Micropropagation of Seed-Derived Clonal Lines of the Endangered Agave marmorata Roezl and Their Compatibility with Endophytes

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Material
2.2. Surface Disinfection of Seeds
2.3. Seed Germination Experiments
2.4. Shoot Multiplication
2.5. Plant Adaptation to Beach Sand-Vermiculite Microcosm Nitrogen-Free
2.6. Phytochemical Compounds Analysis of Selected A. marmorata Clonal Lines
2.6.1. Plant Material and Its Lyophilization
2.6.2. Sample Preparation to Obtain Extracts
2.6.3. Total Flavonoid, Phenolic and Steroidal Saponin Quantification
2.6.4. HPLC-PDA Analysis/Fraction Collection UHPLC-UV
2.6.5. MALDI-TOF Mass Spectrometry Analysis
2.7. Plant Growth Stimulation with Endophytic Bacteria
2.8. Statical Analysis
3. Results
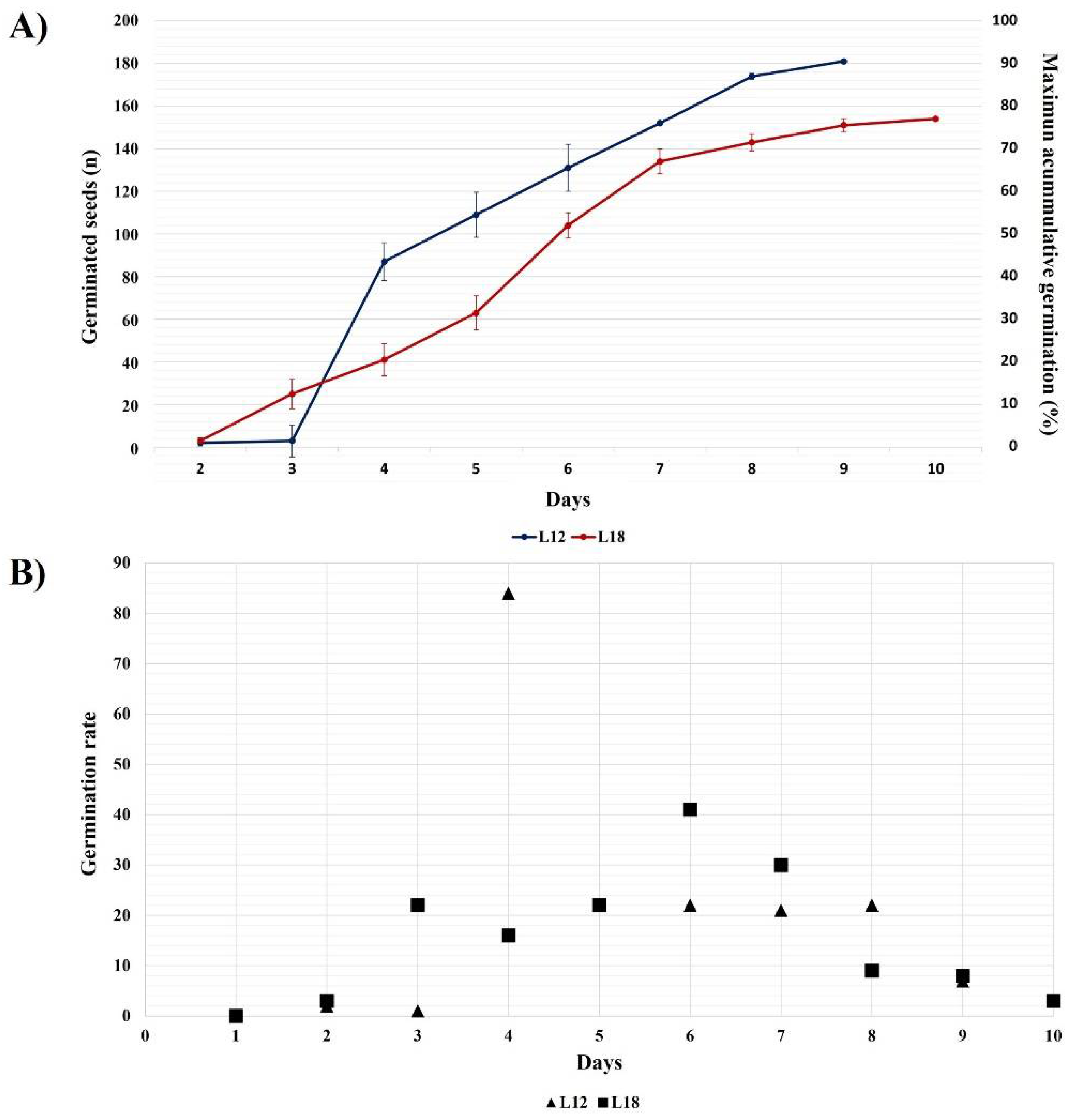
3.1. Seed Germination Rate and Final Germination Percentage
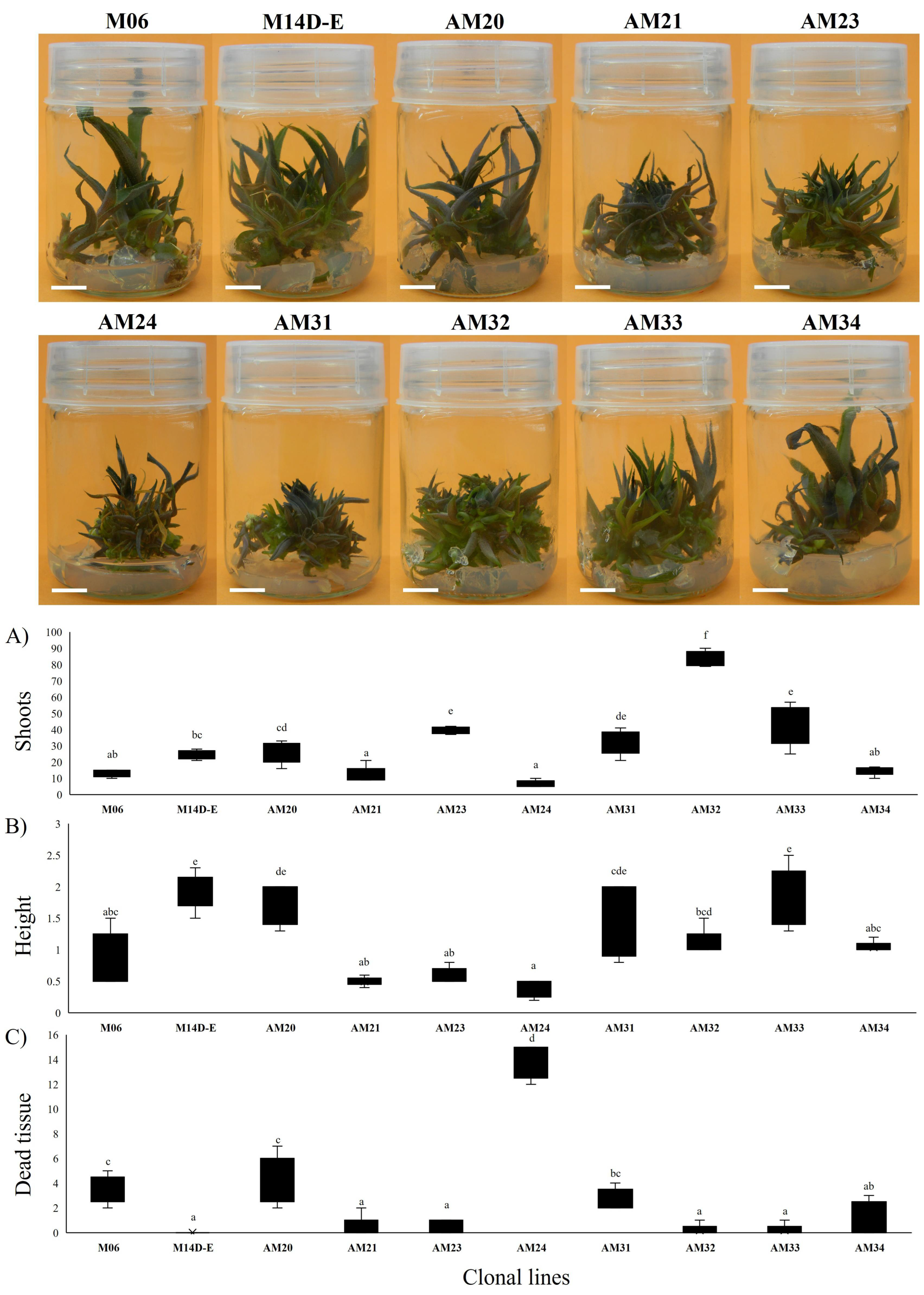
3.2. Seedling Selection for Clonal Propagation
3.3. Shoot Multiplication Induction with BAP in MS without Vitamins
3.4. Comparative Analysis of Total Flavonoid, Phenolic and Saponin Content in the Clonal Lines
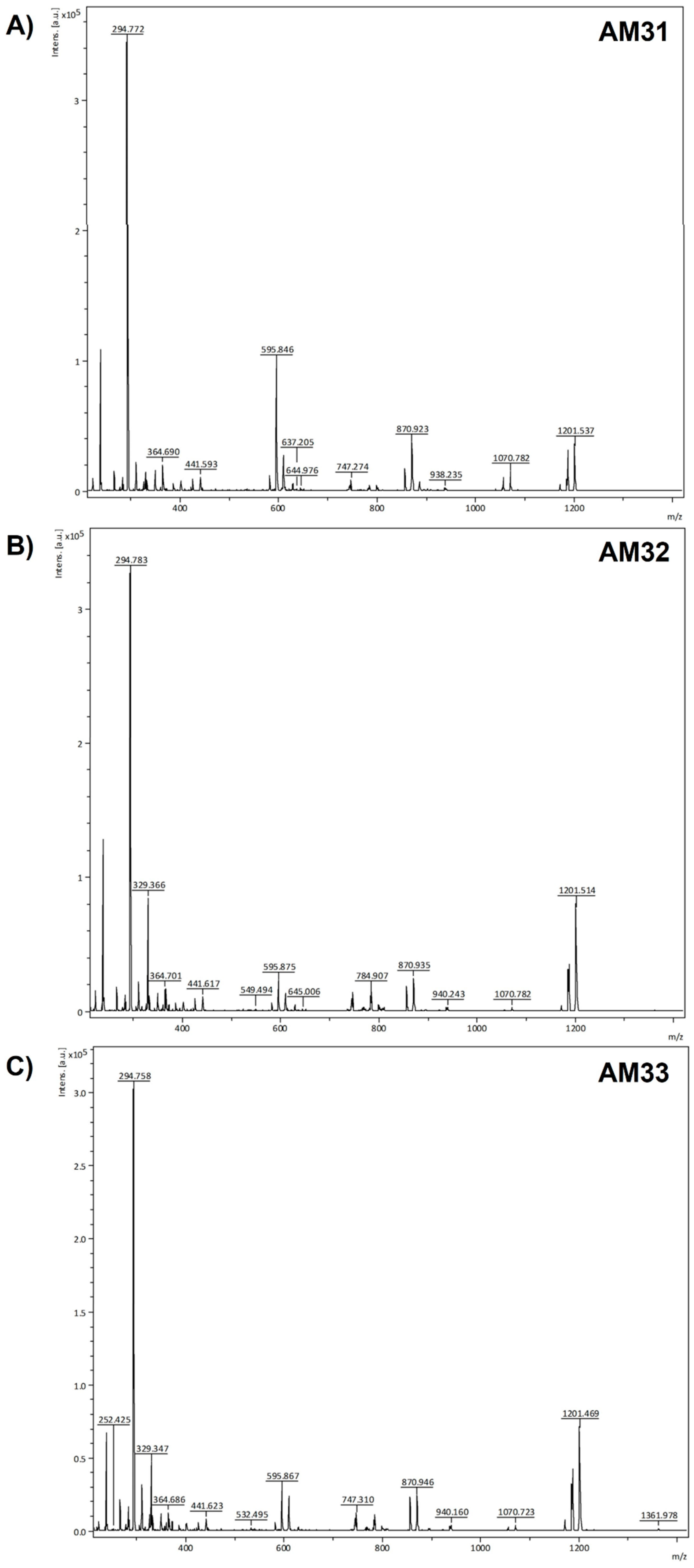
3.5. MALDI-TOF Profiling of the HPLC Collected Fractions from Clonally Propagated Lines of A. marmorata
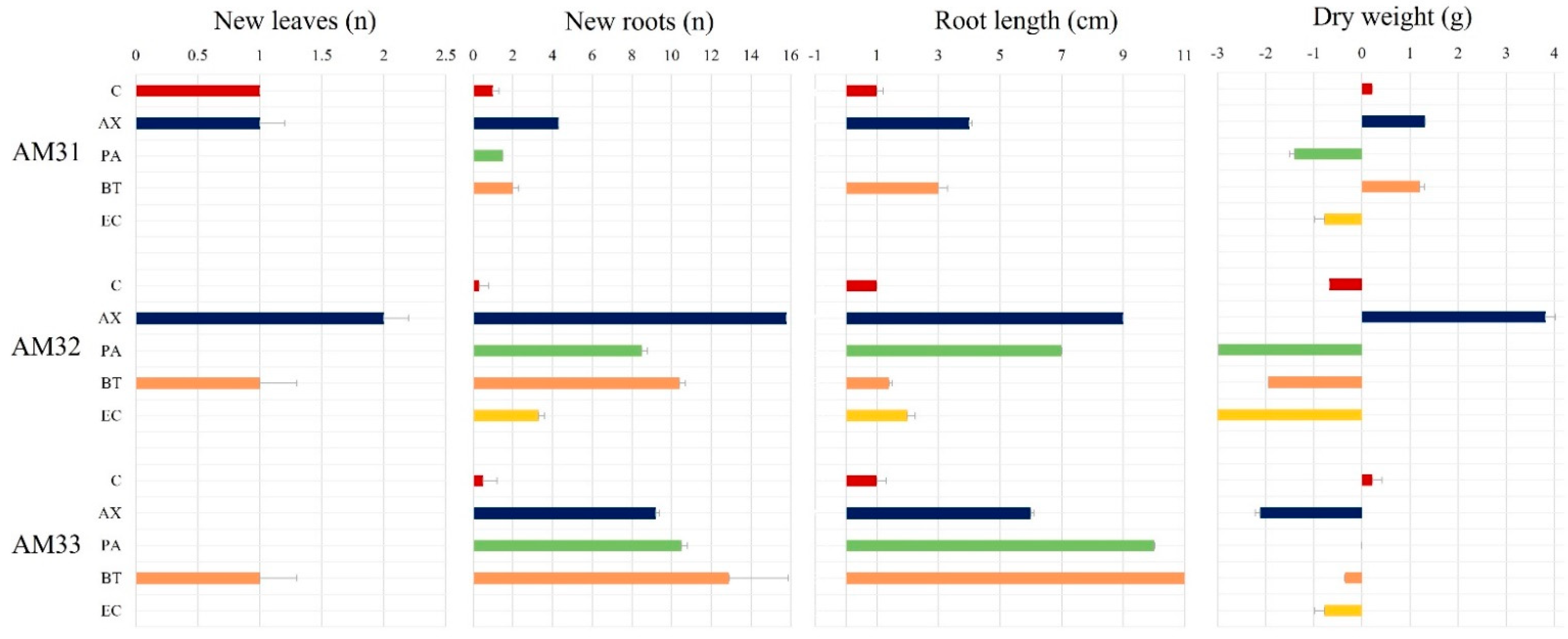
3.6. Plant Growth Parameters and Compatibility of Three Clonal Lines with Endophytic Bacteria
4. Discussion
4.1. Seedling Selection Derived from A. marmorata Seed Germination Capabilities
4.2. Seedling Response to MS+BAP for Clonal Selection
4.3. Shoot Propagation Induced by BAP in MS without Vitamins
4.4. Total Flavonoid, Phenolic and Saponin Content in the 3 Clonal Lines
4.5. Mass Spectrometry Identification of Phytochemicals Compounds Accumulated in the Clonal Lines
4.6. Compatibility of Three Clonal Lines with Endophytic Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentry, H.S. Agaves of Continental North America; University of Arizona Press: Tucson, AZ, USA, 1982; pp. 1–670. ISBN 9780816523955. [Google Scholar]
- Radding, C. The Children of Mayahuel: Agaves, human cultures, and desert landscapes in northern Mexico. Environ. Hist. Durh 2011, 17, 84–115. Available online: https://www.jstor.org/stable/23212617 (accessed on 3 March 2022). [CrossRef]
- Bermúdez-Bazán, M.; Castillo-Herrera, G.A.; Urias-Silvas, J.E.; Escobedo-Reyes, A.; Estarrón-Espinosa, M. Hunting bioactive molecules from the Agave genus: An update on extraction and biological potential. Molecules 2021, 26, 6789. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Delgado, N.C.; Torres-Maravilla, E.; Mayorga-Reyes, L.; Martín, R.; Langella, P.; Pérez-Pastén-Borja, R.; Sánchez-Pardo, M.E.; Bermúdez-Humarán, L.G. Antioxidant and anti-inflammatory properties of probiotic candidate strains isolated during fermentation of Agave (Agave angustifolia Haw). Microorganisms 2021, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, A.V.; Simpson, J.; Clench, M.R.; Gomez-Vargas, A.D.; Ordaz-Ortiz, J.J. Localization and composition of fructans in stem and rhizome of Agave tequilana Weber var. Azul. Front. Plant Sci. 2021, 11, 608850. [Google Scholar] [CrossRef]
- Delfín-Ruíz, M.E.; Calderón-Santoyo, M.; Ragazzo-Sánchez, J.A.; Gómez-Rodriguez, K.; Aguilar-Uscanga, M.G. Ethanol production from enzymatic hydrolysates optimized of Agave tequilana Weber var. Azul and Agave karwinskii bagasses. Bioenerg. Res. 2021, 14, 785–798. [Google Scholar] [CrossRef]
- Holtum, J.A.M.; Chambers, D.; Morgan, T.; Tan, D.K.Y. Agaves as biofuel stock in Australia. GCB. Bioenergy 2011, 3, 58–67. [Google Scholar] [CrossRef]
- Honorato-Salazar, J.A.; Aburto, J.; Amezcua-Allieri, M.A. Agave and Opuntia species as sustainable feedstocks for bioenergy and byproducts. Sustainability 2021, 13, 12263. [Google Scholar] [CrossRef]
- Yan, X.; Corbin, K.R.; Burton, R.; Tan, D.K.Y. Agave: A promising feedstock for biofuels in the water-energy-food environment (WEFE) nexus. J. Clean. Prod. 2020, 261, 121283. [Google Scholar] [CrossRef]
- Consejo Regulador del Tequila. Informe Estadistico; CRT: Zapopan, Mexico, 2021. [Google Scholar]
- Consejo Regulador Mezcal. Informe Estadístico 2020; Consejo Regulador Mezcal: Oaxaca, Mexico, 2020. [Google Scholar]
- Torres-Garcia, I.; Rendon-Sandoval, F.J.; Blancas, J.; Casas, A.; Moreno-Calles, A.I. The genus Agave in agroforestry systems of Mexico. Bot. Sci. 2019, 97, 263–290. [Google Scholar] [CrossRef]
- Sánchez-Teyer, F.; Moreno-Salazr, S.; Esqueda, M.; Barraza, A.; Robert, M.L. Genetic variability of wild Agave angustifolia populations based on AFLP: A basic study for conservation. J. Arid. Envriron. 2009, 73, 611–616. [Google Scholar] [CrossRef]
- Dominguez-Licona, J.M. Diagnóstico de los Recursos Naturales de la Bahía y Micro-cuenca de Cacaluta, Municipio de Santa María Huatulco, Oaxaca; Domínguez-Licona, J.M., Ed.; Universidad del Mar Huatulco: Santa María Huatulco, Mexico, 2008; 453p, ISBN 978-970-9932-03-4. [Google Scholar]
- Flores-Maya, S.; Vargas-Jurado, M.A.; Suarez-Mota, M.E.; Barrera-Ecorcia, H. Karyotype analysis of Agave marmorata and Agave peacockii (Agavaceae) from alluvial terraces of the Zapotitlán River, Puebla, Mexico. Polibotanica 2015, 40, 109–125. [Google Scholar] [CrossRef]
- García-Mendoza, A.J.; Franco-Martínez, I.S.; Sandoval-Gutiérrez, D. Cuatro especies nuevas de Agave (Asparagaceae, Agavoideae) del sur de México. Acta Bot. Mex. 2019, 126, e1461. [Google Scholar] [CrossRef]
- Hanus-Fajerska, E.; Kocot, D.; Wisznieska, A.; Koźmińska, A.; Muszyńska, E. Micropropagation and experimental field cultivation of Pulsatilla turczaninovii Kryl. Et Serg. (Ranunculaceae). Plant Cell Tissue Organ Cult. 2021, 147, 477–489. [Google Scholar] [CrossRef]
- Sumira, A.; Shabeer, A.D.; Ishfq, M.L.; Aabid, H.M.; Azra, N.K. Robust in-vitro protocol for shoot multiplication of Echinacea angustifolia. Curr. Plant Biol. 2021, 28, 100221. [Google Scholar] [CrossRef]
- Lee, J.-H.; Han, T.-H. Micropropagation of the plantlets derived from seeds in the genus Acorus (A. calamus and A. grmineus). Hort. Environ. Biotechnol. 2011, 52, 89–94. [Google Scholar] [CrossRef]
- Milena, T.; Dragana, A.; Aleksandar, C.; Nabil, G.; Angelina, S.; Sladana, J. Advancement in protocol for in vitro seed germination, plant regeneration and cryopreservation of Viola cornuta. 3 Biotech 2019, 9, 17. [Google Scholar] [CrossRef]
- Guarino, F.; Conte, B.; Improta, G.; Sciarrillo, R.; Castiglione, S.; Cicatelli, A.; Guarino, C. Genetic characterization, micropropagation, and potential us for arsenic phytoremediation of Dittrichia viscosa (L.) Greuter. Ecotoxicol. Environ. Saf. 2018, 148, 675–683. [Google Scholar] [CrossRef]
- Barrientos, F.P.; Villegas, M.A.; Vazquez, R.A. Métodos de propagación de agaves. In Biología y Aprovechamiento Integral del henequen y Otros Agaves; Cruz, C., Castillo, L., Robert, M., Ondarza, R.N., Eds.; Centro de Investigación Científica de Yucatán, A.C: Mérida, Mexico, 1985; pp. 91–96. [Google Scholar]
- Ramírez-Tobías, H.M.; Peña-Valdivia, C.B.; Aguirre, J.R.; Reyes-Agüero, J.A.; Sánchez-Urdaneta, A.B.; Valle-Guadarrama, S. Seed germination temperatures of eight Mexican Agave species with an extensive history of human use. Plant Species Biol. 2012, 27, 124–137. [Google Scholar] [CrossRef]
- Jiménez-Valdés, M.; Godínez-Alvarez, H.; Caballero, J.; Lira, R. Population dynamics od Agave marmorata Roezl. under two contrasting management systems in central Mexico. Econ. Bot. 2010, 64, 149–160. [Google Scholar] [CrossRef]
- Peña-Valdivia, C.B.; Sánchez-Urdaneta, A.B.; Aguirre, J.R.; Trejo, C.; Cárdenas, E.; Villegas, A. Temperature and mechanical scarification on seed germination of ‘maguey’ (Agave salmiana Otto ex Salm-Dyck). Seed Sci. Technol. 2006, 34, 47–56. [Google Scholar] [CrossRef]
- Ramírez-Tobías, H.M.; Peña-Valdivia, C.B.; Trejo, C.; Aguirre, R.J.R.; Vaquera, H.H. Seed germination of Agave species as influenced by substrate water potential. Biol. Res. 2014, 47, 11. [Google Scholar] [CrossRef] [PubMed]
- Slazak, B.; Sliwinska, E.; Saluga, M.; Ronikier, M.; Bujak, J.; Slomka, A.; Goransson, U.; Kuta, E. Micropropagation of Viola uliginosa (Violaceae) for endangered species conservation and for somaclonal variation-enhanced cyclotide biosynthesis. Plant Cell Tissue Organ Cult. 2014, 120, 179–190. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Moraes, R.M.; Cerdeira, A.L.; Lourenço, M.V. Using micropropagation to develop medicinal plants into crops. Molecules 2021, 26, 1752. [Google Scholar] [CrossRef]
- Rodríguez-Garay, B.; Rodríguez-Domínguez, J.M. Micropropagation of Agave species. In Plant Cell Culture Protocols; Loyola-Vargas, V., Ochoa-Alejo, N., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; p. 1815. [Google Scholar] [CrossRef]
- Bautista-Montes, E.; Hernández-Soriano, L.; Simpson, J. Advances in the micropropagation and genetic transformation of Agave species. Plants 2022, 11, 1757. [Google Scholar] [CrossRef]
- Domínguez-Rosales, M.S.; Alpuche-Solis, A.G.; Vasco-Mendez, N.L.; Perez-Molphe, B.E. Effect of cytokinins on the in vitro propagation of Mexican Agaves. Rev. Fitoc. Mex. 2008, 33, 317–322. [Google Scholar] [CrossRef]
- Ramirez-Malagon, R.; Boronadenko, A.; Pérez-Moreno, L.; Salas-Araiza, M.; Nuñez-Palenius, H.G.; Ochoa-Alejo, N. In vitro propagation of three Agave species used for liquor distillation and three for landscape. Plant Cell Tissue Organ Cult. 2008, 94, 201–207. [Google Scholar] [CrossRef]
- Rios-Ramírez, S.C.; Enriquez-del Valle, J.R.; Rodríguez-Ortiz, G.; Ruiz-Luna, J. Benzylaminopurine and indol-3 acetic acid concentrations in In vitro proliferation of Agave angustifolia adventitious shoots. Cienc. Investig. Agrar. 2017, 44, 285–294. [Google Scholar] [CrossRef]
- Santacruz-Ruvalcaba, F.; Gutierrez-Pulido, H.; Rodriguez-Garay, B. Efficient in vitro propagation of Agave parrasana Berger. Plant Cell Tissue Organ Cult. 1999, 56, 163–167. [Google Scholar] [CrossRef]
- Delgado-Aceves, L.; Portillo, L.; Folgado, R.; Romo-Paz, F.J.; González-Arnao, M.T. New approaches fro micropropagation and cryopreservation of Agave preacokii, an endangered species. Plant Cell Tissue Organ Cult. 2022, 150, 85–95. [Google Scholar] [CrossRef]
- Beltran-Garcia, M.J.; White, J.F.; Padro, F.M.; Prieto, K.R.; Yamaguchi, L.F.; Torres, M.S.; Kato, M.J.; Medeiros, M.H.G.; Di Mascio, P. Nitrogen acquisition in Agave tequilana from degradation of endophytic bacteria. Sci. Rep. 2014, 4, 6938. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.; Hutter, I.; Döring, M. Commercial use of endophytes in micropropagaction. Acta Hortic. 2017, 1155. [Google Scholar] [CrossRef]
- Murashigue, T.; Skoog, F. A revised medium for rapid growth and bioassays with Tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Portillo, L.; Santacruz-Ruvalcaba, F.; Gutiérrez-Mora, A.; Rodríguez-Garay, B. Somatic embryogenesis in Agave tequilana Weber cultivar azul. In Vitro Cell. Dev. Biol. Plant 2007, 43, 569–575. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetrics methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Nagaraju, Y.; Mahadevaswamy; Naik, N.M.; Gowdar, S.B.; Narayanarao, K.; Satyanarayanarao, K. ACC Deaminase-positive halophilic bacterial isolates with multiple plant growth-promoting traits improve the growth and yield of chickpea (Cicer arietinum L.) under salinity stress. Front. Agron. 2021, 3, 681007. [Google Scholar] [CrossRef]
- ALKahtani, M.D.F.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N.; et al. Isolation and characterization of plant growth promoting endophytic bacteria from desert plants and their application as bioinoculants for sustainable agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Silva, U.C.; Cuadros-Orellana, S.; Silva, D.R.C.; Freitas-Júnior, L.F.; Fernandes, A.C.; Leite, L.R.; Oliverira, C.A.; Dos Santos, V.L. Genomic and phenotypic insights into the potential of rock phosphate solubilizing bacteria to promote millet growth in vivo. Front. Microbiol. 2021, 11, 574550. [Google Scholar] [CrossRef]
- Shipton, W.A.; Baker, A.; Blaney, B.J.; Horwood, P.F.; Warner, J.M.; Pelowa, D.; Greenhill, A.R. Nitrogen fixation associated with sago (Metroxylon sqagu) and some implications. Appl. Microbiol. 2010, 52, 56–61. [Google Scholar] [CrossRef]
- Puente-Garza, C.; Gutierrez-Mora, A.; García-Lara, S. Micropropagation of Agave salmiana: Means to production of antioxidant and bioactive principles. Front. Plant Sci. 2015, 6, 1026. [Google Scholar] [CrossRef]
- Sánchez-Urbina, A.; Ventura-Canseco, L.M.C.; Ayora-Talavera, T.; Abud-Archila, M.; Peez-Farrera, M.A.; Dendooven, L.; Gutierrez-Miceli, F.A. Seed germination an in vitro propagation of Agave grijalvensis and endemic endangered mexican species. Asian J. Plant Sci. 2008, 7, 752–756. [Google Scholar] [CrossRef]
- Cartabia, A.; Sarropoulou, V.; Grigoriadou, K.; Maloupa, E.; Eclerck, S.D. In vitro propagation of Alkanna tinctoria Tausch: A medicinal plant of the Boraginaceae family with high pharmaceutical value. Ind. Crops Prod. 2022, 182, 114860. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, X.; Hu, F.; Yang, H.; Yue, L.; Trigiano, R.N.; Cheng, A.M. Micropropagation of Agave americana. Horst. Sci. 2014, 49, 320–327. [Google Scholar] [CrossRef]
- Garriga-Caraballo, M.; Gonzalez-Oramas, G.; Aleman-García, S.; Abreu-Cruz, E.; Quiroz-Bravo, K.; Caligari, P.S.; Garcia-Gonzalez, R. Management of auxin-cytokinin interactions to improve micropropagation protocol of henequen (Agave fourcroydes Lem.). Chilean J. Agric. Res. 2010, 70, 545–551. [Google Scholar] [CrossRef]
- Sanchez, A.; Coronel-Lara, Z.; Gutierrez, A.; Vargas, G.; Coronado, M.L.; Esqueda, M. Acclimatization and transplantation of Agave angustifolia Haw. vitroplants in wild conditions. Rev. Mex. Cien. Agric. 2020, 11, 7. [Google Scholar] [CrossRef]
- Arzate-Fernández, A.M.; Martínez-Velasco, I.; Alvareza-Aragón, C.; Martinez-Martinez, S.Y.; Norman Mondragon, T.H. Morphogenetic response of two agave species regenerated In vitro. Trop. Subtrop. 2020, 23, 47. [Google Scholar]
- Carimi, F.; Zottini, M.; Formentin, E.; Terzi, M.; Schiavo, F.L. Cytokinins: New apoptotic inducers in plants. Planta 2003, 216, 413–421. [Google Scholar] [CrossRef]
- Carimi, F.; Terzi, M.; De Michele, R.; Zottini, M.; Lo Schiavo, F. High levels of the cytokinin BAP induce PCD by accelerating senescence. Plant Sci. 2004, 166, 963–969. [Google Scholar] [CrossRef]
- Vescovi, M.; Riefler, M.; Gessuti, M.; Novák, O.; Schmülling, T.; Lo Schiavo, F. Programmed cell death induced by high levels of cytokinin in Arabidopsis cultured cells is mediated by the cytokinin receptor CRE1/AHK4. J. Exp. Bot. 2012, 63, 2825–2832. [Google Scholar] [CrossRef]
- Kuniskowska, A.; Byczkowska, A.; Doniak, M.; Kaźmierczak, A. Cytokinins remume: Their signaling and role in programmed cell death in plants. Plant Cell Rep. 2013, 32, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Singla-Pareek, S.L.; Pareek, A. Two component signaling system in plants: Interaction network and specificity in response to stress hormones. Plant Cell Rep. 2021, 40, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Veselova, S.V.; Nuzhnaya, T.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Khusnutdinova, E.K.; Maksimov, I.V. Ethylene-Cytokinin interaction determines early defense response of wheat against Stagonospora nodorum Berk. Biomolecules 2021, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Morreeuw, Z.P.; Castillo-Quiroz, D.; Ríos-González, L.J.; Martínez-Rincón, R.; Estrada, N.; Melchor-Martínez, E.M.; Iqbal, H.M.N.; Parra-Saldívar, R.; Reyes, A.G. High throughput profiling of flavonoid abundance in Agave lechuguilla residue-valorizing under explored mexican plant. Plants 2021, 10, 695. [Google Scholar] [CrossRef] [PubMed]
- Puente-Garza, C.; Espinosa-Leal, C.A.; García-Lara, S. Steroidal saponin and flavonol content and antioxidant activity during sporophyte development of Maguey (Agave salmiana). Plant Foods Hum. Nutr. 2018, 73, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Soto-Castro, D.; Pérez-Herrera, A.; García-Sánchez, E.; Santiago-García, P.A. Identification and quantification of bioactive compounds in Agave potatorum Zucc. Leaves at different stages of development and a preliminary biological assay. Waste Biomass Valorization 2021, 12, 4537–4547. [Google Scholar] [CrossRef]
- Puente-Garza, C.A.; Meza-Miranda, C.; Ochoa-Martínez, D.; García-Lara, S. Effect of In vitro drought stress on phenolic acids, flavonols, saponins, and antioxidant activity in Agave salmiana. Plant Physiol. Biochem. 2017, 115, 400–407. [Google Scholar] [CrossRef]
- Erken, O. Some bioactive metabolites’ response to long-term water stress in red cabbage. Sci. Hortic. 2022, 293, 110731. [Google Scholar] [CrossRef]
- Puente-Garza, C.A.; Espinosa-Leal, C.A.; García-Lara, S. Effect of saline elicitors on saponin production in Agave salmiana plants grown In vitro. Plant Physiol. Biochem. 2021, 162, 476–482. [Google Scholar] [CrossRef]
- Qian, H.; Xu, Z.; Cong, K.; Zhu, X.; Zhang, L.; Wang, J.; Wei, J.; Ji, P. Transcriptomic responses to drought stress in Polygonatum kingianum tuber. BMC Plant Biol. 2021, 21, 537. [Google Scholar] [CrossRef]
- Félix-Vald Ayuso, M.; Pinela, J.; Días, M.I.; Barros, L.; Ivanov, M.; Calhelha, R.C.; Soković, M.; Ramil-Rego, P.; Barreal, M.E.; Gallego, P.P.; et al. Phenolic composition and biological activities of the in vitro cultured endangered Eryngium viviparum. J. Gray. Ind. Crops Prod. 2020, 148, 112325. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Hnatuszki-Konka, K.; Krzemińska, M.; Olszewaka, M.A.; Owczarek, A. Cytokinin-based tissue cultures for stable medicinal plant production: Rregeneration and phytochemical profiling of Salvia bulleyana shoots. Biomolecules 2021, 11, 1513. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Singh, P.; Kaur, K.; Bhandawat, A.; Nogia, P.; Pati, P.K. Development of robust In vitro culture protocol for the propagation of generically and phytochemically stable plants of Withania somnifera (L.). Dunal (Ashwagandha). Ind. Crops Prod. 2021, 166, 113428. [Google Scholar] [CrossRef]
- Liu, B.L.; Fan, Z.B.; Liu, Z.Q.; Qiu, X.H.; Jiang, Y.H. Comparison of phytochemical and antioxidant activities in micropropagated and seed-derived Salvia miltiorrhiza plants. Hort. Sci. 2018, 53, 1038–1044. [Google Scholar] [CrossRef]
- Salih, A.M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Aladallah, N.M.; Alansi, S.; Alshameri, A. Mass propagation of Juniperus procera Hoechst. Ex. Endl. From seedling and screening of bioactive compounds in shoot and callus extract. BMC Plant Biol. 2021, 21, 192. [Google Scholar] [CrossRef] [PubMed]
- Riaji, L.; Chograni, H.; Rejeb, F.B.; Romdhane, M.B.; Masmoudi, A.S.; Cherif, A. Efficient in vitro regeneration of the endangered species Artemisia arborescens L. through direct organogenesis and impact on secondary metabolites production. Hortic. Environ. Biotechnol. 2022, 63, 439–450. [Google Scholar] [CrossRef]
- Puente-Garza, C.; García-Lara, S.; Gutiérrez-Uribe, J.A. Enhnancement of saponins and flavonols by micropropagation of Agave salmiana. Ind. Crops Prod. 2017, 105, 225–230. [Google Scholar] [CrossRef]
- Frison-Norrie, S.; Sporns, P. Identification and quantification of flavonol glycosides in almond seedcoats using MALDI-TOF MS. J. Agric. Food Chem. 2002, 50, 2782–2787. [Google Scholar] [CrossRef]
- Ucar, M.B.; Ucar, G.; Pizzi, A.; Gonultas, O. Characterization of Pinus brutia bark tannin by MALDI-TOF MS and ¹³C NMR. Ind. Crops Prod. 2013, 49, 697–704. [Google Scholar] [CrossRef]
- Wang, J.; Sporns, P. MALDI-TOF MS análisis of food flavonol grycosides. J. Agric. Food Chem. 2000, 48, 1657–1662. [Google Scholar] [CrossRef]
- Alecu, A.; Albu, C.; Litescu, S.C.; Eremia, S.A.V.; Radu, G.L. Phenolic and anthocyanin profile of valea calugareasca red wines by HPLC-PDA-MS and MALDI-TOF analysis. Food Anal. Methods 2016, 9, 300–310. [Google Scholar] [CrossRef]
- Fougère, L.; Da Silva, D.; Destandau, E.; Elfakir, C. TLC-MALDI-TOF-MS-based identification of flavonoid compounds using an inorganic matrix. Phytochem. Anal. 2018, 30, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Husseinv, S.; Dishisha, T.; Soliman, H.A.; Adeleke, R.; Raslan, M. Characterization of growth promoting bacterial endophytes isolated from Artemisia annua L. S. Afric. J. Bot. 2021, 143, 238–247. [Google Scholar] [CrossRef]
- White, J.F.; Chang, X.; Kingsley, K.L.; Zhang, Q.; Chiaranunt, P.; Micci, A.; Velazquez, F.; Elmore, M.; Crane, S.; Li, S.; et al. Endophytic bacteria in grass crop growth promotion and biostimulation. Grass Res. 2021, 1, 5. [Google Scholar] [CrossRef]
- Kanani, P.; Modi, A.; Kumar, A. Biotization of endophytes in micropropagation: A helpful enemy. In Microbial Endophytes: Prospects for Sustainable Agriculture; Kumar, A., Singh, V.P., Eds.; Woodhead Publishing: Sawston, UK, 2020; pp. 357–379. [Google Scholar] [CrossRef]
- Soumare, A.; Diédhiou, A.G.; Arora, N.K.; Al-Ani, L.K.T.; Ngom, M.; Fall, S.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L.; Sy, M.O. Potential role and utilization of plant growth promoting microbes in plant tissue culture. Front. Microbiol. 2021, 12, 649878. [Google Scholar] [CrossRef] [PubMed]
- Tamošiūnė, I.; Stanienė, G.; Haimi, P.; Stanys, V.; Rugienius, R.; Baniulis, D. Endophytic Bacillus and Pseudomonas spp. Modulate apple shoot growth, cellular redox balance, and protein expression under In vitro conditions. Front. Plant. Sci. 2018, 9, 889. [Google Scholar] [CrossRef]
- Sneck, M.E.; Rudgers, J.A.; Young, C.A.; Miller, T.E. Does host outcrossing disrupt compatibility with heritable symbionts? Oikos 2019, 128, 892–903. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef]
- Mukherjee, A.; Gaurav-Kumar, A.; Kumar-Patel, P.; Singh, S.; Kumar-Chouhan, K.; Lepcha, A.; Pereira, A.P.A.; Verma, J.K. Unlocking the potential plant growth-promoting properties of chickpea (Cicer arietinum L.) seed endophytes bio-inoculants for improving soil health and crop production. Land Degrad. Dev. 2021, 32, 4362–4374. [Google Scholar] [CrossRef]
- Lima, A.S.; Prieto, K.R.; Santos, C.S.; Paula-Valerio, H.; Garcia-Ochoa, E.Y.; Huerta-Robles, A.; Beltran-Garcia, M.J.; Di Mascio, P.; Bertotti, M. In-vivo electrochemical monitoring of H2O2 production induced by root-inoculated endophytic bacteria in Agave tequilana leaves. Biosens. Bioelectron. 2018, 99, 108–114. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagy cycle: An oxidative process in plants for nutrient extraction from symbiotic microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef]
- Aguilar-Jimenez, M.; Rodriguez, J.L. Micropropagacion y aclimatación de maguey Potzometl (Agave marmorata Roezl) en la mixteca poblana. Rev. Colomb. Biotecnol. 2018, 20, 124–131. [Google Scholar] [CrossRef]
- Martinez-Rodriguez, A.; Macedo-Raygoza, G.; Huerta-Robles, A.X.; Reyes-Sepulveda, I.; Lozano-Lopez, J.; Garcia-Ochoa, E.Y.; Fierro-Kong, L.; Medeiros, M.H.G.; Di Mascio, P.; White, J.F.; et al. Agave seed endophytes: Ecology and impacts on root architecture, nutrient acquisition, and cold stress tolerance. In Seed Endophytes; Verma, S., White, J., Jr., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Benson, A.; Melvin, J.; Balathandayutham, K.; Sa, T.; Chandrasekaran, R. Role of Achromobacter xylosoxidans AUM54 in Micropropagation of Endangered Medicinal Plant Naravelia zeylanica (L.) DC. J. Plant Growth Regul. 2013, 33, 202–213. [Google Scholar] [CrossRef]
- Joe, M.M.; Islam-Rashedul, M.D.; Karthikeyan, B.; Bradeepa, K.; Sivakumaar, P.K.; Sa, T. Resistance responses or fice-to-rice blast fungus after seed treatment with the endophytic Achromobacter xylosoxidans AUM54 strains. Crop Prot. 2012, 42, 141–148. [Google Scholar] [CrossRef]
- Jana, G.; Yaish, M. Genome analysis of a salinity adapted Achromobacter xylosoxidans rhizobacteria from the date palm. Rhizophere 2021, 19, 100401. [Google Scholar] [CrossRef]
- Leite, M.S.; Furtado-Pinto, T.E.; Rabelo-Centofante, A.; Rubio-Neto, A.; Guimaraes-Silva, F.; Gonçalves-Selari, P.J.R.; Martins, P.F. Acclimatization of Pouteria gardneriana Radlk micropropagated plantlets: Role of In vitro rooting and plant growth-promoting bacteria. Curr. Plant Biol. 2021, 27, 100209. [Google Scholar] [CrossRef]
- Pushpakanth, P.; Krishnamoorthy, R.; Anandham, R.; Senthilkumar, M. Biotization of tissue culture banana plantlets with Methylobacterium salsuginis to enhance the survival and growth under greenhouse and open environment condition. J. Environ. Biol. 2021, 42, 1452–1460. [Google Scholar] [CrossRef]
- Rajamanickam, S.; Karthikeyan, G.; Kavino, M.; Manoranjitham, S.K. Biohardening of micropropagated banana using endophytic bacteria to induce plant growth promotion and restrain rhizome rot disease caused by Pectobacterium carotovorum subsp. carotovorum. Sci. Hort. 2018, 231, 179–187. [Google Scholar] [CrossRef]
- Sharma, P.; Kharkwal, A.C.; Abdin, M.Z.; Varma, A. Piriformospora indica improves micropropagation, growth and phytochemical content of Aloe vera L. plants. Symbiosis 2014, 64, 11–23. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Endophyte Bacteria | ACC | IAA | N-BRIP | NFB |
|---|---|---|---|---|---|
| Bt | Bacillus tequilensis | + | + | + | + |
| Ec | Enterobacter cloacae | + | − | + | + |
| Ax | Achromobacter xylosoxidans | + | − | − | + |
| Pa | Pseudomonas aeruginosa | + | − | − | + |
| Embryonic Leaf | Seedlings | |||||||
|---|---|---|---|---|---|---|---|---|
| Lot | Seed | Germination (d) | Appearance of the Embryonic Leaf (d) | Length (cm) | Width (cm) | Length (cm) | Secondary Roots (n) | Leaves Formed (n) |
| L12 | M06 | 3 | 4 | 3.0 | 0.3 | 3.8 | 2 | 2 |
| M14D-E | 3 | 4 | 3.2 | 0.5 | 4.2 | 3 | 2 | |
| AM20 | 3 | 5 | 3.0 | 0.3 | 3.7 | 2 | 2 | |
| AM21 | 3 | 5 | 3.0 | 0.3 | 4.0 | 2 | 2 | |
| L18 | AM23 | 3 | 5 | 3.0 | 0.4 | 4.0 | 2 | 2 |
| AM24 | 3 | 6 | 3.0 | 0.3 | 4.0 | 2 | 2 | |
| AM31 | 3 | 4 | 3.2 | 0.5 | 4.2 | 3 | 2 | |
| AM32 | 3 | 4 | 3.2 | 0.5 | 4.3 | 3 | 2 | |
| AM33 | 3 | 4 | 3.2 | 0.5 | 4.2 | 2 | 2 | |
| AM34 | 3 | 4 | 3.0 | 0.3 | 4.0 | 2 | 2 | |
| Sample | Total Phenolic Content (mgGAE/g Crude Dried Extract) Mean ± SE | Total Flavonoid Content (mg Quercentin E/g Crude Dried Extract) Mean ± SE | Total Steroidal Saponin Content (mg Diosgenin E/g Crude Dried Extract) Mean ± SE | Total Saponin Content (mg Diosgenin E/g Crude Dried Extract) Mean ± SE |
|---|---|---|---|---|
| AM31 | 15.28 ± 0.78 b | 13.94 ± 0.19 b | 10.79 ± 2.84 a | 42.46 ± 0.05 a |
| AM32 | 12.90 ± 1.38 ab | 11.22 ± 1.31 b | 12.71 ± 3.87 a | 43.59 ± 3.45 a |
| AM33 | 13.89 ± 0.35 ab | 12.20 ± 2.02 b | 10.49 ± 3.81 a | 47.66 ± 2.57 a |
| HAM | 11.20 ± 2.36 a | 1.67 ± 0.10 a | 12.74 ± 0.10 a | 48.42 ± 3.78 a |
| Fraction | tR (min) | (λ nm) | Percentage by Normalization Area (%) | m/z (+)/Ion Assignment | Tentative Identification | Metabolite Class | ||
|---|---|---|---|---|---|---|---|---|
| AM31 | AM32 | AM33 | ||||||
| F1 | 2.42 | 205 | 7.01 ± 4.25 a | 24.19 ± 1.57 b | 8.15 ± 4.21 a | 224, 329, 581 636, 782, 1070, 1200 | Unknown | Mixture of compounds |
| F2 | 2.95 | 275 | 5.17 ± 0.29 a | 3.69 ± 0.92 a | 3.57 ± 0.69 a | 290 [M], 581 [2M+H, dimer], 870 [3M+H, trimer], 1201 [4M+H, tetramer+K] | Catechin | Phenolic |
| F5 | 5.37 | 275 | 8.28 ± 1.84 a | 6.93 ± 0.81 a | 6.97 ± 1.67 a | 171 [M], 343 [2M+H, dimer], 512 [3M+H, trimer], 684 [4M+H, tetramer] | Gallic Acid | Phenolic |
| F7 | 8.47 | 275 | 0.73 ± 0.57 a | 3.32 ± 0.54 b | 3.04 ± 1.27 b | 635 [M+H+Rutinoside] | Isorhamnetin Rutinoside | Flavonoid |
| F8 | 9.89 | 275 | 10.68 ± 6.22 a | 1.49 ± 0.32 b | 1.90 ± 0.04 b | 482 [M+H−Glucuronide] | Myricetin-3-Glucoside | Flavonoid |
| F12 | 17.35 | 275 | 1.20 ± 0.15 a | 5.06 ± 0.56 b | 4.69 ± 0.87 b | 496 [M+H−Glucuronide] | Myricetin-3-Glucuronide | Flavonoid |
| F14 | 19.75 | 205 | 2.86 ± 1.19 a | 0.77 ± 0.42 b | 1.15 ± 0.05 b | 628 [M+H+Rhamnosylglucoside] | Myricetin-3-Rutinoside | Flavonoid |
| F15 | 20.63 | 275 | 0.3 ± 0.17 a | 3.46 ± 0.42 b | 3.85 ± 1.71 b | 180 [M+H], 360 [M+H, dimer], 539 [M+H, trimer],745 [M+H, tetramer] | Caffeic Acid | Phenolic |
| F18 | 25.08 | 518 | 12.99 ± 2.21 a | 15.60 ± 0.51 a | 14.37 ± 5.56 a | 303 [M], 484 [M+Glycoside+Na] | Delphinidin-3-O-Glucoside | Anthocyanin |
| F22 | 27.72 | 275 | 1.80 ± 0.70 a | 1.06 ± 0.09 a | 1.65 ± 0.45 a | 607 [M+H] | Rutin | Flavonoid |
| F23 | 28.23 | 518 | 35.47 ± 2.28 a | 23.08 ± 0.25 b | 21.62 ± 4.52 b | 484 [M+Glycoside+K] 613 [M+2glyc+Na-18] | Cyanidin 3,5-Diglucoside | Anthocyanin |
| F24 | 29.45 | 518 | 5.12 ± 0.70 a | 3.97 ± 1.19 a | 4.44 ± 0.42 a | 484 [M+Glycoside+Na], 496 [M+Glycoside+K], 656 [M+2Glycoside+K] | Delphinidin-3,5-O-Diglucoside | Anthocyanin |
| F25 | 29.87 | 390 | 2.08 ± 0.50 a | 3.95 ± 1.33 a | 4.73 ± 1.23 a | 296 [M], 484 [M+H+Glucoronide+Na] | Quercetin-Glucuronide | Flavonoid |
| F28 | 34.15 | 275 | 2.17 ± 0.33 a | 2.88 ± 1.09 a | 1.75 ± 0.23 a | 224 [M] 455 [2M+H, dimer] 736 [3M+H, trimer] | Sinapic Acid | Phenolic |
| F29 | 34.60 | 275 | 23.79 ± 0.11 a | 12.71 ± 2.98 b | 12.26 ± 0.83 b | 621 [M+H+2Glucoside] | Quercetin-3-O-Diglucoside | Flavonoid |
| F30 | 35.87 | 275 | 5.20 ± 0.62 a | 3.78 ± 1.53 a | 3.71 ± 0.57 a | 465 [M+H+Glucoside] | Quercetin-3-Glucoside | Flavonoid |
| F31 | 38.17 | 275 | 6.53 ± 3.69 a | 5.62 ± 0.46 a | 5.67 ± 0.80 a | 605 [M+H+2Glucoside] 628 [M+H+2 Glucoside+Na] | Kaempferol-Diglucoside | Flavonoid |
| F32 | 38.97 | 275 | 2.53 ± 0.28 a | 0.70 ± 0.11 b | 1.15 ± 0.64 b | 635 [M+H+2Glucoside] | Isorhamnetin-3-O-Di-glucoside | Flavonoid |
| F37 | 49.89 | 275 | 3.51 ± 0.33 a | 5.05 ± 1.63 a | 4.60 ± 0.07 a | 1332 [M+H−4hexose-2pentose] | Tigogenin | Saponin |
| F42 | 56.38 | 275 | 0.55 ± 0.21 a | 1.18 ± 0.61 a | 1.49 ± 0.86 a | 496 [M+H+Glucoronide] | Isorhamnetin-Glucuronide | Flavonoid |
| F44 | 59.42 | 275 | 0.84 ± 0.71 a | 3.63 ± 1.99 ab | 4.87 ± 1.55 b | 473 [M+H+Glucoside] | Isorhamnetin-3-Glucoside | Flavonoid |
| F 47 | 63.43 | 275 | 0.32 ± 0.19 a | 1.01 ± 0.59 a | 1.03 ± 0.51 a | 1185 [M+H−3Hexose-2Pentose] | Hecogenin | Saponin |
| F 48 | 64.53 | 275 | 0.30 ± 0.24 a | 0.85 ± 0.47 a | 1.14 ± 0.39 a | 1056 [M+H−3Hex-Pen] | Hecogenin | Saponin |
| F62 | 84.92 | 205/390 | 4.06 ± 2.14 a | 3.55 ± 0.02 a | 2.76 ± 0.51 a | 496 [M+H+Glucoronide] | Myricetin-3-Glucoronide | Flavonoid |
| F66 | 90.02 | 390 | 6.25 ± 2.47 a | 9.77 ± 0.17 b | 8.01 ± 0.95 ab | 287 [M] 595 [M+Rutinose] | Kaempferol Rutinoside | Flavonoid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Rodriguez, A.; Beltran-Garcia, C.; Valdez-Salas, B.; Santacruz-Ruvalcaba, F.; Di Mascio, P.; Beltran-Garcia, M.J. Micropropagation of Seed-Derived Clonal Lines of the Endangered Agave marmorata Roezl and Their Compatibility with Endophytes. Biology 2022, 11, 1423. https://doi.org/10.3390/biology11101423
Martinez-Rodriguez A, Beltran-Garcia C, Valdez-Salas B, Santacruz-Ruvalcaba F, Di Mascio P, Beltran-Garcia MJ. Micropropagation of Seed-Derived Clonal Lines of the Endangered Agave marmorata Roezl and Their Compatibility with Endophytes. Biology. 2022; 11(10):1423. https://doi.org/10.3390/biology11101423
Chicago/Turabian StyleMartinez-Rodriguez, America, Celia Beltran-Garcia, Benjamin Valdez-Salas, Fernando Santacruz-Ruvalcaba, Paolo Di Mascio, and Miguel J. Beltran-Garcia. 2022. "Micropropagation of Seed-Derived Clonal Lines of the Endangered Agave marmorata Roezl and Their Compatibility with Endophytes" Biology 11, no. 10: 1423. https://doi.org/10.3390/biology11101423