
Impact of the “Flavescence Dorée” Phytoplasma on Xylem Growth and Anatomical Characteristics in Trunks of ‘Chardonnay’ Grapevines (Vitis vinifera)

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
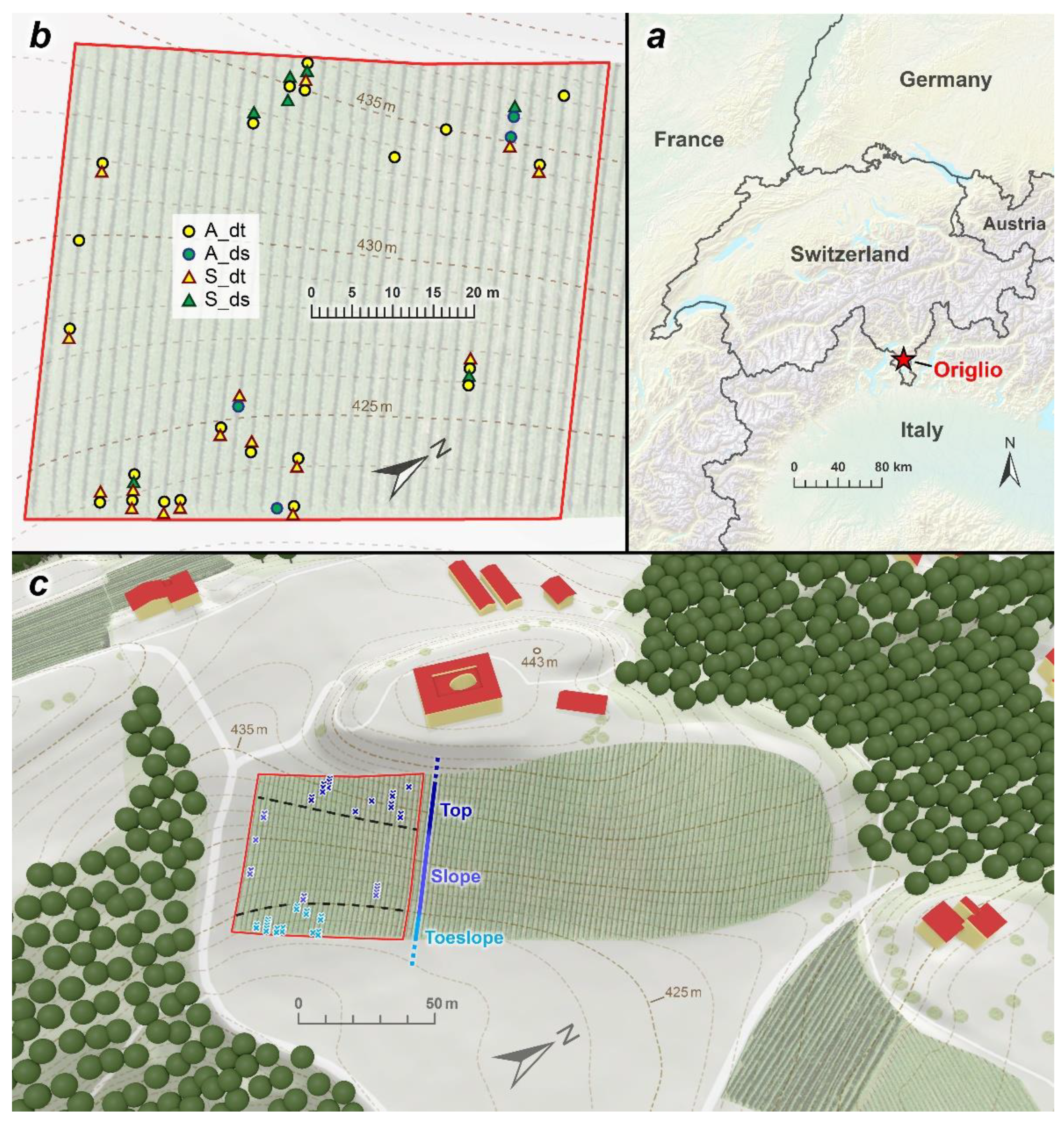
2.1. Study Area and Sampling Design
2.2. Nucleic Acid Extraction
2.3. FDp Detection
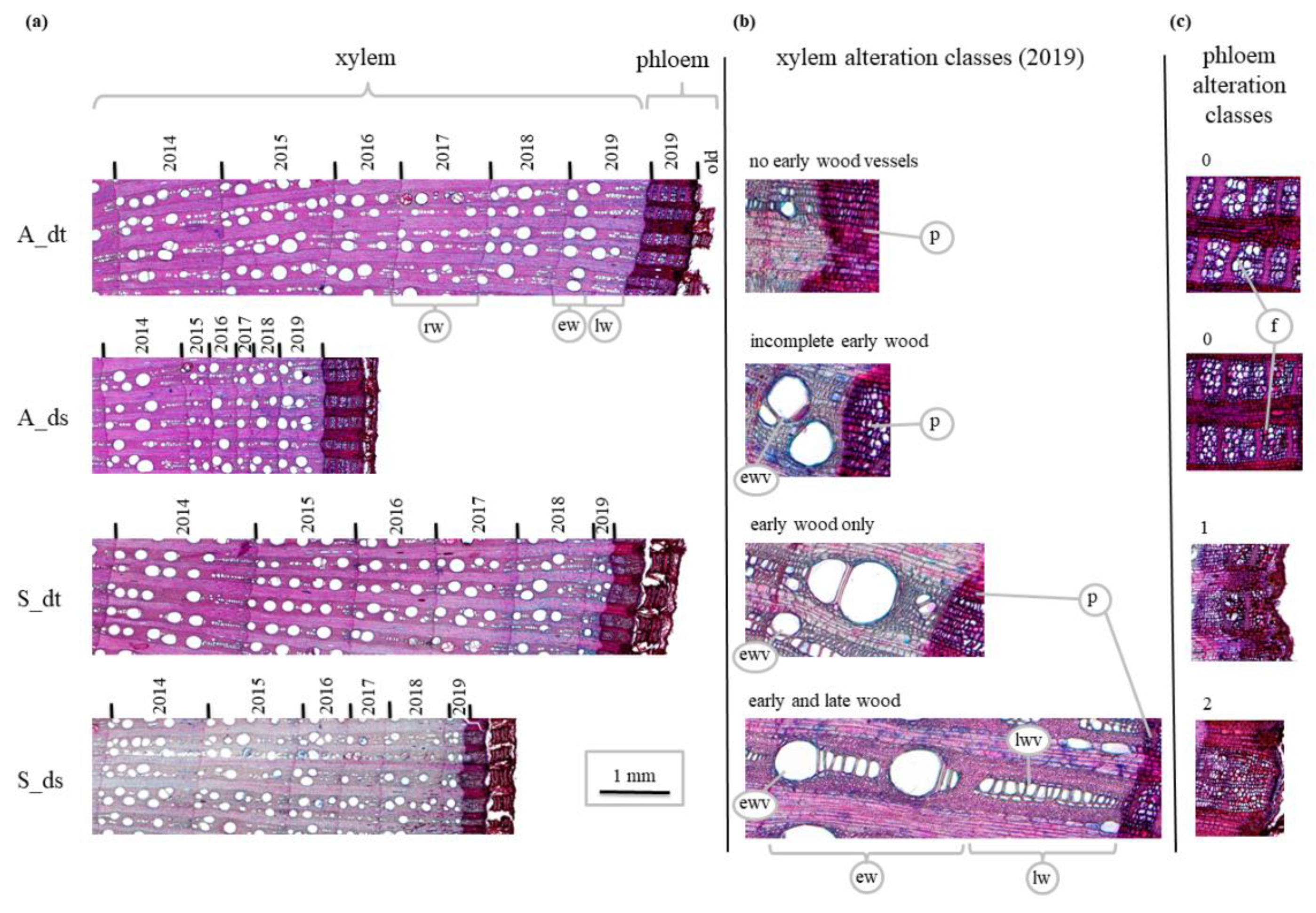
2.4. Core Samples’ Collection and Preparation
2.5. Image Capturing and Processing
2.6. Data Analysis
2.6.1. Assessment of Anatomical Anomalies
2.6.2. Measurement and Standardisation of Annual Ring Widths
2.6.3. Climatic Impact on Growth and Definition of Drought Years
2.6.4. Detection of Drought-Sensitive Specimens
2.6.5. Factors Influencing Xylem Growth Responses
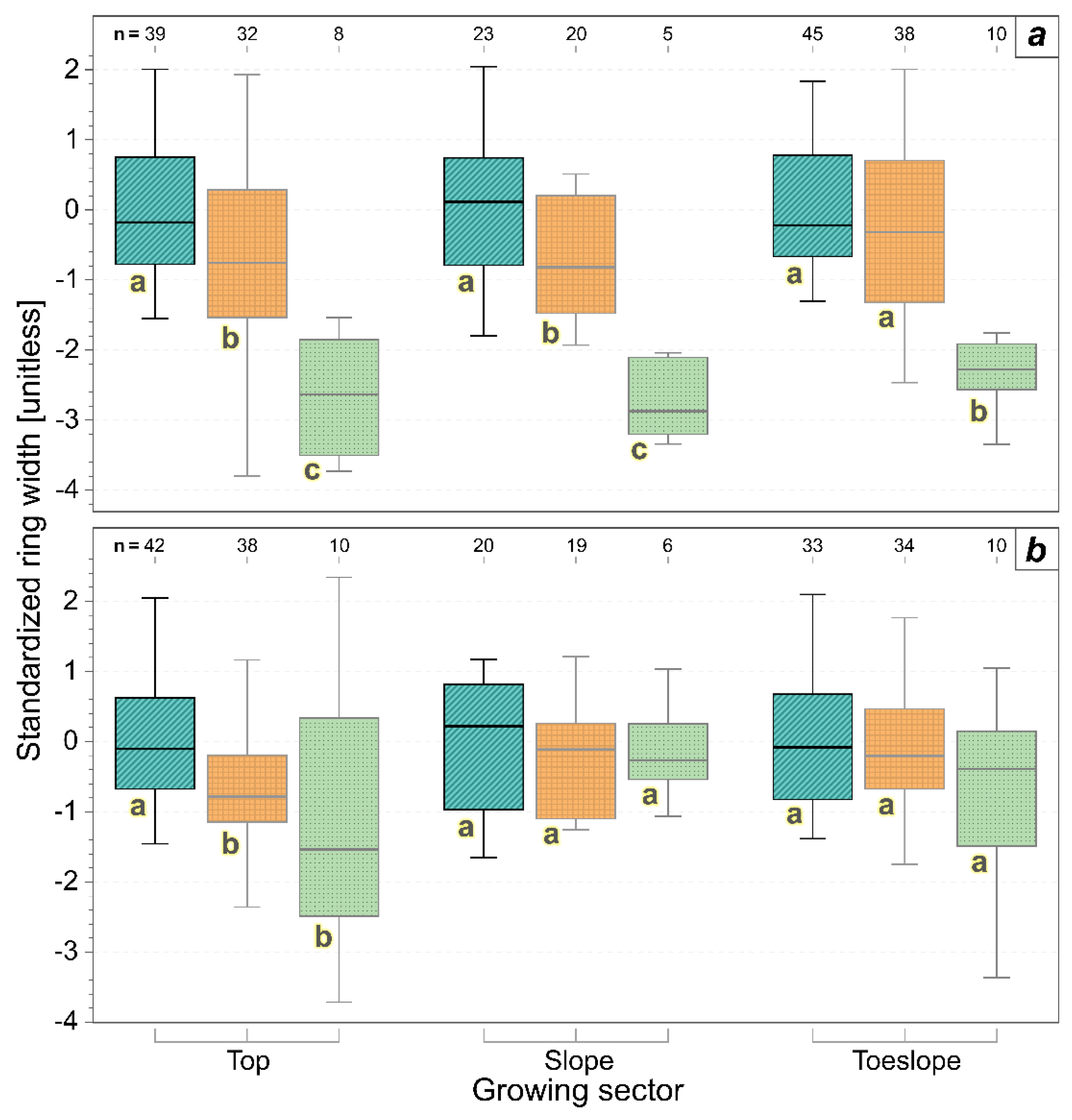
3. Results
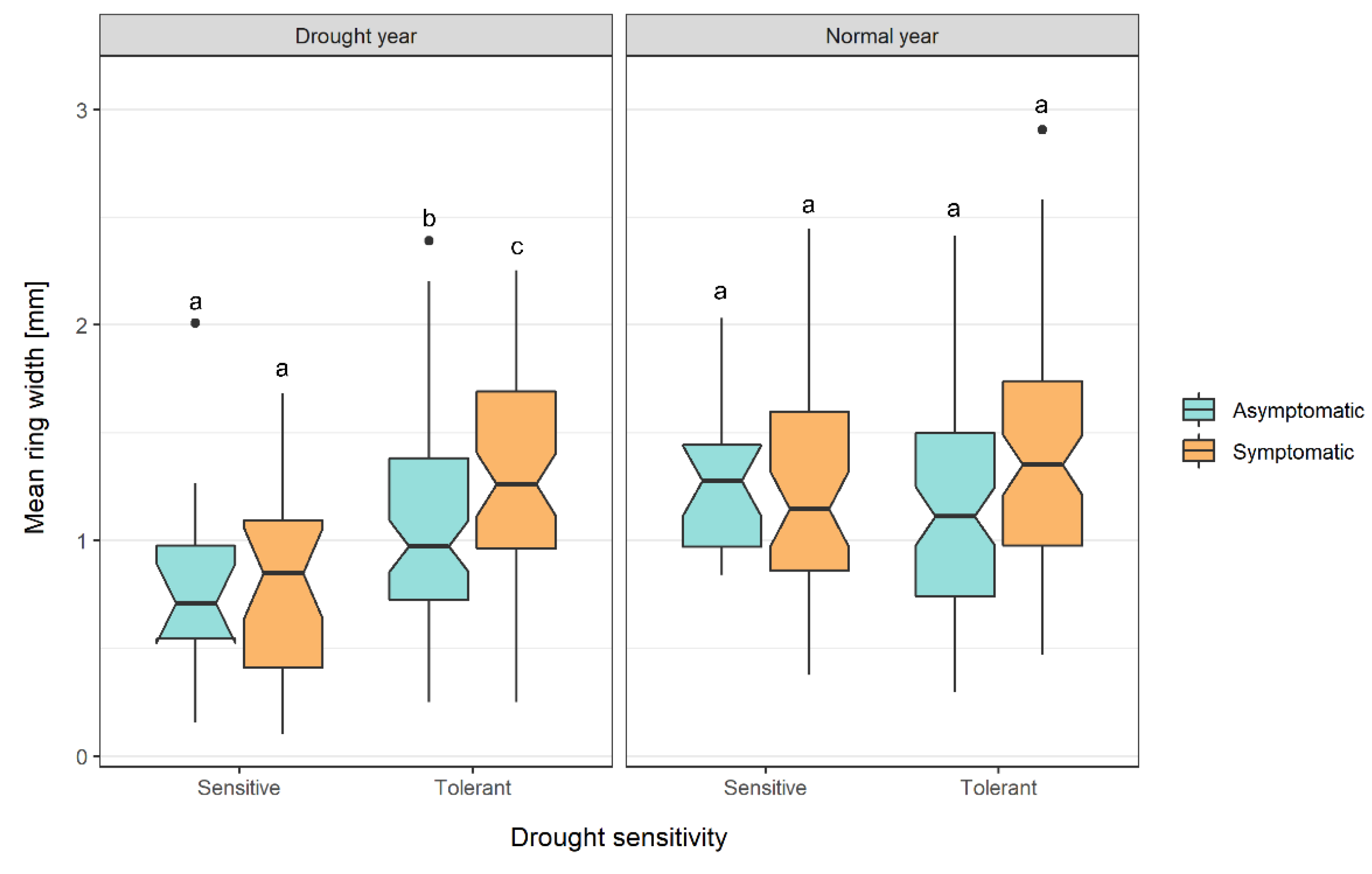
3.1. Drought Years
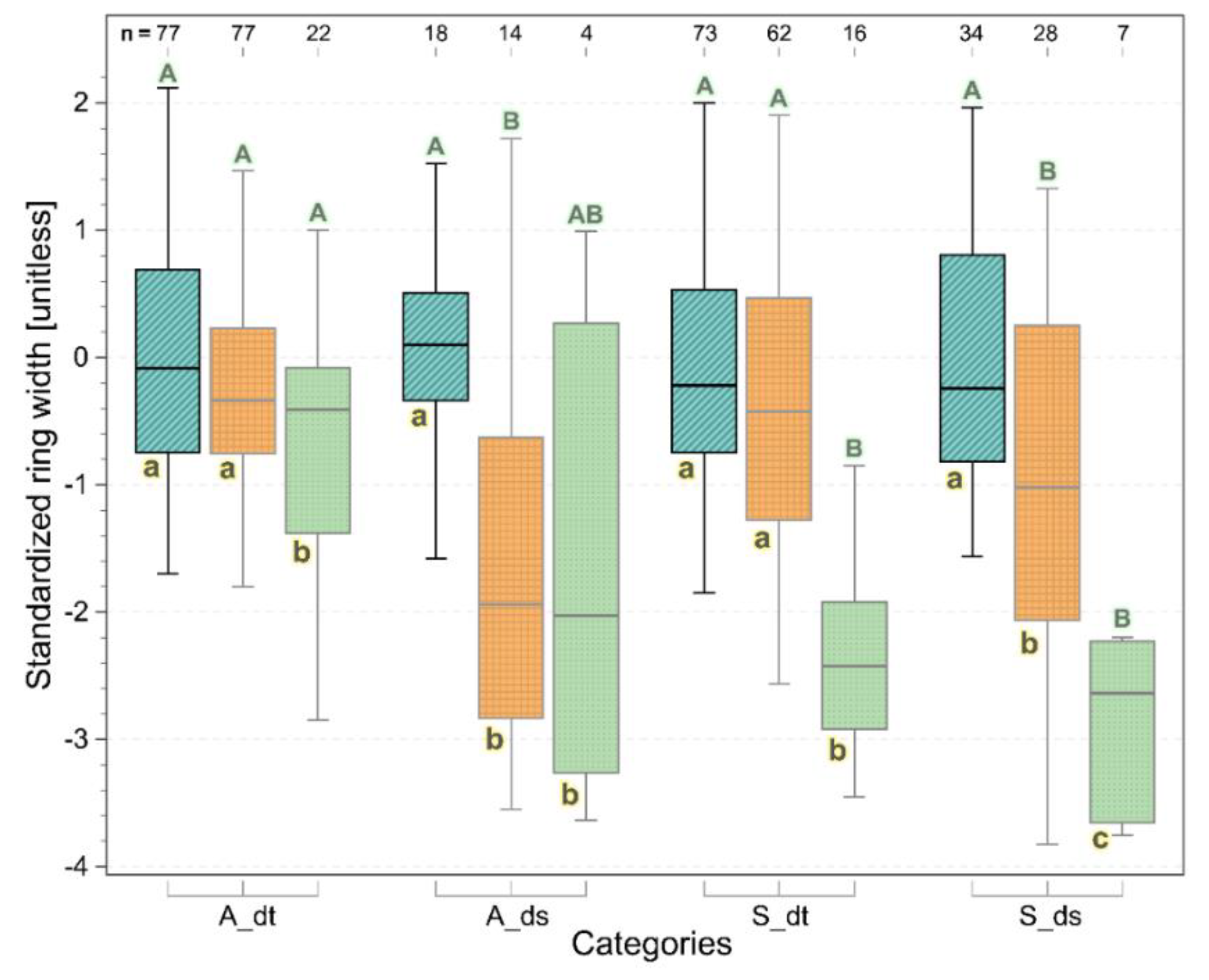
3.2. Annual Xylem Increments and Phloem Anomalies
3.3. Drivers of Xylem Increments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Firrao, G.; Marcone, C.; Bertaccini, A. Phytoplasma Classification. In Proceedings of the 11th SIPaV Annual Meeting, Milano, Italy, 29 September–1 October 2004; p. 299. [Google Scholar]
- Bertaccini, A.; Duduk, B. Phytoplasma and Phytoplasma Diseases: A Review of Recent Research. Phytopathol. Mediterr. 2009, 48, 355–378. [Google Scholar] [CrossRef]
- Tramontini, S.; Delbianco, A.; Vos, S. Pest Survey Card on Flavescence Dorée Phytoplasma and Its Vector Scaphoideus titanus. EFSA Supporting Publ. 2020, 17, 1–36. [Google Scholar] [CrossRef]
- Bressan, A.; Larrue, J.; Boudon-Padieu, E. Patterns of Phytoplasma-Infected and Infective Scaphoideus titanus Leafhoppers in Vineyards with High Incidence of Flavescence Doree. Entomol. Exp. Appl. 2006, 119, 61–69. [Google Scholar] [CrossRef]
- Chuche, J.; Thiéry, D. Biology and Ecology of the Flavescence Dorée Vector Scaphoideus titanus: A Review. Agron. Sustain. Dev. 2014, 34, 381–403. [Google Scholar] [CrossRef] [Green Version]
- EPPO/CABI. Quarantine Pests for Europe, 2nd ed.; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Sugio, A.; MacLean, A.M.; Kingdom, H.N.; Grieve, V.M.; Manimekalai, R.; Hogenhout, S.A. Diverse Targets of Phytoplasma Effectors: From Plant Development to Defense Against Insects. Annu. Rev. Phytopathol. 2011, 49, 175–195. [Google Scholar] [CrossRef]
- Namba, S. Molecular and Biological Properties of Phytoplasmas. Proc. Jpn. Academy. Ser. B Phys. Biol. Sci. 2019, 95, 401–418. [Google Scholar] [CrossRef] [Green Version]
- van Bel, A.J.E.; Musetti, R. Sieve Element Biology Provides Leads for Research on Phytoplasma Lifestyle in Plant Hosts. J. Exp. Bot. 2019, 70, 3737–3755. [Google Scholar] [CrossRef]
- Gallinger, J.; Zikeli, K.; Zimmermann, M.R.; Görg, L.M.; Mithöfer, A.; Reichelt, M.; Seemüller, E.; Gross, J.; Furch, A.C.U. Specialized 16SrX Phytoplasmas Induce Diverse Morphological and Physiological Changes in Their Respective Fruit Crops. PLOS Pathog. 2021, 17, e1009459. [Google Scholar] [CrossRef]
- Caudwell, A. Identification d’une Nouvelle Maladie à Virus de La Vigne «la Flavescence Dorée». Étude Des Phénomènes de Localisation Des Symptômes et de Rétablissement; Annales des épiphyties, Institut national de la recherche agronomique: Paris, France, 1964. [Google Scholar]
- Dermastia, M.; Bertaccini, A.; Constable, F.; Mehle, N. Grapevine Yellows Diseases and Their Phytoplasma Agents—Biology and Detection; Springer B; Springer: Berlin/Heidelberg, Germany; Springer International Publishing AG: Cham, Switzerland, 2017. [Google Scholar]
- Oliveira, M.J.R.A.; Castro, S.; Paltrinieri, S.; Bertaccini, A.; Sottomayor, M.; Santos, C.S.; Vasconcelos, M.W.; Carvalho, S.M.P. “Flavescence Dorée” Impacts Growth, Productivity and Ultrastructure of Vitis vinifera Plants in Portuguese “Vinhos Verdes” Region. Sci. Hortic. 2020, 261, 108742. [Google Scholar] [CrossRef]
- Musetti, R.; Buxa, S.V.; de Marco, F.; Loschi, A.; Polizzotto, R.; Kogel, K.-H.; Van Bel, A.J.E. Phytoplasma-Triggered Ca2+ Influx Is Involved in Sieve-Tube Blockage. Mol. Plant-Microbe Interact. 2013, 26, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Mendgen, K. Untersuchungen Über Eine Vergilbungskrankheit Der Reben an Rhein, Mosel Und Saar (Research on a Yellows Disease of Grapevine in Rhine, Mosel and Saar Regions). Weinberg und Keller 1971, 18, 345–431. [Google Scholar]
- Jelmini, L.; Rizzoli, A.; Jermini, M.; Schumpp, O.; Conedera, M. Phloem and Xylem Modifications of Vitis vinifera Stems in Response to Flavescence Dorée Phytoplasma Infection. Plant Pathol. 2021, 70, 970–979. [Google Scholar] [CrossRef]
- Maggi, F.; Palmano, S.; Bosco, D.; Marzachì, C.; Galetto, L. Space-Time Point Pattern Analysis of Flavescence Dorée Epidemic in a Grapevine Field: Disease Progression and Recovery. Front. Plant Sci. 2017, 7, 1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, A.L.; Rodriguez-Zaccaro, F.D.; Lee, T.F.; Valdovinos, J.; Toschi, H.S.; Martinez, J.A.; Pratt, R.B. Grapevine Xylem Development, Architecture, and Function. In Functional and Ecological Xylem Anatomy; Hacke, U., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 133–162. ISBN 978-3-319-15783-2. [Google Scholar]
- Wimmer, R. Wood Anatomical Features in Tree-Rings as Indicators of Environmental Change. Dendrochronologia 2002, 20, 21–36. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree Growth as Indicator of Tree Vitality and of Tree Reaction to Environmental Stress: A Review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- MeteoSwiss Climate Normals (1981–2010). Available online: https://www.meteoswiss.admin.ch/home/climate/swiss-climate-in-detail/climate-normals/normal-values-per-measured-parameter.html (accessed on 13 March 2022).
- Eveillard, S.; Jollard, C.; Labroussaa, F.; Khalil, D.; Perrin, M.; Desqué, D.; Salar, P.; Razan, F.; Hévin, C.; Bordenave, L.; et al. Contrasting Susceptibilities to Flavescence Dorée in Vitis vinifera, Rootstocks and Wild Vitis Species. Front. Plant Sci. 2016, 7, 1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, C.; Salar, P.; Gillet, J.; Cloquemin, C.; Very, P.; Foissac, X.; Malembic-Maher, S. Triplex Real-Time PCR Assay for Sensitive and Simultaneous Detection of Grapevine Phytoplasmas of the 16SrV and 16SrXII-A Groups with an Endogenous Analytical Control. Vitis 2009, 48, 87–95. [Google Scholar] [CrossRef]
- Russo, M. Electron Microscopy of Grapevine Virus Infections. Phytopathol. Mediterr. 1985, 24, 144–147. [Google Scholar]
- Laboratoire de la santé des végétaux (ANSES). Détection Des Phytoplasmes de La Vigne, Groupe 16SrV (Flavescence Dorée) et Groupe 16SrXII (Bois Noir): MOA 006 Version 2a; Ministère de l’alimentation, de l’agriculture et de la pêche: Paris, France, 2015. [Google Scholar]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A New Tool for Sampling Microcores from Tree Stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Gärtner, H.; Schweingruber, F. Microscopic Preparation Techniques for Plant Stem Analysis; Verlag Dr. Kessel: Remagen-Oberwinter, Germany, 2013; ISBN 378-3-941300-76-7. [Google Scholar]
- Schweingruber, F.H. Dendroökologische Holzanatomie. Anatomische Grundlagen Der Dendrochronologie; WSL; Haupt Verlag: Bern, Switzerland, 2001; ISBN 978-3258063294. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Munitz, S.; Netzer, Y.; Shtein, I.; Schwartz, A. Water Availability Dynamics Have Long-Term Effects on Mature Stem Structure in Vitis vinifera. Am. J. Bot. 2018, 105, 1443–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shtein, I.; Hayat, Y.; Munitz, S.; Harcavi, E.; Akerman, M.; Drori, E.; Schwartz, A.; Netzer, Y. From Structural Constraints to Hydraulic Function in Three Vitis Rootstocks. Trees-Struct. Funct. 2017, 31, 851–861. [Google Scholar] [CrossRef]
- De Micco, V.; Zalloni, E.; Battipaglia, G.; Erbaggio, A.; Scognamiglio, P.; Caputo, R.; Cirillo, C. Rootstock Effect on Tree-Ring Traits in Grapevine under a Climate Change Scenario. IAWA J. 2017, 39, 145–155. [Google Scholar] [CrossRef]
- Quintana-Pulido, C.; Villalobos, L.; Muñoz-Araya, M.; Franck, N.; Pastenes, C. Xylem Structure and Function in Three Grapevine Varieties. Chil. J. Agric. Res. 2018, 78, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-Induced Changes in Development and Function of Grapevine (Vitis spp.) Organs and in Their Hydraulic and Non-Hydraulic Interactions at the Whole-Plant Level: A Physiological and Molecular Update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The Physiology of Drought Stress in Grapevine: Towards an Integrative Definition of Drought Tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Lessio, F.; Alma, A. Dispersal Patterns and Chromatic Response of Scaphoideus titanus Ball (Homoptera Cicadellidae), Vector of the Phytoplasma Agent of Grapevine Flavescence Dorée. Agric. For. Entomol. 2004, 6, 121–127. [Google Scholar] [CrossRef]
- Mazzoni, V.; Trona, F.; Ioriatti, C.; Andrea, L.; Eriksson, A.; Anfora, G. Attractiveness of Different Colours to Scaphoideus titanus Ball (Hemiptera: Cicadellidae) Adults. IOBC/WPRS Bull. 2011, 67, 281–284. [Google Scholar]
- Chuche, J.; Boudon-Padieu, E.; Thiéry, D. Host Preferences of the Leafhopper Scaphoideus titanus, Vector of “Flavescence Dorée” Phytoplasma. Phytopathogenic Mollicutes 2016, 6, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Intrigliolo, D.; Castel, J. Evaluation of Grapevine Water Status from Trunk Diameter Variations. Irrig. Sci. 2007, 26, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Landi, L.; Murolo, S.; Romanazzi, G. Detection of ‘Candidatus Phytoplasma solani’ in Roots from Bois Noir Symptomatic and Recovered Grapevines. Sci. Rep. 2019, 9, 2013. [Google Scholar] [CrossRef] [PubMed]
- Morone, C.; Rossi, V.; Giosuè, S.; Scapin, I.; Gotta, P.; Marzachì, C.; Boveri, M. Epidemiology of Flavescence Dorée in Vineyards in Northwestern Italy. Phytopathology 2007, 97, 1422–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margaria, P.; Ferrandino, A.; Caciagli, P.; Kedrina, O.; Schubert, A.; Palmano, S. Metabolic and Transcript Analysis of the Flavonoid Pathway in Diseased and Recovered Nebbiolo and Barbera Grapevines (Vitis vinifera L.) Following Infection by Flavescence Dorée Phytoplasma. Plant Cell Environ. 2014, 37, 2183–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifico, D.; Margaria, P.; Galetto, L.; Legovich, M.; Abbà, S.; Veratti, F.; Marzachì, C.; Palmano, S. Differential Gene Expression in Two Grapevine Cultivars Recovered from “Flavescence Dorée”. Microbiol. Res. 2019, 220, 72–82. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | June | July | August | September | ySPEI6 |
|---|---|---|---|---|---|
| 2010 | −0.450 | −0.407 | 0.272 | 0.059 | −0.131 |
| 2011 | −2.181 | −1.494 | −1.543 | −1.545 | −1.691 |
| 2012 | −0.688 | −1.137 | −0.684 | −0.254 | −0.692 |
| 2013 | 0.378 | 0.336 | 0.083 | 0.227 | 0.256 |
| 2014 | −0.969 | −0.757 | −1.163 | −1.523 | −1.103 |
| 2015 | −1.134 | −1.849 | −2.048 | −1.881 | −1.728 |
| 2016 | 0.092 | −0.315 | −1.215 | −1.494 | −0.733 |
| 2017 | −1.477 | −1.700 | −1.824 | −1.990 | −1.748 |
| 2018 | −1.297 | −2.309 | −2.580 | −2.836 | −2.255 |
| 2019 | −1.638 | −1.957 | −1.937 | −2.047 | −1.895 |
| Category | Number of Specimens | Xylem Anatomical Features 2019 | Xylem Annual Widths | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No Vessels | Incomplete Early Wood | Early Wood Only | Early and Late Wood | Year Type | Annual Rings | ||||
| N. Obs | Average Width (mm) | SD (mm) | |||||||
| A_dt | 22 | 0 | 3 | 0 | 19 | normal * | 79 | 1.202 | 0.24 |
| drought † | 77 | 1.123 | 0.38 | ||||||
| 2019 | 22 | 1.020 | 0.45 | ||||||
| A_ds | 4 | 0 | 1 | 0 | 3 | normal * | 21 | 1.305 | 0.23 |
| drought † | 14 | 0.776 | 0.07 | ||||||
| 2019 | 4 | 0.751 | 0.52 | ||||||
| S_dt | 16 | 3 | 1 | 12 | 0 | normal * | 77 | 1.443 | 0.27 |
| drought † | 62 | 1.277 | 0.23 | ||||||
| 2019 | 16 | 0.349 | 0.18 | ||||||
| S_ds | 7 | 4 | 0 | 3 | 0 | normal * | 45 | 1.261 | 0.30 |
| drought † | 28 | 0.783 | 0.28 | ||||||
| 2019 | 7 | 0.164 | 0.09 | ||||||
| Category | Growing Sector | N | Samples with Phloem Anomalies in 2019 | ||
|---|---|---|---|---|---|
| 0% | 1–50% | 51–100% | |||
| A_dt | top | 8 | 7 | 1 | 0 |
| slope | 5 | 5 | 0 | 0 | |
| toeslope | 9 | 9 | 0 | 0 | |
| total | 22 | 21 | 1 | 0 | |
| A_ds | top | 2 | 2 | 0 | 0 |
| slope | 1 | 1 | 0 | 0 | |
| toeslope | 1 | 0 | 1 | 0 | |
| total | 4 | 3 | 1 | 0 | |
| S_dt | top | 3 | 0 | 1 | 2 |
| slope | 4 | 0 | 0 | 4 | |
| toeslope | 9 | 0 | 3 | 6 | |
| total | 16 | 0 | 4 | 12 | |
| S_ds | top | 5 | 0 | 0 | 5 |
| slope | 1 | 0 | 1 | 0 | |
| toeslope | 1 | 0 | 0 | 1 | |
| total | 7 | 0 | 1 | 6 | |
| GLM Statistics | Predictor | df | F | p | |||
|---|---|---|---|---|---|---|---|
| Response | R2 | F | p | ||||
| Ring width | 0.22 | 20.72 | < 0.001 | symptom | 1, 382 | 3.98 | <0.05 |
| sector | 2, 382 | 9.88 | <0.001 | ||||
| ySPEI_6 | 1, 382 | 70.76 | <0.001 | ||||
| drought_s * ySPEI_6 | 1, 382 | 7.29 | <0.01 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzoli, A.; Jelmini, L.; Pezzatti, G.B.; Jermini, M.; Schumpp, O.; Debonneville, C.; Marcolin, E.; Krebs, P.; Conedera, M. Impact of the “Flavescence Dorée” Phytoplasma on Xylem Growth and Anatomical Characteristics in Trunks of ‘Chardonnay’ Grapevines (Vitis vinifera). Biology 2022, 11, 978. https://doi.org/10.3390/biology11070978
Rizzoli A, Jelmini L, Pezzatti GB, Jermini M, Schumpp O, Debonneville C, Marcolin E, Krebs P, Conedera M. Impact of the “Flavescence Dorée” Phytoplasma on Xylem Growth and Anatomical Characteristics in Trunks of ‘Chardonnay’ Grapevines (Vitis vinifera). Biology. 2022; 11(7):978. https://doi.org/10.3390/biology11070978
Chicago/Turabian StyleRizzoli, Attilio, Luca Jelmini, Gianni Boris Pezzatti, Mauro Jermini, Olivier Schumpp, Christophe Debonneville, Enrico Marcolin, Patrik Krebs, and Marco Conedera. 2022. "Impact of the “Flavescence Dorée” Phytoplasma on Xylem Growth and Anatomical Characteristics in Trunks of ‘Chardonnay’ Grapevines (Vitis vinifera)" Biology 11, no. 7: 978. https://doi.org/10.3390/biology11070978