
Quorum Sensing in ESKAPE Bugs: A Target for Combating Antimicrobial Resistance and Bacterial Virulence

1
Department of Medical Technology, School of Allied Health Sciences, Walailak University, Nakhon Si Thammarat 80160, Thailand
2
Research Center in Tropical Pathobiology, Walailak University, Nakhon Si Thammarat 80160, Thailand
3
Department of Microbiology and Immunology, Faculty of Tropical Medicine, Mahidol University, Bangkok 10400, Thailand
4
Biomedical Research Incubator Unit, Department of Research, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok 10700, Thailand
*
Author to whom correspondence should be addressed.
Biology 2022, 11(10), 1466; https://doi.org/10.3390/biology11101466
Submission received: 31 August 2022
/
Revised: 22 September 2022
/
Accepted: 30 September 2022
/
Published: 6 October 2022
(This article belongs to the Special Issue Bacterial Biofilms in Complex Infection Environments: Challenges and Novel Therapeutic Perspectives)
Abstract
:Simple Summary
Quorum sensing in ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) bacteria denotes a global threat to public health. The acquisition of antimicrobial resistance genes, virulence production, and biofilm formation by ESKAPE pathogens has reduced the treatment options for serious infections. QS has been well recognized as being involved in the pathogenesis and antibiotic resistance. More understanding of QS mechanistic would also aid in the prediction of underlying or even unknown mechanisms of antimicrobial resistance and bacterial pathogenesis. In this review, we describe the known antibiotic resistance and pathogenesis caused by QS as well as the strategies to control QS in these pathogens.
Abstract
A clique of Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp. (ESKAPE) bugs is the utmost causative agent responsible for multidrug resistance in hospital settings. These microorganisms employ a type of cell–cell communication termed ‘quorum sensing (QS) system’ to mediate population density and synchronously control the genes that modulate drug resistance and pathogenic behaviors. In this article, we focused on the present understanding of the prevailing QS system in ESKAPE pathogens. Basically, the QS component consisted of an autoinducer synthase, a ligand (e.g., acyl homoserine lactones/peptide hormones), and a transcriptional regulator. QS mediated expression of the bacterial capsule, iron acquisition, adherence factors, synthesis of lipopolysaccharide, poly-N-acetylglucosamine (PNAG) biosynthesis, motility, as well as biofilm development allow bacteria to promote an antimicrobial-resistant population that can escape the action of traditional drugs and endorse a divergent virulence production. The increasing prevalence of these harmful threats to infection control, as well as the urgent need for effective antimicrobial strategies to combat them, serve to highlight the important anti-QS strategies developed to address the difficulty of treating microorganisms.
1. Introduction
Bacterial quorum sensing (QS) is the intercellular communication system among microorganisms. This phenomenon is mediated by their specific hormone-like signaling molecules called ‘auto-inducers’, which interact with their cognate receptors, thus allowing bacteria to sense the population densities, orchestrate gene expression, and regulate several physiological activities, such as the expression of antibiotic resistance, virulence determinants, motility, conjugation, plasmid transfer, biofilm formation, and interactions with eukaryotic host cells [1,2,3,4]. Recently, ESKAPE organisms, which encompass six leading causative agents of nosocomial infections (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.), were shown to be capable of ‘escaping’ conventional antimicrobial treatments, thus evolving from microorganisms with multidrug resistance to those with pan-drug resistance and becoming ‘superbugs’ [5].
Virulence factors are the molecules that assist the bacterium in colonizing the host and initiating diseases. Several virulence determinants are involved in the step of attachment, such as pili, fimbria, fibronectin-binding proteins, PNAG, and adhesins. Additionally, in the step of invasion and tissue damage, many exoenzymes and toxins interplay in this mechanism, including glycohydrolases (hyaluronidase), nucleases (DNase), phospholipases (phospholipase C), proteases (collagenase, gelatinase, elastase, oligopeptide permease), lipid A, cytolysins, exotoxins, alpha-toxins, toxic shock syndrome toxin, hemolysins, leucocidins, coagulase, and pigments, etc. [1,2,3,4]. Antibiotic resistance among these pathogens has been considered a major public health concern worldwide [6,7]. The drug resistance mechanisms in ESKAPE organisms include target alteration, enzyme inactivation, porin loss, efflux pump, and biofilm formation [8]. Several studies have found that the QS system regulates antimicrobial resistance and pathogenicity in these pathogens [9]. Generally, the expression of drug resistance phenotypes through the QS system occurs according to the following key steps: synthesis of QS signaling molecules, release of signal ligands to the milieu, sensing of the signal molecules at a high level of cell density, binding to the transcriptional regulator or R-protein, transporting the ligand–receptor complex from the cell, binding of the complex to the promoter region of the target gene, and transcription of drug resistance-associated genes, leading to phenotypic manifestation [10].
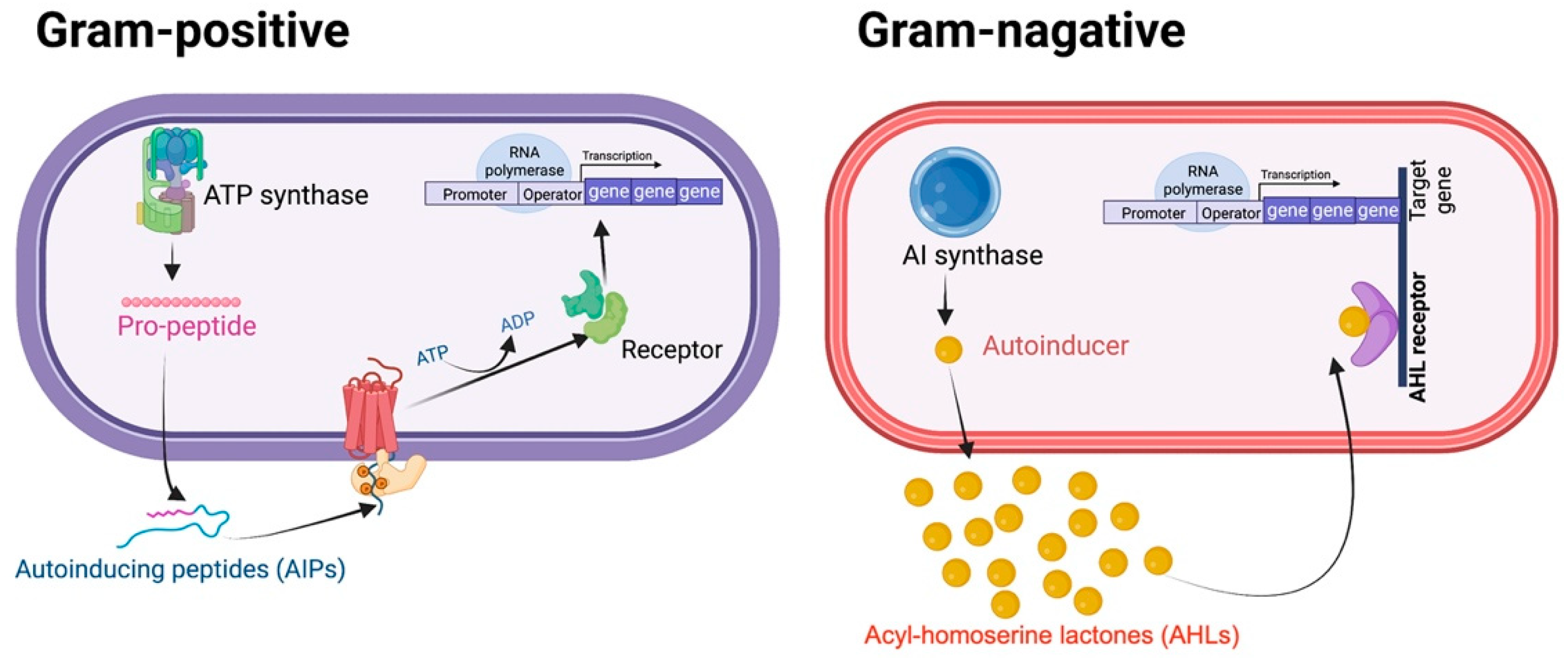
In general, Gram-positive and Gram-negative bacteria use QS to control a diverse array of vital bacterial behaviors and biological characteristics. The QS circuit consists of main components, including autoinducer synthases, signaling ligands (autoinducers or self-inducers), transcriptional regulators, and target genes (Figure 1). Gram-positive bacteria establish oligopeptides, so-called ‘autoinducing peptides’ (AIPs), which are short peptide chains, as the major autoinducers (AIs) [11]. The mature AIPs can interact with the transmembrane histidine kinase receptor, resulting in autophosphorylation of the transcriptional regulator, consequently triggering the expression of the target gene. In contrast, Gram-negative organisms use acylated homoserine lactones as signal molecules [12]. The derivatives of N-acyl homoserine lactones (AHLs), which are classified as autoinducers-1 (AI-1), are predominantly renowned for mediating intraspecies communication among Gram-negative bacteria. Typically, AHL molecules diverge in the length of their carbon chains. Short-chain AHLs (with 4–8 carbon atoms) are able to simply diffuse across the bacterial cell membrane, whereas the long-chain AHLs (with 10–14 carbon atoms) require an active transport system to facilitate their transit through the membrane. Moreover, furanosyl borate diester, which is categorized as an autoinducer-2 (AI-2), is found in both Gram-positive and Gram-negative bacteria and is responsible for interspecies coordination among them [13,14]. To date, several QS inhibitors have been developed to block and/or abolish QS signaling and subsequently prevent pathogenic activities and drug resistance phenotypes. This review focuses on the QS-mediated drug resistance in ESKAPE pathogens and the current state of anti-QS approaches as a potential alternative to traditional therapy in the future.
2. QS-Mediated Drug Resistance and Bacterial Virulence in ESKAPE Pathogens
Nosocomial ESKAPE organisms employ AI synthases to generate and accumulate QS signal molecules, thus synchronizing their specific receptors. Subsequently, they stimulate the expression of target genes and acquire pathogenic and antimicrobial characteristics. Typically, Gram-positive bacteria utilize small peptide signals for sensing through the two-component histidine kinase system, which is mainly of the RNPP (Rap, NprP, PlcR, and PrgX regulatory proteins) and Rgg (transcriptional regulator of glucosyltransferase) type, whereas most Gram-negative bacteria control QS-related gene expression via the N-acyl homoserine lactone (AHL)-mediated LuxR–LuxI homologous system [1,15].
2.1. Enterococcus spp.
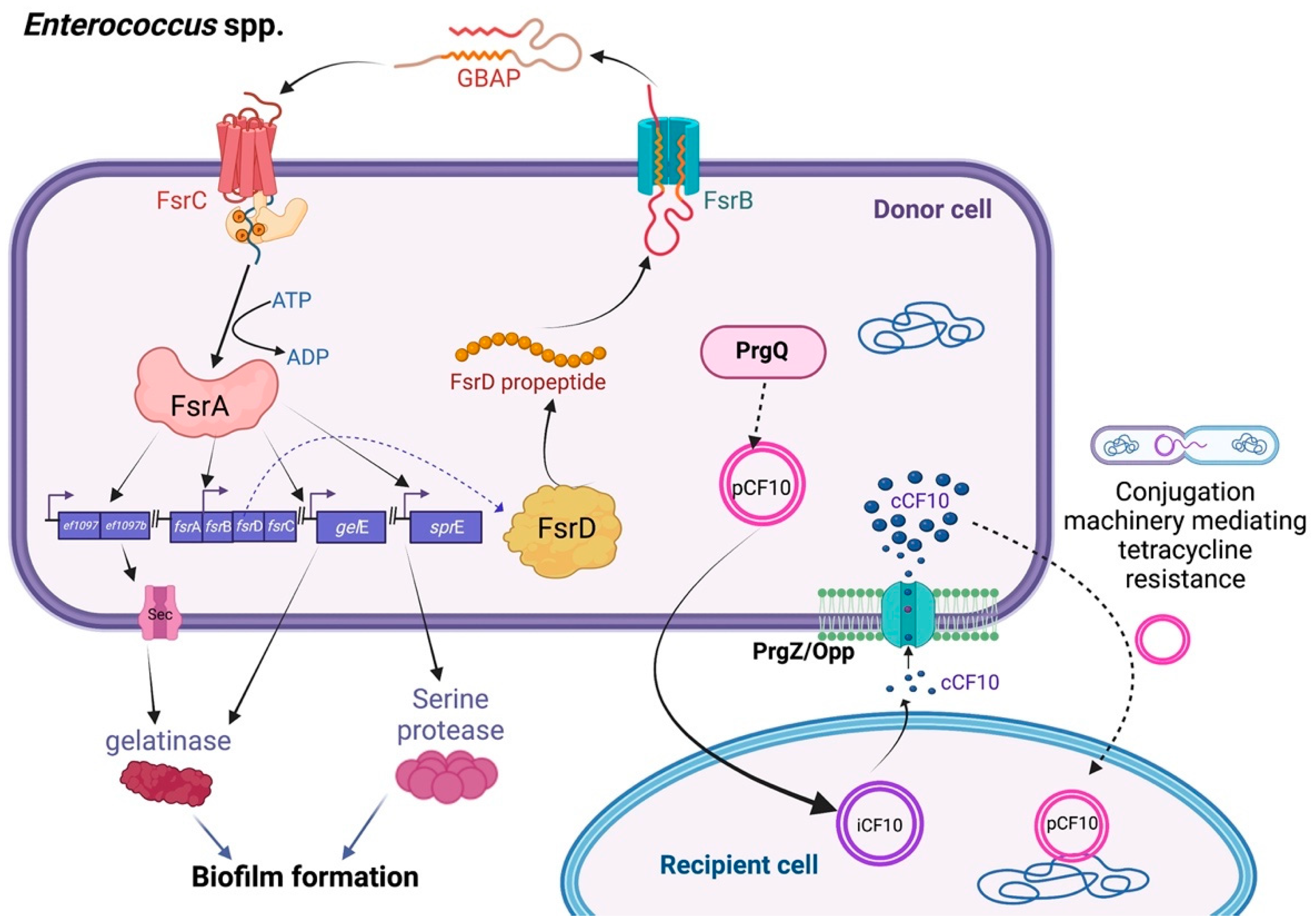
Enterococcus spp., such as E. faecalis and E. faecium, are common infectious pathogens that produce a variety of virulence genes encoding bacteriocins, aggregation factors, and drug resistance determinants [16]. The antibiotic resistance conjugation transfer is mediated by signaling by common QS peptide pheromones functioning as AIs, such as cCF10 (with the sequence ‘LVTLVEV’) produced by PrgQ. The temporary donor cells harbor resistance genes on pCF10, which is the tetracycline resistance conjugative plasmid, in response to the small signal peptides secreted by recipient cells. These AIs can pass through the PrgZ–Opp complex and the ATP-binding cassette transport system, to then accumulate in the extracellular environment and sense the ligand-binding protein, thus upregulating the conjugation genes that trigger conjugation-related mating functions throughout a population. After the bacterial recipients acquire plasmids, they confer the tetracycline resistance phenotype [17,18]. Similarly, the bacteria occupy an octapeptide pheromone known as cAD1 (with the sequence LFSLVLAG) for QS synchronization. Subsequently, the transfer of the mobile elements called pAD1, which encode a cytolysin (hemolysin/bacteriocin) and an aggregation constituent, has been shown to contribute to pathogenicity [19].
Moreover, enterococci, especially E. faecium, possess other AIs, including a cyclic peptide molecule called gelatinase, biosynthesis-activating pheromone (GBAP). These signal molecules can cooperate with their transmembrane cognate receptors, FsrC, thus facilitating the QS activity [20]. The faecal streptococci regulator (Fsr) locus is encoded by the fsrA, fsrB, and fsrC genes via GBAP (Figure 2). This controls the expression of gelatinase, biofilm formation, and the production of serine proteases and enterocin O16 (cytolysin). The LuxS/autoinducer-2 (AI-2) system, which is another QS regulatory system, has also been proposed to play a role in the persistence of infections and biofilm development through interspecies communication. However, the details of this mechanism warrant further exploration [21,22].
2.2. Staphylococcus aureus
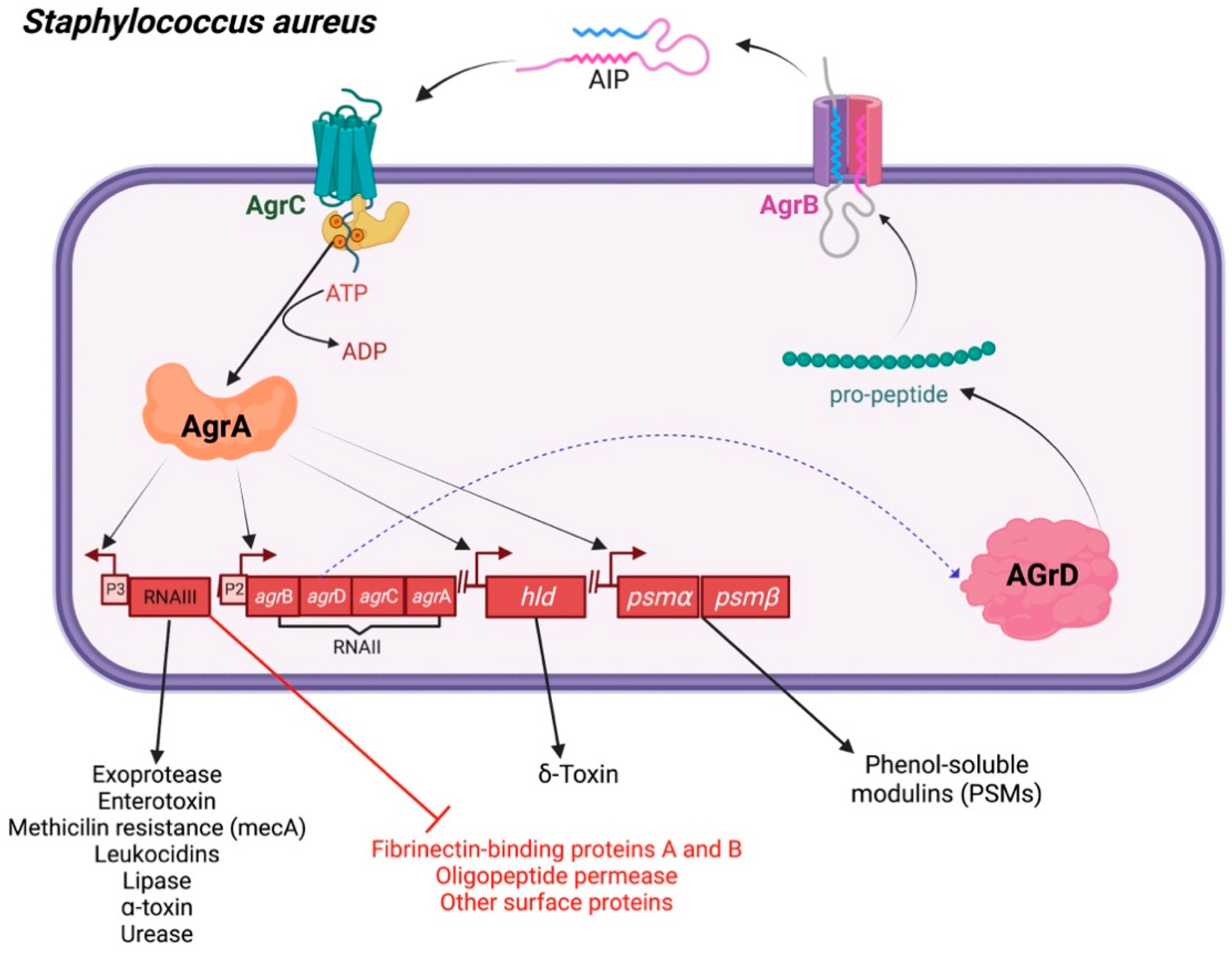
The QS system potentiates S. aureus to cause disease according to the expression of various adhesins, toxins, and substances that disrupt the host’s immune system [23]. Most Gram-positive bacteria utilize a two-component QS cascade, which is controlled by an additional gene regulator (agr) locus [23]. The agr system comprises RNA, RNAII, and RNAIII transcripts. The RNAII operon comprises agr genes, such as agrB, agrD, agrC, and agrA. The signaling pathway is initiated by the production of an agrD-encoding peptide, which is then modified by an integral membrane protein named ‘AgrB’. The altered peptide acts as an ultimate AIP [24,25]. This two-component machinery, composed of AgrA and AgrC, as well as of an AIP-binding domain, participates in histidine kinase transduction. Stimulation of the two-component system turns on the RNAII operon, which acts as a regulator of RNAIII transcription [26]. RNAIII can trigger the generation of α-toxin, while inhibiting the expression of the fibronectin-binding proteins A and B, peptide A, oligopeptide permease, coagulase, and other surface proteins (Figure 3).
Bacteria form biofilms as part of their survival mechanisms. The biofilm layer promotes the bacteria’s tolerance to antibiotics and chemical disinfectants. Molecular mechanisms that have been shown to enhance establishment of bacterial biofilm indicate that biofilm virulence factors likely arise through resistance to phagocytosis and other host immune defense mechanisms resulting in persistence in the host niche rendering the chronic infections [3,4,23]. To date, several reports have addressed the regulation of biofilm formation mediated by the LuxS/AI-2 system. In S. aureus, the homolog of luxS, which encodes AI-2 synthase, promotes ribosome binding factor (rbf) transcription, consequently increasing the production of polysaccharide intercellular adhesion and adhesion, which positively regulate biofilm formation and have been investigated in other bacteria, e.g., Escherichia coli, Salmonella enterica, Klebsiella pneumoniae, and Streptococcus pneumoniae [27].
2.3. Klebsiella pneumoniae
K. pneumoniae primarily exploits the AI-1 QS system for conducting its cellular processes. Due to the lack of luxI gene homologs for LuxI synthase establishment, this strain encodes a LuxR receptor (acronym, ‘SdiA’), but does not produce AHLs [28,29]. This orphan receptor responds to the exogenous AHL produced by other bacterial strains, thus regulating cell division and the expression of virulence factors, such as fimbriae expression, biofilm formation, and production of QS autoinducers in K. pneumoniae [30].
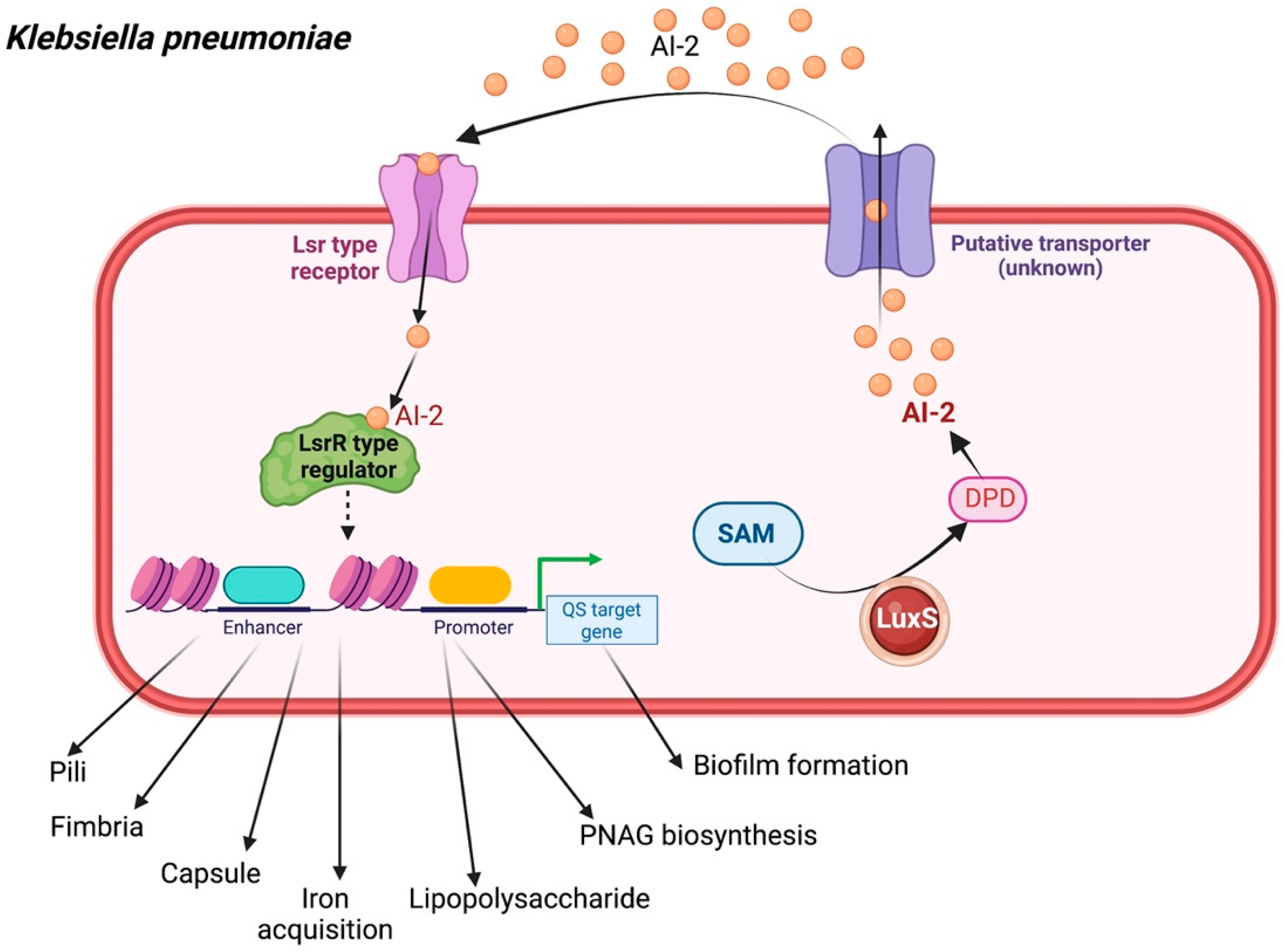
Similar to other ESKAPE organisms, K. pneumoniae also engages in classical QS-mediated bacterial negotiation via the furanosyl borate diester (AI-2) produced by the LuxS synthase and by N-octanoyl homoserine lactone (C8-HSL) and N-3-dodecanoyl-L-homoserine lactone (C12-HSL) [31,32]. When AI-2 is mediated by the LuxS system, the signaling molecule is passed through the transmembrane protein and transferred to the bacterial transporter [33]. Once AI-2 is internalized into the cytoplasm and phosphorylated by LsrK, the phospho-AI-2 further abolishes the LsrR repression of the lipolysis-stimulated lipoprotein receptor (lsr) operon, thus resulting in increased AI-2 uptake. As optimal cell density is reached, the extracellular AI-2 binds to cognate receptors and the signal transduction pathway is activated. The results revealed the expression of the bacterial capsule, iron acquisition, adherence factors, and synthesis of lipopolysaccharide (LPS), which promote the pathogenicity of the agent. Moreover, the study demonstrated the expression of poly-β-1,6-N-acetylglucosamine (PNAG) biosynthesis as well as biofilm development, rendering the drug-resistant K. pneumoniae (Figure 4) [34,35,36].
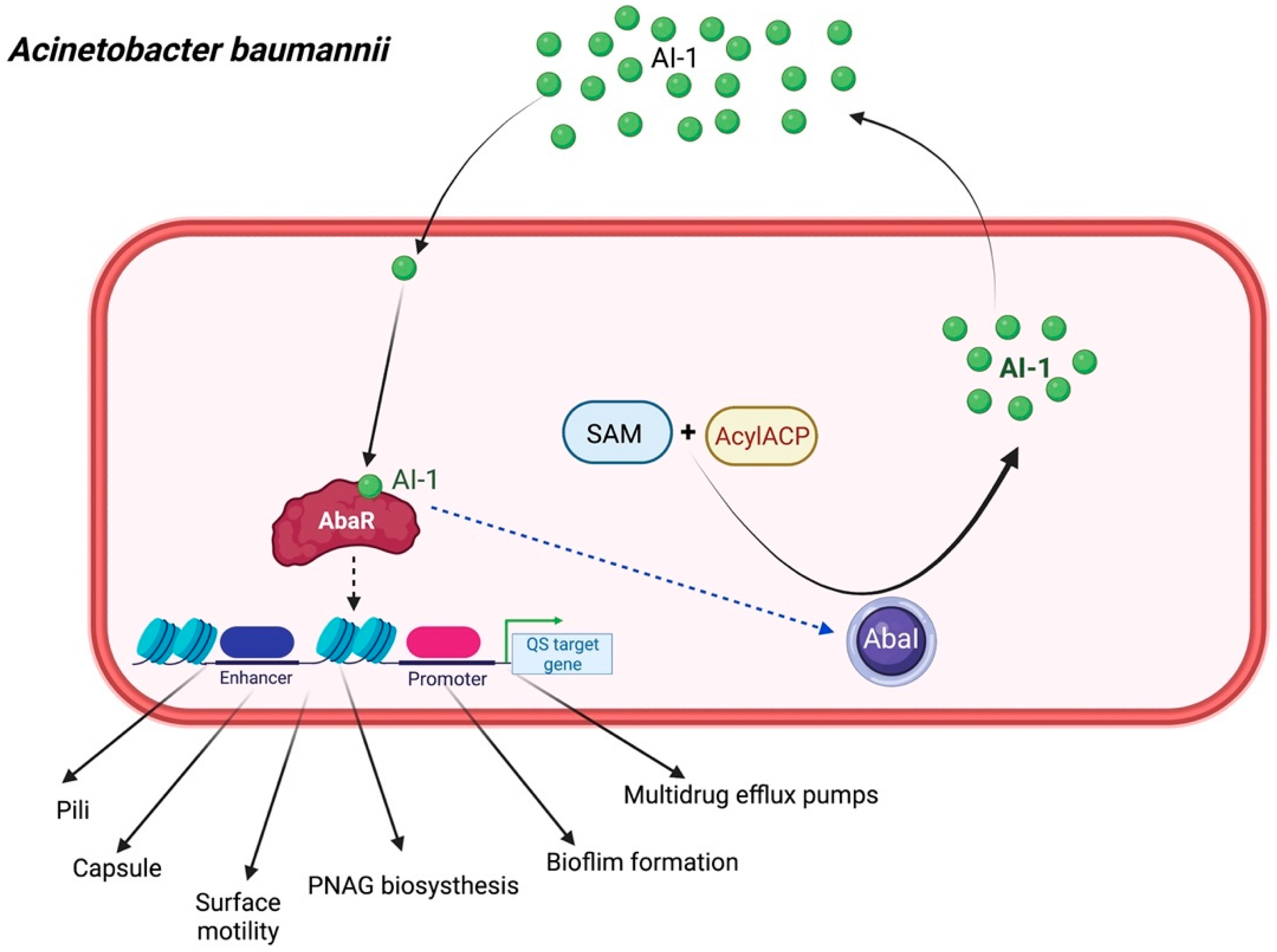
2.4. Acinetobacter baumannii
The single-complex QS machinery in A. baumannii is homologous to the typical LuxI/LuxR system. This machinery mainly comprises AbaI (AI synthase), a signaling AHL molecule, and AbaR (cognate receptor), which is controlled by the abaR/abaI locus [37,38]. When AbaI-generated AHL forms a complex with AbaR, the AbaR–AI complex recognizes a putative lux-box sequence (CTGTAAATTCTTACAG) [39], thus triggering drug resistance, surface motility, and the production of the exopolysaccharide poly-β-1,6-N-acetylglucosamine (PNAG), which is essential for adhesion, aggregation, and biofilm formation (Figure 5) [40]. This organism creates medium-to-long-chain AHLs with their acyl side chains, ranging from C6 to C8 and C10 to C16, such as unsubstituted C10-HSL, C12-HSL, 3-hydroxy-C10-HSL, 3-hydroxy-C12-HSL, unsaturated 3-oxo-C11-HSL, and C14-HSL [41,42]. The most abundant AHL is N-(3-hydroxydodecanoyl)-L-homoserine lactone (3-hydroxy-C12-HSL) [37]. Moreover, the loci located between abaR and abaI encompass a third gene, namely abaM, which controls the expression of an uncharacterized RsaM protein family. Previous evidence showed that AbaM downregulates AHL synthesis in A. baumannii and modulates surface motility and biofilm formation [43]. Moreover, it regulates N-acyl homoserine lactone (AHL)-dependent QS in other beta- and gamma-proteobacteria, such as Burkholderia spp., Halothiobacillus neapolitanus, and Pseudomonas fuscovaginae [44].
Furthermore, many studies have reported the role of the efflux system in the resistance–nodulation–cell division (RND) systems, AdeABC, AdeIJK, and AdeFGH, which share structural similarity with the MexAB pumps of P. aeruginosa. This machinery is used for the transport of QS molecules, virulence factors, and antibiotics in A. baumannii [45,46]. This finding indicated the correlation between efflux-pump-mediated QS and antibiotic resistance in this organism. Basically, iron is required for many physiological activities in pathogens, including DNA replication, transcription, metabolism, and energy generation via respiration. Some reports demonstrated that restricted concentrations of iron can positively regulate QS signaling molecules, thus increasing population persistence and virulence production in the bacteria [38,47,48].
2.5. Pseudomonas aeruginosa
The eradication of P. aeruginosa is frequently challenging because it is resistant to several antibiotics and generates various virulence determinants. The ability of P. aeruginosa to develop a biofilm is another important virulence trait that hampers its removal. Similar to other ESKAPE bacteria, P. aeruginosa uses a small diffusible signal molecule to mediate QS. The predominant QS signals produced by this organism are N-butanoyl-L-homoserine lactone (C4-HSL), 3-oxo-dodecanoyl-L-homoserine lactone (3-oxo-C12-HSL), and 2-heptyl-3-hydroxy-4-quinolone (PQS; Pseudomonas quinolone signal) [28,49,50].
The LasI/LasR and RhlI/RhlR systems drive the production of various virulence elements (such as elastase, pyocyanin pigment, and rhamnolipid biosynthesis) and biofilm development, which triggers the reduction of conventional antibiotic susceptibility [51]. These signal ligands, such as 3-oxo-C12-HSL and C4-HSL, are generated by AI synthases, including LasI and RhlI, respectively. Their cognate receptor proteins are LasR and QscR for 3-oxo-C12-HSL, and RhlR for C4-HSL [52]. In addition, the C4-HSL of P. aeruginosa expands the expression of the MexAB–OprM efflux pump, which confers resistance to traditional beta-lactam drugs [53].
Another QS system that is present in this organism arbitrates a second class of QS signals, including 4-hydroxy-2-alkylquinolines (HAQs) (such as 4-hydroxy-2-heptylquinoline (HHQ) derivatives and 2-heptyl-3,4-dihydroxyquinoline, which is the corresponding dihydroxylated derivative. The HAQ biosynthesis in this pathogen requires a set of genes encoded by the pqsABCDE and phnAB operons. PQS is synthesized by hydroxylation of HHQ by a putative monoxygenase known as PqsH. Both HHQ and PQS act as co-inducing ligands of PqsR, also called multiple virulence factor regulator (MvfR). These activated the QS via their cognate receptor, PqsR [28,52]. Furthermore, PQS plays a role during P. aeruginosa infection by deliberating the expression of virulence factors (for instance, pyocyanin, elastase, lectin, and rhamnolipids; and biofilm development) and provoking the inflammation caused by host immune responses (Figure 6) [54].
2.6. Enterobacter spp.
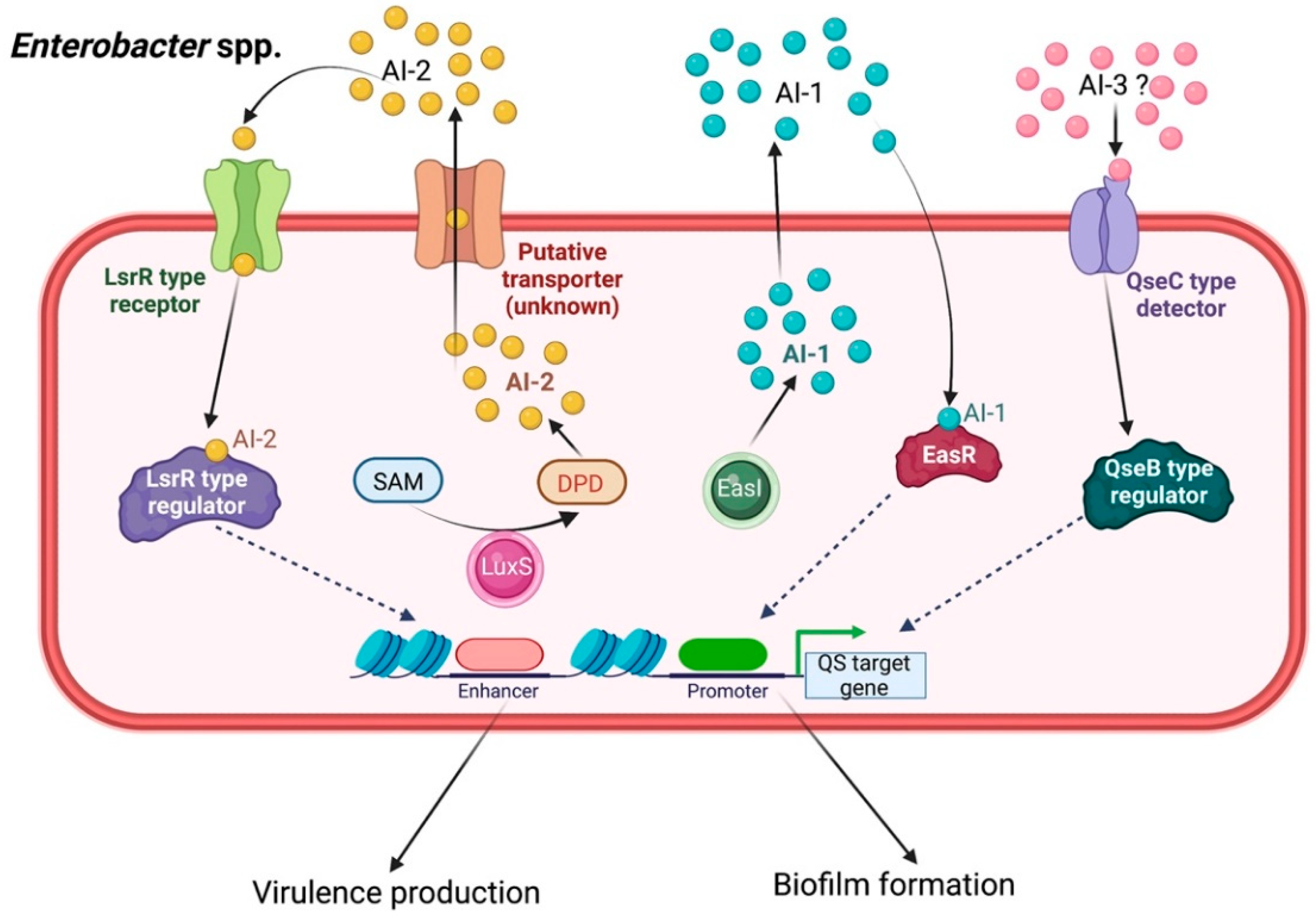
Enterobacter QS signaling, especially in E. aerogenes and E. cloacae, occupies AI-1, AI-2, and AI-3 as the functional signaling ligands [55]. Although little information related to QS-controlled drug resistance and pathogenesis is available in this group of bacteria, several research efforts have been made to elucidate the complicated mechanisms that contribute to the pathogenicity and antimicrobial resistance of diverse Enterobacter genera.
Some evidence has indicated that Enterobacter spp. use C4-HSL and C6-HSLs as QS signals [56]. These are generated by a LuxR homolog, which has been found to decrease bacterial adhesion and downregulate biofilm development [57]. In E. asburiae, the QS pathway is triggered by C4-HSL and C6-HSL, which bind to their cognate EasR receptor protein (the transcriptional regulator luxR homolog), thus triggering QS-associated gene transcription, as well as its related phenotypes and biofilm formation [58].
Intercellular negotiation among Enterobacter spp. also occurs via the AI-2-mediated QS system, as the cognate Lsr-type receptors have been found in strains of E. cloacae, E. cancerogenus, and E. mori [59,60]. Mostly in Enterobacteriaceae, such as E. cloacae and the enterohemorrhagic Escherichia coli O157:H7, the AI-3, epinephrine and norepinephrine, was also reported to modulate QS phenotypes, such as biofilm formation, which is accelerated by the QseC/QseB system (Figure 7) [61].
3. Therapeutic Approaches Targeting QS Systems Counteract Drug Resistance and Virulence in ESKAPE Bugs
Disrupting bacterial QS pathways in ESKAPE bacteria seems to be an attractive broad-spectrum remedial strategy for reducing resistance to antimicrobial agents, silencing bacterial pathogenesis, and promoting susceptibility to host immune defenses without eliciting any evolutionary pressure. To date, several protocols have been proposed as potential anti-QS approaches, such as blocking AI synthases, degrading the AIs, inactivating transcriptional regulators, interfering with the ligand–receptor complex, and incorporation with traditional drugs (Table 1) [62,63].
3.1. Targeting AI Synthase
Several studies have demonstrated that the inhibition of QS signal synthesis can interrupt the initial step of the QS network, which attenuates AHL-mediated virulence and drug resistance phenotypes. In this manner, several studies have reported the use of natural and synthetic compounds that are structural analogues of the substrates for the AHL synthases S-adenosyl methionine (SAM) and acyl-carrier protein (ACP). In P. aeruginosa, sinefungin, butyryl-SAM, and L/D-S-adenosylhomocysteine can attenuate QS-mediated virulence factors and prevent bacterial infection by inhibiting the secretion of AHLs [64,65].
Evidently, triclosan can reduce the production of AHLs by diminishing the production of enoyl-ACP reductase precursors (FabI) in S. aureus [66].
3.2. Sequestration of QS Ligands
The degradation of QS signal molecules in Gram-negative bacteria triggers the inactivation of AHLs, which is mediated by enzymatic activity. The major enzymes identified in many P. aeruginosa research studies include AHL lactonases, acylases, oxidoreductases, and 3-hydroxy-2-methyl-4(1H)-quinolone 2, 4-dioxygenase. AHL lactonases and AHL acylases function by cutting the amide linkage with different lengths of side chains of AHLs and destroying the lactone ring. AHL lactonases, such as lactonase SsoPox, lactonase Aii810, quorum quenching lactonase enzyme AHL-1 (a novel lactonase cloned by bpiB01 and bpiB04), and lactonase AiiK, are found to reduce the production of extracellular proteases and pyocyanin, rhamnolipids, swarming motility, and biofilm formation, and to prevent bacterial infection [67,68,69,70,71,72]. Furthermore, AHL lactonases have been shown to increase bacterial sensitivity to traditional drugs without affecting bacterial metabolic growth [108]. In A. baumannii, the engineered AHL lactonase was also reported to sequester AHL and reduce the A. baumannii biomass-associated biofilms, which increases bacterial sensitivity to antibiotics without affecting the growth of A. baumannii. Many current agents are intended to directly kill pathogenic bacteria by damaging cell membranes or interfering with fundamental protein synthesis. The widespread use of agents has resulted in major microbial resistance problems, and this selection pressure encourages the evolution of microbial resistance. AHL lactonase has less effect on organism development, implying less selection pressure to drive microorganism evolution. This indicates that anti-QS compounds may be used as potential alternatives to traditional medications [109].
Acylases constitute another enzyme type that can block the QS pathway by hydrolyzing the amide bond of AHLs. Previous research found that Aspergillus melleus acylase can degrade C4-LHL, C6-LHL, and 3-oxo-C12-LHL, resulting in decreased pyocyanin synthesis and biofilm formation [73]. Acylase (EC.3.5.1.14) can inactivate AHL inducers, resulting in decreased biofilm biomass [74]. N-acyl homoserine lactone acylase PA2385 can destroy the 3-oxo-C12-HSL and 2-heptyl-3-hydroxy-4 (1H)-quinolone, which lessens elastase and pyocyanin biosynthesis [75].
Regarding oxidoreductases, another enzyme can change the acyl side-chain structure of AHLs, thus interfering with the expression of QS signaling. The BpiB09 oxidoreductase was reported to inhibit the activation of 3-oxo-C12-HSL, causing a reduction in bacterial motility, biofilm formation, and pyocyanin production in P. aeruginosa [76]. Similarly, oxidoreductases immobilized on a glass surface can inhibit bacterial biofilm development and decrease the growth rate in K. pneumoniae [77].
The 3-hydroxy-2-methyl-4 (1H)-quinolone can catalyze the conversion of PQS to N-octanoylanthranilic acid and carbon monoxide, thereby downregulating lectin A, pyocyanin, and rhamnolipid [78]. Dioxygenase has been shown to block the quinolone-mediated QS signals via the degradation of 2-heptyl-3-hydroxy-4 (1H)-quinolone of P. aeruginosa, thus decreasing the generation of pyocyanin, rhamnolipid, and lectin A [78,110].
Another intriguing anti-QS platform arbitrated by antibodies that target the QS signal molecules has also been reported, such as the RS2-1G9QQ antibody, which prevented the stimulation of the mitogen-activated protein kinase p38 and protected murine bone-marrow-derived macrophages from cytotoxic effects [79]. The XYD-11G2 antibody hydrolyses 3-oxo-C12-HSL, leading to the suppression of bacterial QS signals [80]. Moreover, the engineered human single-chain variable fragments inhibit P. aeruginosa 3-oxo-C12-HSL and prevent mammalian cell apoptosis [81].
A previous study of Gram-positive bacteria, including S. aureus, demonstrated that the AP4-24H11 antibody targets autoinducing peptide-4, elicits protective activities in vivo by attenuating pathogenicity in S. aureus-generated-abscess formation in a mouse model, and offers a complete defense against a lethal S. aureus challenge [82]. Furthermore, in vivo studies have demonstrated the inhibitory activity of synthetic RIP (the amide form of the originally isolated one) by reducing S. aureus infections, such as cellulitis, septic arthritis, keratitis, osteomyelitis, and mastitis [83]. The synergistic activity exerted through the combination of synthetic RIP and antibiotics has been reported to act against S. aureus biofilms [84].
3.3. Blocking of QS Transcriptional Regulators
Inactivation of receptors in QS signaling is an effective strategy for deactivating bacterial virulence and infection. Many studies have revealed that, in P. aeruginosa, flavonoids can target the allosteric inhibition of AI-binding receptors, including LasR and RhlR, which affects the transcription of QS-controlled target promoters and suppresses virulence factor production [111]. N-decanoyl-L-homoserine benzyl ester can activate a QS control repressor, therefore attenuating protease and elastase activities, swarming motility, and biofilm development [85]. LasR and RhlR can be inhibited by meta-bromo-thiolactone, thus disarming the production of pyocyanin and biofilm formation [86]. The AHL ligands include A4, 4-bromophenyl-PHL B7, 4-iodo PHL C10, and 3-nitro PHL C14, which bind to TraR, LasR, and LuxR, thus inhibiting the production of virulence factors [87].
Moreover, several researchers have reported the effectiveness of virstatin, which is a tiny organic compound, as an inhibitor that prevents A. baumannii from expressing anoR, a LuxR-type regulator, which is a homologue of the AbaI/AbaR regulatory system. The effectiveness of virstatin as a T4P pili system biogenesis inhibitor in preventing bacterial movement and initiating biofilm formation has been demonstrated [88].
3.4. Alternative Approach for Inhibiting QS Using Probiotics
The treatment or primary prevention of bacterial infections using probiotics has been demonstrated to be successful. However, the effectiveness of these agents has a strain- and disease-specific nature. The benefit of using probiotics as therapeutic agents is that these living microbes frequently have multiple modes of action, such as expressing proteases that specifically destroy Toxin A, inference with toxin attachment sites, immune regulation, and other mechanisms that may include inhibition of QS systems [112].
Studies have shown that certain probiotic strains may interfere with the QS system of ESKAPE bacteria. Another study has shown that Lactobacillus plantarum PA100 can prevent the induction of P. aeruginosa virulence factors by targeting AHL. According to this investigation, the development of biofilm, elastase, and AHL could be inhibited by the acid filtrate and the neutralized filtrate of L. plantarum PA100 [89]. Moreover, C4-HSL and 3-oxo-C12-HSL of P. aeruginosa can be destroyed by cell extracts of L. crustorum ZHG 2-1, which has been reclassified as Companilactobacillus crustorum. Thus, suppression of biofilm formation, loss of swarming and swimming motilities, and reduction of virulence factors (chitinase and protease) were also noted, without altering bacterial growth [90]. In addition, the metabolites of lactic acid bacteria, such as L. lactis NCDC 309, L. rhamnosus MTCC 5897, L. rhamnosus MTCC 5857, L. fermentum MTCC 5898, L. acidophilus NCDC 15, L. delbrueckii subsp. lactis, and L. plantarum NCDC 372, were found to effectively hinder elastase and biofilm production, as well as lasI and rhlI gene expression in P. aeruginosa. It is interesting to note that these supernatants efficiently lower AHL synthesis [91].
In Gram-positive bacteria, the L. reuteri RC-14 strain, which acts as a probiotic and is used to treat toxic shock syndrome, creates the small molecules cyclo (L-Phe-L-Pho) and cyclo (L-Tyr-L-Pro), thus disrupting the QS system of toxic strains of S. aureus. The TSST-1 gene, which encodes the toxin linked to toxic shock syndrome, was suppressed by this interference [91]. The biosurfactants generated by L. plantarum and P. acidilactici decrease the expression of AI-2 in a dose-dependent manner, as well as the cidA, icaA, dltB, agrA, sortaseA, and sarA genes, which are related to biofilm development in S. aureus [93].
3.5. Alternative Approach for Inhibiting QS Using Plant Extracts
Currently, various plant extracts, including the previously mentioned flavonoids and phenolic acids, display potent action against ESKAPE QS [94]. In addition, the previous study showed that eugenol suppressed the synthesis of virulence factors such as elastase, pyocyanin, and biofilm formation in P. aeruginosa via the las and pqs QS systems [95,96,97]. Eugenol also prevented the formation of biofilms and the expression of QS synthase genes, particularly lasI, rhlI, and the rhlA gene [97]. In methicillin-resistant S. aureus, this compound also limits the protease enzymes and pigment production [98]. Another study found that carvacrol (2-methyl-5-(1-methylethyl)-phenol) was effective against biogenesis and QS which lessens the pathogenicity of P. aeruginosa by blocking lasI expression and reducing lasR expression, as well as biofilm growth and surface motility [99,100].
A diterpene known as phytol has shown anti-QS activity. In this manner, this substance suppresses flagella mobilization, restricts the formation of pyocyanin, and inhibits the establishment of the biofilm in P. aeruginosa PAO1 [102,103]. Another terpene with anti-QS properties is sesquiterpene lactones. This compound diminished the QS mediators in P. aeruginosa ATCC 27,853 [104]. Similar to the previous example, oleanolic aldehyde coumarate showed inhibitory actions against P. aeruginosa biofilm by downregulation of lasI/lasR, rhlI/rhlR, and gacA expression [105]. Additionally, other terpenoids, including linalool, hindered the generation of A. baumannii biofilms and altered this strain’s surface adhesion. This trait is associated with linalool’s interference with the QS system [106,107].
Several studies have looked into the anti-QS properties of quercetin and its derivatives [113,114,115,116]. The substance has antibiofilm properties against P. aeruginosa strain PAO1 and inhibits the synthesis of virulence factors such as pyocyanin, protease, and elastase by lowering lasI, lasR, rhlI, and rhlR gene expression levels.
4. Clinical Applications and Future Perspectives
Many anti-QS compounds have been suggested by various researchers to control pathogenesis, infection, and antibiotic resistance in pathogenic organisms, including ESKAPE. Nevertheless, the tolerance, effectiveness, and safety of therapeutic regimens for clinical usage should be investigated. In recent years, the pyrimidine analog 5-fluorouracil (5-FU) is a potent quorum-quencher, inhibiting AI-2 production of MRSA, Staphylococcus epidermidis, E. coli, and Vibrio harveyi, and has gained popularity as an antimetabolite used both topically and systemically for the treatment of actinic keratoses and neoplastic disorders [114]. The anti-QS compounds showed growth inhibition against both Gram-positive and Gram-negative bacteria, and the 5-FU was utilized as an antimetabolite by coating central venous catheters to prevent colonization or infection of a patient’s implanted medical device [115]. When utilized in critically ill patients, central venous catheters externally coated with 5-FU were found to be a safe and effective alternative to catheters externally coated with chlorhexidine and silver sulfadiazine. Another study found that azithromycin, which decreases QS-regulated virulence in P. aeruginosa, could help patients prevent ventilator-associated pneumonia (VAP). The findings suggest that virulence inhibition is a potential antimicrobial strategy due to azithromycin dramatically reducing the high risk of rhamnolipid-dependent VAP [116].
This innovative nonantibiotic therapy, which can suppress the expression of genes related to bacterial pathogenesis, prevent infection, and lessen the possibility of drug resistance in bacterial cells, has been gaining popularity in recent years. Recent studies have discovered numerous anti-QS compounds that can be used to regulate the pathogenic phenotypes of the majority of bacteria and to lessen the pathological harm in a variety of animal infection models [115,117,118,119]. However, anti-QS compounds may be toxic, and their therapeutic impact is not as permanent as that of antibiotics, limiting their widespread adoption. Combining anti-QS medicines with traditional antibiotics can considerably boost therapeutic medication efficacy. In contrast, the development calls for a mechanistic understanding of the QS system’s operation as well as an understanding of its molecular pathways to be used as the main application technique of anti-QS drugs for the treatment of bacterial diseases in the future.
5. Conclusions
The global health problem caused by the introduction of several drug-resistant strains of ESKAPE nosocomial pathogens is extremely concerning. The QS signaling among these pathogens accompanies and elicits antimicrobial susceptibility and the production of bacterial virulence factors. Understanding the QS mechanism in ESKAPE pathogens opens the door to the creation of efficient QS-targeted drugs as substitutes for the drawbacks of traditional therapeutic choices. To prevent future bacterial pathogeneses and the antibiotic resistance caused by these difficult-to-treat pathogens, the complete QS pathways should be blocked or terminated.
Author Contributions
Conceptualization, S.S. and N.I.; software, S.S. and N.S.; validation, N.I. and N.S.; data curation, S.S., N.S. and N.I.; writing—original draft preparation, S.S.; writing—review and editing, S.S. and N.I.; visualization, S.S., N.S. and N.I.; supervision, N.I.; project administration, S.S. and N.I.; funding acquisition, S.S. and N.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Office of the Permanent Secretary, Ministry of Higher Education, Science, Research and Innovation (OPS MHESI), Thailand Science Research and Innovation (TSRI), grant number RGNS 64-205; Mahidol University (Basic Research Fund: fiscal year 2023).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhargava, N.; Sharma, P.; Capalash, N. Quorum sensing in Acinetobacter: An emerging pathogen. Crit. Rev. Microbiol. 2010, 36, 349–360. [Google Scholar] [CrossRef]
- Stacy, D.M.; Welsh, M.A.; Rather, P.N.; Blackwell, H.E. Attenuation of quorum sensing in the pathogen Acinetobacter baumannii using non-native N-Acyl homoserine lactones. ACS Chem. Biol. 2012, 7, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Zarrilli, R. Acinetobacter baumannii virulence determinants involved in biofilm growth and adherence to host epithelial cells. Virulence 2016, 7, 367–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colquhoun, J.M.; Rather, P.N. Insights into mechanisms of biofilm formation in Acinetobacter baumannii and implications for uropathogenesis. Front. Cell. Infect. Microbiol. 2020, 10, 253. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Simões, M. Quorum sensing inhibition by marine bacteria. Mar. Drugs 2019, 17, 427. [Google Scholar] [CrossRef] [Green Version]
- Theuretzbacher, U.; Outterson, K.; Engel, A.; Karlén, A. The global preclinical antibacterial pipeline. Nat. Rev. Microbiol. 2020, 18, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.H.; Wang, L.H.; Xu, J.L.; Zhang, H.B.; Zhang, X.F.; Zhang, L.H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the labyrinth of antibacterial discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaaban, M.; Elgaml, A.; Habib, E.S.E. Biotechnological applications of quorum sensing inhibition as novel therapeutic strategies for multidrug resistant pathogens. Microb. Pathog. 2019, 127, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Diggle, S.P.; Griffin, A.S.; Campbell, G.S.; West, S.A. Cooperation and conflict in quorum-sensing bacterial populations. Nature 2007, 450, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Juárez, I.; Maeda, T.; Mandujano-Tinoco, E.A.; Tomás, M.; Pérez-Eretza, B.; García-Contreras, S.J.; García-Contreras, R. Role of quorum sensing in bacterial infections. World J. Clin. Cases 2015, 3, 575–598. [Google Scholar] [CrossRef] [PubMed]
- Egland, K.A.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Analysis of the LuxR DNA binding region by alanine-scanning mutagenesis. J. Bacteriol. 2001, 183, 382–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clewell, D.B.; Weaver, K.E.; Dunny, G.M.; Coque, T.M.; Francia, M.V.; Hayes, F. Extrachromosomal and mobile elements in enterococci: Transmission, maintenance, and epidemiology. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 309–320. [Google Scholar]
- Dunny, G.M. Enterococcal sex pheromones: Signaling, social behavior, and evolution. Annu. Rev. Genet. 2013, 47, 457–482. [Google Scholar] [CrossRef] [PubMed]
- Varahan, S.; Harms, N.; Gilmore, M.S.; Tomich, J.M.; Hancock, L.E. An ABC transporter is required for secretion of peptide sex pheromones in Enterococcus faecalis. MBio 2014, 5, e01726-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, F.Y.; Clewell, D.B. Identification of the cAD1 sex pheromone precursor in Enterococcus faecalis. J. Bacteriol. 2002, 184, 1880–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, W.; Shepard, B.D.; Gilmore, M.S. Two-component regulator of Enterococcus faecalis cytolysin responds to quorum-sensing autoinduction. Nature 2002, 415, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Hou, B.; Zhang, C. Quorum sensing LuxS/autoinducer-2 inhibits Enterococcus faecalis biofilm formation ability. J. Appl. Oral Sci. 2018, 26, 1–8. [Google Scholar] [CrossRef]
- Oli, A.K.; Javaregowda, P.K.; Jain, A.; Kelmani, C.R. Mechanism Involved in Biofilm Formation of Enterococcus Faecalis. Available online: https://www.intechopen.com/online-first/81571 (accessed on 20 January 2022).
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef] [PubMed]
- Lyon, G.J.; Wright, J.S.; Muir, T.W.; Novick, R.P. Key determinants of receptor activation in the agr autoinducing peptides of Staphylococcus aureus. Biochemistry 2002, 41, 10095–10104. [Google Scholar] [CrossRef] [PubMed]
- Murray, E.J.; Williams, P. Detection of agr-type autoinducing peptides produced by Staphylococcus aureus. In Quorum Sensing; Humana Press: New York, NY, USA, 2018; pp. 89–96. [Google Scholar]
- Kirchdoerfer, R.N.; Garner, A.L.; Flack, C.E.; Mee, J.M.; Horswill, A.R.; Janda, K.D.; Wilson, I.A. Structural basis for ligand recognition and discrimination of a quorum-quenching antibody. J. Biol. Chem. 2011, 286, 17351–17358. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Qiu, S.; Jiang, Q.; Sun, H.; Xue, T.; Cai, G.; Sun, B. AI-2 quorum sensing negatively regulates rbf expression and biofilm formation in Staphylococcus aureus. Int. J. Med. Microbiol. 2017, 307, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C. The QscR quorum-sensing regulon of Pseudomonas aeruginosa: An orphan claims its identity. J. Bacteriol. 2006, 188, 3169–3171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patankar, A.V.; González, J.E. Orphan LuxR regulators of quorum sensing. FEMS Microbiol. Rev. 2009, 33, 739–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, T.; Gomes, A.É.I.; Siqueira, N.M.G.; Assoni, L.; Darrieux, M.; Venter, H.; Ferraz, L.F.C. Sdia, a quorum-sensing regulator, suppresses fimbriae expression, biofilm formation, and quorum-sensing signaling molecules production in Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 597735. [Google Scholar] [CrossRef]
- Balestrino, D.; Haagensen, J.A.; Rich, C.; Forestier, C. Characterization of type 2 quorum sensing in Klebsiella pneumoniae and relationship with biofilm formation. J. Bacteriol. 2005, 187, 2870–2880. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.F.; Purmal, K.; Chin, S.; Chan, X.Y.; Koh, C.L.; Sam, C.K.; Chan, K.G. N-acyl homoserine lactone production by Klebsiella pneumoniae isolated from human tongue surface. Sensors 2012, 12, 3472–3483. [Google Scholar] [CrossRef] [Green Version]
- Herzberg, M.; Kaye, I.K.; Peti, W.; Wood, T.K. YdgG (TqsA) controls biofilm formation in Escherichia coli K-12 through autoinducer 2 transport. J. Bacteriol. 2006, 188, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev. 2001, 15, 1468–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Keersmaecker, S.C.; Sonck, K.; Vanderleyden, J. Let LuxS speak up in AI-2 signaling. Trends Microbiol. 2006, 14, 114–119. [Google Scholar] [CrossRef]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Isolation and characterization of an autoinducer synthase from Acinetobacter baumannii. J. Bacteriol. 2008, 190, 3386–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saipriya, K.; Swathi, C.H.; Ratnakar, K.S.; Sritharan, V. Quorum-sensing system in Acinetobacter baumannii: A potential target for new drug development. J. Appl. Microbiol. 2020, 128, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subhadra, B.; Oh, M.H.; Choi, C.H. Quorum sensing in Acinetobacter: With special emphasis on antibiotic resistance, biofilm formation and quorum quenching. AIMS Microbiol. 2016, 2, 27–41. [Google Scholar] [CrossRef]
- Choi, A.H.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litrán, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-β-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [Green Version]
- Boşgelmez-Tınaz, G.; Ulusoy, S.; Arıdoğan, B.; Eroğlu, F.; Kaya, S. N-butanoyl-L-homoserine lactone (BHL) deficient Pseudomonas aeruginosa isolates from an intensive care unit. Microbiol. Res. 2005, 160, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.; Chiu, S.K.; Hsueh, P.R.; Wang, N.C.; Wang, C.C.; Fang, C.T. Risk factors for healthcare-associated extensively drug-resistant Acinetobacter baumannii infections: A case-control study. PLoS ONE 2014, 9, e85973. [Google Scholar] [CrossRef] [PubMed]
- López-Martín, M.; Dubern, J.F.; Alexander, M.R.; Williams, P. Abam regulates quorum sensing, biofilm formation, and virulence in Acinetobacter baumannii. J. Bacteriol. 2021, 203, e00635-20. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.D.; Sacha, P.T.; Kaczynska, K.; Tryniszewska, E.A. The diversity of aminoglycoside-modifying enzymes among ESBL-positive proteus mirabilis clinical strains. Medtube Sci. 2014, 4, 16–20. [Google Scholar]
- Nemec, A.; Maixnerová, M.; van der Reijden, T.J.; Van den Broek, P.J.; Dijkshoorn, L. Relationship between the AdeABC efflux system gene content, netilmicin susceptibility and multidrug resistance in a genotypically diverse collection of Acinetobacter baumannii strains. J. Antimicrob. Chemother. 2007, 60, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Courvalin, P.; Grillot-Courvalin, C. RND-type efflux pumps in multidrug-resistant clinical isolates of Acinetobacter baumannii: Major role for AdeABC overexpression and AdeRS mutations. Antimicrob. Agents Chemother. 2013, 57, 2989–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modarresi, F.; Azizi, O.; Shakibaie, M.R.; Motamedifar, M.; Mosadegh, E.; Mansouri, S. Iron limitation enhances acyl homoserine lactone (AHL) production and biofilm formation in clinical isolates of Acinetobacter baumannii. Virulence 2015, 6, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii biofilms: Effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, L.A.; McKnight, S.L.; Kuznetsova, M.S.; Pesci, E.C.; Manoil, C. Functions required for extracellular quinolone signaling by Pseudomonas aeruginosa. J. Bacteriol. 2002, 184, 6472–6480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, R.; Vanderleyden, J.; Michiels, J. Quorum sensing and swarming migration in bacteria. FEMS Microbiol. Rev. 2004, 28, 261–289. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, J.T.; Welch, M.; Spring, D.R. Learning the language of bacteria. ACS Chem. Biol. 2007, 2, 715–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Lequette, Y.; Greenberg, E.P. Activity of purified QscR, a Pseudomonas aeruginosa orphan quorum-sensing transcription factor. Mol. Microbiol. 2006, 59, 602–609. [Google Scholar] [CrossRef]
- Maseda, H.; Sawada, I.; Saito, K.; Uchiyama, H.; Nakae, T.; Nomura, N. Enhancement of the mexAB-oprM efflux pump expression by a quorum-sensing autoinducer and its cancellation by a regulator, MexT, of the mexEF-oprN efflux pump operon in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2004, 48, 1320–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Kim, Y.U.; Koh, B.H.; Hwang, S.S.; Kim, S.H.; Lépine, F.; Cho, Y.H.; Lee, G.R. HHQ and PQS, two Pseudomonas aeruginosa quorum-sensing molecules, down-regulate the innate immune responses through the nuclear factor-κB pathway. Immunology 2010, 129, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Lazar, V.; Holban, A.M.; Curutiu, C.; Chifiriuc, M.C. Modulation of quorum sensing and biofilms in less investigated gram-negative ESKAPE pathogens. Front. Microbiol. 2021, 12, 2072. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.Y.; Sulaiman, J.; Chen, J.W.; Yin, W.F.; Chan, K.G. Quorum sensing activity of Enterobacter asburiae isolated from lettuce leaves. Sensors 2013, 13, 14189–14199. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.; Ponraj, P.; Illakkiam, D.; Rajendhran, J.; Gunasekaran, P. Inactivation of the transcriptional regulator-encoding gene sdiA enhances rice root colonization and biofilm formation in Enterobacter cloacae GS1. J. Bacteriol. 2013, 195, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, Y.Y.; How, K.Y.; Yin, W.F.; Chan, K.G. Functional characterization of quorum sensing LuxR-type transcriptional regulator, EasR in Enterobacter asburiae strain L1. PeerJ 2020, 8, e10068. [Google Scholar] [CrossRef]
- Rezzonico, F.; Smits, T.H.; Duffy, B. Detection of AI-2 receptors in genomes of Enterobacteriaceae suggests a role of type-2 quorum sensing in closed ecosystems. Sensors 2012, 12, 6645–6665. [Google Scholar] [CrossRef] [Green Version]
- Tay, S.B.; Yew, W.S. Development of quorum-based anti-virulence therapeutics targeting Gram-negative bacterial pathogens. Int. J. Mol. Sci. 2013, 14, 16570–16599. [Google Scholar] [CrossRef] [Green Version]
- Reading, N.C.; Sperandio, V. Quorum sensing: The many languages of bacteria. FEMS Microbiol. Lett. 2006, 254, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.C.; Huang, C.T. Effects of quorum-sensing deficiency on Pseudomonas aeruginosa biofilm formation and antibiotic resistance. J. Antimicrob. Chemother. 2002, 49, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, P.S.; Rai, V.R. Attenuation of quorum-sensing-dependent virulence factors and biofilm formation by medicinal plants against antibiotic resistant Pseudomonas aeruginosa. J. Tradit. Complement. Med. 2018, 8, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Val, D.L.; Hanzelka, B.L.; Cronan, J.E., Jr.; Greenberg, E.P. Acyl homoserine-lactone quorum-sensing signal generation. Proc. Natl. Acad. Sci. USA 1999, 96, 4360–4365. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Givskov, M. Quorum-sensing inhibitors as anti-pathogenic drugs. Int. J. Med. Microbiol. 2006, 296, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshi, A.; Kim, E.E.; Hwang, K.Y. Structural insights into Staphylococcus aureus enoyl-ACP reductase (FabI), in complex with NADP and triclosan. Proteins Struct. Funct. Genet. 2010, 78, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Hraiech, S.; Hiblot, J.; Lafleur, J.; Lepidi, H.; Papazian, L.; Rolain, J.M.; Raoult, D.; Elias, M.; Silby, M.W.; Bzdrenga, J.; et al. Inhaled lactonase reduces Pseudomonas aeruginosa quorum sensing and mortality in rat pneumonia. PLoS ONE 2014, 9, e107125. [Google Scholar] [CrossRef]
- Guendouze, A.; Plener, L.; Bzdrenga, J.; Jacquet, P.; Rémy, B.; Elias, M.; Lavigne, J.P.; Daudé, D.; Chabrière, E. Effect of quorum quenching lactonase in clinical isolates of Pseudomonas aeruginosa and comparison with quorum sensing inhibitors. Front. Microbiol. 2017, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Liang, M.; Wang, L.; Chen, R.; Li, H.; Liu, X. Aii810, a novel cold-adapted N-acylhomoserine lactonase discovered in a metagenome, can strongly attenuate Pseudomonas aeruginosa virulence factors and biofilm formation. Front. Microbiol. 2017, 8, 1950. [Google Scholar] [CrossRef] [Green Version]
- Sakr, M.M.; Aboshanab, K.M.; Elkhatib, W.F.; Yassien, M.A.; Hassouna, N.A. Overexpressed recombinant quorum quenching lactonase reduces the virulence, motility and biofilm formation of multidrug-resistant Pseudomonas aeruginosa clinical isolates. Appl. Microbiol. Biotechnol. 2018, 102, 10613–10622. [Google Scholar] [CrossRef]
- Schipper, C.; Hornung, C.; Bijtenhoorn, P.; Quitschau, M.; Grond, S.; Streit, W.R. Metagenome-derived clones encoding two novel lactonase family proteins involved in biofilm inhibition in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2009, 75, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhu, J.; Guo, X.; Kong, D.; Zhang, Q.; Zhou, Y.; Ruan, Z. Characterization of AiiK, an AHL lactonase, from Kurthia huakui LAM0618T and its application in quorum quenching on Pseudomonas aeruginosa PAO1. Sci. Rep. 2018, 8, 6013. [Google Scholar] [CrossRef] [Green Version]
- Grover, N.; Plaks, J.G.; Summers, S.R.; Chado, G.R.; Schurr, M.J.; Kaar, J.L. Acylase-containing polyurethane coatings with anti-biofilm activity. Biotechnol. Bioeng. 2016, 113, 2535–2543. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I.; Nam, J.; Hwang, D.S.; Yeon, K.M.; Kim, J. Immobilization and stabilization of acylase on carboxylated polyaniline nanofibers for highly effective antifouling application via quorum quenching. ACS Appl. Mater. Interfaces 2017, 9, 15424–15432. [Google Scholar] [CrossRef] [PubMed]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Caámara, M.; Williams, P.; Quax, W.J. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect. Immun. 2006, 74, 1673–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijtenhoorn, P.; Mayerhofer, H.; Müller-Dieckmann, J.; Utpatel, C.; Schipper, C.; Hornung, C.; Szesny, M.; Grond, S.; Thürmer, A.; Brzuszkiewicz, E.; et al. A novel metagenomic short-chain dehydrogenase/reductase attenuates Pseudomonas aeruginosa biofilm formation and virulence on Caenorhabditis elegans. PLoS ONE 2011, 6, e26278. [Google Scholar] [CrossRef]
- Zhang, X.; Ou-Yang, S.; Wang, J.; Liao, L.; Wu, R.; Wei, J. Construction of antibacterial surface via layer-by-layer method. Curr. Pharm. Des. 2018, 24, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Pustelny, C.; Albers, A.; Büldt-Karentzopoulos, K.; Parschat, K.; Chhabra, S.R.; Cámara, M.; Fetzner, S. Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem. Biol. 2009, 16, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, G.F.; Park, J.; Mee, J.M.; Ulevitch, R.J.; Janda, K.D. The quorum quenching antibody RS2-1G9 protects macrophages from the cytotoxic effects of the Pseudomonas aeruginosa quorum sensing signalling molecule N-3-oxo-dodecanoyl-homoserine lactone. Mol. Immunol. 2008, 45, 2710–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koul, S.; Prakash, J.; Mishra, A.; Kalia, V.C. Potential emergence of multi-quorum sensing inhibitor resistant (MQSIR) bacteria. Indian J. Microbiol. 2016, 56, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Santajit, S.; Seesuay, W.; Mahasongkram, K.; Sookrung, N.; Pumirat, P.; Ampawong, S.; Reamtong, O.; Chongsa-Nguan, M.; Chaicumpa, W.; Indrawattana, N. Human single-chain variable fragments neutralize Pseudomonas aeruginosa quorum sensing molecule, 3O-C12-HSL, and prevent cells from the HSL-mediated apoptosis. Front. Microbiol. 2020, 11, 1172. [Google Scholar] [CrossRef]
- Park, J.; Jagasia, R.; Kaufmann, G.F.; Mathison, J.C.; Ruiz, D.I.; Moss, J.A.; Meijler, M.M.; Ulevitch, R.J.; Janda, K.D. Infection control by antibody disruption of bacterial quorum sensing signaling. Chem. Biol. 2007, 14, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Suresh, M.K.; Biswas, R.; Biswas, L. An update on recent developments in the prevention and treatment of Staphylococcus aureus biofilms. Int. J. Med. Microbiol. 2019, 309, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.X.; Yang, Y.X.; Zhang, Y.Q.; Wang, L.H. A new synthetic ligand that activates QscR and blocks antibiotic-tolerant biofilm formation in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2014, 98, 2565–2572. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geske, G.D.; O’Neill, J.C.; Blackwell, H.E. N-phenylacetanoyl-L-homoserine lactones can strongly antagonize or superagonize quorum sensing in Vibrio fischeri. ACS Chem. Biol. 2007, 2, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nait Chabane, Y.; Mlouka, M.B.; Alexandre, S.; Nicol, M.; Marti, S.; Pestel-Caron, M.; Vila, J.; Jouenne, T.; Dé, E. Virstatin inhibits biofilm formation and motility of Acinetobacter baumannii. BMC Microbiol. 2014, 14, 62. [Google Scholar] [CrossRef] [Green Version]
- Valdez, J.C.; Peral, M.C.; Rachid, M.; Santana, M.; Perdigon, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: The potential use of probiotics in wound treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Cui, T.; Bai, F.; Sun, M.; Lv, X.; Li, X.; Zhang, D.; Du, H. Lactobacillus crustorum ZHG 2-1 as novel quorum-quenching bacteria reducing virulence factors and biofilms formation of Pseudomonas aeruginosa. LWT 2020, 117, 108696. [Google Scholar] [CrossRef]
- Rana, S.; Bhawal, S.; Kumari, A.; Kapila, S.; Kapila, R. pH-dependent inhibition of AHL-mediated quorum sensing by cell-free supernatant of lactic acid bacteria in Pseudomonas aeruginosa PAO1. Microb. Pathog. 2020, 142, 104105. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc. Natl. Acad. Sci. USA 2011, 108, 3360–3365. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Gu, S.; Cui, X.; Shi, Y.; Wen, S.; Chen, H.; Ge, J. Antimicrobial, anti-adhesive and anti-biofilm potential of biosurfactants isolated from Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus aureus CMCC26003. Microb. Pathog. 2019, 127, 12–20. [Google Scholar] [CrossRef]
- Bouyahya, A.; Chamkhi, I.; Balahbib, A.; Rebezov, M.; Shariati, M.A.; Wilairatana, P.; Mubarak, M.S.; Benali, T.; El Omari, N. Mechanisms, anti-quorum-sensing actions, and clinical trials of medicinal plant bioactive compounds against bacteria: A comprehensive review. Molecules 2022, 27, 1484. [Google Scholar] [CrossRef]
- Packiavathy, I.A.S.V.; Agilandeswari, P.; Musthafa, K.S.; Pandian, S.K.; Ravi, A.V. Antibiofilm and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite methyl Eugenol against gram negative bacterial pathogens. Food Res. Int. 2012, 8, 85–92. [Google Scholar] [CrossRef]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 2013, 35, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, P.; Kumar, H.S.V.; Viswanathan, P. Eugenol exhibits anti-virulence properties by competitively binding to quorum sensing receptors. Biofouling 2017, 33, 624–639. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; Letsididi, K.S.; Yu, F.; Pei, Z.; Wang, H.; Letsididi, R. Inhibitive effect of Eugenol and its nanoemulsion on quorum sensing–mediated virulence factors and biofilm formation by Pseudomonas aeruginosa. J. Food Prot. 2019, 82, 379–389. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Baig, M.H. Eugenol inhibits quorum sensing and biofilm of toxigenic MRSA strains isolated from food handlers employed in Saudi Arabia. Biotechnol. Biotechnol. Equip. 2017, 11, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Rodriguez, M.R.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez Tellez, M.A.; Martins, C.M.; Ayala-Zavala, J.F. Carvacrol as potential quorum sensing inhibitor of Pseudomonas aeruginosa and biofilm production on stainless steel surfaces. Food Control 2017, 75, 255–261. [Google Scholar] [CrossRef]
- Tapia-Rodriguez, M.R.; Bernal-Mercado, A.T.; Gutierrez-Pacheco, M.M.; Vazquez-Armenta, F.J.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez-Tellez, M.A.; Nazzaro, F.; Ayala-Zavala, J.F. Virulence of Pseudomonas aeruginosa exposed to Carvacrol: Alterations of the quorum sensing at enzymatic and gene levels. J. Cell Commun. Signal. 2019, 13, 531–537. [Google Scholar] [CrossRef]
- Pejin, B.; Ciric, A.; Glamoclija, J.; Nikolic, M.; Sokovic, M. In vitro anti quorum sensing activity of Phytol. Nat. Prod. Res. 2015, 29, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Devi, K.R.; Kannappan, A.; Pandian, S.K.; Ravi, A.V. Pipervtle and its bioactive metabolite Phytol mitigates quorum sensing mediated virulence factors and biofilm of nosocomial pathogen Serratia marcescens in vitro. J. Ethnopharmacol. 2016, 193, 592–603. [Google Scholar] [CrossRef]
- Amaya, S.; Pereira, J.A.; Borkosky, S.A.; Valdez, J.C.; Bardón, A.; Arena, M.E. Inhibition of quorum sensing in Pseudomonas aeruginosa by Sesquiterpene lactones. Phytomedicine 2012, 19, 1173–1177. [Google Scholar] [CrossRef] [PubMed]
- Rasamiravaka, T.; Vandeputte, O.M.; Pottier, L.; Huet, J.; Rabemanantsoa, C.; Kiendrebeogo, M.; Andriantsimahavandy, A.; Rasamindrakotroka, A.; Stévigny, C.; Duez, P.; et al. Pseudomonas aeruginosa biofilm formation and persistence, along with the production of quorum sensing-dependent virulence factors, are disrupted by a triterpenoid coumarate ester isolated from Dalbergia trichocarpa, a Tropical Legume. PLoS ONE 2015, 10, e0132791. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.; Duarte, A.; Sousa, S.; Domingues, F.C. Study of the major essential oil compounds of Coriandrum sativum against Acinetobacter baumannii and the effect of Linalool on adhesion, biofilms and quorum sensing. Biofouling 2016, 32, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Vega, P.; Xu, Y.; Chen, C.-Y.; Irudayaraj, J. Exploring the anti-quorum sensing activity of ad-limonene nanoemulsion for Escherichia coli O157:H7. J. Biomed. Mater. Res. A 2018, 106, 1979–1986. [Google Scholar] [CrossRef]
- Rémy, B.; Plener, L.; Decloquement, P.; Armstrong, N.; Elias, M.; Daudé, D.; Chabrière, É. Lactonase specificity is key to quorum quenching in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 762. [Google Scholar] [CrossRef]
- Chow, J.Y.; Yang, Y.; Tay, S.B.; Chua, K.L.; Yew, W.S. Disruption of biofilm formation by the human pathogen Acinetobacter baumannii using engineered quorum-quenching lactonases. Antimicrob. Agents Chemother. 2014, 58, 1802–1805. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, F.; Depke, T.; Hoffmann, M.; Empting, M.; Brönstrup, M.; Müller, R.; Blankenfeldt, W. The alkylquinolone repertoire of Pseudomonas aeruginosa is linked to structural flexibility of the FabH-like 2-heptyl-3-hydroxy-4 (1H)-quinolone (PQS) biosynthesis enzyme PqsBC. Chem. Biochem. 2018, 19, 1531–1544. [Google Scholar]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids suppress Pseudomonas aeruginosa virulence through allosteric inhibition of quorum-sensing receptors. J. Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Pejin, B.; Ciric, A.; Glamoclija, J.; Nikolic, M.; Stanimirovic, B.; Sokovic, M. Quercetin potently reduces biofilm formation of the strain Pseudomonas aeruginosa PAO1 in vitro. Curr. Pharm. Biotechnol. 2015, 16, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Gopu, V.; Meena, C.K.; Shetty, P.H. Quercetin influences Quorum Sensing in Food Borne Bacteria: In-Vitro and In-Silico Evidence. PLoS ONE 2015, 10, e0134684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdönmez, D.; Rad, A.Y.; Aksöz, N.; Erdönmez, D.; Rad, A.Y.; Aksöz, N. Anti-Quorum Sensing Potential of Antioxidant Quercetin and Resveratrol. Braz. Arch. Biol. Technol. 2018, 61, e18160756. [Google Scholar] [CrossRef]
- Ouyang, J.; Feng, W.; Lai, X.; Chen, Y.; Zhang, X.; Rong, L.; Sun, F.; Chen, Y. Quercetin Inhibits Pseudomonas aeruginosa Biofilm Formation via the Vfr-Mediated LasIR System. Microb. Pathog. 2020, 149, 104291. [Google Scholar] [CrossRef]
- Sedlmayer, F.; Woischnig, A.K.; Unterreiner, V.; Fuchs, F.; Baeschlin, D.; Khanna, N.; Fussenegger, M. 5-Fluorouracil blocks quorum-sensing of biofilm-embedded methicillin-resistant Staphylococcus aureus in mice. Nucleic Acids Res. 2021, 49, e73. [Google Scholar] [CrossRef] [PubMed]
- Walz, J.M.; Avelar, R.L.; Longtine, K.J.; Carter, K.L.; Mermel, L.A.; Heard, S.O.; 5-FU Catheter Study Group. Anti-infective external coating of central venous catheters: A randomized, noninferiority trial comparing 5-fluorouracil with chlorhexidine/silver sulfadiazine in preventing catheter colonization. Crit. Care Med. 2010, 38, 2095–2102. [Google Scholar]
- Van Delden, C.; Köhler, T.; Brunner-Ferber, F.; François, B.; Carlet, J.; Pechère, J.C. Azithromycin to prevent Pseudomonas aeruginosa ventilator-associated pneumonia by inhibition of quorum sensing: A randomized controlled trial. Intensive Care Med. 2012, 38, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic illustration of quorum sensing systems in bacteria. In Gram-positive bacteria, AIPs engage the receptor and induce the regulator for target gene expression. AIP synthase functions similarly to AI synthase in Gram-negative bacteria. In Gram-negative bacteria, the AHL receptor (LuxR) senses the signaling AHL molecules outside of the cell and forms a contact interaction with them. The AI synthase participates in the synthesis of these signal molecules. When necessary, LuxI (referring to AHL synthase) suppresses the signal. The AHL-receptor complex interacts with the regulator and contributes to the regulation of the target gene. AIP, or auto-inducing peptide; AHL, acyl homoserine lactone.
Figure 1.
Schematic illustration of quorum sensing systems in bacteria. In Gram-positive bacteria, AIPs engage the receptor and induce the regulator for target gene expression. AIP synthase functions similarly to AI synthase in Gram-negative bacteria. In Gram-negative bacteria, the AHL receptor (LuxR) senses the signaling AHL molecules outside of the cell and forms a contact interaction with them. The AI synthase participates in the synthesis of these signal molecules. When necessary, LuxI (referring to AHL synthase) suppresses the signal. The AHL-receptor complex interacts with the regulator and contributes to the regulation of the target gene. AIP, or auto-inducing peptide; AHL, acyl homoserine lactone.

Figure 2.
Diagram of the major quorum sensing pathways of Enterococcus spp. Conjugative transfer in Enterococcus spp. Pheromone signaling occurs between the two cell types and is the mechanism by which the plasmid pCF10 is transferred from donor cells to recipient cells. The chromosomally encoded peptide cCF10, which is secreted by recipient cells and internalized by donor cells, triggers the expression of genes important for the conjugative process. Asc10, also known as “aggregation substance,” is specifically expressed by particular cells and allows a stable interaction between the donor and receivers. The pCF10 plasmid is then transferred from donor cells to recipient cells. The QS system in Fsr and its regulation in Enterococcus spp. FsrB exports and processes the FsrD propeptide (encoded by fsrD) to create the tiny lactone gelatinase biosynthesis-activating pheromone (GBAP). The intracellular response regulator, FsrA, is phosphorylated in response to extracellular GBAP by FsrC, a component of a two-component regulatory system. FsrA then stimulates the expression of the genes ef1097, ef1097b, the fsr locus, gelE (encoding a gelatinase), and sprE (encoding a serine protease), which lead to biofilm formation.
Figure 2.
Diagram of the major quorum sensing pathways of Enterococcus spp. Conjugative transfer in Enterococcus spp. Pheromone signaling occurs between the two cell types and is the mechanism by which the plasmid pCF10 is transferred from donor cells to recipient cells. The chromosomally encoded peptide cCF10, which is secreted by recipient cells and internalized by donor cells, triggers the expression of genes important for the conjugative process. Asc10, also known as “aggregation substance,” is specifically expressed by particular cells and allows a stable interaction between the donor and receivers. The pCF10 plasmid is then transferred from donor cells to recipient cells. The QS system in Fsr and its regulation in Enterococcus spp. FsrB exports and processes the FsrD propeptide (encoded by fsrD) to create the tiny lactone gelatinase biosynthesis-activating pheromone (GBAP). The intracellular response regulator, FsrA, is phosphorylated in response to extracellular GBAP by FsrC, a component of a two-component regulatory system. FsrA then stimulates the expression of the genes ef1097, ef1097b, the fsr locus, gelE (encoding a gelatinase), and sprE (encoding a serine protease), which lead to biofilm formation.

Figure 3.
Hierarchical quorum sensing network in Staphylococcus aureus. Autoinducing peptide (AIP) synthesis and mechanism of action in S. aureus. A two-component signal transduction pathway underlies AIP-mediated quorum sensing in S. aureus (AgrC and AgrA). RNAII and RNAIII regions’ P2 and P3 promoters are correspondingly activated by phosphorylated AgrA. AIP is synthesized as a result of RNAII expression, and RNAIII expression controls the synthesis of exoproteins, virulence genes, and δ-hemolysin genes while inhibiting the synthesis of adhesins.
Figure 3.
Hierarchical quorum sensing network in Staphylococcus aureus. Autoinducing peptide (AIP) synthesis and mechanism of action in S. aureus. A two-component signal transduction pathway underlies AIP-mediated quorum sensing in S. aureus (AgrC and AgrA). RNAII and RNAIII regions’ P2 and P3 promoters are correspondingly activated by phosphorylated AgrA. AIP is synthesized as a result of RNAII expression, and RNAIII expression controls the synthesis of exoproteins, virulence genes, and δ-hemolysin genes while inhibiting the synthesis of adhesins.

Figure 4.
Quorum sensing cascade in Klebsiella pneumoniae. AI-2 dependent regulated QS signaling in K. pneumoniae. AI-2 is expressed by a luxS homolog, as well as N-octanoyl homoserine lactone (C8-HSL) and N-3-dodecanoyl-L-homoserine lactone (N-3-DL-HSL) (C12-HSL). The luxS system is associated with the expression of LPS synthesis-related genes and their expression of capsule, fimbria, pili, LPS, iron acquisition, and PNAG biosynthesis, as well as biofilm formation. SAM, S-adenosyl methionine; DPD, 4,5-dihydroxy-2,3-pentanedione.
Figure 4.
Quorum sensing cascade in Klebsiella pneumoniae. AI-2 dependent regulated QS signaling in K. pneumoniae. AI-2 is expressed by a luxS homolog, as well as N-octanoyl homoserine lactone (C8-HSL) and N-3-dodecanoyl-L-homoserine lactone (N-3-DL-HSL) (C12-HSL). The luxS system is associated with the expression of LPS synthesis-related genes and their expression of capsule, fimbria, pili, LPS, iron acquisition, and PNAG biosynthesis, as well as biofilm formation. SAM, S-adenosyl methionine; DPD, 4,5-dihydroxy-2,3-pentanedione.

Figure 5.
Diagrammatic illustration of quorum sensing signaling in Acinetobacter baumannii. AI-1 mediates the QS cascade in A. baumannii through the LuxR receptor (AbaR) and LuxI synthase (AbaI) systems. This QS system is crucial for capsule biogenesis, bacterial motility, multidrug efflux pumps, and biofilm development capability. AcylACP, Acyl carrier protein.
Figure 5.
Diagrammatic illustration of quorum sensing signaling in Acinetobacter baumannii. AI-1 mediates the QS cascade in A. baumannii through the LuxR receptor (AbaR) and LuxI synthase (AbaI) systems. This QS system is crucial for capsule biogenesis, bacterial motility, multidrug efflux pumps, and biofilm development capability. AcylACP, Acyl carrier protein.

Figure 6.
Quorum sensing machinery in Pseudomonas aeruginosa. The hierarchical organization of the three major QS systems in P. aeruginosa has been recognized as Las, Rhl, and PQS (such as Pseudomonas Quinolone Signal). These systems involve the signal synthases LasI, RhlI, PqsABCDEH, and the receptors LasR, RhlR, and PqsR, respectively. Three autoinducer signaling molecules are employed by these systems. The Las and Rhl systems employ two acyl-homoserine lactones (AHL), 3-oxo-C12 HSL, and C4 HSL, whereas PQS is based on 2-alkyl-4-quinolones (i.e., PQS and HHQ). Besides, the cognate receptors for 3-oxo-C12-HSL are also QscR. All three of these systems are interconnected, and Las is regarded as the global activator. The Las system controls exoprotease, elastase, lectin, pyocyanin synthesis, and biofilm development. The Rhl system triggers rhamnolipid biogenesis. The Pqs system positively regulates the Las and Rhl systems and triggers pyoverdine and pyochelin production. Solid arrows indicate direct control of genotypic or phenotypic signal regulation, while dashed arrows indicate additional or alternative transcriptional control for gene expression.
Figure 6.
Quorum sensing machinery in Pseudomonas aeruginosa. The hierarchical organization of the three major QS systems in P. aeruginosa has been recognized as Las, Rhl, and PQS (such as Pseudomonas Quinolone Signal). These systems involve the signal synthases LasI, RhlI, PqsABCDEH, and the receptors LasR, RhlR, and PqsR, respectively. Three autoinducer signaling molecules are employed by these systems. The Las and Rhl systems employ two acyl-homoserine lactones (AHL), 3-oxo-C12 HSL, and C4 HSL, whereas PQS is based on 2-alkyl-4-quinolones (i.e., PQS and HHQ). Besides, the cognate receptors for 3-oxo-C12-HSL are also QscR. All three of these systems are interconnected, and Las is regarded as the global activator. The Las system controls exoprotease, elastase, lectin, pyocyanin synthesis, and biofilm development. The Rhl system triggers rhamnolipid biogenesis. The Pqs system positively regulates the Las and Rhl systems and triggers pyoverdine and pyochelin production. Solid arrows indicate direct control of genotypic or phenotypic signal regulation, while dashed arrows indicate additional or alternative transcriptional control for gene expression.

Figure 7.
Quorum sensing signaling in Enterobacter spp. Three main QS systems play a role in intercellular communication signaling within Enterobacter spp. mediated through AI-1, AI-2, and AI-3 cascades to induce virulence expression and biofilm formation.
Figure 7.
Quorum sensing signaling in Enterobacter spp. Three main QS systems play a role in intercellular communication signaling within Enterobacter spp. mediated through AI-1, AI-2, and AI-3 cascades to induce virulence expression and biofilm formation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Anti-QS strategies capable of bacterial virulence and antimicrobial resistance.
| Strategies | Anti-QS Agents | Modes of Action | Effect on ESKAPE Organisms | References |
|---|---|---|---|---|
| Inhibition of AI synthases | Sinefungin | Structural analogues of S-adenosyl methionine (SAM) and acyl-carrier protein (ACP), the substrates of AHL synthases | Prevent bacterial infection and diminish QS-mediated virulence factors by blocking P. aeruginosa AHL synthesis | [64,65] |
| Butyryl-SAM | ||||
| L/D-S-adenosylhomocysteine | ||||
| Triclosan | Reduction of the establishment of enoyl-ACP reductase precursors (FabI) | Decrease S. aureus AHL production | [66] | |
| Targeting of QS Ligands | AHL lactonases (such as SsoPox, lactonase Aii810, QQ lactonase enzyme AHL-1, a novel lactonase cloned by bpiB01 and bpiB04 and lactonase AiiK) | Hydrolysis of the AHL lactone ring to form the consequent N-acyl homoserine | Lessen the extracellular proteases and pyocyanin biosynthesis, rhamnolipids, swarming motility and biofilm production and prevent bacterial infection of P. aeruginosa | [67,68,69,70,71,72] |
| Acylases (i.e., N-acyl homoserine lactone acylase PA2385, acylase (EC.3.5.1.14) | Degradation of the AHL amide bond and generation of the corresponding free fatty acid and a lactone ring | Decrease elastase, pyocyanin synthesis and biofilm biomass and formation in P. aeruginosa | [73,74,75] | |
| Oxidoreductases (e.g., BpiB09) | Oxidation and consequent inhibition of signal QS molecules | Slow down bacterial motility and reduce biofilm formation and pyocyanin production in P. aeruginosa; hinder bacterial biofilm development and decrease the growth rate of K. pneumoniae | [76,77] | |
| 3-Hydroxy-2-methyl-4(1H)-quinolone 2, 4-dioxygenase | Catalysis of the conversion of PQS to N-octanoylanthranilic acid and carbon monoxide | Hamper lectin A, pyocyanin and rhamnolipid biosynthesis of P. aeruginosa | [78] | |
| Quorum quenching antibody, RS2-1G9 | Hydrolysis of 3-oxo-C12-HSL | Inhibit the activation of the mitogen- activated protein kinase p38 and protect against the cytotoxic effects of P. aeruginosa on macrophages generated from murine bone marrow | [79] | |
| XYD-11G2 antibody | Hydrolysis of 3-oxo-C12-HSL | Conquest the bacterial QS signals of P. aeruginosa | [80] | |
| Human single-chain variable fragments | Hydrolysis of 3-oxo-C12-HSL | Prevent mammalian cell apoptosis triggered by P. aeruginosa | [81] | |
| AP4-24H11 antibody | Targeting of autoinducing peptide-4 | Induce the protective properties of S. aureus-produced abscess in vivo | [82] | |
| Synthetic RIP | Targeting of autoinducing peptide-4 | Diminish S. aureus infections in vivo | [83] | |
| Blockade of QS Transcriptional Regulators | Flavonoids | Allosteric blockade of the AI-binding receptors LasR and RhlR | Modulate the transcription of QS-controlled target promoters and limit the synthesis of virulence factors in P. aeruginosa | [84] |
| N-decanoyl-L-homoserine benzyl ester | Repression of the quorum sensing control repressor | Impair the production of biofilms, swarming activity, and protease and elastase enzymes in P. aeruginosa | [85] | |
| Meta-bromo-thiolactone | Allosteric blockade of the AI-binding receptors LasR and RhlR | Decrease pyocyanin synthesis and biofilm development in P. aeruginosa | [86] | |
| A4, 4-bromophenyl-PHL B7, 4-iodo PHL C10 and 3-nitro PHL C14 | Blockade of the AI-binding receptors, including TraR, LasR, and LuxR | Suppress the development of virulence factors in P. aeruginosa | [87] | |
| Virstatin | Inhibition of the expression of the anoR gene | Prevent bacterial movement and biofilm formation in A. baumannii | [88] | |
| Probiotics | L. plantarum PA 100 | Blockade of the function and inhibition of the synthesis of acyl homoserine lactones | Diminish biofilm production and elastase activity in P. aeruginosa | [89] |
| C. crustorum ZHG 2-1 | Degradation of C4-HSL and 3-oxo-C12-HSL | Suppress virulence factors (chitinase and protease), reduce swarming and swimming motilities, and inhibit biofilm formation in P. aeruginosa | [90] | |
| Cell-free acidic supernatants L. lactis NCDC 309, L. rhamnosus MTCC 5897, L. rhamnosus MTCC 5857, L. fermentum MTCC 5898, L. acidophilus NCDC 15, L. delbrueckii subsp. lactis, L. plantarum NCDC 372 | Destruction of C4-HSL and 3-oxo-C12-HSL | Inhibit biofilm formation, elastase, and expression of lasI and rhlI in P. aeruginosa | [91] | |
| L. reuteri RC-14 | Inhibition of arg gene expression by bioactive cyclic dipeptides (known as 2,5-diketo-piperazines, or DKPs) | Neutralise S. aureus MN8 toxin TSST-1 synthesis (toxic shock syndrome) | [92] | |
| L. plantarum, P. acidilactici | Downregulation of genes including cidA, icaA, dltB, agrA, sortaseA, and sarA | Suppress the formation of S. aureus biofilm | [93] | |
| Plant extracts | Eugenol | Suppression the expression of las and pqs systems | Prevent biofilm formation of P. aeruginosa PAO1. | [94,95,96,97] |
| Reduction in the level of QS synthase genes, including lasI, rhlI, and rhlA, | Inhibit biofilm growth and regressed virulence production (including pyocyanin, pyocyanin, and elastase) of P. aeruginosa PAO1 | [98] | ||
| Unknown | Limit the production of protease and pigments in MRSA | [99] | ||
| Carvacrol (2-methyl-5-(1-methylethyl)-phenol) | Blocking lasI and lasR expression | Lower the biofilm development and bacterial motility of P. aeruginosa | [100,101] | |
| Phytol | Unknown | Suppress flagella mobilization, restricts the formation of pyocyanin, and inhibits the establishment of the biofilm in P. aeruginosa PAO1 | [102,103] | |
| Sesquiterpene lactones | Unknown | Diminish the QS mediators in P. aeruginosa ATCC 27853 | [104] | |
| Oleanolic aldehyde coumarate | Downregulation of lasI/lasR, rhlI/rhlR, and gacA expression | Decrease the P. aeruginosa ‘s biofilm biogenesis | [105] | |
| Linalool | Unknown | Prevent the establishment of A. baumannii‘s biofilms and alter this strain’s surface adhesion. | [106,107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Santajit, S.; Sookrung, N.; Indrawattana, N. Quorum Sensing in ESKAPE Bugs: A Target for Combating Antimicrobial Resistance and Bacterial Virulence. Biology 2022, 11, 1466. https://doi.org/10.3390/biology11101466
AMA Style
Santajit S, Sookrung N, Indrawattana N. Quorum Sensing in ESKAPE Bugs: A Target for Combating Antimicrobial Resistance and Bacterial Virulence. Biology. 2022; 11(10):1466. https://doi.org/10.3390/biology11101466
Chicago/Turabian StyleSantajit, Sirijan, Nitat Sookrung, and Nitaya Indrawattana. 2022. "Quorum Sensing in ESKAPE Bugs: A Target for Combating Antimicrobial Resistance and Bacterial Virulence" Biology 11, no. 10: 1466. https://doi.org/10.3390/biology11101466
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.