
AMPK and Diseases: State of the Art Regulation by AMPK-Targeting Molecules



1
Experimental Neurology Unit, School of Medicine and Surgery, University of Milano-Bicocca, 20900 Monza, Italy
2
SPILLOproject—Innovative In Silico Solutions for Drug R&D and Pharmacology, 20037 Paderno Dugnano, Italy
*
Author to whom correspondence should be addressed.
Biology 2022, 11(7), 1041; https://doi.org/10.3390/biology11071041
Submission received: 9 June 2022
/
Revised: 4 July 2022
/
Accepted: 6 July 2022
/
Published: 11 July 2022
(This article belongs to the Topic Biomedical Applications of Enzymes)
Abstract
:Simple Summary
AMPK is an enzyme that plays a role in cellular energy mechanisms and controls many metabolic and physiological processes. AMPK is dysregulated in different diseases, making it a potential therapeutic target. AMPK can be activated in a direct or indirect way. In this review, we discuss different AMPK activating compounds and especially focus our attention on those compounds that imitate physiological mechanisms of AMPK activation.
Abstract
5′-adenosine monophosphate (AMP)-activated protein kinase (AMPK) is an enzyme that regulates cellular energy homeostasis, glucose, fatty acid uptake, and oxidation at low cellular ATP levels. AMPK plays an important role in several molecular mechanisms and physiological conditions. It has been shown that AMPK can be dysregulated in different chronic diseases, such as inflammation, diabetes, obesity, and cancer. Due to its fundamental role in physiological and pathological cellular processes, AMPK is considered one of the most important targets for treating different diseases. Over decades, different AMPK targeting compounds have been discovered, starting from those that activate AMPK indirectly by altering intracellular AMP:ATP ratio to compounds that activate AMPK directly by binding to its activation sites. However, indirect altering of intracellular AMP:ATP ratio influences different cellular processes and induces side effects. Direct AMPK activators showed more promising results in eliminating side effects as well as the possibility to engineer drugs for specific AMPK isoforms activation. In this review, we discuss AMPK targeting drugs, especially concentrating on those compounds that activate AMPK by mimicking AMP. These compounds are poorly described in the literature and still, a lot of questions remain unanswered about the exact mechanism of AMP regulation. Future investigation of the mechanism of AMP binding will make it possible to develop new compounds that, in combination with others, can activate AMPK in a synergistic manner.
1. AMPK: An Important Player in Regulating Cellular Processes
AMPK is a heterotrimeric complex composed of one catalytic α-subunit and two regulatory β- and γ-subunits. AMPK can consist of two isoforms, each of α- and β-subunits, and three isoforms of γ-subunit [1]. In mammals exist up to 12 distinct combinations of AMPK complexes. These complexes are shown to differ depending on tissue-, cell-type, regulation mechanism, and biochemical properties [2,3].
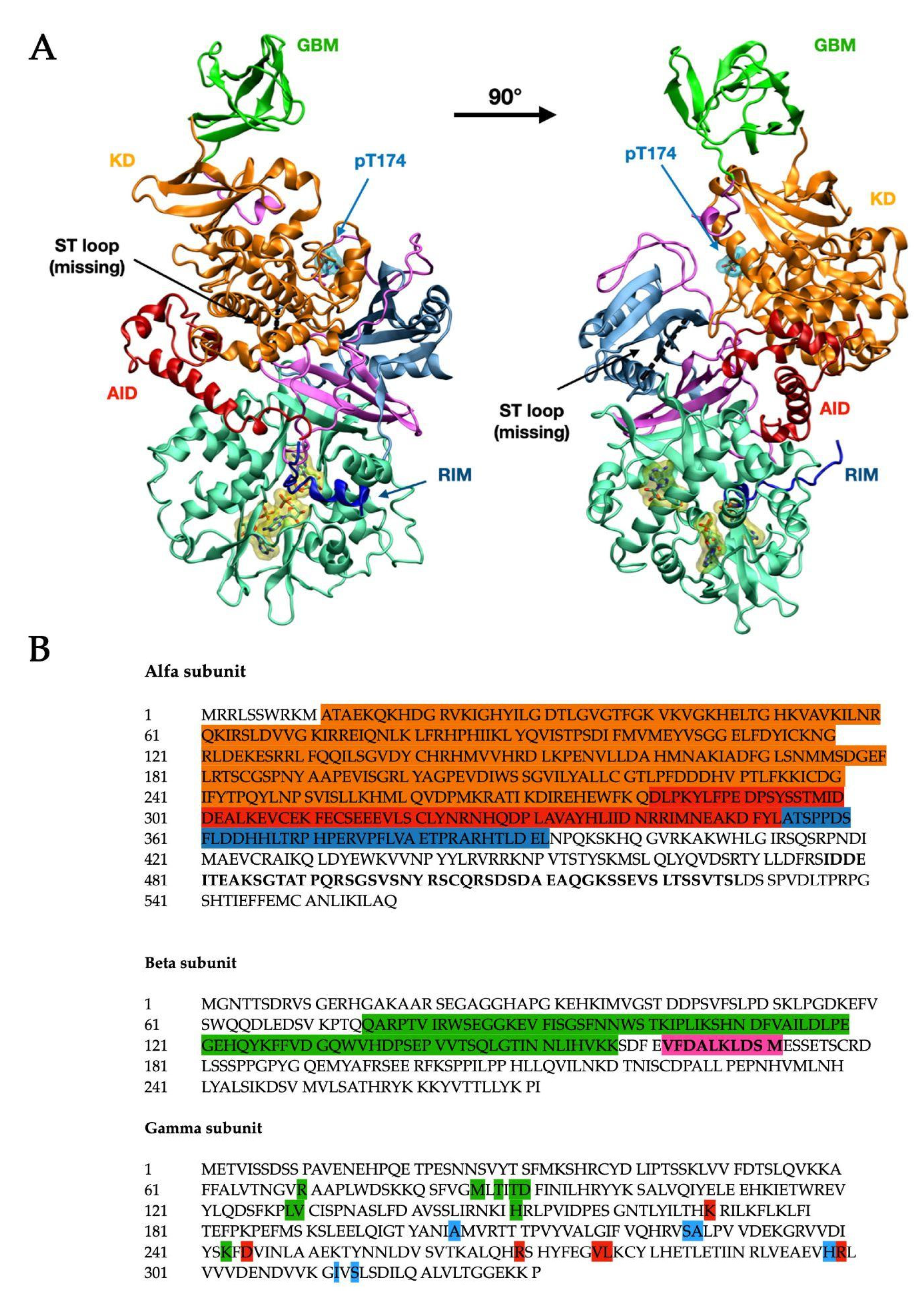
α-subunit (Figure 1 in blue-grey) consists of the N-terminal kinase domain (KD) (Figure 1, in orange) [4] and C terminus that binds the β (Figure 1, in pink) and γ (Figure 1, in light teal green) subunits and comprises important regulatory domains, namely a so-called autoinhibitory domain (AID) (Figure 1, in red), the α-linker (which interacts with the γ-subunit through two regulatory subunit-interacting motifs, RIM motifs—Figure 1, in blue), and a serine/threonine-rich domain (ST loop).
The β-subunit contains two conserved glycogen-binding domains (GBM) (Figure 1, in bright green), involved in glycogen sensing, and a C terminus domain that binds the α and γ-subunits. The β-subunit also includes a site at its N terminus, which supports the targeting of AMPK to cellular membranes [5].
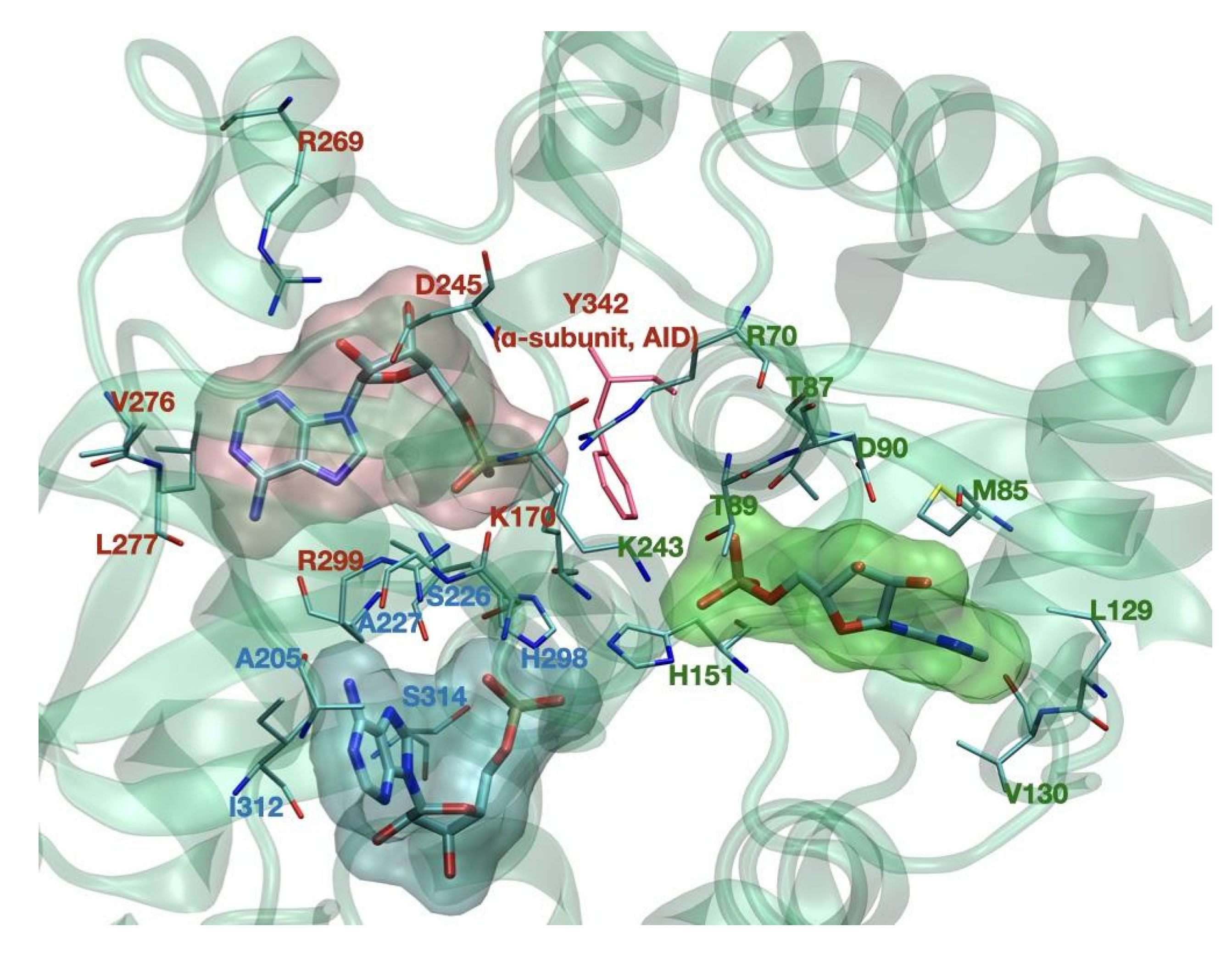
The γ subunit consists of two domains conserved among all species and called Bateman domains [6], which contain two cystathionine β-synthase repeats (CBS) each, as represented in Figure 2. The four CBS repeats function as four universal adenine nucleotide-binding sites and can competitively bind AMP, ADP, or ATP [7]. In mammals, sites 1 and 3 can exchange adenine nucleotides competitively. Site 2 (CBS2) lacks the key aspartate required for nucleotide binding and consequently is non-functional. Site 4 can also bind AMP and ATP but has a higher affinity for AMP [8].
Figure 1.
Structural visualization (A)and primary sequence (B) of human AMPK (PDB ID: 4RER [4]). The structure (A) is composed of the α1 subunit (blue-grey) and the β2 (pink) and γ1 (light teal green, with three bound AMP residues) regulatory subunits. The α subunit presents the kinase domain (KD, orange), the autoinhibitory domain (AID, red), and the residue inhibitory motif (RIM, blue). The black dashed line indicates the position of the ST loop, while the phosphorylated Thr-174 (Thr-172 in α2) is shown with the cyan surface. On the β subunit, the glycogen-binding domain (GBM) is shown in bright green. The numbers of starting and ending residues for each highlighted domain are reported in Table 1. In the primary sequence (B), the domains are highlighted in the same color code: the α1 subunit (UniProt: Q13131) shows the KD domain (orange), AID (red), and RIM (blue), and the sequence for the ST loop has been bolded. The β2 subunit (UniProt: O43741) shows the GBM domain (green) and the sequence for the ADaM site (bolded, highlighted in pink). In the γ1 subunit (UniProt: P54619), all residues belonging to the respective CBM have been highlighted using the same color code as shown in Figure 2 (green: CBM1, red: CBS3, blue: CBS4).
Figure 1.
Structural visualization (A)and primary sequence (B) of human AMPK (PDB ID: 4RER [4]). The structure (A) is composed of the α1 subunit (blue-grey) and the β2 (pink) and γ1 (light teal green, with three bound AMP residues) regulatory subunits. The α subunit presents the kinase domain (KD, orange), the autoinhibitory domain (AID, red), and the residue inhibitory motif (RIM, blue). The black dashed line indicates the position of the ST loop, while the phosphorylated Thr-174 (Thr-172 in α2) is shown with the cyan surface. On the β subunit, the glycogen-binding domain (GBM) is shown in bright green. The numbers of starting and ending residues for each highlighted domain are reported in Table 1. In the primary sequence (B), the domains are highlighted in the same color code: the α1 subunit (UniProt: Q13131) shows the KD domain (orange), AID (red), and RIM (blue), and the sequence for the ST loop has been bolded. The β2 subunit (UniProt: O43741) shows the GBM domain (green) and the sequence for the ADaM site (bolded, highlighted in pink). In the γ1 subunit (UniProt: P54619), all residues belonging to the respective CBM have been highlighted using the same color code as shown in Figure 2 (green: CBM1, red: CBS3, blue: CBS4).

Important knowledge of AMPK activity regulation by AMP is inferred from the experimentally solved (by X-ray diffraction or cryo-EM) 3D structures of the mammalian AMPK complex. Table 2 reports the 3D structures (including the three α, β, and γ subunits) of human AMPK currently (May 2022) available from the RCSB Protein Data Bank. Recent Cryo-EM structures allow for interesting protein-protein interactions, such as 7JHH.
It is suggested that αRIM, which directly binds AMP at CBS3, starts AMPK activation, making this loop function as an adenine nucleotide sensor. In addition, the N-terminus of the loop folds back and is connected to the AID. When AMP is bound, the C-terminus of the AID and the N-terminus of the αRIM loop form an interaction, termed αRIM-1, that is bound to the unoccupied CBS2 site of the γ-subunit [15].
Direct binding of AMP and/or ADP to γ-subunit allosterically activates the complex and promotes the phosphorylation of Thr-174/Thr-172 (Figure 1, cyan surface) by at least three upstream kinases and three phosphatases: namely, liver kinase B1 (LKB1) [16], calcium-/calmodulin-dependent kinase kinase 2 (CaMKK2) [17], TGFβ-activated kinase 1 (TAK1) [18], protein phosphatase 2A (PP2A) [19], protein phosphatase 2C (PP2C) [20] and Mg2+/Mn2+-dependent protein phosphatase 1E (PPM1E) [21].
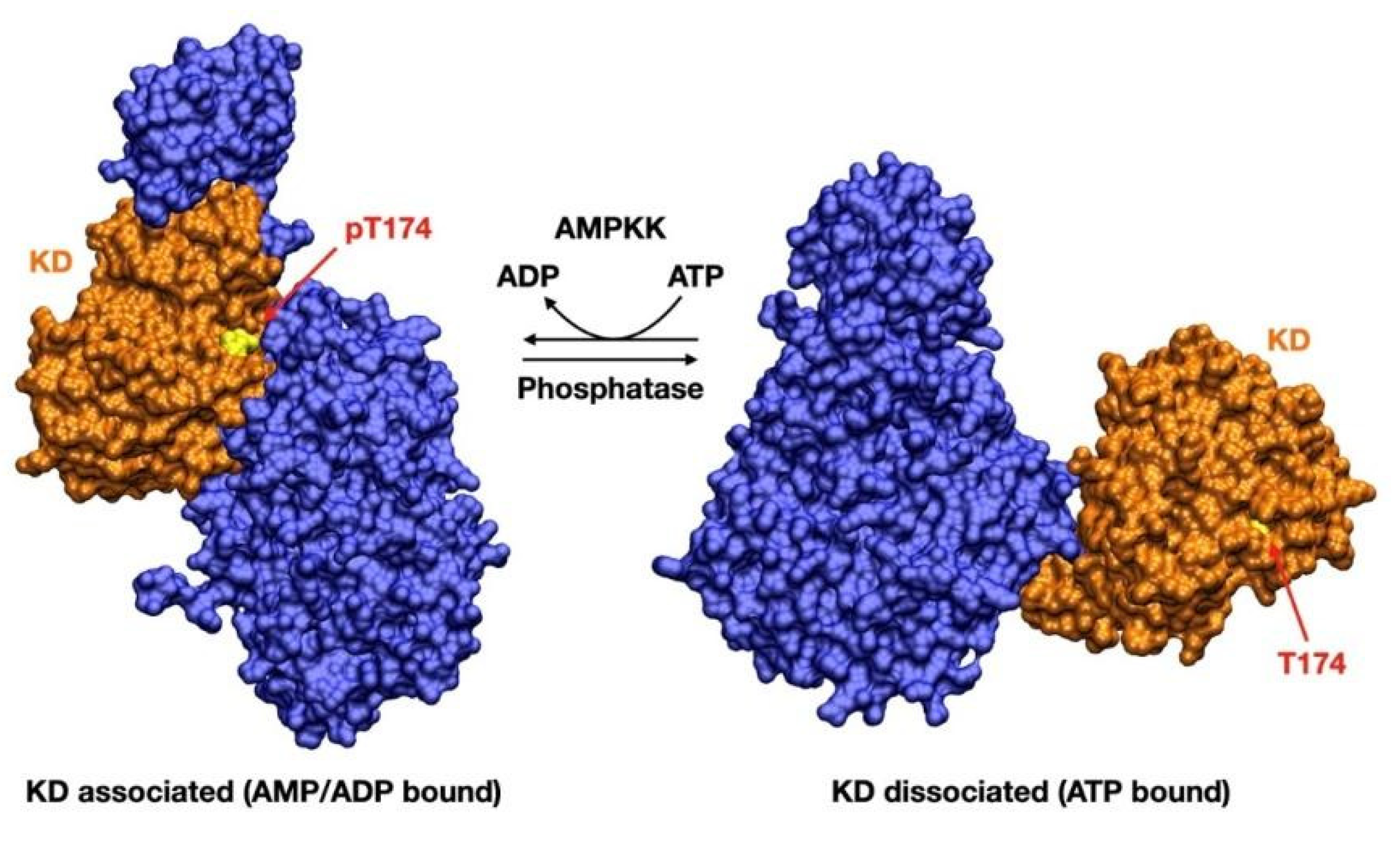
Once AMPK is phosphorylated on Thr-174/Thr-172, AMP preserves AMPK signaling by inhibiting dephosphorylation of pThr-174/172. ATP, on the other hand, competitively inhibits the binding of both AMP and ADP to the γ-subunit, as well as promoting Thr-174/Thr-172 dephosphorylation [22]. The αRIM2 is crucial both for allosteric activation and for protection against dephosphorylation. It is required for sensing the adenine nucleotide binding at γ-subunit and signal transduction to the α-catalytic domain, resulting in either stimulation (AMP) or inhibition (ATP) of AMPK activity [4]. Dephosphorylation of Thr-174/Thr-172 promotes a dramatic conformational change, which brings AMPK from its active (KD associated) to its inactive (KD displaced) state. In this state, the KD is displaced by a distance of around 80 Å from its previous position, thus preventing AMPK catalytic activation [7] (Figure 3).
The ability of the γ subunit to bind AMP, ADP, and ATP allows AMPK to sense the energy state of the cell. Under conditions of high AMP:ATP ratio, AMPK phosphorylates specific enzymes to generate ATP and decrease its consumption; in this way, restoration of energy balance can be reached, acting by turning off energy-consuming processes, such as protein synthesis, and turning on energy-generating processes, mitochondrial biogenesis or glucose metabolism [23]. Cellular ATP concentration is kept at a constant level to ensure adequate ATP supply, which is essential for cellular survival [24].
Due to it, AMPK plays an important role in cell growth and metabolic processes in different tissues such as the liver, muscle, and fat. AMPK directly phosphorylates acetyl-CoA carboxylase 1 (ACC1) and ACC2, suppressing the de novo synthesis of fatty acids and stimulating fatty acids oxidation that regulates overall cellular lipid metabolism [25]. AMPK can induce the inhibitory phosphorylation of the HMG-CoA reductase, which regulates cholesterol synthesis [26]. AMPK stimulates glucose uptake by enhanced GLUT1 and GLUT4 translocation to the plasma membrane and is the main regulator of glucose and fat metabolism in skeletal muscle during exercise [27]. Moreover, AMPK regulates protein synthesis. It is a high energy-consuming process that needs to be inhibited during cellular stress to preserve intracellular ATP. AMPK inhibits cap-dependent translation during both initiation and elongation steps in protein biosynthesis [28]. AMPK downregulates ribosomal RNA synthesis by inducing the inhibitory phosphorylation of transcription initiation factor 1A [29,30]. A recent study suggested that AMPK stimulates cap-independent and IRES-dependent translation of Hif-1α during energy stress to activate the expression of genes important for cell survival [31].
Moreover, different studies have shown that AMPK regulates autophagy processes [32]. First, AMPK directly phosphorylates and activates ULK1 to induce autophagy. Second, AMPK indirectly activates ULK1 by inhibiting mTORC1, which phosphorylates and inhibits ULK1 to disrupt the ULK1–AMPK interaction [33]. This double regulation of ULK1 and mTORC1 eliminates damaged mitochondria and maintains mitochondrial integrity during nutrient starvation [34].
It is also suggested that AMPK regulates mitochondrial homeostasis. AMPK upregulates several antioxidant genes, such as those encoding superoxide dismutase and uncoupling protein 2, which reduces superoxide levels and thioredoxin (TRX), a disulfide reductase, by phosphorylating and activating FOXO, so playing an important role in antioxidant defense during oxidative stress [35]. Some studies suggested that AMPK targets NRF2 and induces antioxidant defense [36].
Considering AMPK’s central role in regulating important cellular metabolic pathways, it has been proposed that it may have therapeutic importance for treating obesity, insulin resistance, type 2 diabetes (T2D), non-alcoholic fatty liver disease (NAFLD), and cardiovascular disease (CVD) [36]. Recently, AMPK has been gaining enormous attention as a target in Alzheimer’s Disease drug research. AMPK activators seem to be able to influence Aβ accumulation, tau aggregation, and oxidative stress [37]. Moreover, strong interest is given to targeting AMPK in cancer. It has been shown that AMPK activation can inhibit tumor cell proliferation and cell growth [38]. In 2003, the discovery of the tumor suppressor LKB1 as the major upstream kinase of AMPK established a link between an energy regulator and cancer pathogenesis, suggesting that the tumor suppressor functions of LKB1 could be mediated by AMPK [39]. Among other mechanisms, several studies showed a correlation between AMPK activation by numerous agents and COX-2 inhibition in different cancers [40]. Moreover, AMPK has been shown to induce phosphorylation of p53 in response to metabolic stress, which is required to initiate AMPK-dependent cell-cycle arrest [41]. Acetyl-CoA carboxylase (ACC) is a well-established downstream target of AMPK involved in lipid metabolism. In several cases, cancer cell proliferation and survival are dependent on ACC activity and inhibiting ACC results in apoptosis [42].
2. AMPK Activators
Given that AMPK activation regulates a lot of cellular physiological and pathological processes, it is not surprising that the development of new AMPK activating compounds has been intensively explored. The mechanisms by which compounds activate AMPK can be divided into three classes: (1) compounds that activate AMPK indirectly by increasing intracellular AMP and ADP levels; (2) compounds that activate AMPK directly by selectively binding to its domains; (3) compounds that mimic AMP or ATP and are thus able to bind AMPK at the γ-subunit.
2.1. Indirect AMPK Activators
Indirect AMPK activation can be caused by the intracellular accumulation of calcium or AMP. Because AMPK activity is regulated by phosphorylation and dephosphorylation events, the relationships between calcium and upstream kinases or phosphatases play a crucial role. For example, in muscle cells, the increase in cytosolic calcium affects AMPK activation and further influences GLUT-4 gene expression, a skeletal muscle-specific glucose transporter that mediates both insulin and contraction-stimulated glucose transport [43].
AMPK can be activated by compounds that inhibit ATP synthesis as depletion of ATP always causes an increase in AMP and ADP. In cells that are primarily using glycolysis to generate ATP, AMPK is activated by inhibitors of glycolysis such as 2-deoxyglucose. A much larger class of activators is those that inhibit mitochondrial ATP synthesis by inhibiting the respiratory chain, such as metformin, phenformin, antimycin A, oligomycin, and resveratrol [44,45,46]. These agents increase cellular AMP:ATP and ADP:ATP ratios. It is, however, obvious that compounds that inhibit mitochondrial function inhibit oxygen uptake, while those that inhibit glycolysis reduce lactate output and cause extracellular acidification [47]. Due to this, it is expected that different indirect agents, that act mostly by increasing the concentration of AMP, may lack specificity and trigger different unwanted side effects. For example, it has been shown that inhibition of the respiratory chain induced by metformin and phenformin develops life-threatening cases of lactic acidosis that results in phenformin being withdrawn from clinical use [48,49]. Orally administered metformin is effectively absorbed from the gastrointestinal tract to the portal vein. As a first-pass route, the liver is exposed to a high concentration of the drug, which causes gastrointestinal side effects (diarrhea, nausea, abdominal discomfort, anorexia), limiting its use in many patients [47].
There are many other compounds that activate AMPK by inhibiting mitochondrial ATP synthesis, for example, resveratrol, whose pro-oxidative effect leads to cellular oxidative stress limiting its dosage [50]. Moreover, sorafenib can target vascular endothelial growth factor (VEGF) [51], leading to undesirable cardiovascular events like hypertension, bleeding as well as other gastrointestinal disturbances and hand–foot skin reactions [4].
2.2. Direct AMPK Activators
Because of indirect AMPK activation side effects, particular attention has been focused on compounds that activate AMPK directly. It is assumed that direct activation of AMPK does not change the AMP:ATP ratio or alter oxygen uptake and does not inhibit mitochondrial function. Direct AMPK activating compounds can be distinguished into two groups: AMP mimetics, which mimic AMP and activate AMPK similarly to physiological ligands, or non-nucleoside activators that bind AMPK at some other sites.
It is shown that some small-molecule activators can bind the allosteric drug and metabolite (ADaM) site between the KD in α-subunit and CBM in β-subunit [24]. ADaM site agonists can activate AMPK both by direct allosteric kinase activation or by protection from dephosphorylation. Adenine nucleotides are strongly present in the cells and are involved in many regulative processes and therefore are poorly selective as AMPK activators [52]. On the contrary, ADaM site activators can be more selective for AMPK and are promising as therapeutic drugs. However, their ineffective activation of β2-subunit, which is the predominant AMPK isoform in the human liver and skeletal muscle is a limitation of many ADaM site agonists [52].
A-769662 was the first molecule for AMPK direct activation developed by Abbott Laboratories in 2006. The development of this direct AMPK activator demonstrated that AMPK activation with non-nucleotide ligands is possible and stimulated to study new approaches for AMPK activation. A-769662 allosterically activates AMPK on the α subunit without Thr-172 phosphorylation [53]. Other direct AMPK activators, compound 991 [14] and MT 63–78 [54], are reported to be 5–10-fold more potent than A-769662. Furthermore, another compound, salicylate, shows structural similarity with A-769662 and binds at a site that overlaps with the A-769662 targeting site. Acetyl salicylate (aspirin) is a derivative that is easier to take orally than salicylate and rapidly breaks down to salicylate upon entering circulation [55]. All four compounds bind the ADaM site and exhibit specificity toward AMPKs that comprise β1 isoforms rather than the β2, giving the possibility to develop isoform-specific AMPK activators.
Discovering new direct AMPK activators appeared very promising in the reduction of lactic acidosis side effects. Vincent et al. compared 6 typical direct (salicylate, A-769662) and indirect (metformin, phenformin, AICAR, 2DG) AMPK agonists that affect cell proliferation. AMPK agonists showed contrasting effects on glycolytic metabolism. Metformin and phenformin increased both glucose consumption and lactate production, while AICAR and 2DG treatment reduced it. On the contrary, salicylate and A-769662 showed no significant changes. These results indicate that indirect AMPK activation may cause unfavorable side effects that can be avoided by direct AMPK activation [56].
Direct AMPK activators showed promising results in eliminating side effects observed from indirect AMPK activation. However, of note are few reports indicating that A-769662 can interfere with various biological pathways unrelated to AMPK through multiple off-target effects [57,58]. For example, it has been shown that A-769662 inhibits the function of the 26S proteasome [57] and Na+/K+-ATPase [58] activity by an AMPK-independent mechanism. This is calling into question the use of A-769662 as a specific AMPK activator. PT-1 is another compound that directly activates AMPK and promising results have shown increasing AMPK phosphorylation [59]. Unfortunately, besides binding directly to the AMPK α1 subunit, it also induces indirect activation of AMPK via inhibition of the mitochondrial respiratory chain complex, which increases cellular AMP:ATP or ADP:ATP ratios [60].
Another potent, direct, allosteric AMPK activator, MK-8722, can activate 12 mammalian AMPK complexes, mediating AMPK activation in skeletal muscle and inducing robust, durable, insulin-independent glucose uptake and glycogen synthesis, leading to chronically sustainable improvements in glucose homeostasis [61]. However, it has also been noted that it can induce reversible cardiac hypertrophy and increase cardiac glycogen [62]. Although increased cardiac glycogen content and hypertrophy were observed without any changes in electro-cardiogram and apparent functional cardiac sequelae, any safety issues associated with AMPK activators remain to be determined.
2.3. AMP Mimicking AMPK Activators
Direct AMPK activators are a good step forward compared to indirect activators; however, their safety aspects need to be considered. Due to this, particular attention is given to compounds that interact with AMPK in its AMP binding site, imitating physiological mechanisms of AMPK activation. Small molecules can mimic cellular AMP and trigger a conformational change and further activation of AMPK by Thr-172 phosphorylation at the α subunit without any significant change in cellular ATP, ADP, or AMP levels [63,64]. The binding of AMP to AMPK can regulate three processes in a synergistic manner: a) promotion of Thr-172 phosphorylation; b) protection against Thr-172 dephosphorylation, and c) allosteric activation of the phosphorylated kinase, making the final response very sensitive to even small changes in AMP [65]. However, very little literature can be found that discusses AMP mimicking compounds and the mechanisms of AMPK activation.
5-aminoimidazole-4-carboxamide riboside (AICAR) is considered to be an AMP mimetic, an adenosine analog taken up by the cell and phosphorylated by adenosine kinase to become ZMP (5-aminoimidazole-4-carboxamide ribonucleotide), which then mimics the activating effect of AMP on AMPK (without affecting the intracellular AMP:ATP ratio) [66]. ZMP is a competitive binder of the natural metabolite AMP and binds to the same CBS domains in AMPK without altering oxygen uptake and inhibiting mitochondrial function, like other AMPK activators. ZMP can accumulate to millimolar concentrations in cells and directly activates AMPK [66]. Even a long-term treatment with AICAR has shown to be beneficial; it enhances glucose tolerance, improves the lipid profile, and reduces systolic blood pressure in an insulin-resistant animal model [67].
Another AMPK activating compound, Cordycepin (30-deoxyadenosine), was reported to be able to enter cells via adenosine transporters. Cordycepin is a prodrug that activates AMPK by being converted by cellular metabolism into the AMP analog cordycepin monophosphate (CoMP) and, in cell-free assays, mimics all three effects of AMP on AMPK [68].
The exciting possibility of a safe ADP mimetic AMPK activator is encouraged by the recently reported by Steneberg et al. [69] successful completion of phase I trials for the novel small molecule O304. O304 is considered to be an ADP mimetic that protects pAMPK against dephosphorylation by the same mechanism as ADP without reducing cellular ATP. However, the exact site of O304 binding has not been reported yet. It is shown that O304, like AMPK activators PF-793 and MK-8722, increases glucose uptake in ex vivo skeletal myotubes in an AMPK-dependent but insulin-independent manner. In mice, O304 increases pAMPK levels and stimulates glucose uptake in skeletal muscle and potently reduces hyperglycemia, hyperinsulinemia, and insulin resistance without inducing cardiac hypertrophy [69].
Activator-3, a recently described AMP mimetic, is a potent AMPK activator that can significantly enhance glucose consumption, increase lipid profiles, reduce body weight, and negative metabolic impact of a high sucrose diet [70]. It interacts with R70 and R152 of the CBS1 domain of the γ subunit near the AMP binding site. A molecular modeling study showed that Activator-3 and AMP likely share a common AMPK activation mechanism. Moreover, it is shown to display a good pharmacokinetic profile in rat blood plasma with low brain penetration [70].
Screening of a chemical library of 1200 AMP mimetics has identified 5-(5-hydroxyl-isoxazol-3-yl)-furan-2-phosphonic acid, termed Compound-2 (C2), and its prodrug C13, as potent allosteric activators of AMPK [71], that has been reported to be >20-fold more potent than A-769662 [72] and more than two orders of magnitude more potent than AMP. Although the precise C2-binding site has not been identified yet, Hunter and colleagues [73] indicated that C2 competes with AMP for binding the γ subunit without causing any significant change in adenine nucleotide levels. However, some further studies suggest that C2 has a dual activation mechanism; Hunter et al. suggested that C2, unlike AICAR or ZMP, is an α1-selective AMPK activator.
Langedorf et al. have described the X-ray crystal structure of full-length α2β1γ1 isoform co-crystallized with C2 and AMP, where they revealed that two molecules of C2 can bind within γ-subunit. In this study, two distinct drug binding sites on AMPK have been identified, one located at the classic α-kinase domain/β-CBM (the so-called ADaM site, see above and Table 1) and the other one within the solvent-accessible core of the γ-subunit (named γ-pSite-1 and γ-pSite-4, respectively). The discovery of γ-subunit C2-binding sites represents a new unknown direction for drug design. This finding can be important for the development of synergistic activation of unphosphorylated α1-AMPK independent of AMP and upstream phosphorylation events [13].
Identification of different activation sites on AMPK raised the idea of dual AMPK targeting, which has been discussed in some studies demonstrating potential results. It has been shown that AMP mimetic compounds and allosteric activators in combination can produce a synergistic effect on AMPK phosphorylation and catalytic activity. Scott et al. have demonstrated that A-769662 and AMP can activate AMPK in a synergistic way (more than 1000-fold) [74]. C2 can synergize with A-769662 as well to activate dephosphorylated AMPK [13].
Ducommun et al. have analyzed the effect of direct AMPK activator A-769662 and the AMP mimetic AICAR, demonstrating their synergistic action on AMPK activation. It is suggested that ZMP binding to AMPK causes the conformational changes of the AMPK complex that facilitates A-769662 binding at β1 but also to β2 AMPK isoforms, which is not occurring when treated by these compounds alone. These data suggest that AMPK activation by two synergistic molecules is more advantageous and allows one to achieve a better physiological effect [75].
Unfortunately, the argument for dual AMPK activation is still poorly analyzed and described in the literature. This is mostly due to the fact that a lot of questions are still unanswered about the exact mechanism of AMPK regulation and activation by AMP. So it remains important to understand the complete composition of the AMP binding site and its mechanism of AMPK regulation to discover new potential compounds that can be used alone or in combination with other AMPK activators. Table 3 summarizes different characteristics of AMPK activators that can be useful for future investigations.
3. Gap of Knowledge for AMP Mimicking Drug Development
Although the various crystal structures obtained over the last few years have yielded considerable insight into the mechanism of AMPK regulation by adenine nucleotides, several questions remain.
As mentioned, AMPK senses adenine nucleotides level by AMP, ADP, and ATP binding to CBS1, CBS3, and CBS4 sites in its γ-subunit. However, it is still quite unknown how nucleotides bind to a single site, which nucleotides occupy each site under physiological conditions, and how binding to these sites interferes with each other. Mutagenesis studies demonstrate that CBS3 and CBS4 are important for AMPK allosteric activation. Mutation at I312D on CBS4 showed to inhibit direct allosteric AMPK activation by AMP [8]. It has been further suggested that CBS4 can sense AMP and is important for stabilizing AMP binding at CBS3. CBS4, therefore, allows AMP exchange at CBS3 at nucleotide concentrations that sense cellular energy conditions [52]. In another study, AMPK crystal structure analysis suggests that stabilization is predominantly due to CBS4 blocking the His298 side chain. The His298 backbone directly binds the phosphate group of AMP at CBS3 and blocks the near key residue Arg299, which forms two hydrogen bonds with the AMP phosphate group and two with the AMP adenine ring at CBS3. In addition, αRIM Glu364 blocks CBS3 Arg70 and Lys170, which forms salt bridges with the phosphate group of AMP at CBS3. The phosphate group of AMP carries two negative charges to interact with the three positively charged residues stabilized by αRIM, CBS4, and the partially positive His298 backbone [52]. Beside CBS repeats, α-RIM motives also play an essential role in AMPK regulation. It has been shown that mutating the key α-RIM1 residues largely inhibited the AMP dependence. Mutational analyses demonstrated that the three regulatory elements in the α-subunit, AID, α-RIM1, and α-RIM2, are indispensable for the allosteric activation of AMPK [15]; however, the full mechanism of this regulation remains to be analyzed. Detailed representation of this domain, as well as that of the ADaM site, in available structures, remains to be resolved, given the flexibility of these molecular regions.
Furthermore, it remains unclear AMP, ADP, and ATP differential effects on AMPK containing different γ1/γ2/γ3 isoforms. Unfortunately, most literature that describes full-length AMPK crystal structures studies mainly the γ1 isoform and explains poorly the exact roles of the three nucleotide-binding sites and their regulation mechanisms. The sequences of the CBS repeats in the γ1, γ2, and γ3 isoforms are highly conserved, but it seems possible that small differences of 1, 3, and 4 sites may influence nucleotides affinities and can be important in regulation. It is notable that γ2 and γ3 subunit isoforms have unrelated to each other N-terminal extensions of approximately 250 and 180 residues, respectively, which are not present in γ1. Although the functions of these N-terminal extensions are still uncertain, it is suggested that the N-terminal extensions may be essential for AMPK targeting at specific subcellular locations [88].
Furthermore, it would be important to understand how AMP binding inhibits Thr-172 dephosphorylation [65]. One of the described mechanism suggests that when AMP is bound to site 3, the γ-subunit create interactions with a few amino acids within the α-linkers α-RIM1 and α-RIM2, which interact with the unoccupied site 2 and the AMP molecule bound at site 3, respectively. The binding of the α-RIM motifs to the γ subunit obstructs the flexibility of the α-linker, resulting in stronger interaction of nucleotide-binding, which as a result protects Thr-172 from dephosphorylation. Interestingly, the same mechanism is expected when ADP binds to site 3, suggesting that in some conditions, ADP might be important for AMPK activation [89]. However, it is believed that AMP is a more sensitive controller of AMPK activity than ADP; some studies show that in certain conditions, ADP is a more important activator than AMP. Namely, Coccimiglio et al., in their publication, validated ADP and AMP binding by applying mathematical models suggesting that in skeletal muscle cells during exercise, AMPK activity dynamics are determined principally by ADP and not AMP [90].
It is also interesting what is the exact mechanism of ADP and AMP competition with ATP for γ subunit binding when the cellular ATP amount is usually much higher. A possible explanation for it can be that the Mg-ATP2− binds with a 10-fold lower affinity compared to free ATP4−. Main ATP in the cell is present as the Mg-ATP2− form and AMP, ADP may have to compete only with the free ATP4−, rather than with Mg-ATP2− complex [23].
4. Conclusions
The role of AMPK in maintaining homeostasis in cells and organs made it to be one of the important therapeutic targets in the treatment of different diseases such as obesity, inflammation, diabetes, and above all, cancer. Targeting AMPK studies have taken into account different generations of drug compounds, starting from those that activate AMPK indirectly to more specific direct AMPK activators that target AMPK at the ADaM site or mimic AMP. While indirect AMPK activators hold promise for the treatment of some diseases, it remains uncertain of the different effects of systemic and chronic AMPK activation. Moreover, the complexity of the different AMPK subunit isoforms combinations in tissues and their regulatory properties remains a challenge. In the last decades, significant progress has been made to understand the molecular mechanisms of AMPK regulation that should help develop more potent and specific drugs. Nowadays, the AMPK regulatory motive, the so-called ADaM site, is the most analyzed and discussed in the literature. Different compounds have been developed that target AMPK in this site, and some show promising results. In this review, however, we wanted to pay attention to the other important AMPK activation sites, namely AMP binding sites, their physiological activation site, and the potential for developing AMPK activation compounds that target exactly these sites. AMP binding sites are less analyzed in the literature and still, many questions remain about their exact regulation. Wide knowledge of both AMP and ADaM site mechanisms may facilitate the design of novel therapeutics as well as will open new opportunities in developing new dual synergistic AMPK activation treatment strategies.
Author Contributions
Conceptualization, analysis of literature, O.T., M.M., and A.D.D.; writing—original draft preparation, O.T., M.M., and A.D.D.; writing—review and editing, O.T., M.M., A.D.D. and G.N.; supervision, G.N.; project administration, G.N. All authors have read and agreed to the published version of the manuscript.
Funding
This reserch received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheung, P.C.; Salt, I.P.; Davies, S.P.; Hardie, D.G.; Carling, D. Characterization of AMP-activated protein kinase gamma-subunit isoforms and their role in AMP binding. Biochem. J. 2000, 346 Pt 3, 659–669. [Google Scholar] [CrossRef]
- Thornton, C.; Snowden, M.A.; Carling, D. Identification of a novel AMP-activated protein kinase beta subunit isoform that is highly expressed in skeletal muscle. J. Biol. Chem. 1998, 273, 12443–12450. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, D.; Gao, G.; Michell, B.J.; Widmer, J.; Mitchelhill, K.; Teh, T.; House, C.M.; Witters, L.A.; Kemp, B.E. Mammalian 5’-AMP-activated protein kinase non-catalytic subunits are homologs of proteins that interact with yeast Snf1 protein kinase. J. Biol. Chem. 1994, 269, 29343–29346. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Zhou, X.E.; Ke, J.; de Waal, P.W.; Gu, X.; Tan, M.H.; Wang, D.; Wu, D.; Xu, H.E.; et al. Structural basis of AMPK regulation by adenine nucleotides and glycogen. Cell Res. 2015, 25, 50–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, A.; Ghilagaber, S.; Nikolaev, A.; Hardie, D.G. The glycogen-binding domain on the AMPK beta subunit allows the kinase to act as a glycogen sensor. Cell Metab. 2009, 9, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A. The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem. Sci. 1997, 22, 12–13. [Google Scholar] [CrossRef]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nature 2011, 472, 230–233. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; Zhang, Y.Y.; Yan, S.F.; Neumann, D.; Schlattner, U.; Wang, Z.X.; Wu, J.W. AMP-activated protein kinase undergoes nucleotide-dependent conformational changes. Nat. Struct. Mol. Biol. 2012, 19, 716–718. [Google Scholar] [CrossRef]
- Yan, Y.; Mukherjee, S.; Harikumar, K.G.; Strutzenberg, T.S.; Zhou, X.E.; Suino-Powell, K.; Xu, T.H.; Sheldon, R.D.; Lamp, J.; Brunzelle, J.S.; et al. Structure of an AMPK complex in an inactive, ATP-bound state. Science 2021, 373, 413–419. [Google Scholar] [CrossRef]
- Calabrese, M.F.; Rajamohan, F.; Harris, M.S.; Caspers, N.L.; Magyar, R.; Withka, J.M.; Wang, H.; Borzilleri, K.A.; Sahasrabudhe, P.V.; Hoth, L.R.; et al. Structural basis for AMPK activation: Natural and synthetic ligands regulate kinase activity from opposite poles by different molecular mechanisms. Structure 2014, 22, 1161–1172. [Google Scholar] [CrossRef] [Green Version]
- Ngoei, K.R.W.; Langendorf, C.G.; Ling, N.X.Y.; Hoque, A.; Varghese, S.; Camerino, M.A.; Walker, S.R.; Bozikis, Y.E.; Dite, T.A.; Ovens, A.J.; et al. Structural Determinants for Small-Molecule Activation of Skeletal Muscle AMPK α2β2γ1 by the Glucose Importagog SC4. Cell Chem. Biol. 2018, 25, 728–737.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Zhou, X.E.; Novick, S.J.; Shaw, S.J.; Li, Y.; Brunzelle, J.S.; Hitoshi, Y.; Griffin, P.R.; Xu, H.E.; Melcher, K. Structures of AMP-activated protein kinase bound to novel pharmacological activators in phosphorylated, non-phosphorylated, and nucleotide-free states. J. Biol. Chem. 2019, 294, 953–967. [Google Scholar] [CrossRef] [Green Version]
- Langendorf, C.G.; Ngoei, K.R.W.; Scott, J.W.; Ling, N.X.Y.; Issa, S.M.A.; Gorman, M.A.; Parker, M.W.; Sakamoto, K.; Oakhill, J.S.; Kemp, B.E. Structural basis of allosteric and synergistic activation of AMPK by furan-2-phosphonic derivative C2 binding. Nat. Commun. 2016, 7, 10912. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Sanders, M.J.; Carmena, D.; Bright, N.J.; Haire, L.F.; Underwood, E.; Patel, B.R.; Heath, R.B.; Walker, P.A.; Hallen, S.; et al. Structural basis of AMPK regulation by small molecule activators. Nat. Commun. 2013, 4, 3017. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.J.; Wang, J.; Zhao, R.Q.; Wang, Z.X.; Wu, J.W. Coordinated regulation of AMPK activity by multiple elements in the α-subunit. Cell Res. 2013, 23, 1237–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, R.J.; Kosmatka, M.; Bardeesy, N.; Hurley, R.L.; Witters, L.A.; DePinho, R.A.; Cantley, L.C. The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc. Natl. Acad. Sci. USA 2004, 101, 3329–3335. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Pan, D.A.; Mustard, K.J.; Ross, L.; Bain, J.; Edelman, A.M.; Frenguelli, B.G.; Hardie, D.G. Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinase. Cell Metab. 2005, 2, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Neumann, D. Is TAK1 a Direct Upstream Kinase of AMPK? Int. J. Mol. Sci. 2018, 19, 2412. [Google Scholar] [CrossRef] [Green Version]
- Joseph, B.K.; Liu, H.Y.; Francisco, J.; Pandya, D.; Donigan, M.; Gallo-Ebert, C.; Giordano, C.; Bata, A.; Nickels, J.T. Inhibition of AMP Kinase by the Protein Phosphatase 2A Heterotrimer, PP2APpp2r2d. J. Biol. Chem. 2015, 290, 10588–10598. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.P.; Helps, N.R.; Cohen, P.T.; Hardie, D.G. 5’-AMP inhibits dephosphorylation, as well as promoting phosphorylation, of the AMP-activated protein kinase. Studies using bacterially expressed human protein phosphatase-2C alpha and native bovine protein phosphatase-2AC. FEBS Lett. 1995, 377, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.B.; Liu, Y.Y.; Cheng, L.B.; Lu, J.W.; Zeng, P.; Lu, P.H. AMPKα phosphatase Ppm1E upregulation in human gastric cancer is required for cell proliferation. Oncotarget 2017, 8, 31288–31296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willows, R.; Sanders, M.J.; Xiao, B.; Patel, B.R.; Martin, S.R.; Read, J.; Wilson, J.R.; Hubbard, J.; Gamblin, S.J.; Carling, D. Phosphorylation of AMPK by upstream kinases is required for activity in mammalian cells. Biochem. J. 2017, 474, 3059–3073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, D.G. AMP-activated protein kinase: An energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonora, M.; Patergnani, S.; Rimessi, A.; De Marchi, E.; Suski, J.M.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinergic Signal. 2012, 8, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fediuc, S.; Gaidhu, M.P.; Ceddia, R.B. Regulation of AMP-activated protein kinase and acetyl-CoA carboxylase phosphorylation by palmitate in skeletal muscle cells. J. Lipid Res. 2006, 47, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, S.; Zhai, A.; Zhang, B.; Tian, G. AMPK-Mediated Regulation of Lipid Metabolism by Phosphorylation. Biol. Pharm. Bull. 2018, 41, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.A.; Roach, W.G.; Keller, S.R.; Lane, W.S.; Lienhard, G.E. Inhibition of GLUT4 translocation by Tbc1d1, a Rab GTPase-activating protein abundant in skeletal muscle, is partially relieved by AMP-activated protein kinase activation. J. Biol. Chem. 2008, 283, 9187–9195. [Google Scholar] [CrossRef] [Green Version]
- Bolster, D.R.; Crozier, S.J.; Kimball, S.R.; Jefferson, L.S. AMP-activated protein kinase suppresses protein synthesis in rat skeletal muscle through down-regulated mammalian target of rapamycin (mTOR) signaling. J. Biol. Chem. 2002, 277, 23977–23980. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, S.; Bierhoff, H.; Cado, I.; Weber, A.; Tiebe, M.; Grummt, I.; Voit, R. AMP-activated protein kinase adapts rRNA synthesis to cellular energy supply. Proc. Natl. Acad. Sci. USA 2009, 106, 17781–17786. [Google Scholar] [CrossRef] [Green Version]
- Leprivier, G.; Remke, M.; Rotblat, B.; Dubuc, A.; Mateo, A.R.; Kool, M.; Agnihotri, S.; El-Naggar, A.; Yu, B.; Somasekharan, S.P.; et al. The eEF2 kinase confers resistance to nutrient deprivation by blocking translation elongation. Cell 2013, 153, 1064–1079. [Google Scholar] [CrossRef] [Green Version]
- Mizrachy-Schwartz, S.; Cohen, N.; Klein, S.; Kravchenko-Balasha, N.; Levitzki, A. Up-regulation of AMP-activated protein kinase in cancer cell lines is mediated through c-Src activation. J. Biol. Chem. 2011, 286, 15268–15277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamargo-Gómez, I.; Mariño, G. AMPK: Regulation of Metabolic Dynamics in the Context of Autophagy. Int. J. Mol. Sci. 2018, 19, 3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Egan, D.F.; Shackelford, D.B.; Mihaylova, M.M.; Gelino, S.; Kohnz, R.A.; Mair, W.; Vasquez, D.S.; Joshi, A.; Gwinn, D.M.; Taylor, R.; et al. Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science 2011, 331, 456–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Assefa, B.T.; Tafere, G.G.; Wondafrash, D.Z.; Gidey, M.T. The Bewildering Effect of AMPK Activators in Alzheimer’s Disease: Review of the Current Evidence. BioMed Res. Int. 2020, 2020, 9895121. [Google Scholar] [CrossRef]
- Hardie, D.G.; Alessi, D.R. LKB1 and AMPK and the cancer-metabolism link—ten years after. BMC Biol. 2013, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Boudeau, J.; Reid, J.L.; Mustard, K.J.; Udd, L.; Mäkelä, T.P.; Alessi, D.R.; Hardie, D.G. Complexes between the LKB1 tumor suppressor, STRAD alpha/beta and MO25 alpha/beta are upstream kinases in the AMP-activated protein kinase cascade. J. Biol. 2003, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.K.; Park, S.Y.; Kim, Y.M.; Park, O.J. Regulatory effect of the AMPK-COX-2 signaling pathway in curcumin-induced apoptosis in HT-29 colon cancer cells. Ann. N Y Acad. Sci. 2009, 1171, 489–494. [Google Scholar] [CrossRef]
- Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell 2005, 18, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Beckers, A.; Organe, S.; Timmermans, L.; Scheys, K.; Peeters, A.; Brusselmans, K.; Verhoeven, G.; Swinnen, J.V. Chemical inhibition of acetyl-CoA carboxylase induces growth arrest and cytotoxicity selectively in cancer cells. Cancer Res. 2007, 67, 8180–8187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Scheffler, T.L.; Gunawan, A.M.; Shi, H.; Zeng, C.; Hannon, K.M.; Grant, A.L.; Gerrard, D.E. Chronic elevated calcium blocks AMPK-induced GLUT-4 expression in skeletal muscle. Am. J. Physiol. Cell Physiol. 2009, 296, C106–C115. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, E. Metformin-Induced Mitochondrial Complex I Inhibition: Facts, Uncertainties, and Consequences. Front. Endocrinol. (Lausanne) 2018, 9, 753. [Google Scholar] [CrossRef] [PubMed]
- Bridges, H.R.; Jones, A.J.; Pollak, M.N.; Hirst, J. Effects of metformin and other biguanides on oxidative phosphorylation in mitochondria. Biochem. J. 2014, 462, 475–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrigal-Perez, L.A.; Ramos-Gomez, M. Resveratrol Inhibition of Cellular Respiration: New Paradigm for an Old Mechanism. Int. J. Mol. Sci. 2016, 17, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, D.G. AMP-activated protein kinase as a drug target. Annu. Rev. Pharm. Toxicol. 2007, 47, 185–210. [Google Scholar] [CrossRef]
- Vial, G.; Detaille, D.; Guigas, B. Role of Mitochondria in the Mechanism(s) of Action of Metformin. Front. Endocrinol. 2019, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- García Rubiño, M.E.; Carrillo, E.; Ruiz Alcalá, G.; Domínguez-Martín, A.; Marchal, J.A.; Boulaiz, H. Phenformin as an Anticancer Agent: Challenges and Prospects. Int. J. Mol. Sci. 2019, 20, 3316. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Vara-Ciruelos, D.; Russell, F.M.; Hardie, D.G. The strange case of AMPK and cancer: Dr Jekyll or Mr Hyde? Open Biol. 2019, 9, 190099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Bridges, M.D.; Yan, Y.; de Waal, P.W.; Zhou, X.E.; Suino-Powell, K.M.; Xu, H.E.; Hubbell, W.L.; Melcher, K. Conformational heterogeneity of the allosteric drug and metabolite (ADaM) site in AMP-activated protein kinase (AMPK). J. Biol. Chem. 2018, 293, 16994–17007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, M.J.; Ali, Z.S.; Hegarty, B.D.; Heath, R.; Snowden, M.A.; Carling, D. Defining the mechanism of activation of AMP-activated protein kinase by the small molecule A-769662, a member of the thienopyridone family. J. Biol. Chem. 2007, 282, 32539–32548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadra, G.; Photopoulos, C.; Tyekucheva, S.; Heidari, P.; Weng, Q.P.; Fedele, G.; Liu, H.; Scaglia, N.; Priolo, C.; Sicinska, E.; et al. A novel direct activator of AMPK inhibits prostate cancer growth by blocking lipogenesis. EMBO Mol. Med. 2014, 6, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Dandapani, M.; Hardie, D.G. AMPK: Mediating the metabolic effects of salicylate-based drugs? Trends Endocrinol. Metab. 2013, 24, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, E.E.; Coelho, P.P.; Blagih, J.; Griss, T.; Viollet, B.; Jones, R.G. Differential effects of AMPK agonists on cell growth and metabolism. Oncogene 2015, 34, 3627–3639. [Google Scholar] [CrossRef] [Green Version]
- Moreno, D.; Knecht, E.; Viollet, B.; Sanz, P. A769662, a novel activator of AMP-activated protein kinase, inhibits non-proteolytic components of the 26S proteasome by an AMPK-independent mechanism. FEBS Lett. 2008, 582, 2650–2654. [Google Scholar] [CrossRef] [Green Version]
- Benziane, B.; Björnholm, M.; Lantier, L.; Viollet, B.; Zierath, J.R.; Chibalin, A.V. AMP-activated protein kinase activator A-769662 is an inhibitor of the Na(+)-K(+)-ATPase. Am. J. Physiol. Cell Physiol. 2009, 297, C1554–C1566. [Google Scholar] [CrossRef] [Green Version]
- Pang, T.; Zhang, Z.S.; Gu, M.; Qiu, B.Y.; Yu, L.F.; Cao, P.R.; Shao, W.; Su, M.B.; Li, J.Y.; Nan, F.J.; et al. Small molecule antagonizes autoinhibition and activates AMP-activated protein kinase in cells. J. Biol. Chem. 2008, 283, 16051–16060. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.E.; Ross, F.A.; Kleinert, M.; Sylow, L.; Knudsen, J.R.; Gowans, G.J.; Hardie, D.G.; Richter, E.A. PT-1 selectively activates AMPK-γ1 complexes in mouse skeletal muscle, but activates all three γ subunit complexes in cultured human cells by inhibiting the respiratory chain. Biochem. J. 2015, 467, 461–472. [Google Scholar] [CrossRef]
- Feng, D.; Biftu, T.; Romero, F.A.; Kekec, A.; Dropinski, J.; Kassick, A.; Xu, S.; Kurtz, M.M.; Gollapudi, A.; Shao, Q.; et al. Discovery of MK-8722: A Systemic, Direct Pan-Activator of AMP-Activated Protein Kinase. ACS Med. Chem. Lett. 2018, 9, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.W.; Guan, H.P.; Ehrhart, J.; Petrov, A.; Prahalada, S.; Tozzo, E.; Yang, X.; Kurtz, M.M.; Trujillo, M.; Gonzalez Trotter, D.; et al. Systemic pan-AMPK activator MK-8722 improves glucose homeostasis but induces cardiac hypertrophy. Science 2017, 357, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, S.A.; Davison, M.; Woods, A.; Davies, S.P.; Beri, R.K.; Carling, D.; Hardie, D.G. Characterization of the AMP-activated protein kinase kinase from rat liver and identification of threonine 172 as the major site at which it phosphorylates AMP-activated protein kinase. J. Biol. Chem. 1996, 271, 27879–27887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, S.C.; Woods, A.; Jones, N.A.; Davison, M.D.; Carling, D. The regulation of AMP-activated protein kinase by phosphorylation. Biochem. J. 2000, 345 Pt 3, 437–443. [Google Scholar] [CrossRef]
- Grahame Hardie, D. Regulation of AMP-activated protein kinase by natural and synthetic activators. Acta Pharm. Sin. B 2016, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corton, J.M.; Gillespie, J.G.; Hawley, S.A.; Hardie, D.G. 5-aminoimidazole-4-carboxamide ribonucleoside. A specific method for activating AMP-activated protein kinase in intact cells? Eur. J. Biochem. 1995, 229, 558–565. [Google Scholar] [CrossRef]
- Buhl, E.S.; Jessen, N.; Pold, R.; Ledet, T.; Flyvbjerg, A.; Pedersen, S.B.; Pedersen, O.; Schmitz, O.; Lund, S. Long-term AICAR administration reduces metabolic disturbances and lowers blood pressure in rats displaying features of the insulin resistance syndrome. Diabetes 2002, 51, 2199–2206. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Ross, F.A.; Russell, F.M.; Atrih, A.; Lamont, D.J.; Hardie, D.G. Mechanism of Activation of AMPK by Cordycepin. Cell Chem. Biol. 2020, 27, 214–222.e4. [Google Scholar] [CrossRef] [Green Version]
- Steneberg, P.; Lindahl, E.; Dahl, U.; Lidh, E.; Straseviciene, J.; Backlund, F.; Kjellkvist, E.; Berggren, E.; Lundberg, I.; Bergqvist, I.; et al. PAN-AMPK activator O304 improves glucose homeostasis and microvascular perfusion in mice and type 2 diabetes patients. JCI Insight 2018, 3, e99114. [Google Scholar] [CrossRef]
- Bung, N.; Surepalli, S.; Seshadri, S.; Patel, S.; Peddasomayajula, S.; Kummari, L.K.; Kumar, S.T.; Babu, P.P.; Parsa, K.V.L.; Poondra, R.R.; et al. 2-[2-(4-(trifluoromethyl)phenylamino)thiazol-4-yl]acetic acid (Activator-3) is a potent activator of AMPK. Sci. Rep. 2018, 8, 9599. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Galeno, J.E.; Dang, Q.; Nguyen, T.H.; Boyer, S.H.; Grote, M.P.; Sun, Z.; Chen, M.; Craigo, W.A.; van Poelje, P.D.; MacKenna, D.A.; et al. A Potent and Selective AMPK Activator That Inhibits de Novo Lipogenesis. ACS Med. Chem. Lett. 2010, 1, 478–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cool, B.; Zinker, B.; Chiou, W.; Kifle, L.; Cao, N.; Perham, M.; Dickinson, R.; Adler, A.; Gagne, G.; Iyengar, R.; et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006, 3, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, R.W.; Foretz, M.; Bultot, L.; Fullerton, M.D.; Deak, M.; Ross, F.A.; Hawley, S.A.; Shpiro, N.; Viollet, B.; Barron, D.; et al. Mechanism of action of compound-13: An α1-selective small molecule activator of AMPK. Chem. Biol. 2014, 21, 866–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.W.; Ling, N.; Issa, S.M.; Dite, T.A.; O’Brien, M.T.; Chen, Z.P.; Galic, S.; Langendorf, C.G.; Steinberg, G.R.; Kemp, B.E.; et al. Small molecule drug A-769662 and AMP synergistically activate naive AMPK independent of upstream kinase signaling. Chem. Biol. 2014, 21, 619–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducommun, S.; Ford, R.J.; Bultot, L.; Deak, M.; Bertrand, L.; Kemp, B.E.; Steinberg, G.R.; Sakamoto, K. Enhanced activation of cellular AMPK by dual-small molecule treatment: AICAR and A769662. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E688–E696. [Google Scholar] [CrossRef] [Green Version]
- Dunn, D.M.; Munger, J. Interplay Between Calcium and AMPK Signaling in Human Cytomegalovirus Infection. Front. Cell Infect. Microbiol. 2020, 10, 384. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Park, W.H.; You, B.R. Antimycin A induces death of the human pulmonary fibroblast cells via ROS increase and GSH depletion. Int. J. Oncol. 2016, 48, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Giaccari, A.; Solini, A.; Frontoni, S.; Del Prato, S. Metformin Benefits: Another Example for Alternative Energy Substrate Mechanism? Diabetes Care 2021, 44, 647–654. [Google Scholar] [CrossRef]
- Smith, R.M.; Peterson, W.H.; Mccoy, E. Oligomycin, a new antifungal antibiotic. Antibiot. Chemother. 1954, 4, 962–970. [Google Scholar]
- Kobayashi, K.; Nishino, C.; Ohya, J.; Sato, S.; Mikawa, T.; Shiobara, Y.; Kodama, M.; Nishimoto, N. Oligomycin E, a new antitumor antibiotic produced by Streptomyces sp. MCI-2225. J. Antibiot. 1987, 40, 1053–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, A.D.; Balteau, M.; Gélinas, R.; Renguet, E.; Ginion, A.; de Meester, C.; Sakamoto, K.; Balligand, J.L.; Bontemps, F.; Vanoverschelde, J.L.; et al. A-769662 potentiates the effect of other AMP-activated protein kinase activators on cardiac glucose uptake. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1619–H1630. [Google Scholar] [CrossRef] [Green Version]
- Bultot, L.; Jensen, T.E.; Lai, Y.C.; Madsen, A.L.; Collodet, C.; Kviklyte, S.; Deak, M.; Yavari, A.; Foretz, M.; Ghaffari, S.; et al. Benzimidazole derivative small-molecule 991 enhances AMPK activity and glucose uptake induced by AICAR or contraction in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E706–E719. [Google Scholar] [CrossRef] [Green Version]
- Yeasmin, F.; Choi, H.W. Natural Salicylates and Their Roles in Human Health. Int. J. Mol. Sci. 2020, 21, 9049. [Google Scholar] [CrossRef] [PubMed]
- Višnjić, D.; Lalić, H.; Dembitz, V.; Tomić, B.; Smoljo, T. AICAr, a Widely Used AMPK Activator with Important AMPK-Independent Effects: A Systematic Review. Cells 2021, 10, 1095. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, M.; Steneberg, P.; Nyrén, R.; Edlund, H. AMPK activator O304 improves metabolic and cardiac function, and exercise capacity in aged mice. Commun. Biol. 2021, 4, 1306. [Google Scholar] [CrossRef]
- Ross, F.A.; Jensen, T.E.; Hardie, D.G. Differential regulation by AMP and ADP of AMPK complexes containing different γ subunit isoforms. Biochem. J. 2016, 473, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Coccimiglio, I.F.; Clarke, D.C. ADP is the dominant controller of AMP-activated protein kinase activity dynamics in skeletal muscle during exercise. PLoS Comput. Biol. 2020, 16, e1008079. [Google Scholar] [CrossRef]
Figure 2.
CBS binding sites of the γ subunit. Key binding residues are noted in green for CBS1, in red for CBS3, and in blue for CBS4. Tyr-342, pictured in red, belongs to the AID domain of the α subunit.
Figure 2.
CBS binding sites of the γ subunit. Key binding residues are noted in green for CBS1, in red for CBS3, and in blue for CBS4. Tyr-342, pictured in red, belongs to the AID domain of the α subunit.

Figure 3.
Displacement of the kinase domain (KD) from the associated, active (AMP/ADP bound) conformation. Disassociation is triggered by the substitution of an AMP/ADP residue with Mg-ATP2− one: this brings to a steric clash with the protecting αRIM2 loop, favoring the disassociation of the KD. Phosphorylated Thr-174 (Thr-172 in α2) thus finds itself in an unprotected state, allowing its subsequent dephosphorylation, which in turn inactivates AMPK [4,7,9]. The structures used were 4RER [4] for the associated, active state and 7JHG [9] for the disassociated, inactive state.
Figure 3.
Displacement of the kinase domain (KD) from the associated, active (AMP/ADP bound) conformation. Disassociation is triggered by the substitution of an AMP/ADP residue with Mg-ATP2− one: this brings to a steric clash with the protecting αRIM2 loop, favoring the disassociation of the KD. Phosphorylated Thr-174 (Thr-172 in α2) thus finds itself in an unprotected state, allowing its subsequent dephosphorylation, which in turn inactivates AMPK [4,7,9]. The structures used were 4RER [4] for the associated, active state and 7JHG [9] for the disassociated, inactive state.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Residue numbers for each highlighted domain.
| Subunit | Domain | Residues (aa) | Color Code (Figure 1) |
|---|---|---|---|
| Alfa | Kinase Domain | 11–281 [4] | Orange |
| AID | 282–353 [4] | Red | |
| RIMST loop (black dashed line) | 354–392 [4] 477–528 [9] | Blue | |
| Beta | GBM | 75–157 [4] | Bright green |
| Allosteric drug and metabolite binding site—ADaM site (see below in the text—not completely resolved) | 162–171 [10] | n/a |
Table 2.
Experimentally determined 3D structures of human AMPK, including the three α, β, and γ subunits, are available (May 2022) from the RCSB Protein Data Bank (https://www.rcsb.org/—accessed on 28 June 2022 ).
Table 2.
Experimentally determined 3D structures of human AMPK, including the three α, β, and γ subunits, are available (May 2022) from the RCSB Protein Data Bank (https://www.rcsb.org/—accessed on 28 June 2022 ).
| PDB ID | Title | Subunits | Method | Release Date | Reference |
|---|---|---|---|---|---|
| 7JHG | Cryo-EM structure of ATP-bound fully inactive AMPK in complex with Dorsomorphin (Compound C) and Fab-nanobody | α1; β2; γ1 | Cryo-EM | 2021 | [9] |
| 7JHH | Cryo-EM structure of ATP-bound fully inactive AMPK in complex with Fab and nanobody | α1; β2; γ1 | Cryo-EM | 2021 | [9] |
| 7JIJ | ATP-bound AMP-activated protein kinase | α1; β2; γ1 | X-ray diffraction | 2021 | [9] |
| 7M74 | ATP-bound AMP-activated protein kinase | α1; β2; γ1 | Cryo-EM | 2021 | [9] |
| 6B2E | Structure of full-length human AMPK (a2b2g1) in complex with a small molecule activator SC4 | α2; β2; γ1 | X-ray diffraction | 2018 | [11] |
| 6B1U | Structure of full-length human AMPK (a2b2g1) in complex with a small molecule activator SC4 | α2; β1; γ1 | X-ray diffraction | 2018 | [11] |
| 6C9F | AMP-activated protein kinase bound to pharmacological activator R734 | α1; β1; γ1 | X-ray diffraction | 2018 | [12] |
| 6C9G | AMP-activated protein kinase bound to pharmacological activator R739 | α1; β1; γ1 | X-ray diffraction | 2018 | [12] |
| 6C9H | Non-phosphorylated AMP-activated protein kinase bound to pharmacological activator R734 | α1; β1; γ1 | X-ray diffraction | 2018 | [12] |
| 6C9J | AMP-activated protein kinase bound to pharmacological activator R734 | α1; β1; γ1 | X-ray diffraction | 2018 | [12] |
| 5ISO | Structure of full-length human AMPK (non-phosphorylated at T-loop) in complex with a small molecule activator, a benzimidazole derivative (991) | α2; β1; γ1 | X-ray diffraction | 2017 | to be published |
| 5EZV | X-ray crystal structure of AMP-activated protein kinase alpha-2/alpha-1 RIM chimaera (alpha-2(1–347)/alpha-1(349–401)/alpha-2(397-end) beta-1 gamma-1) co-crystallized with C2 (5-(5-hydroxyl-isoxazol-3-yl)-furan-2-phosphonic acid) | α2/α1; β1; γ1 | X-ray diffraction | 2016 | [13] |
| 4ZHX | Novel binding site for allosteric activation of AMPK | α2; β1; γ1 | X-ray diffraction | 2016 | [13] |
| 4RER | Crystal structure of the phosphorylated human alpha1 beta2 gamma1 holo-AMPK complex bound to AMP and cyclodextrin | α1; β2; γ1 | X-ray diffraction | 2014 | [4] |
| 4REW | Crystal structure of the non-phosphorylated human alpha1 beta2 gamma1 holo-AMPK complex | α1; β2; γ1 | X-ray diffraction | 2014 | [4] |
| 4CFE | Structure of full-length human AMPK in complex with a small molecule activator, a benzimidazole derivative (991) | α2; β1; γ1 | X-ray diffraction | 2013 | [14] |
| 4CFF | Structure of full-length human AMPK in complex with a small molecule activator, a thienopyridone derivative (A-769662) | α2; β1; γ1 | X-ray diffraction | 2013 | [14] |
Table 3.
Summary and characteristics of AMPK activating compounds.
| Mechanism of action | Drug name | Treatment | Disadvantage | Advantage | Ref. | |
|---|---|---|---|---|---|---|
| Indirect AMPK activation | intracellular accumulation of Ca2+ | upstream regulation of (CaMKK2) | calcium-AMPK signaling regulates Human Cytomegalovirus (HCMV) infection | disruption of Ca2+ balance can lead to various side effects. | Ca2+ plays an essential role in regulating many signaling pathways and cellular processes, such as cell growth and differentiation | [76,77] |
| inhibit mitochondrial ATP synthesis by inhibiting the respiratory chain | antimycin A | anti-tumoral | lack of specificity, quite toxic for normal cells | therapeutic advantage for the treatment of tumors | [78] | |
| metformin | type II diabetes | gastrointestinal side effects (diarrhea, nausea, abdominal discomfort) | glucose-lowering efficacy, modest body weight reduction, easy combination with almost any other glucose-lowering agent | [79] | ||
| phenformin | type II diabetes | develops life-threatening cases of lactic acidosis | potential effect on cancer treatment | [49] | ||
| oligomycin | anti-fungal, anti-tumoral | high lactate accumulating in the blood, urine | therapeutic advantage for the treatment of tumors | [80,81] | ||
| resveratrol | anti-inflammatory, anti-oxidative, antitumoral, neurological, cardiovascular diseases, diabetes, NAFLD, obesity | nausea, vomiting, diarrhea, and liver dysfunction in patients with NAFLD | high range of treatment applications, may promote heart health, lower cholesterol, promote brain health, slow cancer growth | [50] | ||
| Direct AMPK activation | ADaM site | A-769662 | cardiovascular disorders | can have few off-target effects, ineffective activation of β2-subunit | reduction of lactic acidosis, reduces infarct size, allows a better recovery of contractile function during reperfusion | [82] |
| ADaM site | compound 991 | skeletal muscle glucose uptake, type II diabetes, obesity | activate β1-isoform 10 times stronger than β2 | 5-10-fold more potent than A-769662 in activating AMPK, minimal side effects | [83] | |
| ADaM site | MT 63–78 | anti-tumoral | low-affinity binding to β2 subunit | effective at low concentration | [54] | |
| ADaM site | salicylate | relieve pain and inflammation, reduce fever, prevent excessive blood clotting | difficult breathing, diarrhea, nausea, vomiting | lower risks of cancer, heart disease, and diabetes | [84] | |
| contradictive information | PT-1 | lower hepatic lipid content, type II diabetes, obesity | selective for γ1 and not γ3 isoform | promising AMPK activator, minimal side effects | [59,60] | |
| ADAM site | MK-8722 | increase glucose uptake into skeletal muscle, type II diabetes | induce reversible cardiac hypertrophy and increase cardiac glycogen | activate 12 AMPK complexes, induce robust, durable, insulin-independent glucose uptake and glycogen synthesis | [62] | |
| AMPK mimicking compounds | phosphorylated by adenosine kinase to ZMP, binds the same CBS domains as AMP | AICAR | anti-inflammatory, skeletal muscle glucose uptake, cardiovascular diseases | poor oral bioavailability, may have AMPK-independent effects | long-term treatment without side effects, reduce myocardial infarction | [85] |
| AMP analog CoMP binds γ1 subunit | Cordycepin | anti-tumoral, type II diabetes, obesity, anti-fungal, anti-inflammatory, antioxidant, anti-aging, antiviral, hepato-protective | mild gastrointestinal side effects | structure similarity with adenosine makes it an important bioactive component, a wide variety of positive effect | [68,86] | |
| mimics ADP, suppresses pAMPK. dephosphorylation | O304 | type II diabetes, obesity, cardiovascular diseases, peripheral microvascular perfusion | no particular side effects in clinical trials | reduces hyperglycemia, and hyperinsulinemia without inducing cardiac hypertrophy, mimics the beneficial effects of exercise, shows good safety | [87] | |
| interacts with R70 and R152 of the CBS1 domain on γ-subunit near AMP binding site. | Activator-3 | type II diabetes, obesity | mode of activation of Activator-3 is not completely understood | significantly enhance glucose consumption, increase lipid profiles, good pharmacokinetic profile in blood plasma, low brain penetration | [70] | |
| bind γ -subunit near AMP binding site | Compound-2 | metabolic disorders, obesity, and type II diabetes | selectivity for α1 rather than α2 subunit | >20-fold more potent than A-769662 and more than two orders of magnitude more potent than AMP, does not affect any of other AMP activating enzymes | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tarasiuk, O.; Miceli, M.; Di Domizio, A.; Nicolini, G. AMPK and Diseases: State of the Art Regulation by AMPK-Targeting Molecules. Biology 2022, 11, 1041. https://doi.org/10.3390/biology11071041
AMA Style
Tarasiuk O, Miceli M, Di Domizio A, Nicolini G. AMPK and Diseases: State of the Art Regulation by AMPK-Targeting Molecules. Biology. 2022; 11(7):1041. https://doi.org/10.3390/biology11071041
Chicago/Turabian StyleTarasiuk, Olga, Matteo Miceli, Alessandro Di Domizio, and Gabriella Nicolini. 2022. "AMPK and Diseases: State of the Art Regulation by AMPK-Targeting Molecules" Biology 11, no. 7: 1041. https://doi.org/10.3390/biology11071041
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.