.; et al. Ancient role of vasopressin/oxytocin-type neuropeptides as regulators of feeding revealed in an echinoderm. BMC Biol. 2019, 17, 60. [Google Scholar] [CrossRef] [Green Version]
- Elphick, M.R. The protein precursors of peptides that affect the mechanics of connective tissue and/or muscle in the echinoderm Apostichopus japonicus. PLoS ONE 2012, 7, e44492. [Google Scholar] [CrossRef]
- Lin, M.; Egertová, M.; Zampronio, C.G.; Jones, A.M.; Elphick, M.R. Pedal peptide/orcokinin-type neuropeptide signaling in a deuterostome: The anatomy and pharmacology of starfish myorelaxant peptide in Asterias rubens. J. Comp. Neurol. 2017, 525, 3890–3917. [Google Scholar] [CrossRef]
- Lin, M.; Egertova, M.; Zampronio, C.G.; Jones, A.M.; Elphick, M.R. Functional characterization of a second pedal peptide/orcokinin-type neuropeptide signaling system in the starfish Asterias rubens. J. Comp. Neurol. 2018, 526, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Whittington, I.D.; Cribb, B.W. Adhesive secretions in the Platyhelminthes. Adv. Parasitol. 2001, 48, 101–224. [Google Scholar] [CrossRef] [PubMed]
- Flammang, P.; Santos, R.; Haesaerts, D. Echinoderm adhesive secretions: From experimental characterization to biotechnological applications. In Echinodermata; Progress in Molecular and Subcellular Biology (Marine Molecular Biotechnology); Matranga, V., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 39, pp. 201–220. [Google Scholar] [CrossRef]
- Terenina, N.; Kreshchenko, N.; Mochalova, N.; Movsesyan, S. Serotonin and neuropeptide FMRFamide in the attachment organs of trematodes. Helminthologia 2018, 55, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieger, E.; Robert, N.S.; Calcino, A.; Wanninger, A. Ancestral role of ecdysis-related neuropeptides in animal life cycle transitions. Curr. Biol. 2021, 31, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Mulrenin, E.M.; Witkin, J.W.; Silverman, A.J. Embryonic development of the gonadotropin-releasing hormone (GnRH) system in the chick: A spatio-temporal analysis of GnRH neuronal generation, site of origin, and migration. Endocrinology 1999, 140, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Gurbuz, A.S.; Gode, F.; Uzman, M.S.; Ince, B.; Kaya, M.; Ozcimen, N.; Ozcimen, E.E.; Acar, A. GnRH agonist triggering affects the kinetics of embryo development: A comparative study. J. Ovarian Res. 2016, 9, 22. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Cong, X.; Liu, H.; Wang, Y.; Storey, K.B.; Chen, M. Nervous System Development and Neuropeptides Characterization in Embryo and Larva: Insights from a Non-Chordate Deuterostome, the Sea Cucumber Apostichopus japonicus. Biology 2022, 11, 1538. https://doi.org/10.3390/biology11101538
Zheng Y, Cong X, Liu H, Wang Y, Storey KB, Chen M. Nervous System Development and Neuropeptides Characterization in Embryo and Larva: Insights from a Non-Chordate Deuterostome, the Sea Cucumber Apostichopus japonicus. Biology. 2022; 11(10):1538. https://doi.org/10.3390/biology11101538
Chicago/Turabian StyleZheng, Yingqiu, Xiao Cong, Huachen Liu, Yixin Wang, Kenneth B. Storey, and Muyan Chen. 2022. "Nervous System Development and Neuropeptides Characterization in Embryo and Larva: Insights from a Non-Chordate Deuterostome, the Sea Cucumber Apostichopus japonicus" Biology 11, no. 10: 1538. https://doi.org/10.3390/biology11101538