

Roles of Gut Microbiome in Bone Homeostasis and Its Relationship with Bone-Related Diseases
by
 , , ,
, , ,
Nina Zemanova
1,
Radoslav Omelka
1,*,
Vladimira Mondockova
1,
Veronika Kovacova
2 and
Monika Martiniakova
2,* 1
Department of Botany and Genetics, Faculty of Natural Sciences and Informatics, Constantine the Philosopher University in Nitra, 949 74 Nitra, Slovakia
2
Department of Zoology and Anthropology, Faculty of Natural Sciences and Informatics, Constantine the Philosopher University in Nitra, 949 74 Nitra, Slovakia
*
Authors to whom correspondence should be addressed.
Biology 2022, 11(10), 1402; https://doi.org/10.3390/biology11101402
Submission received: 12 August 2022
/
Revised: 14 September 2022
/
Accepted: 19 September 2022
/
Published: 26 September 2022
(This article belongs to the Special Issue Gut Microbiome in Health and Disease)
Abstract
:Simple Summary
In recent years, there has been increasing evidence that communication between the skeletal system and the gut microbiome (GM) can influence bone health and that the GM is a key regulator of bone homeostasis. Here, we review the roles of GM in bone homeostasis. In addition, the relationship between GM composition and selected bone-related diseases (osteoporosis, osteoarthritis, rheumatoid arthritis, diabetes mellitus, obesity and bone cancer) is presented. It is also emphasized that a probiotic supplementation can play an important role in suppressing the symptoms of each of these diseases.
Abstract
The extended microbial genome—the gut microbiome (GM)—plays a significant role in host health and disease. It is able to influence a number of physiological functions. During dysbiosis, GM is associated with the development of various chronic diseases with impaired bone quality. In general, GM is important for bone homeostasis and can affect it via several mechanisms. This review describes the roles of GM in bone homeostasis through influencing the immune and endocrine functions, short-chain fatty acids production, calcium absorption and the gut–brain axis. The relationship between GM composition and several bone-related diseases, specifically osteoporosis, osteoarthritis, rheumatoid arthritis, diabetes mellitus, obesity and bone cancer, is also highlighted and summarized. GM manipulation may become a future adjuvant therapy in the prevention of many chronic diseases. Therefore, the beneficial effects of probiotic therapy to improve the health status of individuals with aforementioned diseases are provided, but further studies are needed to clearly confirm its effectiveness. Recent evidence suggests that GM is responsible for direct and indirect effects on drug efficacy. Accordingly, various GM alterations and interactions related to the treatment of bone-related diseases are mentioned as well.
1. Introduction
The gut microbiota is a unique and dynamic community of microorganisms residing in a specific environment—the gastrointestinal tract (GIT) [1]. The gut microbiome (GM), often used as a synonym for gut microbiota, is defined as a set of all genomic elements of a specific microbiota, representing an “extended genome” of millions of microbial genes. This microbial community consists not only of bacteria but also of archaea, viruses and unicellular eukaryotes [2].
The extended microbial genome—the microbiome—plays an important role in host health and disease [3]. The microbiome is able to affect numerous physiological functions, mostly through microbial metabolites, (e.g., nutrient and xenobiotic metabolism, protection against pathogens and regulation of the immune system and enteric nervous system) [4]. However, the gut microbiota is not stable over time, and factors that have been shown to alter the GM include aging, diet, environment or chronic treatment with oral antibiotics [5]. In general, decreased microbial diversity can be associated with dysbiosis (defined as an abnormal ratio of commensal and pathogenic bacteria), as there is a link between microbiome dysbiosis and various diseases [6,7].
As mentioned above, GM is able to produce multiple compounds that reach the circulation; hence, they can influence the functions of distal organs [8]. Nowadays, there is a growing interest in revealing the role of the microbiota in human health. Interestingly, there is a link between GM and skeletal health, and the microbiome also contributes to the regulation of bone homeostasis [9]. The GM–bone axis refers to the communication between microbial community and their metabolites on bone health. New findings support the information that microbiota is able to impact bone mineral density (BMD) and strength parameters. This can lead to a possibility to use beneficial bacteria as a future adjuvant probiotic therapy [10].
In this review, special emphasis is given to the relationship between GM composition and several chronic diseases consistent with impaired bone mass and bone quality, such as osteoporosis, osteoarthritis, rheumatoid arthritis, diabetes mellitus, obesity and bone cancer. Favorable impacts of probiotic therapy to alleviate symptoms in the above-mentioned disorders are also emphasized. The roles of GM in bone homeostasis through several mechanisms are reported as well. After all, various GM changes and interactions associated with the treatment of bone-related diseases are listed.
2. Materials and Methods
For this review, the PubMed database was used for the search of scientific articles. All searches were up to date as of 2022, and the search was conducted between March and August 2022. The search terms used included: “gut microbiome”, “bone-related diseases”, “bone health”, “bone diseases”, “bone homeostasis”, “osteoporosis”, “diabetes mellitus”, “osteoarthritis”, “rheumatoid arthritis”, “bone cancer”, “primary bone cancer”, “obesity”, “probiotics”, “probiotic supplementation”, “treatment” and “therapy”. Additionally, the combination of aforementioned keywords was used, and these combinations were connected with “and” (combinations of basic term “gut microbiome” and relevant disease, supplement or treatment) and “or” (between terms). The articles not related to the aims of the review were excluded. After analyzing the abstracts, the full text of each paper was checked. Only articles written in English were selected.
3. Composition and Functions of Gut Microbiome
GM is a dynamic system that is able to change over time. The first and most important step is a vertical transmission of maternal microbiota. This microbiota transmission and, subsequently, the colonization process of infants are influenced by the mode of delivery. Infants born by vaginal delivery, the most common mode of delivery, have colonization reflecting the maternal vaginal flora (Lactobacillus and Prevotella species). On the other hand, the microbiome of infants born by cesarean delivery consists of epidermal species (such as Clostridium, Staphylococcus, Propionobacterium and Corynebacterium) rather than vaginal ones, with lower numbers of Bacteroides and Bifidobacterium [11]. The GM undergoes rapid changes during infancy and early childhood and may also be formed by other factors, such as diet and antibiotics [12]. Another important factor influencing the GM of infants is the mode of feeding. The breastfed infant microbiome has a significantly higher numbers of Bifidobacteria and Lactobacillus and lower numbers of Bacteroides, Staphylococcus and Enterobacteriaceae in comparison with formula-fed infants. In addition, breast milk is a source of oligosaccharides that have a prebiotic effect [13]. Several studies [12,14,15] indicate that the GM continues to develop with a gradual increase in microbial diversity into adolescence. The pre-adolescent microbiome is enriched with Bifidobacterium spp., Faecalibacterium spp. and members of the Lachnospiraceae family [12]. In healthy adults, the microbiome is considered stable until old age (65 or older) and is shaped by environmental factors, such as diet and medication, rather than the genetics [12,16]. For example, animal-based diet is able to increase the abundance of bile-tolerant microorganisms (Alistipes, Bilophila and Bacteroides), reflecting an increased need of protein fermentation. On the other hand, in herbivorous mammals, there is a decreased level of Firmicutes (Roseburia, Eubacterium rectale and Ruminococcus bromii) metabolizing dietary plant polysaccharides [17]. In addition, diets high in fat, mainly in saturated fats, increase the abundance of Firmicutes, Proteobacteria and Bilophila spp. [18]. The microbial composition can also be affected by specific drugs, as several studies have reported [19]. Interestingly, changes in composition and diversity are associated with biological (functional) age, independent of chronological age (e.g., co-abundance module consisting of Ruminococcus, Coprobacillus and Eggerthella genera becomes abundant). However, with increasing biological age, the microbiota richness decreases [20].
In healthy individuals, most microbes inhabiting the GIT belong to two major phyla—Firmicutes and Bacteroidetes—representing more than 90% of relative abundance of the gut microbiota, followed by Actinobacteria, Proteobacteria, Synergistetes, Fusobacteria and others [21]. The Firmicutes phylum includes genera such as Bacillus, Lactobacillus, Clostridium, Enterococcus and Ruminicoccus. The Bacteroidetes phylum predominantly consists of Bacteroides and Prevotella [22]. However, even among the healthy individuals, the composition of the microbiota may vary, because each individual has their own unique microbiome. Hence, to define a healthy GM, we need to look at the functional level, as most people carry the same amount of bacterial genes involved in metabolic pathways [23].
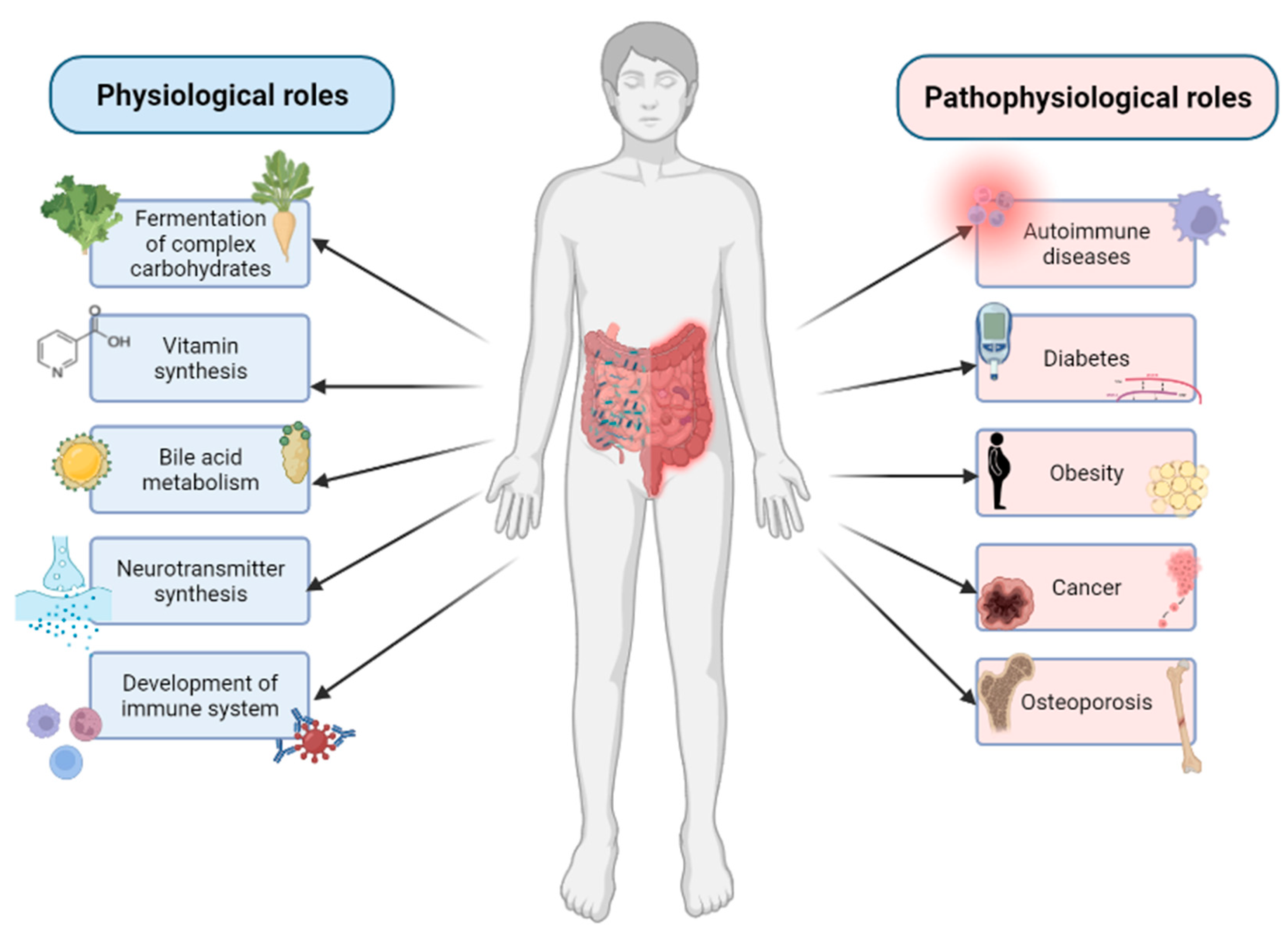
The GM is able to affect the host organism and play a significant role in many physiological functions (Figure 1).
For example, specific dominant bacterial species are known for the fermentation of complex dietary carbohydrates, and some of more nutritionally specialized bacteria are also able to initiate degradation of plant cell walls, starch particles and mucin [24]. The gut microbiota can also affect the largest immunological organ—the GIT. GIT is not only the main line of defense against pathogens, limiting their direct contact with the epithelium, but the bacterial community of GIT is also necessary for the development of a fully functional immune system [25]. In addition, the intestinal microbiota is capable to affect the central nervous system through multiple bidirectional pathways involving neural, endocrine and immune signaling and is known as gut–brain axis. The microbiota can produce neurotransmitters and secrete neuroactive metabolites [26]. It can also synthesize certain vitamins, namely vitamin K and B group vitamins, such as thiamine (B1), riboflavin (B2), niacin (B3), pantothenic acid (B5), pyridoxine (B6), biotin (B7), folate (B9) and cobalamin (B12). The synthesis of these vitamins is not only important for intestinal bacteria, but there is also a metabolic and physiological significance of these pathways in mammals [27]. The GM also plays an important role in the bile acid metabolism. Gut bacteria contribute to the salvage of bile salts that escape active transport in the distal ileum. In addition, secondary bile acids (deoxycholic acid and lithocholic acid) are produced solely by bacteria in the human large intestine [28]. However, during dysbiosis, the GM is associated with development of various metabolic and immune diseases, including obesity, diabetes, inflammatory bowel disease, Crohn’s disease, cancer, allergies and osteoporosis (Figure 1) [7].
4. The Role of Gut Microbiome in Bone Homeostasis
The skeleton forms a structural support of vertebrates with the ability to resist the mechanical forces. It has mechanisms to grow and change its shape and size, as bone tissue is continuously formed and remodeled via two separate processes—bone resorption and bone formation—managed by osteoclasts (OCs) and osteoblasts (Obs) [29,30]. Bone homeostasis is a dynamic process, regulated by complex molecular pathways, where Ocs resorb bone tissue and Obs produce new bone tissue; thus, the net bone mass is maintained. Additionally, bone marrow endothelial cells were found to have a role in regulating bone homeostasis [31]. The bone is also a major source of inorganic ions and actively participates in calcium and phosphate balance [30]. In addition, the bone is a metabolically active organ and is important for the immune system, as immune cells originate from bone marrow, where many of them also mature [32]. Generally, GM can affect bone homeostasis by influencing immune function, host metabolism, hormone secretion and the gut–brain axis.
4.1. GM Influencing Bone Homeostasis via Immune Function
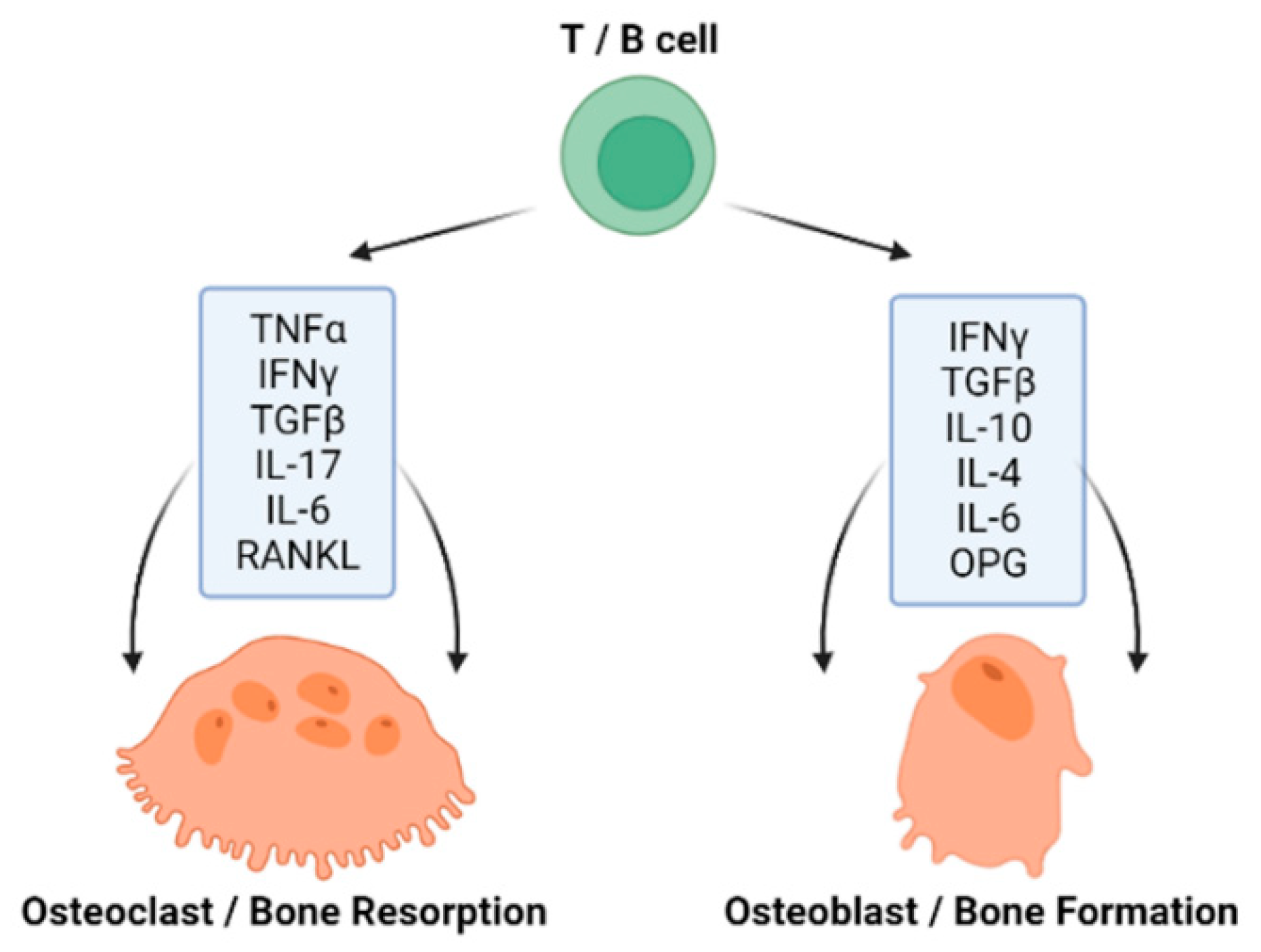
The immune system represents a complex network of cells, compounds and processes that defend and protect the organism from foreign antigens and is tightly connected with bone homeostasis (Figure 2) [33]. An acquired immunity and the skeletal system have been recorded to evolve simultaneously, as there are molecules, such as receptor activator of nuclear factor κ-B ligand (RANKL) and the tumor necrosis factor (TNF) family, which have a significant role in both systems [34]. T cells (T-lymphocytes) are involved in both bone resorption and bone formation through the expression of pro- and anti-inflammatory cytokines, as well as RANKL and OPG [35]. Interestingly, B cells (B-lymphocytes) also produce the key osteoclastogenic cytokine RANKL and its physiological inhibitor osteoprotegerin (OPG) [36,37]. The process of OCs formation requires specific cytokines, primarily RANKL; macrophage colony-stimulating factor (M-CSF); TNF-α; interleukin (IL)-1, IL-7, IL-17, IL-23 and IL-6; interferon (IFN) γ and transforming growth factor-β (TGF-β). OBs are able to express the parathyroid hormone (PTH) receptor, which, upon ligand binding, can activate OC activity. OBs also produce RANKL and OPG. While RANKL stimulates osteoclastogenesis, OPG inhibits this effect [38]. The process of OB formation also requires specific T-lymphocyte cytokines such as IL-10, IL-4, IL-6 and IFN γ [37].
The GM has a great impact on the host organism, so it is not surprising that it also contributes to the regulation of bone homeostasis. This homeostasis is also affected by the immune system; hence, there is a close link between immunity and gut microbiota [9]. The relationship between gut microbiota, immune system and bone health is a relatively new area of study, as Ohlsson and Sjögren [39] even proposed a new interdisciplinary research field called “osteomicrobiology”, combining bone physiology, gastroenterology, immunology and microbiology.
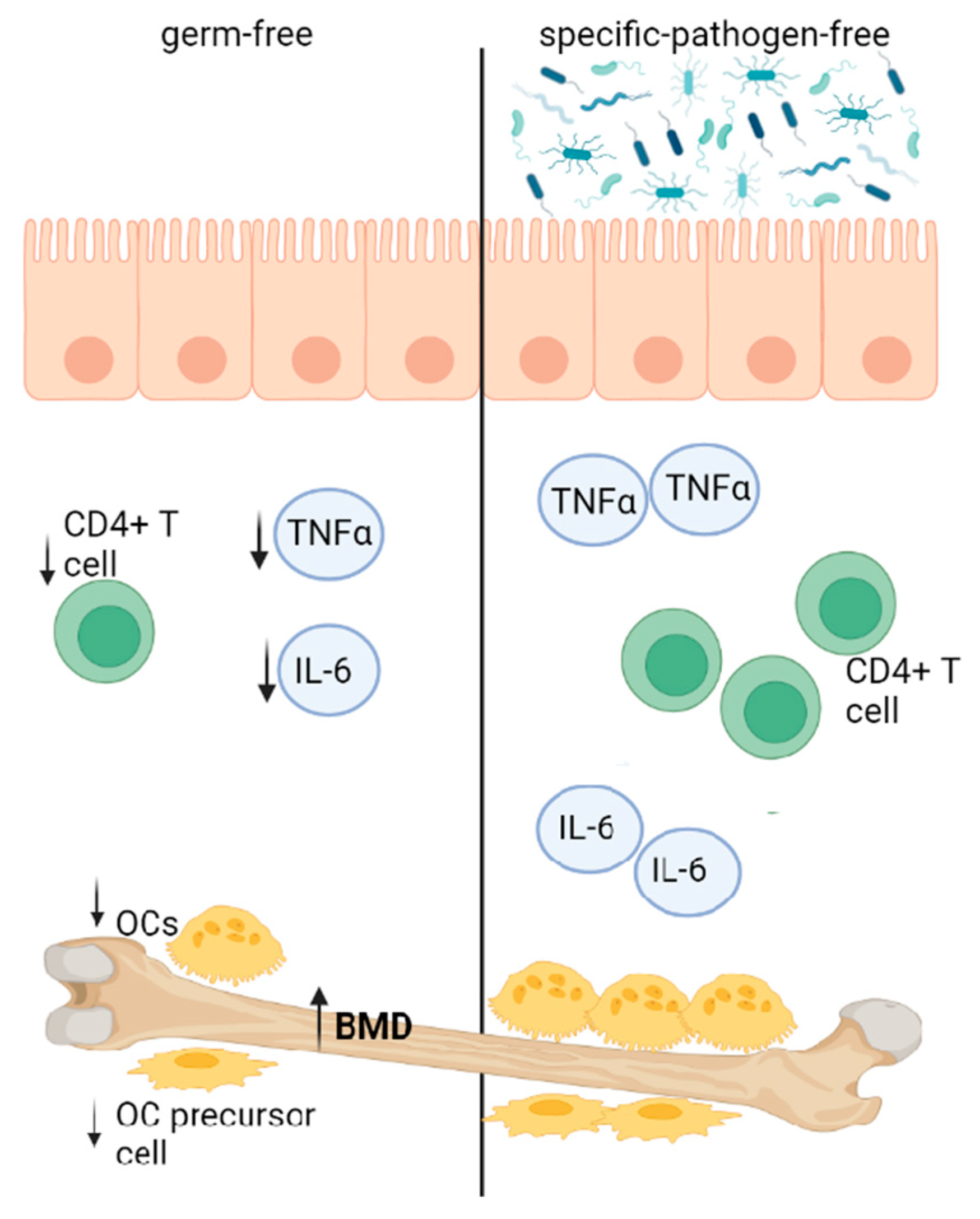
In general, the gut microbiota can affect bone quality and quantity. In the study by Sjögren et al. [40], germ-free (GF) mice exhibited increased trabecular BMD. A higher bone mass was associated with decreased numbers of OCs, CD4+ T cells and OC precursor cells and lowered TNFα and IL-6 expression. Interestingly, the colonization of GF mice resulted in normal bone mass levels (Figure 3).
Another study looked at the possibility of innate immune signaling pathways mediating the effect of the microbiota. This was investigated in GF and CONV-R (conventionally raised) mice with the targeted inactivation of MYD88, NOD1 or NOD2 [43]. NOD1 and NOD-2 are specialized NOD-like receptors, and they have an important role in recognizing pathogens and activating immune responses [44]. On the other hand, myeloid differentiation factor 88 (MyD88) is used by toll-like receptors (TLRs) and activates NF-kB and mitogen-activated protein kinases (MAPKs) to induce the production of inflammatory cytokines [45]. The study has shown that the cortical bone mass was elevated in wild-type and MyD88-deficient GF mice compared to the corresponding CONV-R mice. This increase was not detected in the Nod1−/− or Nod2−/− GF mice. The bone expression of TNFα and RANKL was reduced in GF mice (compared to CONV-R wild-type mice). However, this trend was not observed in Nod1−/− or Nod2−/− mice. This suggests that the effect of gut microbiota on the bone mass depends on the NOD1 and NOD2 signaling pathways [43]. On the contrary, Schwarzer and colleagues [46] demonstrated that wild-type microbiota is associated with optimal systemic somatic growth, as the microbiota influences skeletal growth. The bone growth parameters were decreased in GF animals, although the cortical BMD was not affected. In addition, insulin-like growth factor 1 (IGF-1) was significantly reduced. These contradictory findings may be a result of several differences between animals (different sex, age or strain) in published research.
4.2. GM Influencing Bone Homeostasis via Short-Chain Fatty Acids Production
Microbial products such as bacterial fermentation molecules—short-chain fatty acids (SCFAs)—can also affect bone homeostasis. SCFAs (mainly acetate, propionate and butyrate) are products of carbohydrate fermentation that escape digestion and absorption in the small intestine [47].
Wallimann and colleagues [48] showed that butyrate can significantly reduce OC formation and resorption activity. They also demonstrated that butyrate supplementation can reduce systemic IL-6 levels in a murine model of osteotomy. In another study, Lucas et al. [49] also wanted to determine whether SCFAs are able to affect bone metabolism in two murine models of inflammatory arthritis. Butyrate and propionate have been shown to have a beneficial impact on bone homeostasis. The effect of these SCFAs depends on metabolic reprogramming of pre-osteoclasts, resulting in enhanced glycolysis at the expense of oxidative phosphorylation, leading to the downregulation of essential osteoclast genes such as TRAF6 and NFATc1. SCFAs can also help to alleviate inflammation by inducing Treg cells.
4.3. GM Influencing Bone Homeostasis via Absorption of Calcium
GM can affect the absorption of nutrients related to skeletal development, such as calcium and vitamin D. The skeleton contains 99% of all calcium in the body [29]. The integrity of calcium and phosphate homeostatic mechanisms mediated by parathyroid hormone, fibroblast growth factor 23 and vitamin D is required for normal bone formation, metabolism and repair [50]. In the distal intestine (mostly the ileum), approximately 70–80% of the ingested calcium is absorbed and is facilitated by vitamin D—specifically, by 1, 25-dihydroxyvitamin D3. Calcium absorption involves a calcium influx, translocation of calcium through the enterocyte and basolateral extrusion of calcium by the intestinal plasma membrane pump [51].
A couple decades ago, it has been shown that the presence of SCFAs affected the epithelial tissue and promoted the calcium absorption from the large rat intestine in vitro (using an Ussing chamber) [52]. However, this study did not support a popular theory that a reduced pH environment (caused by the fermentation of prebiotic fiber to SCFAs) can help to elevate the absorption of calcium, because treating the rat colonic segments with HCl did not increase the calcium transport. Several studies have been focused on the effects of prebiotic supplementation (prebiotic dietary fibers, which are fermented by bacteria to SCFAs) and its interactions with mineral absorption. Bryk et al. [53] analyzed the effect of supplementing diets with the GOS/FOS® mixture (a prebiotic mixture of 90% of short-chain galacto-oligosaccharides (GOS) and 10% long-chain fructo-oligosaccharides (FOS)) in growing rats. This mixture increased the bone mineralization and BMD due to higher calcium, phosphorus and magnesium absorptions. Another study investigated the impact of dietary inulin in gluten-free diet and its ability to influence the absorption of calcium [54]. Interestingly, the effect of inulin on mineral utilization depended on the dietary calcium intake from diet. Increased relative calcium absorption from inulin consumption was observed in rats fed with a calcium-restricted gluten-free diet. In addition, some studies also suggest the beneficial effects of prebiotics on calcium absorption in humans. For example, Whisner and colleagues [55] showed that soluble corn fiber (a nondigestible carbohydrate) can increase calcium absorption efficiency in adolescent males and females. A positive impact of a product rich in transgalacto-oligosaccharides (TOS) consumption on calcium absorption was also recorded in postmenopausal women [56]. The increase of calcium absorption was not accompanied by increased urinary excretion, meaning that TOS may indirectly elevate the uptake of calcium by bones and/or inhibit bone resorption.
4.4. GM Influencing Bone Homeostasis via Endocrine Function
The endocrine system (including parathyroid hormone, PTH-related peptide, calcitonin, vitamins A and D, estrogens, androgens and growth hormone) controls a balance between bone formation and bone loss and the maintenance of calcium and phosphate homeostasis [30]. In addition, there is evidence that osteocalcin is also required to stimulate bone mineral maturation [30].
Estrogen receptors are expressed in various tissues, including the brain, bone and adipose tissue; hence, estrogen is able to affect a variety of physiological responses, such as BMD [57]. Li et al. [58] showed that GM modulates the inflammatory responses (upregulated osteoclastogenic cytokines, such as TNFα, RANKL and IL-17) caused by sex steroid deficiency, which leads to trabecular bone loss in a murine model. On the other hand, in GF mice, sex steroid deficiency did not increase osteoclastogenic cytokines production. Additionally, an increase of epithelial barrier permeability and decreased expression of gut epithelial tight junction proteins were recorded.
Vitamin D is mainly known for its role in bone homeostasis (calcium and phosphate absorption), but it has various direct and indirect regulatory effects on the immune system (promoting Treg cells, inhibiting differentiation of Th1 and Th17 cells, impairing the development and function of B cells and reducing monocyte activation) [59]. It has been shown that the GM can be altered by vitamin D. According to Ooi and colleagues [60], the absence of a vitamin D receptor and the ability to produce an active form of vitamin D are associated with inflammation of the intestine, which is caused by the expansion of Proteobacteria phylum (including the Helicobacteraceae family members). Hence, vitamin D supplementation could potentially protect against gastrointestinal injury. Another study, conducted by Luthold et al. [61], supported the effect of vitamin D on the commensal bacterial composition. Interestingly, vitamin D is also able to directly inhibit mycobacterial growth in culture [62].
Osteocalcin is a non-collagenous protein secreted by OBs containing three glutamate residues that can be carboxylated. This modification is catalyzed by γ-glutamyl carboxylase, depending on vitamin K, O2 and CO2 as the cofactors, supplied by the vitamin K cycle and circulation [63]. Carboxylated osteocalcin is essential for the alignment of apatite crystals and optimal bone strength [64]. As the GM has been suggested to contribute to the maintenance of vitamin K, Wagatsuma et al. [65] aimed to clarify the association between GM and alternative indicator of vitamin K deficiency (serum undercarboxylated osteocalcin concentration) in patients with Crohn’s disease. They recorded a significant negative correlation between serum undercarboxylated osteocalcin and mean Chao1 index (determining the alpha diversity).
4.5. GM Influencing Bone Homeostasis via the Gut–Brain Axis
The GM, namely certain bacteria (Streptococcus, Corynebacterium and E. coli), can produce various molecules, including monoamine neurotransmitters such as serotonin, which is a key neurotransmitter within both the enteric and central nervous systems [8].
Peripheral serotonin, produced primarily by the intestinal flora, functions as a hormone inhibiting OB proliferation through 5-hydroxytryptamine receptor 1B and the cAMP response element-binding protein, causing the decrease of BMD and bone formation [66]. On the other hand, brain-derived serotonin appears to be a positive regulator of bone development, acting via sympathetic tone, as well as a molecule regulating food intake and energy expenditure [67]. Supporting these findings, Sjögren et al. [40] showed that GF mice compared to CONV-R mice had lower levels of serum serotonin and an increased relative volume of trabecular bone. However, the colonization of GF mice led to a normalization of trabecular bone mass, but it has no significant effect on the serum serotonin levels.
In addition, tryptophan is a precursor to serotonin, and changes in the tryptophan levels were recorded in bone metabolic diseases [68]. The discovery of the exact role of tryptophan or tryptophan metabolites in bone may result in finding a new effective therapy for management of bone-related diseases. For example, D-tryptophan is a newly identified product from probiotic bacteria [69]; however, more studies are needed to clarify its effectiveness as prebiotic supplement.
5. The Relationship between Gut Microbiome and Bone-Related Diseases
Various diseases are accompanied by changes in GM composition [6]. Bone-related diseases can also be linked to altered microbiome, with significant changes in levels of phyla Firmicutes, Bacteroidetes and Actinobacteria [35]. Additionally, recent studies are focusing on the potential positive effects of GM modulators, such as probiotic and prebiotic supplementations, in both human and animal models. The following text describes in more detail the association between GM and several bone-related diseases, including osteoporosis, osteoarthritis and rheumatoid arthritis, diabetes mellitus (DM), obesity and bone cancer.
5.1. Osteoporosis
Osteoporosis is the most common skeletal disorder, affecting approximately 200 million individuals worldwide [50]. It is characterized by reduced BDM, the microstructural degradation of bone tissue and increased risk of fractures [69]. At the microstructural level, increased bone resorption and decreased bone formation simultaneously occur, leading to a decline in bone mass. The treatment of osteoporosis can be divided into four categories: lifestyle changes (e.g., dietary adjustment, exercise, smoking cessation and avoiding heavy alcohol consumption); nutritional supplements (increased calcium and vitamin D intake); pharmaceutical intervention-antiresorptive agents (e.g., bisphosphonates, selective estrogen receptor modulators, calcitonin and hormone therapy) and anabolic agents (such as parathyroid hormone analogues and strontium ranelate) and surgical management [70,71,72]. In addition, some studies support the idea of importance of circadian rhythmicity for bone health, as sleep disruption and circadian misalignment have been associated with lower BMD and increased fracture risk [73]. Although melatonin is mostly known as an important regulatory factor of circadian rhythm, there is a possibility of its function as a protector in bone injury and osteoporosis; therefore, melatonin supplementation might positively affect bone tissue [74]. Melatonin is secreted from the pineal gland; however, it is also synthesized in the intestines by enterochromaffin cells and gut microbes, where it acts locally rather than entering systemic circulation. There is more melatonin in the gut than in the pineal glands, which proves its importance to gut function [75]. Since melatonin is synthesized from tryptophan with the intermediate serotonin, some drugs such as selective serotonin reuptake inhibitors (SSRIs; the most commonly prescribed antidepressants) can interfere with melatonin production.
Several studies have evaluated the abundance and diversity of bacterial populations in the intestine of patients with osteoporosis (Table 1). Based on these, adults with osteoporosis appear to have a reduced diversity of microorganisms, with an increase in certain species such as Fusobacterium, Dialister, Faecalibacterium and Tolumonas and a decrease in Bacteroides and Roseburia spp. [76,77,78,79]. The GM may influence the risk of osteoporosis through effects on endogenous estrogens. The ratio of estrogen metabolites to parent estrogen was associated with relative abundances of a number of taxa in the class Clostridia (including the order Clostridiales and the family Ruminococcaceae), whereas the genus Bacteroides was inversely associated with this ratio [80]. Another study found [81] that a number of unique bacterial species is directly associated with systemic estrogens; thus, the GM could affect the risk for estrogen-related conditions in older adults.
The gut microbiota is able to affect bone health by modulating inflammatory actions caused by a lack of sex steroids. This deficiency can be observed in postmenopausal osteoporosis, where is mainly caused by ovarian function cessation, while a decline in estrogen levels results in increased bone resorption. In general, the GM regulates estrogens through secretion of β-glucuronidase, which deconjugates estrogens into their active forms. When this process is disrupted by dysbiosis, the decrease in deconjugation results in a reduction of circulating estrogens [57]. In ovariectomized (OVX) mice, elevated gut permeability; expanded Th17 cells and upregulated levels of osteoclastogenic cytokines (such as TNFα, RANKL and IL-17) were reported. On the contrary, sex steroid deficiencies did not affect the osteoclastogenic cytokines in GF mice [58].
Interestingly, there is also a close relationship between GM and vitamin D3. A recent study [82] has shown that vitamin D3 supplementation correlated with the alteration of gut microbiota towards a decrease in Firmicutes to Bacteroidetes ratio in adults with vitamin D insufficiency/deficiency.
Recently, there has been a great interest in probiotic supplementation and its positive effects on health. In general, probiotics are living microorganisms (species such as Lactobacilli, Bifidobacteria, Enterococcus and Bacillus subtilis) that are intended to have health benefits to the host [83]. Li and colleagues [84] focused on the effect of probiotic supplementation on bone loss in OVX mice. The treatment of OVX mice with Lactobacillus rhamnosus GG or commercially available probiotic supplement VSL#3 (containing eight strains of bacteria—Bifidobacterium breve, B. longum, B. infantis, Lactobacillus acidophilus, L. plantarum, L. paracasei, L. bulgaricus and Streptococcus thermophilus) reduced the gut permeability, intestinal and bone marrow inflammation and protected against bone loss [84]. Another study focused on the impact of probiotic treatment, using a mix of three Lactobacillus strains (L. paracasei DSM 13434, L. plantarum DSM 15312 and L. plantarum DSM 15313) in early postmenopausal women [85]. In general, Lactobacillus treatment reduced the lumbar spine BMD (LS-BMD) loss compared to the placebo group. Britton et al. [86] revealed a beneficial effect of Lactobacillus reuteri ATCC PTA 6475 treatment in OVX mice. In these mice, a significant decrease of osteoclastogenesis (via the suppression of bone marrow CD4+ T lymphocytes), as well as bone resorption markers and activators (tartrate resistant acid phosphatase 5 (TRAcP5) and RANKL), were recorded. Moreover, there is also a study investigating the effect of the probiotic Bacillus subtilis C-3102 on BMD in healthy postmenopausal women [77]. The data suggest that 12 weeks of the probiotic supplementation resulted in a significant increase in relative abundance of the genus Bifidobacterium and in a significant decrease in relative abundance of the genus Fusobacterium. Additionally, there was an increase in total hip BMD in the L. reuteri-treated group, which correlates with a decrease of bone resorption markers (urinary type I collagen crosslinked N-telopeptide and TRAcP5b).

{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary table of various GM alterations associated with bone-related diseases and the effects of probiotic therapy used in animal and human studies.
Table 1.
Summary table of various GM alterations associated with bone-related diseases and the effects of probiotic therapy used in animal and human studies.
| Disease | GM Alterations Associated with the Disease | Probiotic Therapy Used | Effects of Probiotic Therapy in Studies | References |
|---|---|---|---|---|
| osteoporosis | HS: increase in Fusobacterium, Dialister, Faecalibacterium, Tolumonas, Bacteroides, Parabacteroides, Adlercreutzia, Lactobacillus; decrease in Roseburia, Clostridia, Methanobrevibacter, Romboutsia, Turicibacter, Lachnospira | AS: Lactobacillus rhamnosus, L. acidophilus, L. plantarum, L. paracasei, L. bulgaricus, L. reuteri, Bifidobacterium breve, B. longum, B. infantis, Streptococcus thermophilus HS: Lactobacillus paracasei, L. plantarum, Bacillus subtilis | AS: reduced gut permeability, intestinal and bone marrow inflammation; decrease of osteoclastogenesis and bone resorption markers HS: reduced lumbar spine BMD loss; increased total hip BMD; increased Bifidobacterium | [76,87,88,89] |
| osteoarthritis | HS: increase in Clostidium, Gemmiger, Klebsiella, Akkermansia, Lactobacillus, Betaproteobacteria, Streptococcus, Bilophila, Desulfovibrio decrease in Bifidobacterium, Faecalibacterium, Bacteroides, Prevotella, Alistipes, Clostridium, Parabacteroides, Roseburia | AS: Clostridium butyricum, Lactobacillus acidophilus, L. casei, L. fermentum, L. paracasei, Streptococcus thermophilus, Bifidobacterium longum, B. bifidum, B. breve, L. rhamnosus, L. plantarum, L. helveticus, L. salivarius | AS: preserved knee cartilage and synovial membrane; reduced fibrous tissue; decreased cartilage damage; lowered inflammatory and bone metabolism markers in serum; increased levels of IFN-γ and glycosaminoglycans; alleviated pain; increased Bifidobacterium and Roseburia; decreased Closteridium leptum, Akkemansia muciniphila | [90,91,92,93,94,95,96,97,98,99,100] |
| rheumatoid arthritis | HS: increase in Prevotella, Clostridium, Ruminococcus, Lactobacillus, Collinsella, Eggerthella decrease in Bacteroides, Haemophillus, Firmicutes, Faecalibacterium, | AS: Lactobacillus casei HS: L. acidophilus, L. casei, B. bifidum | AS: inhibited joint swelling, lowered arthritis scores, prevented bone destruction; downregulated pro-inflammatory cytokines; increased L. acidophilus HS: improved Disease Activity Score, decreased serum insulin | [101,102,103,104,105,106] |
| type 1 diabetes mellitus | HS: increase in Bacteroides, Veillonella, Alistipes, Klebsiella, Enterococcus, Clostridium, Staphylococcus, Streptococcus, Sarcina, Corynebacterium, Barnesiella, Haemophilus, Ruminococcus, Blautia decrease in Bifidobacterium, Lactobacillus, Escherichia, Prevotella, Akkermansia, Eubacterium, Roseburia, Faecalibacterium, Lachnospira | AS: Lactobacillus brevis, L. reuteri, L. lactis, L. kefiranofaciens, L. kefiri, Bifidobacterium, Streptococcus thermophilus HS: L. paracasei, L. plantarum, L. acidophilus, L. delbrueckii, B. longum, B. infantis, B. breve, Streptococcus thermophiles | AS: reduced blood glucose levels via gamma-aminobutyric acid; elevated innate response; reduced intestinal inflammation; suppressed osteoblast Wnt pathway; stimulated secretion of anti-inflammatory cytokines HS: decrease in glycated hemoglobin, decline in total and bolus insulin; ameliorated conditions of metabolic syndrome | [107,108,109,110,111,112] |
| type 2 diabetes mellitus | HS: increase in Ruminococcus, Fusobacterium, Blautia, Bacteroides, Clostridium, Eggerthella, Escherichia decrease in Bifidobacterium, Bacteroides, Faecalibacterium, Akkermansia, Roseburia, Firmicutes | AS: Lactobacillus rhamnosus, L. casei, L. plantarum, L. acidophilus, L. paracasei, Bifidobacterium bifidum HS: L. casei, L. reuteri, L. acidophilus, Bifidobacterium lactis | AS: decreased fasting and postprandial blood glucose; improved insulin sensitivity and reduced lipid accumulation by stimulating adiponectin secretion; reduced plasma lipids and pro-inflammatory cytokines HS: decreased fasting and postprandial blood glucose; reduced plasma lipids and pro-inflammatory cytokines | [109,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129] |
| obesity | HS: increase in Firmicutes, Acidaminococcus, Anaerococcus, Catenibacterium, Dialister, Dorea, Escherichia-Shigella, Eubacterium, Fusobacterium, Megasphera, Prevotella, Roseburia, Streptococcus, Sutterella, Staphylococcus, Clostridium, Lactobacillus decrease in Bacteroidetes, Bifidobacterium, Eggerthella | AS: Lactobacillus gasseri, L. plantarum, L. curvatus, L. reuteri, L. acidophilus, L. paracasei, L. bulgaricus, Bifidobacterium breve, B. pseudocatenulatum, B. longum, B. infantis, Streptococcus thermophiles HS: L. acidophilus, L. rhamnosus, L. gasseri, L. salivarius, L. casei, L. amylovorus L. fermentum, L. plantarum, Enterococcus faecium, Streptococcus thermophilus, Bifidobacterium lactis | AS: decreased weight, body fat and leptin; decreased insulin resistance, triglyceridemia, cholesterolemia; increased trabecular bone volume and bone mechanical strength; improved osteoblast mineralization HS: decreased LDL cholesterol, body weight, BMI, visceral and subcutaneous fat, waist and hip circumference; increased plasma adiponectin | [130,131,132,133,134,135,136,137] |
GM—gut microbiome; AS—animal studies; HS—human studies; BMD—bone mineral density; IFN—γ–interferon γ; LDL—low-density lipoprotein; BMI—body mass index.
5.2. Inflammatory Bone-Related Diseases
Inflammatory diseases involving bones and joints are very common in the world, as 14 million people worldwide suffer from rheumatoid arthritis. Primarily, joint diseases are characterized by a systemic osteoporosis and increased fracture rates [138]. Generally, these disorders are linked by the presence of an inflammatory process targeting the joints with adverse effects on the structure and function [139]. From this group, osteoarthritis and rheumatoid arthritis are described in more detail.
5.2.1. Osteoarthritis
Osteoarthritis (OA) can be defined by joint symptoms, structural pathology or both [140]. It is characterized by degeneration of articular cartilage, leading to joint pain and disability [141]. OA is a multifactorial disorder; thus, both the systemic and local factors (e.g., age, sex, ethnic characteristics, BMD, genetics, obesity, joint injury and muscle weakness) must be taken into account [142]. The treatment of OA falls into four categories—nonpharmacological (exercise and modification of lifestyle); pharmacological (e.g., acetaminophen, non-steroidal anti-inflammatory drugs and intra-articular injections of corticosteroids or hyaluronic acid); complementary and alternative (e.g., glucosamine and chondroitin supplements and balneotherapy) and surgical ones. The treatment should begin with the least invasive therapy, and all patients should receive their treatment from the first two categories [99].
Several studies examined the differences in GM compositions between patients with OA and healthy individuals (Table 1). Chen et al. [93] revealed a decrease of Bifidobacterium longum and Faecalibacterium prausnitzii and an increase of Clostidium spp. in OA patients. This study correlated with the results of Bonato et al. [94], who noted that the Clostridium genus was upregulated in various studies. Interestingly, Wang and colleagues [95] analyzed stool samples from overweight OA patients and overweight healthy individuals by 16S rRNA gene sequencing. Their findings showed that the relative abundance of nine genera differed between the groups, as the genera of Gemmiger, Klebsiella, Akkermansia and Lactobacillus were enriched in OA patients, while Bacteroides, Prevotella, Alistipes, Clostridium XI and Parabacteroides were enriched in the control group.
There are numerous researchers suggesting the positive effects of probiotic supplementation in rats with OA. Sim et al. [96] found that Clostridium butyricum therapy prevented OA symptoms, preserved knee cartilage and synovial membrane and significantly decreased the amount of fibrous tissue. This supplementation also significantly lowered the serum levels of inflammatory and bone metabolism markers (cyclooxygenase-2, leukotriene B4, cartilage oligomeric matrix protein and IL-6) and increased the levels of IFN-γ and glycosaminoglycans. Another study [97] revealed that the combination with a probiotic complex (consisting of Lactobacillus acidophilus, L. casei, L. fermentum, L. paracasei, Streptococcus thermophilus, Bifidobacterium longum, B. bifidum, B. breve, L. rhamnosus, L. plantarum, L. helveticus and L. salivarius); rosavin (natural product found in Cinnamomum iners, Cinnamomum aromaticum and others) and zinc ameliorated pain by preventing cartilage damage in rats with OA. Additionally, the expression of proinflammatory cytokines was decreased. Lee and colleagues [98] showed that Lactobacillus acidophilus supplementation alleviated OA-associated pain and delayed the progression of this disease by inhibiting the levels of proinflammatory cytokines in the joints. Interestingly, the oral administration of L. casei together with type II collagen and glucosamine more effectively reduced pain, cartilage destruction and lymphocyte infiltration than the individual treatment of glucosamine or L. casei. Additionally, the expression of pro-inflammatory cytokines (such as IL-1β, IL-2, IL-6, IL-12, IL17, TNF-α and IFN-γ) and matrix metalloproteinases was decreased, while anti-inflammatory cytokines (IL-4 and IL-10) were increased [100]. According to a preliminary clinical study [143], Streptococcus thermophilus (TCI633), a bacterium able to produce hyaluronate in GIT, could improve the degeneration of knee osteoarthritis in humans.
5.2.2. Rheumatoid Arthritis
Rheumatoid arthritis (RA) is a systemic inflammatory autoimmune disease characterized by synovial inflammation and hyperplasia; auto-antibody production and systemic features (cardiovascular, pulmonary and psychological disorders) [144]. It is also responsible for joints destructions associated with bone complications that include periarticular bone loss, bone erosions and systemic osteoporosis [139]. The most important factors in the management of RA are the early diagnosis, prompt disease-modifying anti-rheumatic drugs (DMARDs) therapy and monitoring of patients to increase the likelihood of remission. The current available drug therapy includes non-steroidal anti-inflammatory drugs (NSAIDs), glucocorticoids and DMARDs (such as methotrexate, Janus kinase inhibitors, TNF inhibitors, IL-6 inhibitors and B-cell depleting drugs). Non-pharmacological treatments such as physical therapy; lifestyle changes (e.g., smoking cessation, attaining ideal body weight and exercise) and surgical management belong to important treatment resources as well [102].
Scher et al. [103] identified the increase in Prevotella copri accompanied by a reduction in Bacteroides and a loss of beneficial microbes in untreated patients with RA. Zhang et al. [104] detected dysbiosis in the gut in these patients; however, it was partially resolved after the treatment. In the oral samples, there was an increase in Porphyromonas gingivalis, while the gut microbiota had elevated numbers of Clostridium spp., Ruminococcus spp. and Lactobacillus spp. and lower numbers of Haemophillus and Firmicutes. The information is presented in Table 1.
There is increasing evidence that probiotic supplementation could reverse a microbial disorder in patients with RA. The L. casei (ATCC334) treatment of adjuvant-induced arthritis (AIA) in rats inhibited joint swelling, lowered the arthritis scores and prevented bone destruction, while the relative abundance of Lactobacillus strains, which is decreased in AIA rats, was restored to normal, and the level of L. acidophilus was even increased. In addition, L. casei supplementation caused a downregulation of proinflammatory cytokines [105]. Another study also showed that L. casei supplementation reduced the levels of proinflammatory cytokines (TNF-α, IL-6 and IL-12) and elevated the level of regulatory cytokine IL-10 in RA individuals [106]. Zamani and colleagues [145] studied the effects of probiotic supplementation (L. acidophilus, L. casei and B. bifidum) on the clinical and metabolic status of RA patients. This supplementation improved the Disease Activity Score in 28 joints (scoring system to evaluate disease activity and treatment response in RA). Additionally, a significant decrease in the serum insulin levels was observed.
5.3. Diabetes Mellitus
Diabetes mellitus (DM) represents a worldwide public health issue, prevailing at approximately 450 million adults [146]. It is a chronic metabolic disease characterized by impaired insulin production/secretion or the action of insulin when an organism is unable to use insulin effectively. The vast majority of DM cases fall into two broad etiopathogenetic categories—type 1 and type 2 DM [147]. Type 1 DM (T1DM) is an autoimmune disease characterized by hyperglycemia, where elevated blood glucose levels are caused by insulin deficiency as a consequence of pancreatic β-cells loss. The aim in the management of T1DM is to promote healthy living and glycemic control. Pharmacological treatment is mainly focused on insulin therapy (insulin and insulin analogues); however, it also includes nutritional awareness to reduce the risk of cardiovascular disease and obesity and exercise to improve insulin sensitivity, lipid metabolism and blood pressure [148]. Type 2 DM (T2DM) is a multifactorial disease characterized by the dysregulation of carbohydrate, lipid and protein metabolism due to impaired insulin secretion by pancreatic β-cells, insulin resistance in skeletal muscle, liver and adipose tissue or both [149]. T2DM management includes lifestyle intervention (e.g., diet, exercise, moderate alcohol consumption and reduced sodium intake), along with pharmacological treatment, such as insulin sensitizers, insulin secretagogues, incretin-based therapies, sodium–glucose cotransporter 2 inhibitors and α-glucosidase inhibitors [150,151]. If antidiabetic treatment fails to normalize the levels of glycated hemoglobin (Hb1Ac), patients may be treated with insulin; however, high doses are often required [149].
In general, both T1DM and T2DM have a harmful impact on the bone quality, which manifests itself in a higher fracture risk [152]. Interestingly, low BMD is observed in T1DM, while BMD may not be affected in patients with T2DM. One study suggested [153] that T2DM is associated with a fracture risk, despite higher BMD levels and thicker femoral cortices in narrower bones. Another study [154] revealed that there is no relationship between T2DM and low BMD. Additionally, it was shown that even BMD was not significantly affected, and the incidence of osteoporosis was higher in T2DM patients [155]. Interestingly, Napoli and colleagues [156] demonstrated that the bone turnover markers (C-terminal telopeptide of type I collagen (CTX), osteocalcin and procollagen type 1 N-terminal propeptide (P1NP)) did not predict the incident fracture risk in T2DM patients. On the other hand, a study conducted by Starup-Linde et al. [157] showed that patients with T2DM displayed significantly lower levels of CTX and P1NP.
The pathophysiology of low BMD in T1DM can be explained by various effects such as reduced insulin signaling [114], growth hormone (GH) disorder and decreased IGF-1 [115], calcium and vitamin D [116] levels or elevated levels of proinflammatory cytokines [117].
It has been reported that the GM is altered in patients with DM (Table 1), as a relative abundance of several bacterial taxa was observed [109]. The most common alterations in patients with T1DM include bacterial species such as Bacteroides spp., Streptococcus spp., Clostridium spp., Bifidobacterium spp., Prevotella spp., Staphylococcus spp., Blautia spp., Faecalibacterium spp., Roseburia spp. and Lactobacillus spp. [107]. Pellegrini and colleagues [118] discovered an association between inflammatory markers and specific bacterial taxa. They found an increased gene expression of the cytokines CCL13, CCL19 and CCL22; chemokine receptor CCR2; cyclooxygenase 2; IL-4 receptor; CD68; pentraxin-3; TNF-α and vascular endothelial growth factor A. Their immunohistochemical analysis (performed on biopsy samples of the duodenal mucosa) also confirmed T1DM—a specific inflammatory condition. In addition, the duodenal mucosal microbiome included increased Firmicutes and a Firmicutes/Bacteroidetes ratio and a decrease in Proteobacteria and Bacteroidetes. Another study confirmed that T1DM is associated with a reduced microbiota diversity (with a significant increase in the relative abundance of Bacteroides, Ruminococcus, Veillonella, Blautia and Streptococcus genera and a lower relative abundance of Bifidobacterium, Roseburia, Faecalibacterium and Lachnospira). Many studies, focusing on the relationship between GM and T2DM, have provided diverse findings. Several researchers [119,120,121,122,123,124,125,126] indicated that the genera of Bifidobacterium, Bacteroides, Faecalibacterium, Akkermansia and Roseburia were negatively associated with T2DM, and the genera of Ruminococcus, Fusobacterium and Blautia were positively associated. Interestingly, the metagenome-wide association study based on the deep next-generation shotgun sequencing of stool sample DNA by Qin et al. [110] showed that patients with T2DM had only moderate bacterial dysbiosis, but there was a decline in butyrate-producing bacteria. Moreover, the findings of this study suggested a “functional dysbiosis” rather than the existence of a specific microbial species that is directly related to the pathophysiology of T2DM.
There is a growing interest in probiotic supplementation for the management and treatment of DM. Kumar et al. [111] investigated the effect of probiotic therapy (multi-strain probiotic Visbiome® containing L. paracasei, L. plantarum, L. acidophilus, L. delbrueckii, B. longum, B. infantis, B. breve and Streptococcus thermophilus) in children with T1DM. They found a significant decrease in HbA1c (glycated hemoglobin) and decline in the total and bolus insulin dose. Other studies [112,127] examined the impacts of Lactobacillus rhamnosus GG and Bifidobacterium lactis Bb12 on β-cell function in children with newly diagnosed T1DM. However, no significant effect in maintaining the residual pancreatic β-cell function was found. The beneficial impacts of probiotic supplementation in patients with T2DM could be mediated by improving the gut integrity and peripheral insulin sensitivity, decreasing the systemic levels of LPS (lipopolysaccharide) and increasing the incretins [128]. Interestingly, L. rhamnosus GG improved the insulin sensitivity and reduced the lipid accumulation by stimulating adiponectin secretion in an animal model [129]. Kobyliak and colleagues [158] revealed that probiotic therapy using the multi-probiotic “Symbiter” (containing 14 probiotic bacteria genera) modestly improved the insulin resistance in individuals with T2DM.
5.4. Obesity
Overweight and obesity are identified as abnormal or excessive fat accumulation that can lead to impaired health. Nowadays, the prevalence of obesity is on the rise, as well as obesity-related diseases [159]. Excessive amounts of fat are associated with cardiovascular disease, T2DM, hypertension, stroke, dyslipidemia and some types of cancers and might be a risk factor for osteoporosis and fragility fractures [160,161]. Adipose tissue is not just a passive reservoir of fat, but it is now considered as an active endocrine organ involved in the modulation of energy homeostasis. It secretes numerous cytokines, such as IL-6 and TNF-α and fat-derived mediators, including resistin, leptin and adiponectin, which are also involved in bone metabolism. In addition, adipose tissue is also a source of aromatase (which catalyzes the synthesis of estrogens); hence, it is an important source of estrogen in postmenopausal women [161]. The interventions for weight control and the treatment of obesity include dietary programs; medical nutrition therapy; exercise/physical activity; behavior therapy; pharmacotherapy (e.g., orlistat, phentermine/topiramate, lorcaserin, naltrexone/bupropion and liraglutide); bariatric surgery or a combination of these treatments [133].
Pinart and colleagues [134] in their review focused on the composition of GM in obese and non-obese adults. Interestingly, in 9 out of 22 studies, there was a significantly lower alpha diversity in obese adults; however, 7 studies did not reveal a significant difference. In obese adults, significantly more Firmicutes and fewer Bacteroidetes were observed. Additionally, lower relative abundances of Bifidobacterium and Eggerthella and higher abundances of Acidaminococcus, Anaerococcus, Catenibacterium, Dialister, Dorea, Escherichia-Shigella, Eubacterium, Fusobacterium, Megasphera, Prevotella, Roseburia, Streptococcus and Sutterella were recorded in obese adults. In some studies, the genera Staphylococcus and Clostridium, which belong to the phylum Firmicutes, have been shown to be positively associated with obesity. The phylum Firmicutes contains many butyrate-producing species, and the increase in butyrate and acetate synthesis may contribute to an increase in energy harvest in obese subjects [130]. In addition, Bifidobacterium has been shown to have an inverse relationship with obesity in pregnant women and children, possibly due to the deconjugation of bile acids, which may reduce fat absorption [135,136]. Table 1 provides this information.
Manipulation of the GM can be a novel approach in preventing pathological bone loss in obese patients. Behera et al. [137] examined the effect of probiotic supplementation (previously mentioned commercially available probiotics VSL#3®) on mitochondrial biogenesis and bone homeostasis in obese mice fed a high-fat diet. The results showed that probiotic therapy increased the trabecular bone volume and bone mechanical strength. The treatment also prevented gut inflammation and improved osteoblast mineralization, as there was an increase of mitochondrial transcription factor A expression in osteoblasts by promoting Kdm6b/Jmjd3 histone demethylase, which inhibits H3K27me3 epigenetic methylation at the Tfam promoter. Another study reported a loss of beneficial Bifidobacteria in obese mice, while the proinflammatory species were increased [162]. Inflammation can further cause a macrophage migration to the synovium accelerating knee OA. Additionally, oligofructose (nondigestible prebiotic fiber) supplementation restored the key microbes (particularly Bifidobacterium pseudolongum), reduced the colonic macrophage cell signature and decreased the systemic and knee joint inflammation [162].
5.5. Bone Cancer
Primary bone cancers are rare, and they represent a heterogeneous group of malignancies, including osteosarcoma, chondrosarcoma and Ewing sarcoma, as the most common forms of bone cancer. The pathogenesis and origin of most bone tumors is unclear [163]. However, it has been hypothesized that osteosarcoma originate from malignant primitive mesenchymal cells that differentiate into osteoblasts, and Ewing sarcoma could be derived from undifferentiated, primitive neuroectodermal or neural crest cells. Chondrosarcoma is a cartilage-producing bone tumor, occurring mostly in the pelvis, proximal long bones, ribs, scapula and vertebrae, and its malignant cell origin are chondrocytes [164]. In addition, bone metastases are a common complication of many types of cancers occurring in patients with breast, prostate, lung, renal or thyroid cancers [165]. The treatment options for bone tumors include chemotherapy (for osteosarcoma–cisplatin, doxorubicin, methotrexate; for Ewing sarcoma–cyclophosphamide, doxorubicin and etoposide); radiation therapy and surgical management [164].
Wenhui and colleagues [166] compared the GM profiles from breast cancer patients with no metastasis (w/oBMs), breast cancer patients with bone metastasis (BMs) and control individuals (Cs). They found that the community diversity was the lowest in patients with BMs. In these patients, a lack of Megamonas and Akkermansia was noted. Streptococcus, Campylobacter and Moraxellaceae showed higher abundances in the w/oBM and BM groups compared to the Cs. However, there are not enough studies focusing on the GM composition in bone cancer. The therapeutic value of probiotic consumption by showing an increase in cancer inhibitors and a decrease in cancer inducers has been revealed [167], but very few or no reports exist on the treatment of bone tumors with probiotics.
6. The Role of Gut Microbiome in Drug Response
The response to drugs can vary widely between individuals, and the role of GM in this variability is increasingly appreciated. A growing number of reports in the literature increase the level of evidence that commonly prescribed drugs that significantly affect bone homeostasis may interact with other physiological systems, such as the GM, which is essential for optimal bone function. Given the growing body of evidence linking healthy GM to bone homeostasis, the therapeutic benefits of commonly prescribed drugs and supplements, many of which are known to alter the GM [168,169]. The mechanisms of action may include drug disposition via microbial metabolism, interference by microbial metabolites or the modification of host enzymes. The current knowledge has been obtained from both animal and human studies. Unfortunately, the effect on the gut microbiome is unknown for several drugs, including bisphosphonates, approved selective estrogen receptor modulators (SERMs, e.g., raloxifene) and approved selective estrogen receptor downregulators (SERDs, e.g., fulvestrant). Selected pharmacological drugs for the treatment of bone-related diseases and also their impacts on GM are summarized in Table 2.
7. Conclusions
The GM plays a key role in numerous physiological functions. It can also influence bone homeostasis via several mechanisms, (e.g., immune and endocrine functions, host metabolism and the gut–brain axis). T- and B-lymphocytes can regulate bone remodeling through the expression of cytokines, as well as RANKL and OPG. SCFAs are able to reduce OC formation and resorption activity. The components of the endocrine system control a balance between bone formation and bone loss, and the maintenance of calcium and phosphate homeostasis. Specific molecules, such as brain-derived serotonin, can positively regulate bone development.
During dysbiosis, GM is consistent with the development of various chronic diseases that can also adversely affect the bone quality and bone health. Significant changes in the GM composition in osteoporosis, osteoarthritis, rheumatoid arthritis, diabetes, obesity and bone cancer were highlighted in this review and summarized in Table 1. GM manipulation may become a future adjuvant therapy in the prevention of many chronic diseases. Therefore, the favorable impacts of probiotic supplementation in each of the above-mentioned diseases are reported in Table 1, but further studies are needed to clearly clarify its effectiveness. Recent evidence suggests that GM is responsible for the direct and indirect effects on drug efficacy. For this reason, various GM changes and interactions associated with the treatment of bone-related diseases are listed in Table 2.
Author Contributions
Conceptualization, R.O. and M.M.; methodology, N.Z., R.O. and M.M.; formal analysis, V.M. and V.K.; writing—original draft preparation, N.Z., R.O. and M.M.; writing—review and editing, R.O. and M.M.; visualization, N.Z. and R.O.; supervision, R.O. and M.M. and project administration, R.O. and M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education, Science, Research and Sport of the Slovak Republic, grant numbers VEGA 1/0444/20 and VEGA 1/0416/22.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rastelli, M.; Cani, P.D.; Knauf, C. The Gut Microbiome Influences Host Endocrine Functions. Endocr. Rev. 2019, 40, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, V.; Salvatore, F. The role of the gut microbiome in the healthy adult status. Clin. Chim. Acta 2015, 451, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Awany, D.; Allali, I.; Dalvie, S.; Hemmings, S.; Mwaikono, K.S.; Thomford, N.E.; Gomez, A.; Mulder, N.; Chimusa, E.R. Host and Microbiome Genome-Wide Association Studies: Current State and Challenges. Front. Genet. 2019, 9, 637. [Google Scholar] [CrossRef]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized Metabolites from the Microbiome in Health and Disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef]
- Hernandez, C.J.; Guss, J.D.; Luna, M.; Goldring, S.R. Links Between the Microbiome and Bone. J. Bone Miner. Res. 2016, 31, 1638–1646. [Google Scholar] [CrossRef]
- Belizário, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef]
- Claus, S.P.; Guillou, H.; Ellero-Simatos, S. The gut microbiota: A major player in the toxicity of environmental pollutants? Npj Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut Microbiota: The Neglected Endocrine Organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Sapra, L.; Tiwari, A.; Mishra, P.K.; Sharma, S.; Srivastava, R.K. “Osteomicrobiology”: The Nexus Between Bone and Bugs. Front. Microbiol. 2022, 12, 812466. [Google Scholar] [CrossRef] [PubMed]
- Villa, C.R.; Ward, W.E.; Comelli, E.M. Gut microbiota-bone axis. Crit. Rev. Food Sci. Nutr. 2015, 57, 1664–1672. [Google Scholar] [CrossRef]
- Gritz, E.C.; Ebhandari, V. The Human Neonatal Gut Microbiome: A Brief Review. Front. Pediatr. 2015, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Hollister, E.B.; Riehle, K.; Luna, R.A.; Weidler, E.M.; Rubio-Gonzales, M.; Mistretta, T.-A.; Raza, S.; Doddapaneni, H.V.; Metcalf, G.A.; Muzny, D.M.; et al. Structure and function of the healthy pre-adolescent pediatric gut microbiome. Microbiome 2015, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.-F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Agans, R.; Rigsbee, L.; Kenche, H.; Michail, S.; Khamis, H.J.; Paliy, O. Distal gut microbiota of adolescent children is different from that of adults. FEMS Microbiol. Ecol. 2011, 77, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Kulka, T.; Cheng, J.; Ringel, Y.; Salojärvi, J.; Carroll, I.; Palva, A.; De Vos, W.M.; Satokari, R. Intestinal Microbiota in Healthy U.S. Young Children and Adults—A High Throughput Microarray Analysis. PLoS ONE 2013, 8, e64315. [Google Scholar] [CrossRef]
- McBurney, M.I.; Davis, C.; Fraser, C.M.; Schneeman, B.O.; Huttenhower, C.; Verbeke, K.; Walter, J.; Latulippe, M.E. Establishing What Constitutes a Healthy Human Gut Microbiome: State of the Science, Regulatory Considerations, and Future Directions. J. Nutr. 2019, 149, 1882–1895. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Matenchuk, B.A.; Mandhane, P.J.; Kozyrskyj, A.L. Sleep, circadian rhythm, and gut microbiota. Sleep Med. Rev. 2020, 53, 101340. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Kim, S.; Jazwinski, S.M. The Gut Microbiota and Healthy Aging: A Mini-Review. Gerontology 2018, 64, 513–520. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Lankelma, J.M.; Nieuwdorp, M.; De Vos, W.M.; Wiersinga, W.J. The gut microbiota in internal medicine: Implications for health and disease. Neth. J. Med. 2015, 73, 61–68. [Google Scholar]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Cox, L.M.; Weiner, H.L. Microbiota Signaling Pathways that Influence Neurologic Disease. Neurotherapeutics 2018, 15, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Reid, I.R. (Ed.) Metabolic Bone Disease; Baillière’s clinical endocrinology and metabolism; Baillière Tindall: London, UK, 1997; ISBN 978-0-7020-2340-8. [Google Scholar]
- Safadi, F.F.; Barbe, M.F.; Abdelmagid, S.M.; Rico, M.C.; Aswad, R.A.; Litvin, J.; Popoff, S.N. Bone Structure, Development and Bone Biology. In Bone Pathology; Khurana, J.S., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 1–50. [Google Scholar] [CrossRef]
- Cui, Y.; Li, Z.; Guo, Y.; Qi, X.; Yang, Y.; Jia, X.; Li, R.; Shi, J.; Gao, W.; Ren, Z.; et al. Bioinspired Nanovesicles Convert the Skeletal Endothelium-Associated Secretory Phenotype to Treat Osteoporosis. ACS Nano 2022, 16, 11076–11091. [Google Scholar] [CrossRef]
- Janeway, C. (Ed.) Immunobiology: The Immune System in Health and Disease, 5th ed.; [Animated CD-ROM Inside]; Garland Publ.: New York, NY, USA, 2001; ISBN 978-0-8153-3642-6. [Google Scholar]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef] [Green Version]
- Tsukasaki, M.; Takayanagi, H. Osteoimmunology: Evolving concepts in bone–immune interactions in health and disease. Nat. Rev. Immunol. 2019, 19, 626–642. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yu, X.-J.; Yu, L.-L.; Tian, F.-W.; Zhao, J.-X.; Zhang, H.; Chen, W.; Zhai, Q.-X. The influence of gut microbiome on bone health and related dietary strategies against bone dysfunctions. Food Res. Int. 2021, 144, 110331. [Google Scholar] [CrossRef] [PubMed]
- Titanji, K. Beyond Antibodies: B Cells and the OPG/RANK-RANKL Pathway in Health, Non-HIV Disease and HIV-Induced Bone Loss. Front. Immunol. 2017, 8, 1851. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.L.; Rios-Arce, N.D.; Schepper, J.D.; Parameswaran, N.; McCabe, L.R. The Potential of Probiotics as a Therapy for Osteoporosis. Microbiol. Spectr. 2017, 5, 213–233. [Google Scholar] [CrossRef] [PubMed]
- Mori, G.; D’Amelio, P.; Faccio, R.; Brunetti, G. The Interplay between the Bone and the Immune System. Clin. Dev. Immunol. 2013, 2013, 1–16. [Google Scholar] [CrossRef]
- Ohlsson, C.; Sjögren, K. Effects of the gut microbiota on bone mass. Trends Endocrinol. Metab. 2015, 26, 69–74. [Google Scholar] [CrossRef]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef]
- Pacifici, R. T cells, osteoblasts, and osteocytes: Interacting lineages key for the bone anabolic and catabolic activities of parathyroid hormone. Ann. N. Y. Acad. Sci. 2015, 1364, 11–24. [Google Scholar] [CrossRef]
- MacPherson, A.J.; Martinic, M.; Harris, N.L. The functions of mucosal T cells in containing the indigenous commensal flora of the intestine. Experientia 2002, 59, 2088–2096. [Google Scholar] [CrossRef]
- Ohlsson, C.; Nigro, G.; Boneca, I.G.; Bäckhed, F.; Sansonetti, P.; Sjögren, K. Regulation of bone mass by the gut microbiota is dependent on NOD1 and NOD2 signaling. Cell. Immunol. 2017, 317, 55–58. [Google Scholar] [CrossRef]
- Moreira, L.O.; Zamboni, D.S. NOD1 and NOD2 Signaling in Infection and Inflammation. Front. Immunol. 2012, 3, 328. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.; Baron, B. The Role of Toll-Like Receptors in Autoimmune Diseases through Failure of the Self-Recognition Mechanism. Int. J. Inflamm. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Schwarzer, M.; Makki, K.; Storelli, G.; Machuca-Gayet, I.; Srutkova, D.; Hermanova, P.; Martino, M.E.; Balmand, S.; Hudcovic, T.; Heddi, A.; et al. Lactobacillus plantarum strain maintains growth of infant mice during chronic undernutrition. Science 2016, 351, 854–857. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, A.; Magrath, W.; Pugliese, B.; Stocker, N.; Westermann, P.; Heider, A.; Gehweiler, D.; Zeiter, S.; Claesson, M.J.; Richards, R.G.; et al. Butyrate Inhibits Osteoclast Activity In Vitro and Regulates Systemic Inflammation and Bone Healing in a Murine Osteotomy Model Compared to Antibiotic-Treated Mice. Mediat. Inflamm. 2021, 2021, 8817421. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 376. [Google Scholar] [CrossRef]
- Oton-Gonzalez, L.; Mazziotta, C.; Iaquinta, M.R.; Mazzoni, E.; Nocini, R.; Trevisiol, L.; D’Agostino, A.; Tognon, M.; Rotondo, J.C.; Martini, F. Genetics and Epigenetics of Bone Remodeling and Metabolic Bone Diseases. Int. J. Mol. Sci. 2022, 23, 1500. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Dhawan, P.; Porta, A.; Mady, L.J.; Seth, T. Vitamin D and intestinal calcium absorption. Mol. Cell. Endocrinol. 2011, 347, 25–29. [Google Scholar] [CrossRef]
- Mineo, H.; Hara, H.; Tomita, F. Short-chain fatty acids enhance diffusional Ca transport in the epithelium of the rat cecum and colon. Life Sci. 2001, 69, 517–526. [Google Scholar] [CrossRef]
- Bryk, G.; Coronel, M.Z.; Pellegrini, G.G.; Mandalunis, P.; Rio, M.E.; De Portela, M.L.P.M.; Zeni, S.N. Effect of a combination GOS/FOS® prebiotic mixture and interaction with calcium intake on mineral absorption and bone parameters in growing rats. Eur. J. Nutr. 2015, 54, 913–923. [Google Scholar] [CrossRef]
- Krupa-Kozak, U.; Markiewicz, L.H.; Lamparski, G.; Juśkiewicz, J. Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner. Nutrients 2017, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Martin, B.R.; Nakatsu, C.H.; A Story, J.; Macdonald-Clarke, C.J.; McCabe, L.D.; McCabe, G.P.; Weaver, C.M. Soluble Corn Fiber Increases Calcium Absorption Associated with Shifts in the Gut Microbiome: A Randomized Dose-Response Trial in Free-Living Pubertal Females. J. Nutr. 2016, 146, 1298–1306. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, E.G.; Schoterman, M.H.C.; Muijs, T. Transgalactooligosaccharides Stimulate Calcium Absorption in Postmenopausal Women. J. Nutr. 2000, 130, 2938–2942. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen–gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Malik, T.A.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex steroid deficiency–associated bone loss is microbiota dependent and prevented by probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef]
- Yamamoto, E.A.; Jørgensen, T.N. Relationships Between Vitamin D, Gut Microbiome, and Systemic Autoimmunity. Front. Immunol. 2020, 10, 13. [Google Scholar] [CrossRef]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D Regulates the Gut Microbiome and Protects Mice from Dextran Sodium Sulfate–Induced Colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef]
- Luthold, R.V.; Fernandes, G.R.; Franco-De-Moraes, A.C.; Folchetti, L.G.; Ferreira, S.R.G. Gut microbiota interactions with the immunomodulatory role of vitamin D in normal individuals. Metabolism 2017, 69, 76–86. [Google Scholar] [CrossRef]
- Greenstein, R.J.; Su, L.; Brown, S.T. Vitamins A & D Inhibit the Growth of Mycobacteria in Radiometric Culture. PLoS ONE 2012, 7, e29631. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2015, 82, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Functions of Osteocalcin in Bone, Pancreas, Testis, and Muscle. Int. J. Mol. Sci. 2020, 21, 7513. [Google Scholar] [CrossRef] [PubMed]
- Wagatsuma, K.; Yamada, S.; Ao, M.; Matsuura, M.; Tsuji, H.; Iida, T.; Miyamoto, K.; Oka, K.; Takahashi, M.; Tanaka, K.; et al. Diversity of Gut Microbiota Affecting Serum Level of Undercarboxylated Osteocalcin in Patients with Crohn’s Disease. Nutrients 2019, 11, 1541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, Y.; Wang, Y.; Ren, X.; Han, J. The impact of the intestinal microbiome on bone health. Intractable Rare Dis. Res. 2018, 7, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Karsenty, G. The two faces of serotonin in bone biology. J. Cell Biol. 2010, 191, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Michalowska, M.; Znorko, B.; Kaminski, T.; Oksztulska-Kolanek, E.; Pawlak, D. New insights into tryptophan and its metabolites in the regulation of bone metabolism. J. Physiol. Pharmacol. 2015, 66, 779–791. [Google Scholar]
- Kepert, I.; Fonseca, J.; Müller, C.; Milger, K.; Hochwind, K.; Kostric, M.; Fedoseeva, M.; Ohnmacht, C.; Dehmel, S.; Nathan, P.; et al. D-tryptophan from probiotic bacteria influences the gut microbiome and allergic airway disease. J. Allergy Clin. Immunol. 2017, 139, 1525–1535. [Google Scholar] [CrossRef]
- Seely, K.D.; Kotelko, C.A.; Douglas, H.; Bealer, B.; Brooks, A.E. The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. Int. J. Mol. Sci. 2021, 22, 9452. [Google Scholar] [CrossRef]
- Martiniakova, M.; Babikova, M.; Omelka, R. Pharmacological agents and natural compounds: Available treatments for osteoporosis. J. Physiol. Pharmacol. 2020, 71, 307–320. [Google Scholar] [CrossRef]
- Cui, Y.; Guo, Y.; Kong, L.; Shi, J.; Liu, P.; Li, R.; Geng, Y.; Gao, W.; Zhang, Z.; Fu, D. A bone-targeted engineered exosome platform delivering siRNA to treat osteoporosis. Bioact. Mater. 2021, 10, 207–221. [Google Scholar] [CrossRef]
- Swanson, C.M.; Kohrt, W.M.; Buxton, O.M.; Everson, C.A.; Wright, K.P.; Orwoll, E.S.; Shea, S.A. The importance of the circadian system & sleep for bone health. Metabolism 2018, 84, 28–43. [Google Scholar] [CrossRef]
- Lu, K.; Dong, S.; Wu, X.; Jin, R.; Chen, H. Probiotics in Cancer. Front. Oncol. 2021, 11, 638148. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Zhang, J.; Reiter, R.J.; Ma, X. Melatonin mediates mucosal immune cells, microbial metabolism, and rhythm crosstalk: A therapeutic target to reduce intestinal inflammation. Med. Res. Rev. 2020, 40, 606–632. [Google Scholar] [CrossRef]
- He, J.; Xu, S.; Zhang, B.; Xiao, C.; Chen, Z.; Si, F.; Fu, J.; Lin, X.; Zheng, G.; Yu, G.; et al. Gut microbiota and metabolite alterations associated with reduced bone mineral density or bone metabolic indexes in postmenopausal osteoporosis. Aging 2020, 12, 8583–8604. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus subtilis C-3102 on bone mineral density in healthy postmenopausal Japanese women: A randomized, placebo-controlled, double-blind clinical trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, J.; Li, X.; Sun, Q.; Qin, P.; Wang, Q. Compositional and functional features of the female premenopausal and postmenopausal gut microbiota. FEBS Lett. 2019, 593, 2655–2664. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xie, Z.; Sun, J.; Huang, S.; Chen, Y.; Li, C.; Sun, X.; Xia, B.; Tian, L.; Guo, C.; et al. Gut Microbiome Reveals Specific Dysbiosis in Primary Osteoporosis. Front. Cell. Infect. Microbiol. 2020, 10, 160. [Google Scholar] [CrossRef]
- Fuhrman, B.; Feigelson, H.S.; Flores, R.; Gail, M.H.; Xu, X.; Ravel, J.; Goedert, J.J. Associations of the Fecal Microbiome With Urinary Estrogens and Estrogen Metabolites in Postmenopausal Women. J. Clin. Endocrinol. Metab. 2014, 99, 4632–4640. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef]
- Charoenngam, N.; Shirvani, A.; Kalajian, T.A.; Song, A.; Holick, M.F. The Effect of Various Doses of Oral Vitamin D3 Supplementation on Gut Microbiota in Healthy Adults: A Randomized, Double-blinded, Dose-response Study. Anticancer. Res. 2020, 40, 551–556. [Google Scholar] [CrossRef]
- Locantore, P.; Del Gatto, V.; Gelli, S.; Paragliola, R.M.; Pontecorvi, A. The Interplay between Immune System and Microbiota in Osteoporosis. Mediat. Inflamm. 2020, 2020, 3686749. [Google Scholar] [CrossRef]
- Li, X.; Wang, N.; Yin, B.; Fang, D.; Jiang, T.; Fang, S.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Effects of Lactobacillus plantarum CCFM0236 on hyperglycaemia and insulin resistance in high-fat and streptozotocin-induced type 2 diabetic mice. J. Appl. Microbiol. 2016, 121, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.-A.; Curiac, D.; Ahrén, I.L.; Hansson, F.; Niskanen, T.M.; Sjögren, K.; Ohlsson, C. Probiotic treatment using a mix of three Lactobacillus strains for lumbar spine bone loss in postmenopausal women: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet Rheumatol. 2019, 1, e154–e162. [Google Scholar] [CrossRef]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri Treatment Prevents Bone Loss in a Menopausal Ovariectomized Mouse Model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- A Rettedal, E.; Ilesanmi-Oyelere, B.L.; Roy, N.C.; Coad, J.; Kruger, M.C. The Gut Microbiome Is Altered in Postmenopausal Women With Osteoporosis and Osteopenia. JBMR Plus 2021, 5, 10452. [Google Scholar] [CrossRef]
- Li, S.; Mao, Y.; Zhou, F.; Yang, H.; Shi, Q.; Meng, B. Gut microbiome and osteoporosis. Bone Jt. Res. 2020, 9, 524–530. [Google Scholar] [CrossRef]
- Ding, K.; Hua, F.; Ding, W. Gut Microbiome and Osteoporosis. Aging Dis. 2020, 11, 438–447. [Google Scholar] [CrossRef]
- Favazzo, L.J.; Hendesi, H.; Villani, D.A.; Soniwala, S.; Dar, Q.-A.; Schott, E.M.; Gill, S.R.; Zuscik, M. The gut microbiome-joint connection: Implications in osteoarthritis. Curr. Opin. Rheumatol. 2020, 32, 92–101. [Google Scholar] [CrossRef]
- Binvignat, M.; Sokol, H.; Mariotti-Ferrandiz, E.; Berenbaum, F.; Sellam, J. Osteoarthritis and gut microbiome. Jt. Bone Spine 2021, 88, 105203. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, C.; Zhang, Y.; Zhang, W.; Doherty, M.; Yang, T.; Zhai, G.; Obotiba, A.D.; Lyu, H.; Zeng, C.; et al. Association Between Gut Microbiota and Symptomatic Hand Osteoarthritis: Data From the Xiangya Osteoarthritis Study. Arthritis Rheumatol. 2021, 73, 1656–1662. [Google Scholar] [CrossRef]
- Chen, J.; Wang, A.; Wang, Q. Dysbiosis of the gut microbiome is a risk factor for osteoarthritis in older female adults: A case control study. BMC Bioinform. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Bonato, A.; Zenobi-Wong, M.; Barreto, G.; Huang, Z. A systematic review of microbiome composition in osteoarthritis subjects. Osteoarthr. Cartil. 2022, 30, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, H.; Jiang, Q.; Zhu, Y.Z. The gut microbiome as non-invasive biomarkers for identifying overweight people at risk for osteoarthritis. Microb. Pathog. 2021, 157, 104976. [Google Scholar] [CrossRef] [PubMed]
- Sim, B.-Y.; Choi, H.-J.; Kim, M.-G.; Jeong, D.-G.; Lee, D.-G.; Yoon, J.-M.; Kang, D.-J.; Park, S.; Ji, J.-G.; Joo, I.-H.; et al. Effects of ID-CBT5101 in Preventing and Alleviating Osteoarthritis Symptoms in a Monosodium Iodoacetate-Induced Rat Model. J. Microbiol. Biotechnol. 2018, 28, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Lee, S.H.; Jhun, J.; Choi, J.; Jung, K.; Cho, K.H.; Kim, S.J.; Yang, C.W.; Park, S.-H.; Cho, M.-L. The Combination of Probiotic Complex, Rosavin, and Zinc Improves Pain and Cartilage Destruction in an Osteoarthritis Rat Model. J. Med. Food 2018, 21, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kwon, J.Y.; Jhun, J.; Jung, K.; Park, S.-H.; Yang, C.W.; Cho, Y.; Kim, S.J.; Cho, M.-L. Lactobacillus acidophilus ameliorates pain and cartilage degradation in experimental osteoarthritis. Immunol. Lett. 2018, 203, 6–14. [Google Scholar] [CrossRef]
- Sinusas, K. Osteoarthritis: Diagnosis and treatment. Am. Fam. Physician 2012, 85, 49–56. [Google Scholar]
- So, J.-S.; Song, M.-K.; Kwon, H.-K.; Lee, C.-G.; Chae, C.-S.; Sahoo, A.; Jash, A.; Lee, S.H.; Park, Z.Y.; Im, S.-H. Lactobacillus casei enhances type II collagen/glucosamine-mediated suppression of inflammatory responses in experimental osteoarthritis. Life Sci. 2011, 88, 358–366. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Burmester, G.R.; Pope, J.E. Novel treatment strategies in rheumatoid arthritis. Lancet 2017, 389, 2338–2348. [Google Scholar] [CrossRef]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Guo, R.; Ju, Y.; Wang, Q.; Zhu, J.; Xie, Y.; Zheng, Y.; Li, T.; Liu, Z.; Lu, L.; et al. A single bacterium restores the microbiome dysbiosis to protect bones from destruction in a rat model of rheumatoid arthritis. Microbiome 2019, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaghef-Mehrabany, E.; Alipour, B.; Homayouni-Rad, A.; Sharif, S.-K.; Asghari-Jafarabadi, M.; Zavvari, S. Probiotic supplementation improves inflammatory status in patients with rheumatoid arthritis. Nutrition 2014, 30, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, P.; Hasanzadeh, S.; Tahvildari, A.; Farsi, Y.; Arbabi, M.; Mota, J.F.; Sechi, L.A.; Nasiri, M.J. Is there any association between gut microbiota and type 1 diabetes? A systematic review. Gut Pathog. 2019, 11, 1–10. [Google Scholar] [CrossRef]
- Mishra, S.; Wang, S.; Nagpal, R.; Miller, B.; Singh, R.; Taraphder, S.; Yadav, H. Probiotics and Prebiotics for the Amelioration of Type 1 Diabetes: Present and Future Perspectives. Microorganisms 2019, 7, 67. [Google Scholar] [CrossRef]
- Knudsen, J.K. Gut Microbiota in Bone Health and Diabetes. Curr. Osteoporos. Rep. 2021, 18, 462–479. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, R.; Rohilla, L.; Jacob, N.; Yadav, J.; Sachdeva, N. A high potency multi-strain probiotic improves glycemic control in children with new-onset type 1 diabetes mellitus: A randomized, double-blind, and placebo-controlled pilot study. Pediatr. Diabetes 2021, 22, 1014–1022. [Google Scholar] [CrossRef]
- Groele, L.; Szajewska, H.; Szypowska, A. Effects of Lactobacillus rhamnosus GG and Bifidobacterium lactis Bb12 on beta-cell function in children with newly diagnosed type 1 diabetes: Protocol of a randomised controlled trial. BMJ Open 2017, 7, e017178. [Google Scholar] [CrossRef]
- Salgaço, M.K.; Oliveira, L.G.S.; Costa, G.N.; Bianchi, F.; Sivieri, K. Relationship between gut microbiota, probiotics, and type 2 diabetes mellitus. Appl. Microbiol. Biotechnol. 2019, 103, 9229–9238. [Google Scholar] [CrossRef]
- Irwin, R.; Lin, H.; Motyl, K.J.; McCabe, L. Normal Bone Density Obtained in the Absence of Insulin Receptor Expression in Bone. Endocrinology 2006, 147, 5760–5767. [Google Scholar] [CrossRef] [PubMed]
- Raisingani, M.; Preneet, B.; Kohn, B.; Yakar, S. Skeletal growth and bone mineral acquisition in type 1 diabetic children; abnormalities of the GH/IGF-1 axis. Growth Horm. IGF Res. 2017, 34, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wongdee, K.; Krishnamra, N.; Charoenphandhu, N. Derangement of calcium metabolism in diabetes mellitus: Negative outcome from the synergy between impaired bone turnover and intestinal calcium absorption. J. Physiol. Sci. 2016, 67, 71–81. [Google Scholar] [CrossRef]
- Fatima, N.; Faisal, S.M.; Zubair, S.; Ajmal, M.; Siddiqui, S.S.; Moin, S.; Owais, M. Role of Pro-Inflammatory Cytokines and Biochemical Markers in the Pathogenesis of Type 1 Diabetes: Correlation with Age and Glycemic Condition in Diabetic Human Subjects. PLoS ONE 2016, 11, e0161548. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, S.; Sordi, V.; Bolla, A.M.; Saita, D.; Ferrarese, R.; Canducci, F.; Clementi, M.; Invernizzi, F.; Mariani, A.; Bonfanti, R.; et al. Duodenal Mucosa of Patients With Type 1 Diabetes Shows Distinctive Inflammatory Profile and Microbiota. J. Clin. Endocrinol. Metab. 2017, 102, 1468–1477. [Google Scholar] [CrossRef]
- Greer, R.L.; Dong, X.; de Moraes, A.; Zielke, R.A.; Fernandes, G.R.; Peremyslova, E.; Vasquez-Perez, S.; Schoenborn, A.A.; Gomes, E.P.; Pereira, A.C.; et al. Akkermansia muciniphila mediates negative effects of IFNγ on glucose metabolism. Nat. Commun. 2016, 7, 13329. [Google Scholar] [CrossRef]
- Sedighi, M.; Razavi, S.; Navab-Moghadam, F.; Khamseh, M.E.; Alaei-Shahmiri, F.; Mehrtash, A.; Amirmozafari, N. Comparison of gut microbiota in adult patients with type 2 diabetes and healthy individuals. Microb. Pathog. 2017, 111, 362–369. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut microbiota dysbiosis associated with glucose metabolism disorders and the metabolic syndrome in older adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Biagi, E.; Soverini, M.; Consolandi, C.; Quercia, S.; Severgnini, M.; Peano, C.; Turroni, S.; Rampelli, S.; Pozzilli, P.; et al. Modulation of gut microbiota dysbioses in type 2 diabetic patients by macrobiotic Ma-Pi 2 diet. Br. J. Nutr. 2016, 116, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Groele, L.; Szajewska, H.; Szalecki, M.; Świderska, J.; Wysocka-Mincewicz, M.; Ochocińska, A.; Stelmaszczyk-Emmel, A.; Demkow, U.; Szypowska, A. Lack of effect of Lactobacillus rhamnosus GG and Bifidobacterium lactis Bb12 on beta-cell function in children with newly diagnosed type 1 diabetes: A randomised controlled trial. BMJ Open Diabetes Res. Care 2021, 9, e001523. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Sun, X.; Li, J.; Li, Z.; Hu, Q.; Li, L.; Hao, X.; Song, M.; Li, C. Using probiotics for type 2 diabetes mellitus intervention: Advances, questions, and potential. Crit. Rev. Food Sci. Nutr. 2019, 60, 670–683. [Google Scholar] [CrossRef]
- Kim, S.-W.; Park, K.-Y.; Kim, B.; Kim, E.; Hyun, C.-K. Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production. Biochem. Biophys. Res. Commun. 2013, 431, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Fontané, L.; Benaiges, D.; Goday, A.; Llauradó, G.; Pedro-Botet, J. Influencia de la microbiota y de los probióticos en la obesidad. Clínica E Investig. En Arterioscler. 2018, 30, 271–279. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef]
- Montan, P.D.; Sourlas, A.; Olivero, J.; Silverio, D.; Guzman, E.; Kosmas, C.E. Pharmacologic therapy of obesity: Mechanisms of action and cardiometabolic effects. Ann. Transl. Med. 2019, 7, 393. [Google Scholar] [CrossRef]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalliomäki, M.; Collado, M.C.; Salminen, S.; Isolauri, E. Early differences in fecal microbiota composition in children may predict overweight. Am. J. Clin. Nutr. 2008, 87, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; Ison, J.; Voor, M.J.; Tyagi, N. Probiotics Stimulate Bone Formation in Obese Mice via Histone Methylations. Theranostics 2021, 11, 8605–8623. [Google Scholar] [CrossRef]
- Briot, K.; Geusens, P.; Em Bultink, I.; Lems, W.F.; Roux, C. Inflammatory diseases and bone fragility. Osteoporos. Int. 2017, 28, 3301–3314. [Google Scholar] [CrossRef]
- Roux, C. Osteoporosis in inflammatory joint diseases. Osteoporos. Int. 2010, 22, 421–433. [Google Scholar] [CrossRef]
- Arden, N.; Nevitt, M.C. Osteoarthritis: Epidemiology. Best Pract. Res. Clin. Rheumatol. 2006, 20, 3–25. [Google Scholar] [CrossRef]
- Wei, Z.; Li, F.; Pi, G. Association Between Gut Microbiota and Osteoarthritis: A Review of Evidence for Potential Mechanisms and Therapeutics. Front. Cell. Infect. Microbiol. 2022, 12, 812596. [Google Scholar] [CrossRef]
- Zhang, Y.; Jordan, J.M. Epidemiology of Osteoarthritis. Clin. Geriatr. Med. 2010, 26, 355–369. [Google Scholar] [CrossRef]
- Lyu, J.-L.; Wang, T.-M.; Chen, Y.-H.; Chang, S.-T.; Wu, M.-S.; Lin, Y.-H.; Kuan, C.-M. Oral intake of Streptococcus thermophilus improves knee osteoarthritis degeneration: A randomized, double-blind, placebo-controlled clinical study. Heliyon 2020, 6, e03757. [Google Scholar] [CrossRef]
- McInnes, I.B. The Pathogenesis of Rheumatoid Arthritis. New Engl. J. Med. 2011, 15, 2205–2219. [Google Scholar] [CrossRef] [PubMed]
- Zamani, B.; Golkar, H.R.; Farshbaf, S.; Emadi-Baygi, M.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akhavan, R.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z. Clinical and metabolic response to probiotic supplementation in patients with rheumatoid arthritis: A randomized, double-blind, placebo-controlled trial. Int. J. Rheum. Dis. 2016, 19, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [Google Scholar] [CrossRef]
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 Diabetes mellitus. Nat. Rev. Dis. Primers 2017, 3, 17016. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Prim. 2015, 1, 15019. [Google Scholar] [CrossRef]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef]
- Blahova, J.; Martiniakova, M.; Babikova, M.; Kovacova, V.; Mondockova, V.; Omelka, R. Pharmaceutical Drugs and Natural Therapeutic Products for the Treatment of Type 2 Diabetes Mellitus. Pharmaceuticals 2021, 14, 806. [Google Scholar] [CrossRef]
- Isidro, M.L. Bone Disease in Diabetes. Curr. Diabetes Rev. 2010, 6, 144–155. [Google Scholar] [CrossRef]
- Oei, L.; Zillikens, M.C.; Dehghan, A.; Buitendijk, G.H.; Castaño-Betancourt, M.C.; Estrada, K.; Stolk, L.; Oei, E.H.; van Meurs, J.B.; Janssen, J.A.; et al. High Bone Mineral Density and Fracture Risk in Type 2 Diabetes as Skeletal Complications of Inadequate Glucose Control. Diabetes Care 2013, 36, 1619–1628. [Google Scholar] [CrossRef]
- Qiu, J.; Li, C.; Dong, Z.; Wang, J. Is diabetes mellitus a risk factor for low bone density: A systematic review and meta-analysis. BMC Endocr. Disord. 2021, 21, 65. [Google Scholar] [CrossRef]
- Asokan, A.G.; Jaganathan, J.; Philip, R.; Soman, R.R.; Sebastian, S.T.; Pullishery, F. Evaluation of bone mineral density among type 2 diabetes mellitus patients in South Karnataka. J. Nat. Sci. Biol. Med. 2017, 8, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, N.; Conte, C.; Eastell, R.; Ewing, S.K.; Bauer, D.C.; Strotmeyer, E.S.; Black, D.M.; Samelson, E.J.; Vittinghoff, E.; Schwartz, A.V. Bone Turnover Markers Do Not Predict Fracture Risk in Type 2 Diabetes. J. Bone Miner. Res. 2020, 35, 2363–2371. [Google Scholar] [CrossRef] [PubMed]
- Starup-Linde, J.; Lykkeboe, S.; Handberg, A.; Vestergaard, P.; Høyem, P.; Fleischer, J.; Hansen, T.K.; Poulsen, P.L.; Laugesen, E. Glucose variability and low bone turnover in people with type 2 diabetes. Bone 2021, 153, 116159. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Kyriienko, D.; Komissarenko, I. Effect of alive probiotic on insulin resistance in type 2 diabetes patients: Randomized clinical trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2018, 12, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; He, C.; He, W.; Yang, M.; Luo, X.; Li, C. Obesity and Bone Health: A Complex Link. Front. Cell Dev. Biol. 2020, 8, 600181. [Google Scholar] [CrossRef]
- Must, A.; Spadano, J.; Coakley, E.H.; Field, A.E.; Colditz, G.; Dietz, W.H. The Disease Burden Associated With Overweight and Obesity. JAMA 1999, 282, 1523–1529. [Google Scholar] [CrossRef]
- Greco, E.; Lenzi, A.; Migliaccio, S. The obesity of bone. Ther. Adv. Endocrinol. Metab. 2015, 6, 273–286. [Google Scholar] [CrossRef]
- Schott, E.M.; Farnsworth, C.W.; Grier, A.; Lillis, J.A.; Soniwala, S.; Dadourian, G.H.; Bell, R.D.; Doolittle, M.L.; Villani, D.A.; Awad, H.; et al. Targeting the gut microbiome to treat the osteoarthritis of obesity. JCI Insight 2018, 3, 95997. [Google Scholar] [CrossRef]
- Biermann, J.S.; Adkins, D.R.; Agulnik, M.; Benjamin, R.S.; Brigman, B.; Butrynski, J.E.; Cheong, D.; Chow, W.; Curry, W.T.; Frassica, D.A.; et al. Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2013, 11, 36. [Google Scholar]
- Ferguson, J.L.; Turner, S.P. Bone Cancer: Diagnosis and Treatment Principles. Am. Fam. Physician 2018, 98, 205–213. [Google Scholar] [PubMed]
- Weilbaecher, K.N.; Guise, T.A.; McCauley, L.K. Cancer to bone: A fatal attraction. Nat. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Wenhui, Y.; Zhongyu, X.; Kai, C.; Zhaopeng, C.; Jinteng, L.; Mengjun, M.; Zepeng, S.; Yunshu, C.; Peng, W.; Yanfeng, W.; et al. Variations in the Gut Microbiota in Breast Cancer Occurrence and Bone Metastasis. Front. Microbiol. 2022, 13, 894283. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Goyal, A. Evolving Roles of Probiotics in Cancer Prophylaxis and Therapy. Probiotics Antimicrob. Proteins 2013, 5, 59–67. [Google Scholar] [CrossRef]
- Das, M.; Cronin, O.; Keohane, D.M.; Cormac, E.M.; Nugent, H.; Nugent, M.; Molloy, C.; O’Toole, P.W.; Shanahan, F.; Molloy, M.G.; et al. Gut microbiota alterations associated with reduced bone mineral density in older adults. Rheumatology 2019, 58, 2295–2304. [Google Scholar] [CrossRef]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Zeibich, L.; Koebele, S.V.; Bernaud, V.E.; Ilhan, Z.E.; Dirks, B.; Northup-Smith, S.N.; Neeley, R.; Maldonado, J.; Nirmalkar, K.; Files, J.A.; et al. Surgical Menopause and Estrogen Therapy Modulate the Gut Microbiota, Obesity Markers, and Spatial Memory in Rats. Front. Cell. Infect. Microbiol. 2021, 11, 702628. [Google Scholar] [CrossRef]
- Laborda-Illanes, A.; Sanchez-Alcoholado, L.; Dominguez-Recio, M.E.; Jimenez-Rodriguez, B.; Lavado, R.; Comino-Méndez, I.; Alba, E.; Queipo-Ortuño, M.I. Breast and Gut Microbiota Action Mechanisms in Breast Cancer Pathogenesis and Treatment. Cancers 2020, 12, 2465. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, I.Y.; Bae, E.-K.; Jeon, C.H.; Ahn, K.-S.; Cha, H.-S. Selective estrogen receptor modulator lasofoxifene suppresses spondyloarthritis manifestation and affects characteristics of gut microbiota in zymosan-induced SKG mice. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Rogers, M.A.M.; Aronoff, D.M. The influence of non-steroidal anti-inflammatory drugs on the gut microbiome. Clin. Microbiol. Infect. 2016, 22, 178.e1–178.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, D.; Law, H.K.-W.; Wu, Y.; Zhu, G.-H.; Huang, W.-Y.; Kang, Y. Integrative Analysis of Gut Microbiota and Fecal Metabolites in Rats after Prednisone Treatment. Microbiol. Spectr. 2021, 9, e00650-21. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza-García, O.; Castro-Alarcón, N.; Pérez-Rubio, G.; Guzmán-Guzmán, I.P. DMARDs–Gut Microbiota Feedback: Implications in the Response to Therapy. Biomolecules 2020, 10, 1479. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Jiang, X.; Chu, W. Shifts in the gut microbiota of mice in response to dexamethasone administration. Int. Microbiol. 2020, 23, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, J.; Gaurav, C.; Muhammad, U.; Chen, Y.; Li, X.; Chen, J.; Wang, Z. CUMS and dexamethasone induce depression-like phenotypes in mice by differentially altering gut microbiota and triggering macroglia activation. Gen. Psychiatry 2021, 34, e100529. [Google Scholar] [CrossRef] [PubMed]
- Kihl, P.; Krych, L.; Buschard, K.; Wesley, J.D.; Kot, W.; Hansen, A.K.; Nielsen, D.S.; Von Herrath, M.G. Oral insulin does not alter gut microbiota composition of NOD mice. Diabetes/Metab. Res. Rev. 2018, 34, e3010. [Google Scholar] [CrossRef]
- Tong, X.; Xu, J.; Lian, F.; Yu, X.; Zhao, Y.; Xu, L.; Zhang, M.; Zhao, X.; Shen, J.; Wu, S.; et al. Structural Alteration of Gut Microbiota during the Amelioration of Human Type 2 Diabetes with Hyperlipidemia by Metformin and a Traditional Chinese Herbal Formula: A Multicenter, Randomized, Open Label Clinical Trial. mBio 2018, 9, e02392-17. [Google Scholar] [CrossRef]
- Wu, H.; Esteve, E.; Tremaroli, V.; Khan, M.T.; Caesar, R.; Mannerås-Holm, L.; Ståhlman, M.; Olsson, L.M.; Serino, M.; Planas-Fèlix, M.; et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat. Med. 2017, 23, 850–858. [Google Scholar] [CrossRef]
- Ke, J.; An, Y.; Cao, B.; Lang, J.; Wu, N.; Zhao, D. Orlistat-Induced Gut Microbiota Modification in Obese Mice. Evidence-Based Complement. Altern. Med. 2020, 2020, 9818349. [Google Scholar] [CrossRef]
- Huang, J.; Wei, S.; Jiang, C.; Xiao, Z.; Liu, J.; Peng, W.; Zhang, B.; Li, W. Involvement of Abnormal Gut Microbiota Composition and Function in Doxorubicin-Induced Cardiotoxicity. Front. Cell. Infect. Microbiol. 2022, 12, 808837. [Google Scholar] [CrossRef]
Figure 1.
The GM has various roles in the host organism. It can affect various physiological roles, such as fermentation of complex carbohydrates [24], immune system [25], synthesis of neuroactive compounds [26], vitamin synthesis [27] and bile acid metabolism [28]. It can also affect many pathophysiological (during dysbiosis) roles [7]. (Created with BioRender.com).
Figure 1.
The GM has various roles in the host organism. It can affect various physiological roles, such as fermentation of complex carbohydrates [24], immune system [25], synthesis of neuroactive compounds [26], vitamin synthesis [27] and bile acid metabolism [28]. It can also affect many pathophysiological (during dysbiosis) roles [7]. (Created with BioRender.com).

Figure 2.
Lymphocytes can regulate bone remodeling through the expression of cytokines, as well as RANKL and OPG. (Created with BioRender.com).
Figure 2.
Lymphocytes can regulate bone remodeling through the expression of cytokines, as well as RANKL and OPG. (Created with BioRender.com).

Figure 3.
Mice lacking GM exhibited increased trabecular BMD, which was associated with decreased OCs, CD4+ T cells and OC precursor cells and lowered TNFα and IL-6 expression [40]. T cells are known to secrete osteoclastogenic cytokines such as TNF- α and IL-6 [41]. Intestinal bacteria are needed to develop the immune system. In GF mice, the mucosal immune system is undeveloped, having reduced the lamina propria CD4+ T cells. Moreover, the spleen and lymph nodes are also affected, as they are relatively structureless with poorly formed B- and T-cell zones [42]. (Created with BioRender.com).
Figure 3.
Mice lacking GM exhibited increased trabecular BMD, which was associated with decreased OCs, CD4+ T cells and OC precursor cells and lowered TNFα and IL-6 expression [40]. T cells are known to secrete osteoclastogenic cytokines such as TNF- α and IL-6 [41]. Intestinal bacteria are needed to develop the immune system. In GF mice, the mucosal immune system is undeveloped, having reduced the lamina propria CD4+ T cells. Moreover, the spleen and lymph nodes are also affected, as they are relatively structureless with poorly formed B- and T-cell zones [42]. (Created with BioRender.com).

Table 2.
Summary table of various GM alterations and interactions related to the treatment of bone-related diseases.
Table 2.
Summary table of various GM alterations and interactions related to the treatment of bone-related diseases.
| Disease | Selected Drugs | GM Alterations Associated with the Treatment | Interactions between the Treatment and GM | References |
|---|---|---|---|---|
| osteoporosis | bisphosphonates | not known | supposed GM-related immunosuppression via inhibition of the mevalonate pathway | [70] |
| estrogen therapy | AS: increase in Clostridium, Turicibacter, Romboutsia, Parabacteroides decrease in Kineothrix, Ruminococcus, Muribaculum | estrogen promotes microbiota, which improves T-regulatory cell function and suppresses inflammation; GM influences estrogen metabolism | [170,171] | |
| raloxifene (SERMs) | not known | lasofoxifene, a SERM not approved for clinical use, affects the composition and biodiversity of GM | [172] | |
| osteoarthritis | ibuprofen (NSAIDs) | HS: increase in families Propionibacteriaceae, Pseudomonadaceae, Puniceicoccaceae, Rikenellaceae, Acidaminococcaceae, Enterococcaceae, Erysipelotrichaceae | NSAIDs inhibit some positive effects of microbiota through enzyme inhibition. | [173] |
| prednisone (glucocorticoids) | AS: increase in Anaerobacterium decrease in Eisenbergiella, Alistipes, Clostridium | microbiota-derived SCFAs might be involved in the pharmacology of prednisone | [174] | |
| rheumatoid arthritis | methotrexate | HS: increase in Bacteroides, Faecalibacterium, Clostridia decrease in Haemophilus, Enterobacteriales, Bacteroidetes | GM includes dihydrofolate reductase enzyme, so it can modulate methotrexate metabolism; at the same time, methotrexate can modulate microbial metabolism | [175] |
| dexamethasone (glucocorticoids) | AS: increase in Lachnospiraceae, Oscillibacter, Ruminococcaceae, Ruminiclostridium, Anaerotruncus, Butyricicoccus, Enterobacteriaceae, Escherichia Shigella, Gammaproteobacteria, Enterobacteriales, Anaerofustis, Erysipelotrichaceae decrease in Lactobacillus, Enterorhabdus, Pseudomonas, Clostridium | GM directly mediates the therapeutic efficiency and side effects of glucocorticoids | [176,177] | |
| T1DM | insulin | not known | oral insulin administration had no effect on GM, possibly due to the insulin degradation in the intestine, and its low bioavailability | [178] |
| T2DM | metformin | HS: increase in Megamonas, Escherichia/Shigella, Klebsiella, Blautia, Fusobacterium, Bifidobacterium, Intestinibacter decrease in Alistipes, Bacteroidetes | metformin clinical benefits are partly mediated by bacteria-specific mechanisms such as glucose-SGLT1-sensing glucoregulatory pathway | [179,180] |
| obesity | orlistat | AS: increase in Pseudomonas, Rhodococcus, Roseburia, Acetivibrio | orlistat showed enrichment in genes involved in the endocrine and lipid metabolism including ALA metabolism; GM plays a role in the metabolism of ALA to conjugated linolenic acid, which was documented to have antiadipogenic effects | [181] |
| bone cancer | doxorubicin | AS: increase in Faecalibaculum, Dubosiella, Lachnospiraceae decrease in Allobaculum, Muribaculum, Lachnoclostridium | doxorubicin application is accompanied by variation of metabolism processes such as amino acid metabolism, glycan biosynthesis and metabolism, lipid metabolism, and other secondary metabolites | [182] |
GM—gut microbiome; AS—animal studies; HS—human studies; T1DM—type 1 diabetes mellitus; T2DM—type 2 diabetes mellitus; SERMs—selective estrogen receptor modulators; NSAIDs—non-steroidal anti-inflammatory drugs; SCFAs—short-chain fatty acids; SGLT1—sodium/glucose cotransporter 1; ALA—alpha-linolenic acid.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zemanova, N.; Omelka, R.; Mondockova, V.; Kovacova, V.; Martiniakova, M. Roles of Gut Microbiome in Bone Homeostasis and Its Relationship with Bone-Related Diseases. Biology 2022, 11, 1402. https://doi.org/10.3390/biology11101402
AMA Style
Zemanova N, Omelka R, Mondockova V, Kovacova V, Martiniakova M. Roles of Gut Microbiome in Bone Homeostasis and Its Relationship with Bone-Related Diseases. Biology. 2022; 11(10):1402. https://doi.org/10.3390/biology11101402
Chicago/Turabian StyleZemanova, Nina, Radoslav Omelka, Vladimira Mondockova, Veronika Kovacova, and Monika Martiniakova. 2022. "Roles of Gut Microbiome in Bone Homeostasis and Its Relationship with Bone-Related Diseases" Biology 11, no. 10: 1402. https://doi.org/10.3390/biology11101402
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.