
Analysis of Physiological and Transcriptomic Differences between a Premature Senescence Mutant (GSm) and Its Wild-Type in Common Wheat (Triticum aestivum L.)


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Material Preparation and Agronomic Analysis
2.2. Measurement of Photosynthetic Performance
2.3. Analysis of Senescence-Related Physiological Parameters
2.4. RNA Extraction, Library Construction, and Sequencing
2.5. Bioinformatics Analysis of RNA-Seq Data
2.6. Quantitative Reverse-Transcription PCR
3. Results
3.1. Manifestations of the Premature Senescence Mutant (GSm)
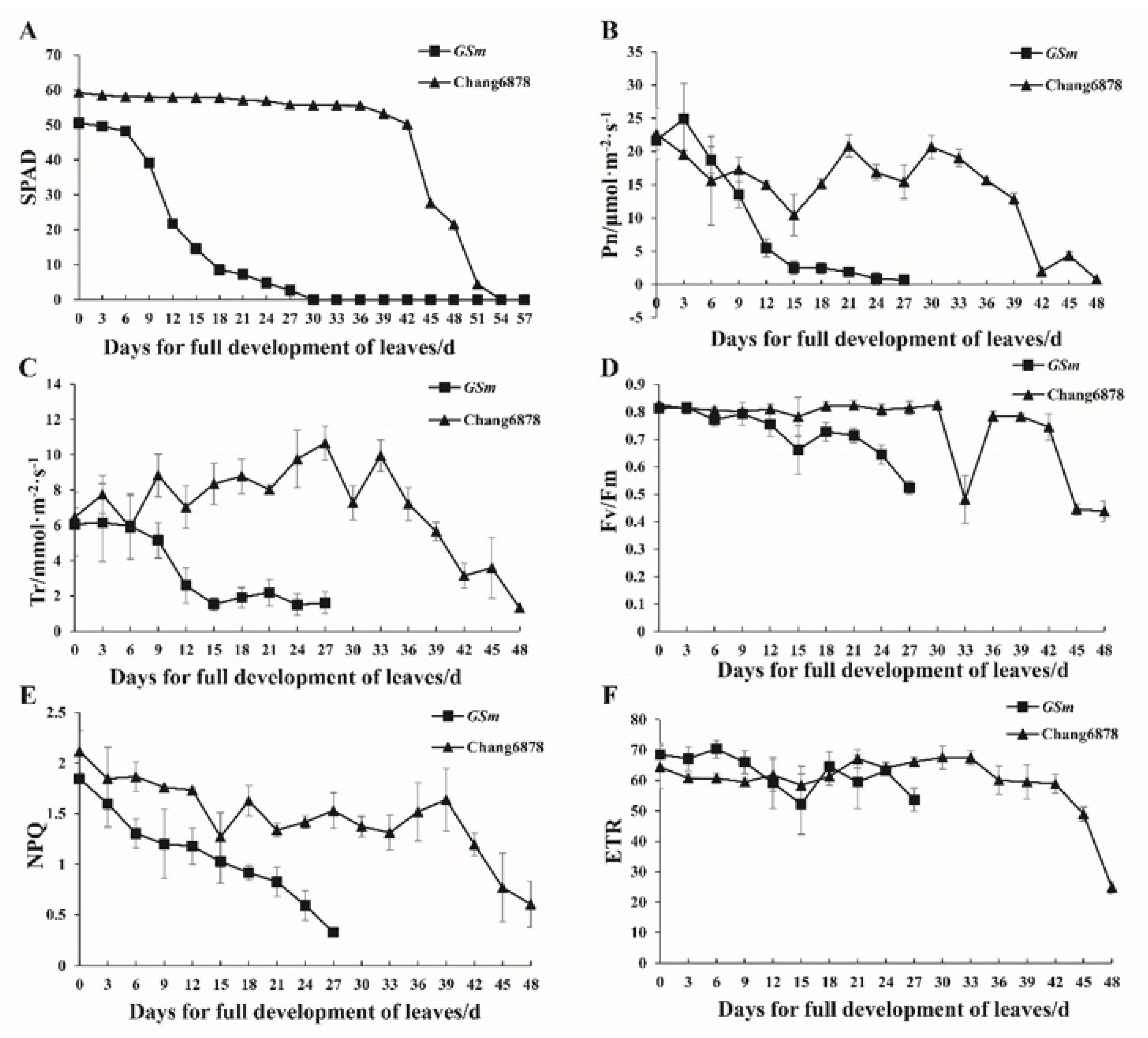
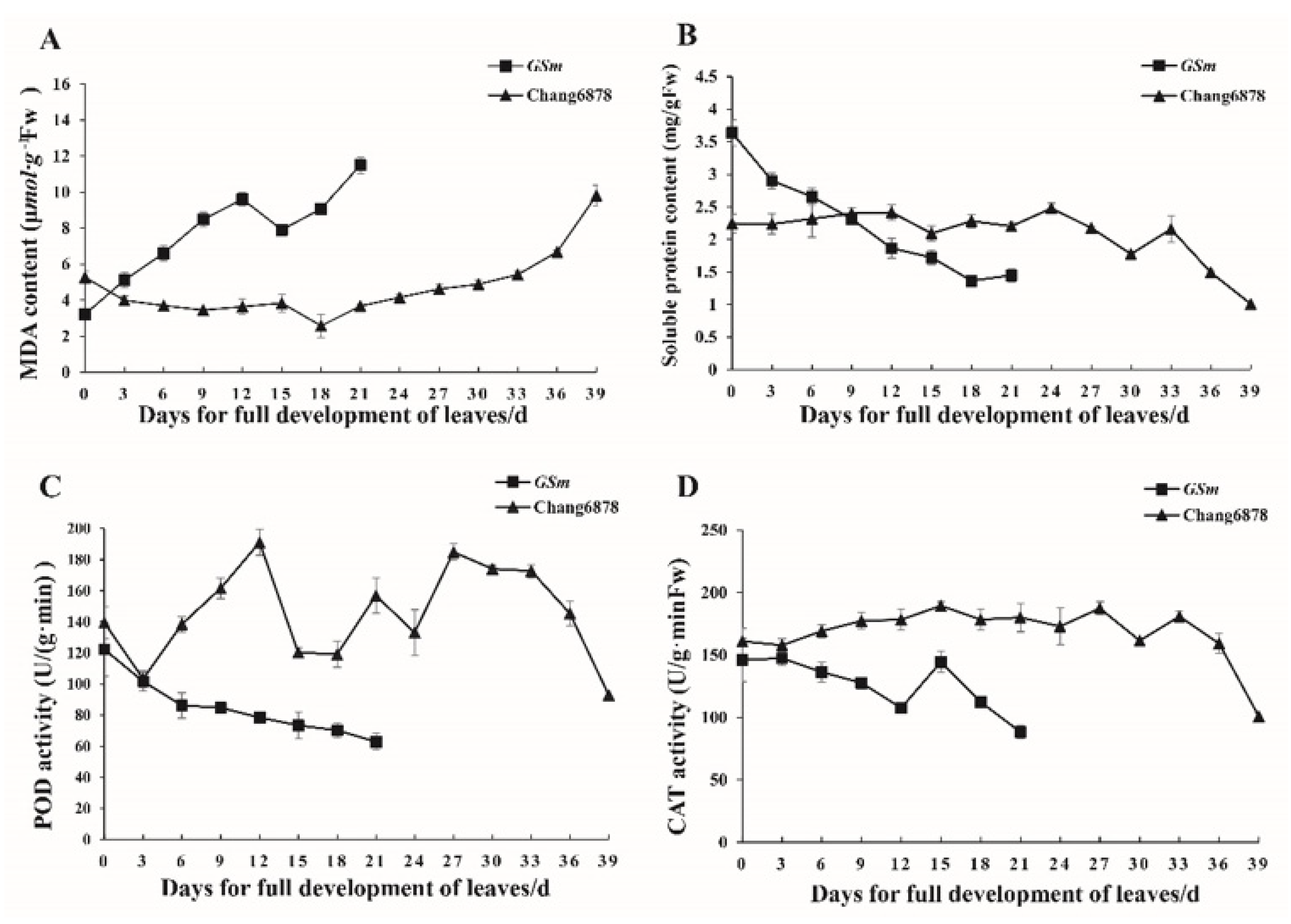
3.2. Differences in Physiological Traits between the Premature Senescence Mutant (GSm) and Its Wild-Type
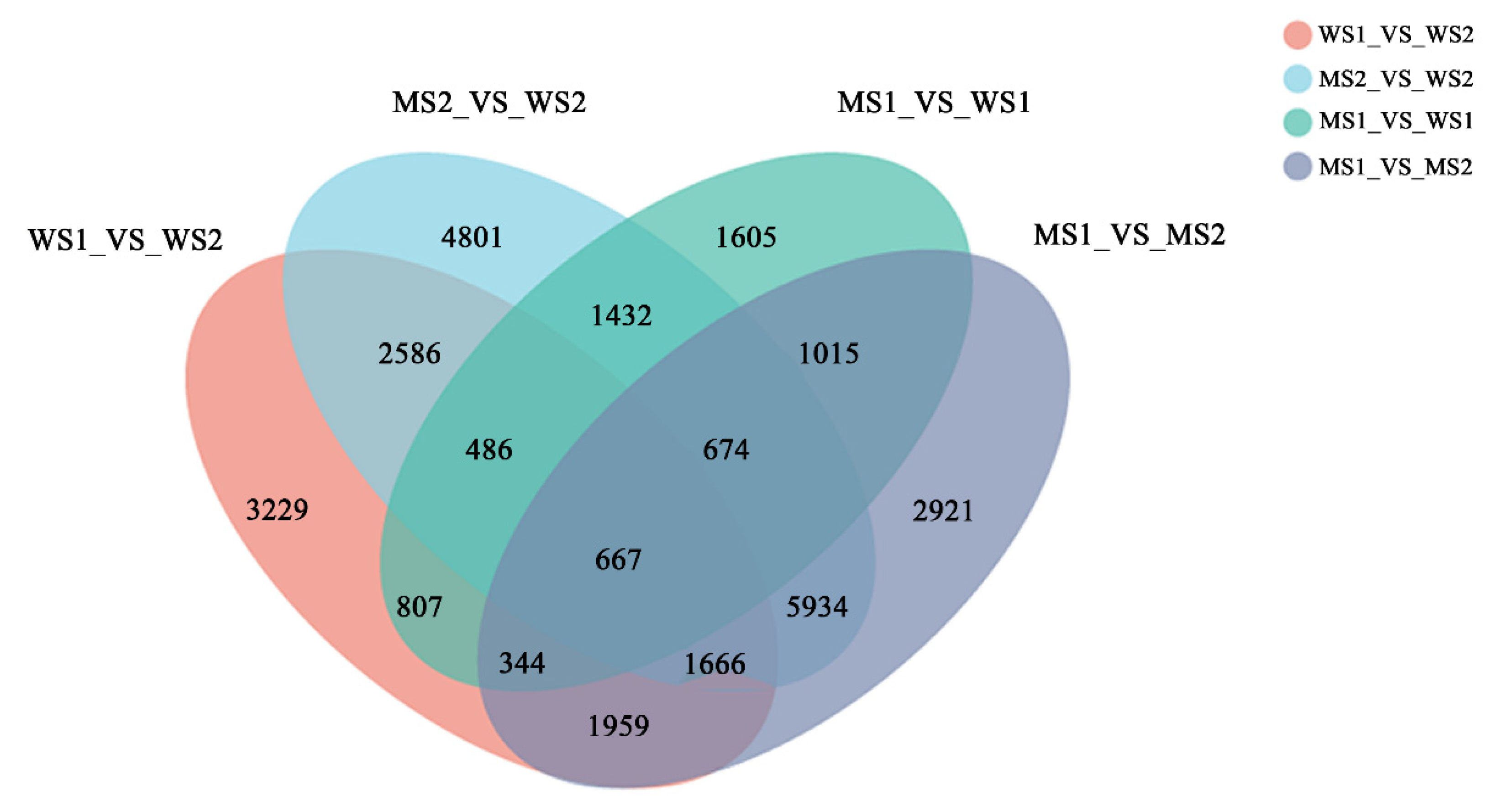
3.3. RNA Sequencing Analysis and Identificaiton of Differentially Expressed Genes
3.4. GO Enrichment and KEGG Pathway Analysis of Differentially Expressed Genes
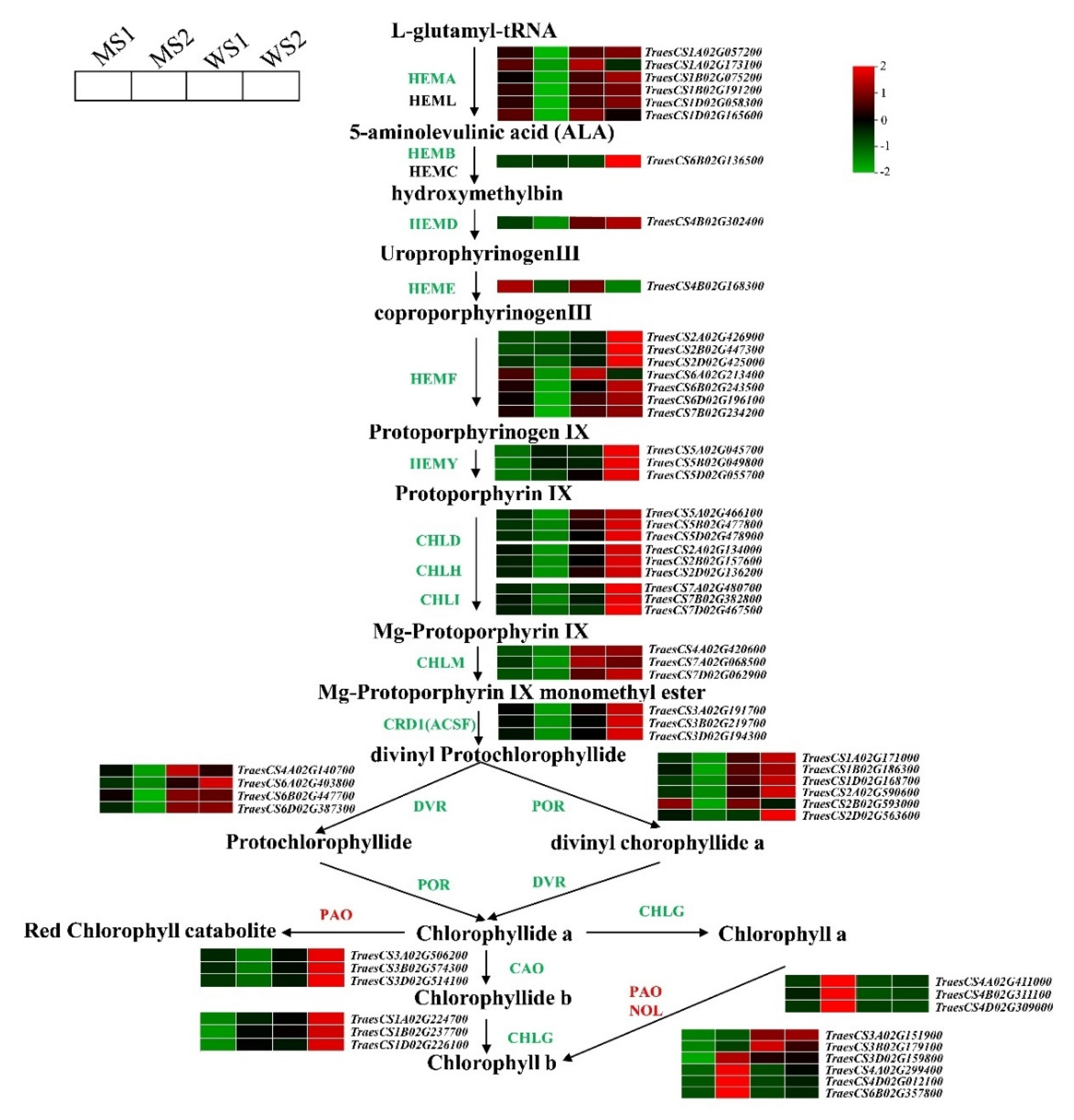
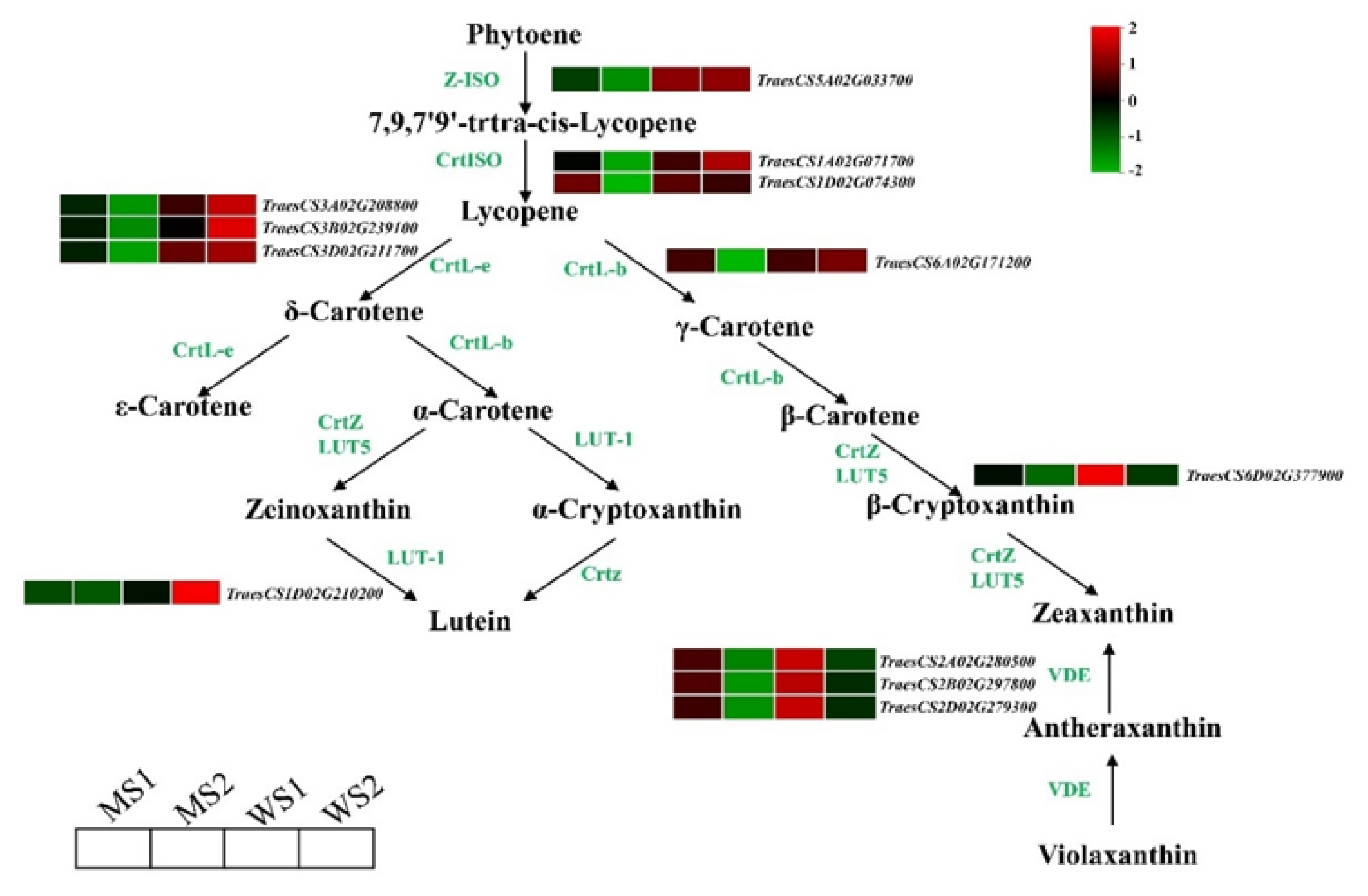
3.5. Differentially Expressed Genes Involved in Chlorophyll and Carotenoid Biosynthesis and Photosynthesis
3.6. Differentially Expressed Genes Related to Antioxidative Metabolism and Protein Processing, Transportation
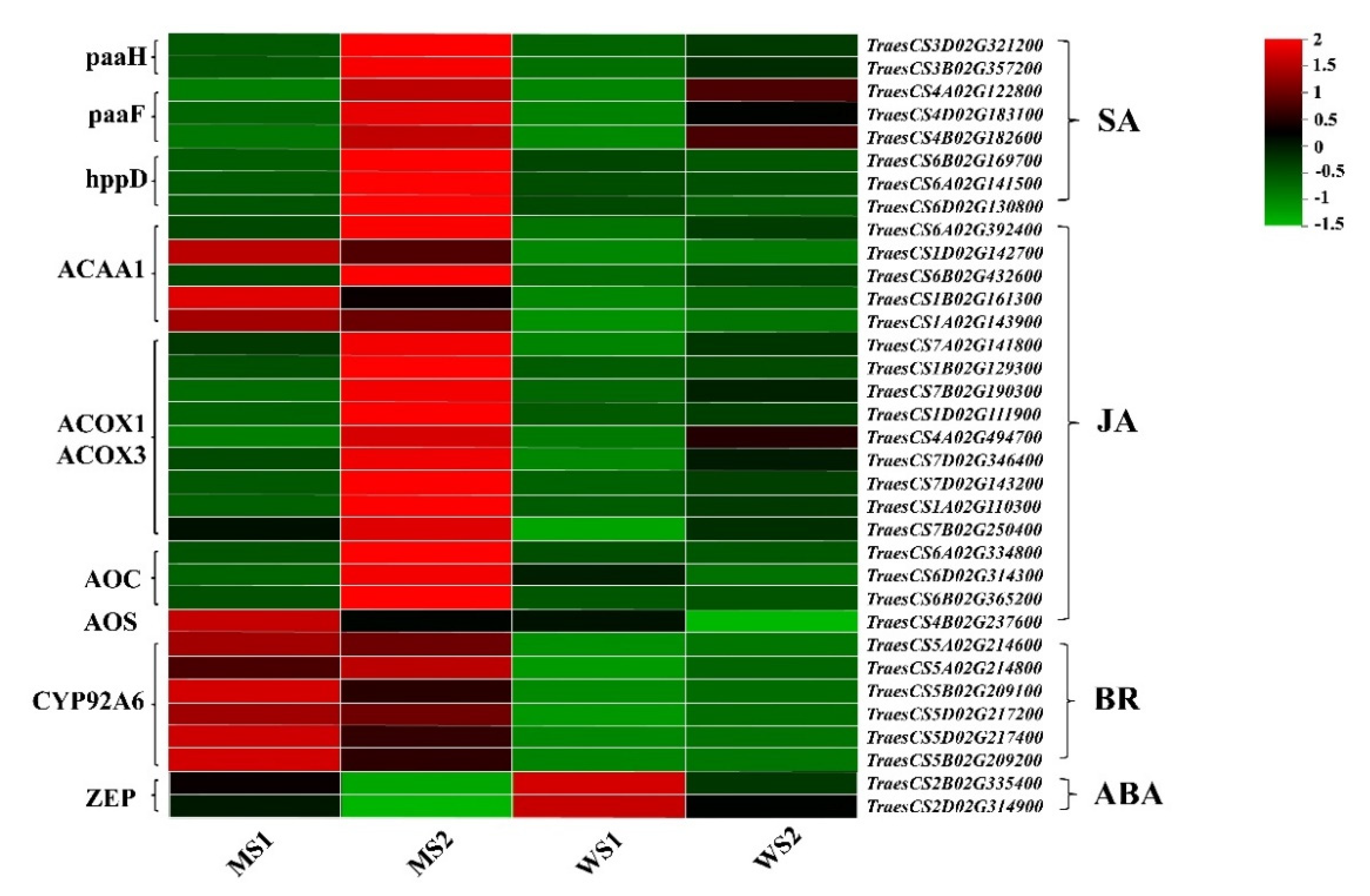
3.7. Differentially Expressed Genes Associated with Hormone Signaling
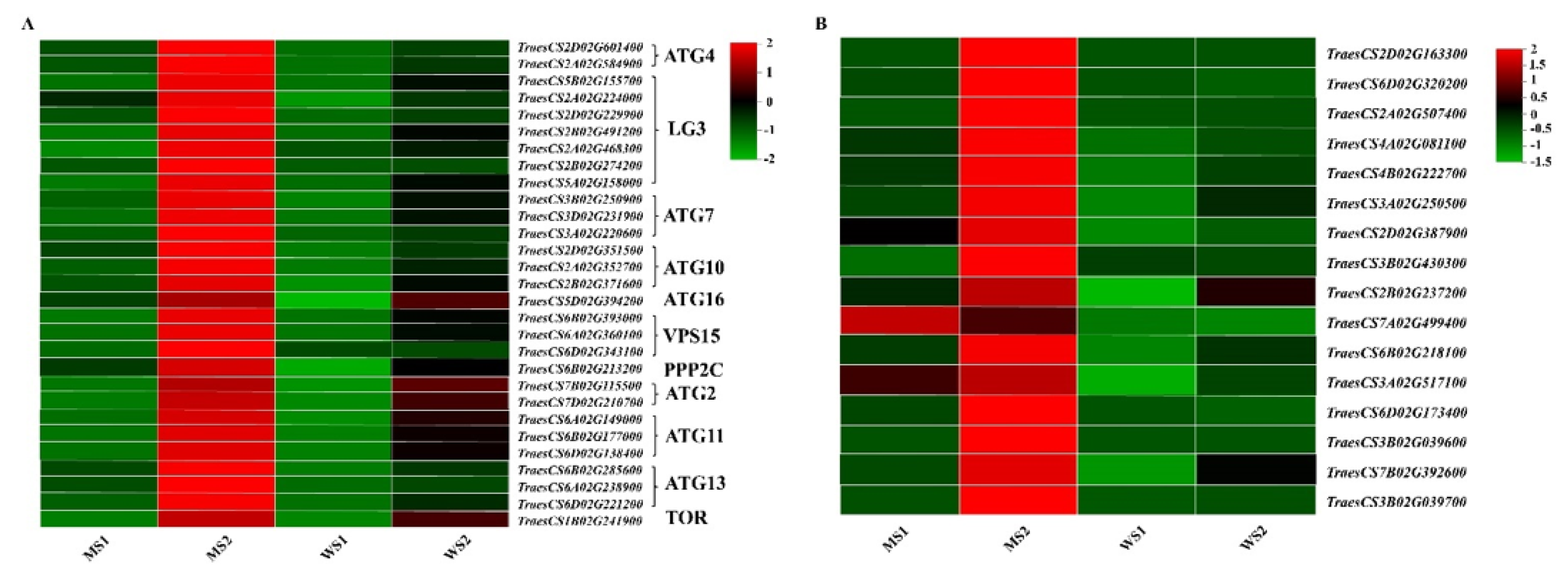
3.8. Differentially Expressed Genes Participating in Autophagy and Hydrolysis of Senescent Leaves
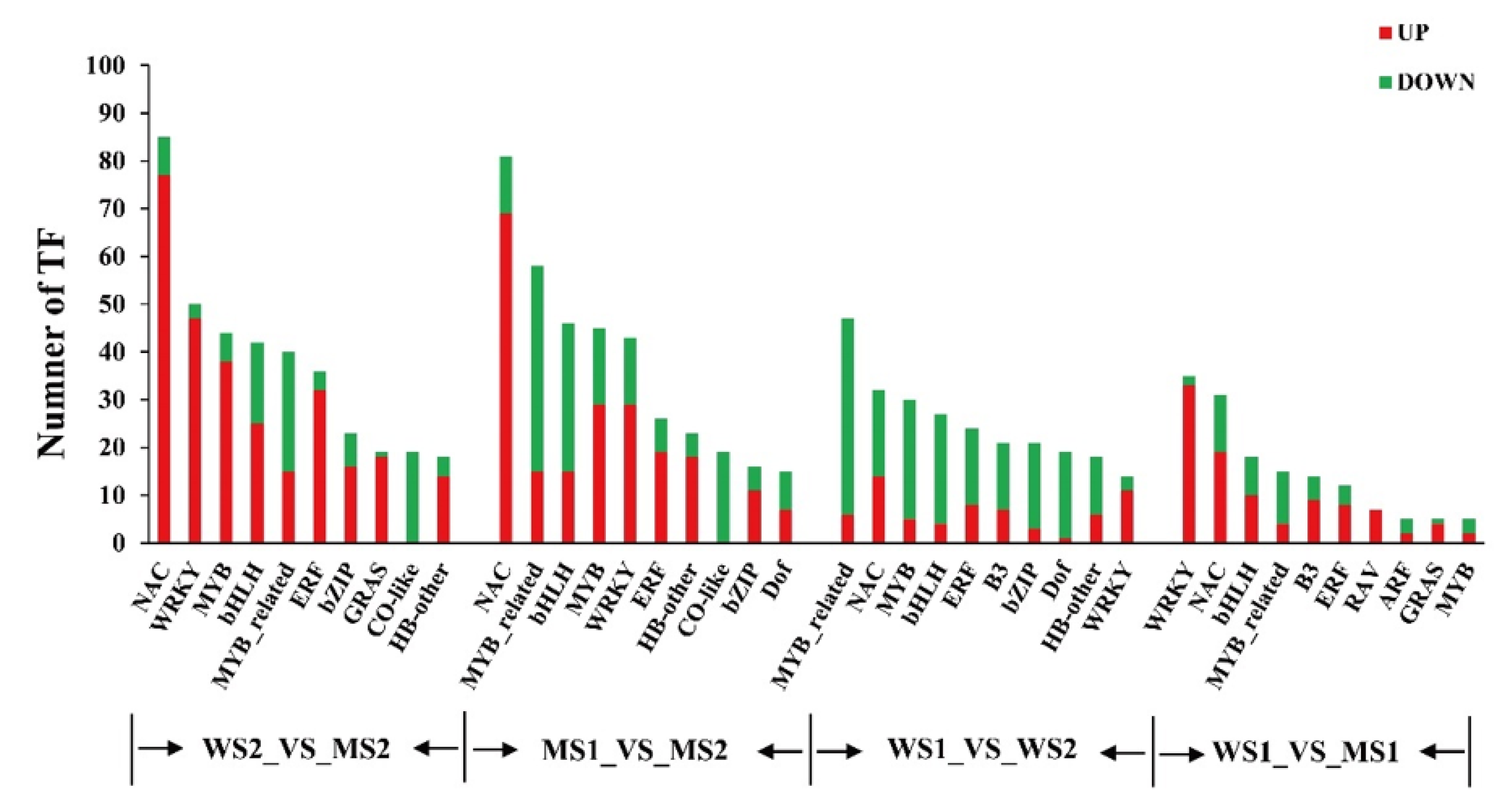
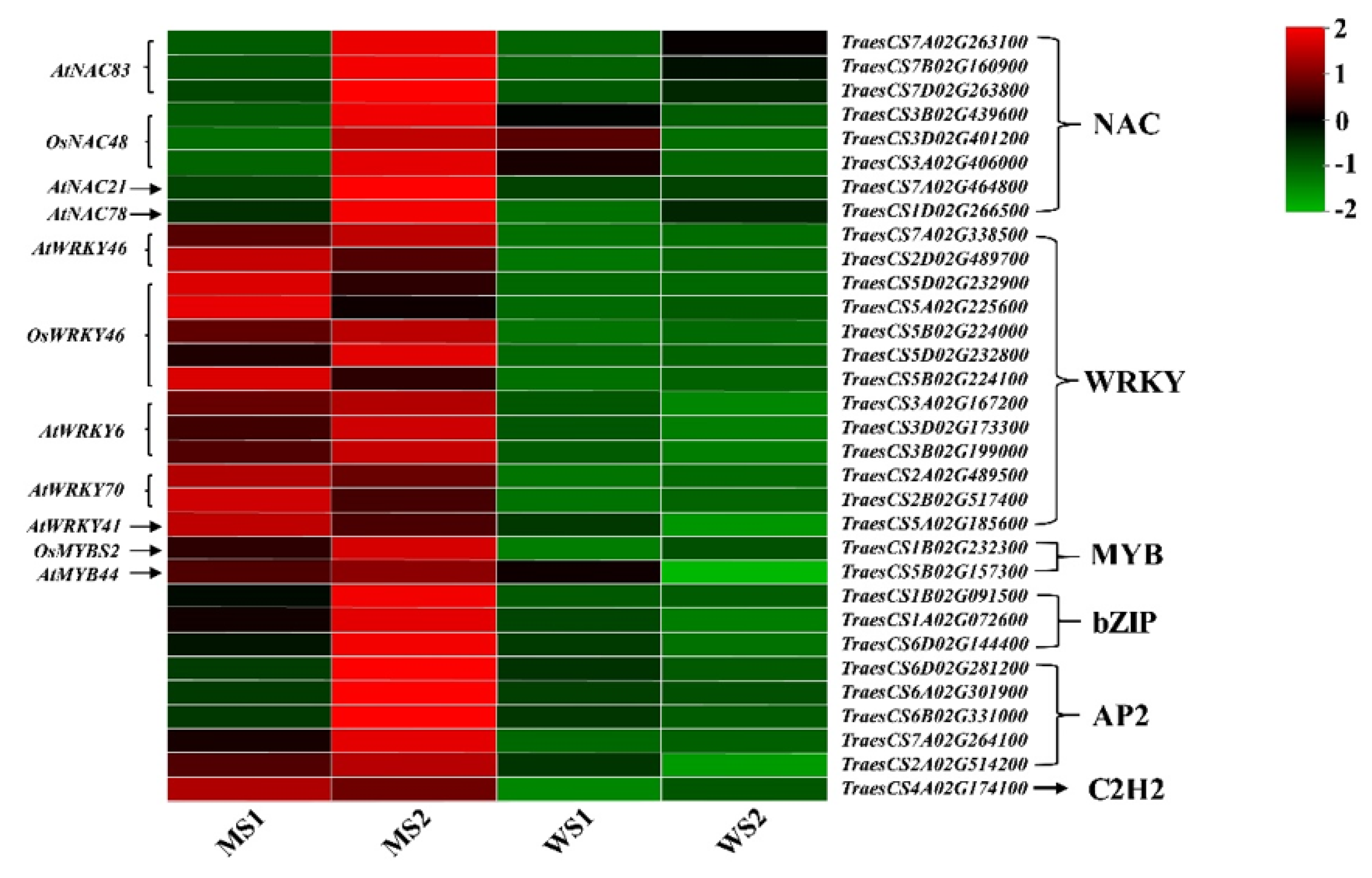
3.9. Transcription Factor Family Members in Differentially Expressed Genes
4. Discussion
4.1. Agronomic, Yield, and Physiological Traits Are Distinctively Different between the Common Wheat Premature Senescence Mutant and Its Wild-Type
4.2. Photosynthesis-Related Genes Are Differentially Expressed in the Leaves of Common Wheat Premature Senescence Mutant and Its Wild-Type
4.3. Antioxidant System-Related Genes Are Differentially Expressed between the Common Wheat Premature Senescence Mutant and Its Wild-Type
4.4. Plant Hormones and Autophagy Play an Essential Regulatory Role in Leaf Senescence
4.5. Transcription Factors Play a Vital Role in Senescence Regulation in Common Wheat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sultana, N.; Islam, S.; Juhasz, A.; Ma, W. Wheat leaf senescence and its regulatory gene network. Crop J. 2021, 9, 703–717. [Google Scholar] [CrossRef]
- Wu, X.-Y.; Kuai, B.-K.; Jia, J.-Z.; Jing, H.-C. Regulation of Leaf Senescence and Crop Genetic ImprovementF. J. Integr. Plant Biol. 2012, 54, 936–952. [Google Scholar] [CrossRef] [PubMed]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef]
- Sang, X.-C.; Fang, L.-K.; Vanichpakorn, Y.; Ling, Y.-H.; Du, P.; Zhao, F.-M.; Yang, Z.-L.; He, G.-H. Physiological character and molecular mapping of leaf-color mutant wyv1 in rice (Oryza sativa L.). Genes Genomics. 2010, 32, 123–128. [Google Scholar] [CrossRef]
- Distelfeld, A.; Avni, R.; Fischer, A. Senescence, nutrient remobilization, and yield in wheat and barley. J. Exp. Bot. 2014, 65, 3783–3798. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant Mol. Biol. 2013, 82, 603–622. [Google Scholar] [CrossRef]
- Ischebeck, T.; Zbierzak, A.M.; Kanwischer, M.; Dörmann, P. A Salvage Pathway for Phytol Metabolism in Arabidopsis. J. Biol. Chem. 2006, 281, 2470–2477. [Google Scholar] [CrossRef]
- Girondé, A.; Etienne, P.; Trouverie, J.; Bouchereau, A.; Le Cahérec, F.; Leport, L.; Orsel, M.; Niogret, M.-F.; Nesi, N.; Carole, D.; et al. The contrasting N management of two oilseed rape genotypes reveals the mechanisms of proteolysis associated with leaf N remobilization and the respective contributions of leaves and stems to N storage and remobilization during seed filling. BMC Plant Biol. 2015, 15, 59. [Google Scholar] [CrossRef]
- Uauy, C.; Brevis, J.C.; Dubcovsky, J. The high grain protein content gene Gpc-B1 accelerates senescence and has pleiotropic effects on protein content in wheat. J. Exp. Bot. 2006, 57, 2785–2794. [Google Scholar] [CrossRef]
- Yang, J.; Worley, E.; Udvardi, M. A NAP-AAO3 regulatory module promotes chlorophyll degradation via ABA biosynthesis in Arabidopsis leaves. Plant Cell 2014, 26, 4862–4874. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Luo, S.; Araújo, W.L.; Schlicke, H.; Rothbart, M.; Yu, J.; Fan, T.; Fernie, A.R.; Grimm, B.; Luo, M. Virus-induced gene silencing of pea CHLI and CHLD affects tetrapyrrole biosynthesis, chloroplast development and the primary metabolic network. Plant Physiol. Biochem. 2013, 65, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.-I.; Sueda, K.; Burgyan, J.; Masuta, C. A Viral Satellite RNA Induces Yellow Symptoms on Tobacco By Targeting a Gene Involved in Chlorophyll Biosynthesis Using the RNA Silencing Machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Xu, X.; He, C.; Liu, C.; Liu, Q.; Dong, R.; Liu, T.; Wang, L. Fine Mapping and Candidate Gene Analysis of the Leaf-Color Gene ygl-1 in Maize. PLoS ONE. 2016, 11, e0153962. [Google Scholar] [CrossRef] [PubMed]
- Prina, A.R.; Arias, M.C.; Lainez, V.; Landau, A.; Maldonado, S. A cytoplasmically inherited mutant controlling early chloroplast development in barley seedlings. Theor. Appl. Genet. 2003, 107, 1410–1418. [Google Scholar] [CrossRef]
- Jarvis, P.; Dörmann, P.; Peto, C.A.; Lutes, J.; Benning, C.; Chory, J. Galactolipid deficiency and abnormal chloroplast development in the Arabidopsis MGD synthase 1 mutant. Proc. Natl. Acad. Sci. USA 2000, 97, 8175–8179. [Google Scholar] [CrossRef]
- Holding, D.R.; Springer, P.S.; Coomber, S.A. The Chloroplast and Leaf Developmental Mutant, pale cress, Exhibits Light-conditional Severity and Symptoms Characteristic of its ABA Deficiency. Ann. Bot. 2000, 86, 953–962. [Google Scholar] [CrossRef]
- Gupta, R.; Jiao, S.; Zhao, S.; Meeley, R.B.; Williams, R.W.; Taramino, G.; Feng, D.; Li, G.; Liu, J.; Allen, S.M.; et al. The maize premature senesence2 encodes for PHYTOCHROME-DEPENDENT LATE-FLOWERING and its expression modulation improves agronomic traits under abiotic stresses. Plant Direct. 2020, 4, e00295. [Google Scholar] [CrossRef]
- Wu, H.-B.; Wang, B.; Chen, Y.; Liu, Y.-G.; Chen, L. Characterization and fine mapping of the rice premature senescence mutant ospse1. Theor. Appl. Genet. 2013, 126, 1897–1907. [Google Scholar] [CrossRef]
- Aung, K.; Lin, S.-I.; Wu, C.-C.; Huang, Y.-T.; Su, C.-L.; Chiou, T.-J. pho2, a Phosphate Overaccumulator, Is Caused by a Nonsense Mutation in a MicroRNA399 Target Gene. Plant Physiol. 2006, 141, 1000–1011. [Google Scholar] [CrossRef]
- Kong, Z.; Li, M.; Yang, W.; Xu, W.; Xue, Y. A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice. Plant Physiol. 2006, 141, 1376–1388. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, Z.; Li, L.; Tang, S.; Wu, J.-L. Genetic and Physio-Biochemical Characterization of a Novel Premature Senescence Leaf Mutant in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2018, 19, 2339. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wu, X.; Liu, G.; Zhang, Z.; Chao, J.; Li, Z.; Guo, Y.; Sun, Y. Characterization and Mapping of a Novel Premature Leaf Senescence Mutant in Common Tobacco (Nicotiana tabacum L.). Plants 2019, 8, 415. [Google Scholar] [CrossRef]
- Fracheboud, Y.; King, P.J. An auxin-auxotrophic mutant of Nicotiana plumbaginifolia. Mol. Gen. Genet. MGG 1991, 227, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-M.; Zhang, X.-W.; Liu, Y.-W.; Li, K.; Tan, Q.; Zhou, S.; Wang, G.; Zhou, C.-J. A WRKY transcription factor, TaWRKY42-B, facilitates initiation of leaf senescence by promoting jasmonic acid biosynthesis. BMC Plant Biol. 2020, 20, 444. [Google Scholar] [CrossRef]
- Jiang, Y.; Liang, G.; Yang, S.; Yu, D. Arabidopsis WRKY57 Functions as a Node of Convergence for Jasmonic Acid- and Auxin-Mediated Signaling in Jasmonic Acid-Induced Leaf Senescence. Plant Cell 2014, 26, 230–245. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, S.; Chen, Y.; Li, D.; Yu, D. Arabidopsis WRKY45 Interacts with the DELLA Protein RGL1 to Positively Regulate Age-Triggered Leaf Senescence. Mol. Plant 2017, 10, 1174–1189. [Google Scholar] [CrossRef]
- Gao, S.; Gao, J.; Zhu, X.; Song, Y.; Li, Z.; Ren, G.; Zhou, X.; Kuai, B. ABF2, ABF3, and ABF4 Promote ABA-Mediated Chlorophyll Degradation and Leaf Senescence by Transcriptional Activation of Chlorophyll Catabolic Genes and Senescence-Associated Genes in Arabidopsis. Mol. Plant 2016, 9, 1272–1285. [Google Scholar] [CrossRef]
- Markwell, J.; Osterman, J. Occurrence of Temperature-Sensitive Phenotypic Plasticity in Chlorophyll-Deficient Mutants of Arabidopsis thaliana. Plant Physiol. 1992, 98, 392–394. [Google Scholar] [CrossRef]
- Pasini, L.; Bruschini, S.; Bertoli, A.; Mazza, R.; Fracheboud, Y.; Marocco, A. Photosynthetic performance of cold-sensitive mutants of maize at low temperature. Physiol. Plant. 2005, 124, 362–370. [Google Scholar] [CrossRef]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.-I.; Asensi-Fabado, M.A.; Munné-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yan, M.; Zhu, X.; Zhang, T.; Shen, W.; Yu, P.; Wang, Y.; Sang, X.; Yu, G.; Zhao, B.; et al. Gene mapping of starch accumulation and premature leaf senescence in the ossac3 mutant of rice. Euphytica 2018, 214, 177. [Google Scholar] [CrossRef]
- Huang, Q.N.; Shi, Y.F.; Zhang, X.B.; Song, L.X.; Feng, B.H.; Wang, H.M.; Xu, X.; Li, X.H.; Guo, D.; Wu, J.L. Single base substitution in OsCDC48 is responsible for premature senescence and death phenotype in rice. J. Integr. Plant Biol. 2016, 58, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gong, P.; Li, K.; Huang, F.; Cheng, F.; Pan, G. A single cytosine deletion in the OsPLS1 gene encoding vacuolar-type H+-ATPase subunit A1 leads to premature leaf senescence and seed dormancy in rice. J. Exp. Bot. 2016, 67, 2761–2776. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.; Xu, B.; Li, Y.; Ma, Y.; Wang, G. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of Anthurium andraeanum ‘Sonate’. Front. Plant Sci. 2015, 6, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Wang, P.; Wang, S.A.; Ma, L.; Li, L.; Yang, R.; Ma, Y.; Wang, Q. Comprehensive transcriptome analysis discovers novel candidate genes related to leaf color in a Lagerstroemia indica yellow leaf mutant. Genes Genom. 2015, 37, 851–863. [Google Scholar] [CrossRef]
- Chai, M.; Guo, Z.; Shi, X.; Li, Y.; Tang, J.; Zhang, Z. Dissecting the Regulatory Network of Leaf Premature Senescence in Maize (Zea mays L.) Using Transcriptome Analysis of ZmELS5 Mutant. Genes 2019, 10, 944. [Google Scholar] [CrossRef]
- Li, Z.; Pan, X.; Guo, X.; Fan, K.; Lin, W. Physiological and Transcriptome Analyses of Early Leaf Senescence for ospls1 Mutant Rice (Oryza sativa L.) during the Grain-Filling Stage. Int. J. Mol. Sci. 2019, 20, 1098. [Google Scholar] [CrossRef]
- Wu, H.; Shi, N.; An, X.; Liu, C.; Fu, H.; Cao, L.; Feng, Y.; Sun, D.; Zhang, L. Candidate Genes for Yellow Leaf Color in Common Wheat (Triticum aestivum L.) and Major Related Metabolic Pathways according to Transcriptome Profiling. Int. J. Mol. Sci. 2018, 19, 1594. [Google Scholar] [CrossRef]
- Zhang, Q.; Xia, C.; Zhang, L.; Dong, C.; Liu, X.; Kong, X. Transcriptome Analysis of a Premature Leaf Senescence Mutant of Common Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2018, 19, 782. [Google Scholar] [CrossRef]
- Li, F.; Zhang, X.; Song, B.; Li, J.; Shang, Z.; Guan, J. Combined effects of 1-MCP and MAP on the fruit quality of pear (Pyrus bretschneideri Reld cv. Laiyang) during cold storage. Sci. Hortic. 2013, 164, 544–551. [Google Scholar] [CrossRef]
- Saher, S.; Piqueras, A.; Hellin, E.; Olmos, E. Hyperhydricity in micropropagated carnation shoots: The role of oxidative stress. Physiol. Plant. 2004, 120, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, R.; Zong, W. Bisphenol S Interacts with Catalase and Induces Oxidative Stress in Mouse Liver and Renal Cells. J. Agric. Food Chem. 2016, 64, 6630–6640. [Google Scholar] [CrossRef] [PubMed]
- Kochba, J.; Lavee, S.; Spiegel-Roy, P. Differences in peroxidase activity and isoenzymes in embryogenic ane non-embryogenic ‘Shamouti’ orange ovular callus lines1. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Xu, J.; Rao, Y.-C.; Zeng, Y.-J.; Liu, H.-J.; Zheng, T.-T.; Zhang, G.-H.; Hu, J.; Guo, L.-B.; Qian, Q.; et al. Cloning and functional analysis of pale-green leaf (PGL10) in rice (Oryza sativa L.). Plant Growth Regul. 2016, 78, 69–77. [Google Scholar] [CrossRef]
- Jung, K.-H.; Hur, J.; Ryu, C.-H.; Choi, Y.; Chung, Y.-Y.; Miyao, A.; Hirochika, H.; An, G. Characterization of a Rice Chlorophyll-Deficient Mutant Using the T-DNA Gene-Trap System. Plant Cell Physiol. 2003, 44, 463–472. [Google Scholar] [CrossRef]
- Pattanayak, G.K.; Tripathy, B.C. Modulation of biosynthesis of photosynthetic pigments and light-harvesting complex in wild-type and gun5 mutant of Arabidopsis thaliana during impaired chloroplast development. Protoplasma 2016, 253, 747–752. [Google Scholar] [CrossRef]
- Xie, Q.; Liang, Y.; Zhang, J.; Zheng, H.; Dong, G.; Qian, Q.; Zuo, J. Involvement of a Putative Bipartite Transit Peptide in Targeting Rice Pheophorbide a Oxygenase into Chloroplasts for Chlorophyll Degradation during Leaf Senescence. J. Genet. Genom. 2016, 43, 145–154. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Chyzhykova, O.A.; Palladina, T.O. The role of amino acids and sugars in supporting of osmotic homeostasis in maize seedlings under salinization conditions and treatment with synthetic growth regulators. Ukr. Kyi Biokhimichnyi Zhurnal 2006, 78, 124–129. [Google Scholar]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Zhu, S.-S.; Zhao, Z.-C.; Pu, Z.-G.; Liu, T.-Z.; Luo, S.; Zhang, X. Gene Mapping and Cloning of a Premature Leaf Senescence Mutant psls1 in Rice. Acta Agron. Sin. 2017, 43, 51–62. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, Y.-Y.; Zheng, J.-M.; Cheng, Z.-J.; Jiang, K.-F.; Yang, L.; Cao, Y.-J.; You, S.-M.; Wan, J.-M.; Zheng, J.-K. Genetic Analysis and Fine Mapping of a Premature Leaf Senescence Mutant in Rice (Orzya sativa L.). Acta Agron. Sin. 2014, 40, 2070. [Google Scholar] [CrossRef]
- Li, H.; Wang, G.; Liu, S.; An, Q.; Zheng, Q.; Li, B.; Li, Z. Comparative changes in the antioxidant system in the flag leaf of early and normally senescing near-isogenic lines of wheat (Triticum aestivum L.). Plant Cell Rep. 2014, 33, 1109–1120. [Google Scholar] [CrossRef]
- Qiu, K.; Li, Z.; Yang, Z.; Chen, J.; Wu, S.; Zhu, X.; Gao, S.; Gao, J.; Ren, G.; Kuai, B.; et al. EIN3 and ORE1 Accelerate Degreening during Ethylene-Mediated Leaf Senescence by Directly Activating Chlorophyll Catabolic Genes in Arabidopsis. PLoS Genet. 2015, 11, e1005399. [Google Scholar] [CrossRef]
- Pauwels, L.; Morreel, K.; De Witte, E.; Lammertyn, F.; Van Montagu, M.; Boerjan, W.; Inzé, D.; Goossens, A. Mapping methyl jasmonate-mediated transcriptional reprogramming of metabolism and cell cycle progression in cultured Arabidopsis cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385. [Google Scholar] [CrossRef]
- Schommer, C.; Palatnik, J.F.; Aggarwal, P.; Chételat, A.; Cubas, P.; Farmer, E.E.; Nath, U.; Weigel, D. Control of jasmonate biosynthesis and senescence by miR319 targets. PLoS Biol. 2008, 6, e230. [Google Scholar] [CrossRef]
- He, K.; Gou, X.; Yuan, T.; Lin, H.; Asami, T.; Yoshida, S.; Russell, S.D.; Li, J. BAK1 and BKK1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr. Biol. 2007, 17, 1109–1115. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Z.-Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 Accumulates in the Nucleus in Response to Brassinosteroids to Regulate Gene Expression and Promote Stem Elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef]
- Hung, K.T.; Kao, C.H. Hydrogen peroxide is necessary for abscisic acid-induced senescence of rice leaves. J. Plant Physiol. 2004, 161, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- van der Graaff, E.; Schwacke, R.; Schneider, A.; Desimone, M.; Flügge, U.I.; Kunze, R. Transcription analysis of arabidopsis membrane transporters and hormone pathways during developmental and induced leaf senescence. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, J.L.; Thomas, H. Leaf Senescence. In Nucleic Acids and Proteins in Plants I: Structure, Biochemistry and Physiology of Proteins; Boulter, D., Parthier, B., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 592–636. [Google Scholar]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.K.; Hickman, R.; Hill, C.; Kiddle, S.J.; Kim, Y.-S.; Penfold, C.A.; Jenkins, D.J.; et al. High-Resolution Temporal Profiling of Transcripts during Arabidopsis Leaf Senescence Reveals a Distinct Chronology of Processes and Regulation[C][W][OA]. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [PubMed]
- Sağlam-Çağ, S. The Effect of Epibrassinolide on Senescence in Wheat Leaves. Biotechnol. Biotechnol. Equip. 2007, 21, 63–65. [Google Scholar] [CrossRef]
- Lin, M.; Pang, C.; Fan, S.; Song, M.; Wei, H.; Yu, S. Global analysis of the Gossypium hirsutum L. Transcriptome during leaf senescence by RNA-Seq. BMC Plant Biol. 2015, 15, 43. [Google Scholar] [CrossRef] [PubMed]
- Borrill, P.; Harrington, S.A.; Simmonds, J.; Uauy, C. Identification of Transcription Factors Regulating Senescence in Wheat through Gene Regulatory Network Modelling. Plant Physiol. 2019, 180, 1740–1755. [Google Scholar] [CrossRef] [PubMed]
- Schippers, J.H.M.; Schmidt, R.; Wagstaff, C.; Jing, H.-C. Living to Die and Dying to Live: The Survival Strategy behind Leaf Senescence. Plant Physiol. 2015, 169, 914–930. [Google Scholar] [CrossRef]
- Schippers, J.H.M. Transcriptional networks in leaf senescence. Curr. Opin. Plant Biol. 2015, 27, 77–83. [Google Scholar] [CrossRef]
- Bresson, J.; Bieker, S.; Riester, L.; Doll, J.; Zentgraf, U. A guideline for leaf senescence analyses: From quantification to physiological and molecular investigations. J. Exp. Bot. 2018, 69, 769–786. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef]
- Guo, W.F.; Liu, D.C.; Yang, L.; Zhuang, X.; Zhang, J.J.; Wang, S.S.; Liu, Y. Cloning and Expression Analysis of New Stress-resistant NAC83 Gene from Citrus. Acta Hortic. Sin. 2015, 42, 445–454. [Google Scholar]
- Mao, Y.; Xu, J.; Wang, Q.; Li, G.; Tang, X.; Liu, T.; Feng, X.; Wu, F.; Li, M.; Xie, W.; et al. A natural antisense transcript acts as a negative regulator for the maize drought stress response gene ZmNAC48. J. Exp. Bot. 2021, 72, 2790–2806. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, C.; Xue, F.; Zhang, H.; Ji, W. Wheat NAC transcription factor TaNAC29 is involved in response to salt stress. Plant Physiol. Biochem. 2015, 96, 356–363. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y. Overexpression of TaNAC2D Displays Opposite Responses to Abiotic Stresses between Seedling and Mature Stage of Transgenic Arabidopsis. Front. Plant Sci. 2016, 7, 1754. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Zhang, G.; Liu, X.-Y.; Deng, L.; Cai, G.-L.; Zhang, Y.; Wang, X.-J.; Zhao, J.; Huang, L.-L.; Kang, Z.-S. Characterization of a novel wheat NAC transcription factor gene involved in defense response against stripe rust pathogen infection and abiotic stresses. Mol. Biol. Rep. 2010, 37, 3703–3712. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Fan, S.; Song, M.; Pang, C.; Guo, Y.; Li, L.; Wei, H.; Yu, S. Characterization and Function Analysis of GhNAC78,a Transcription Factor Gene in Upland Cotton(Gossypium hirsutum L.). Cotton Sci. 2014, 140, 270–273. [Google Scholar]
- Wang, F.; Hou, X.; Tang, J.; Wang, Z.; Wang, S.; Jiang, F.; Li, Y. A novel cold-inducible gene from Pak-choi (Brassica campestris ssp. chinensis), BcWRKY46, enhances the cold, salt and dehydration stress tolerance in transgenic tobacco. Mol. Biol. Rep. 2012, 39, 4553–4564. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Xu, X.Y.; Yu, D.Q.; Li, G.X.; Zhang, S.Q.; Zheng, S.J. Transcription factor WRKY46 regulates osmotic stress responses and stomatal movement independently in Arabidopsis. Plant J. 2014, 79, 13–27. [Google Scholar] [CrossRef]
- Hu, Y.; Dong, Q.; Yu, D. Arabidopsis WRKY46 coordinates with WRKY70 and WRKY53 in basal resistance against pathogen Pseudomonas syringae. Plant Sci. 2012, 185–186, 288–297. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Z.; Dong, J.; Wang, T. Overexpression of MtWRKY76 increases both salt and drought tolerance in Medicago truncatula. Environ. Exp. Bot. 2016, 123, 50–58. [Google Scholar] [CrossRef]
- Raineri, J.; Ribichich, K.F.; Chan, R.L. The sunflower transcription factor HaWRKY76 confers drought and flood tolerance to Arabidopsis thaliana plants without yield penalty. Plant Cell Rep. 2015, 34, 2065–2080. [Google Scholar] [CrossRef]
- Robatzek, S.; Somssich, I.E. Targets of AtWRKY6 regulation during plant senescence and pathogen defense. Genes Dev. 2002, 16, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-H.; Yie, S.W.; Hwang, D.-J. Heterologous expression of OsWRKY6 gene in Arabidopsis activates the expression of defense related genes and enhances resistance to pathogens. Plant Sci. 2011, 181, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Robatzek, S.; Somssich, I.E. A new member of the Arabidopsis WRKY transcription factor family, AtWRKY6, is associated with both senescence- and defence-related processes. Plant J. 2001, 28, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Knoth, C.; Ringler, J.; Dangl, J.L.; Eulgem, T. Arabidopsis WRKY70 is required for full RPP4-mediated disease resistance and basal defense against Hyaloperonospora parasitica. Mol. Plant-Microbe Interact. 2007, 20, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Kariola, T.; Palva, E.T. WRKY70 modulates the selection of signaling pathways in plant defense. Plant J. 2006, 46, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 Transcription Factors Are Involved in Brassinosteroid-Regulated Plant Growth and Drought Responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wei, Y.; Deng, Z.; Dai, L.; Yang, H.; Dejun, L.I. Cloning and Expression Analysis of a WRKY Transcription Factor Gene HbWRKY41 in Hevea brasiliensis. Acta Agric. Boreali-Sin. 2017, 32, 91–96. [Google Scholar]
- Chen, Y.-S.; David Ho, T.-H.; Liu, L.; Lee Ding, H.; Lee, C.-H.; Chen, Y.-R.; Lin, S.-Y.; Lu, C.-A.; Yu, S.-M. Sugar starvation-regulated MYBS2 and 14-3-3 protein interactions enhance plant growth, stress tolerance, and grain weight in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Chao, Y.-C.; Tseng, T.-W.; Huang, C.-K.; Lo, P.-C.; Lu, C.-A. Two MYB-related transcription factors play opposite roles in sugar signaling in Arabidopsis. Plant Mol. Biol. 2017, 93, 299–311. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agronomic Traits | Wild Type | Mutant |
|---|---|---|
| Effective tiller/plant | 6.4 ± 1.96 | 2.60 ± 0.49 * |
| Plant height (cm) | 62.12 ± 3.3 | 49.40 ± 2.87 * |
| NO. of upper sterile spikelets | 0 ± 0 | 0.20 ± 0.4 |
| NO. of lower sterile spikelets | 0 ± 0 | 1.60 ± 0.49 * |
| Panicle length (cm) | 14.56 ± 0.29 | 9.78 ± 0.72 * |
| NO.of grains per panicle | 100.40 ± 9.02 | 33.80 ± 2.79 * |
| NO. of fruiting spikelets | 23.40 ± 0.49 | 16.60 ± 0.80 * |
| Main spike grain weight (g) | 2.93 ± 0.55 | 0.76 ± 0.23 * |
| Grain yield per plant (g) | 13.75 ± 4.40 | 2.49 ± 0.73 * |
| 1000-grainweight (g) | 30.16 ± 0.53 | 14.77 ± 0.33 * |
| Flag leaf length (cm) | 15.75 ± 1.89 | 16.06 ± 1.28 |
| Flag leaf width (cm) | 1.5 ± 0.16 | 1.32 ± 0.12 * |
| Second leaf length (cm) | 20.66 ± 2.64 | 20.44 ± 1.30 |
| Second leaf width (cm) | 1.38 ± 0.11 | 1.22 ± 0.07 * |
| Varirty | Curve Parameters | Characteristic | |||||
|---|---|---|---|---|---|---|---|
| k | a | b | MRS | TMRS/d | Ts/d | To/d | |
| GSm | 63.047 | 0.1047 | −0.1404 | 3.26 | 16.07 | 2.84 | 24.13 |
| Chang6878 | 57.781 | 1.82 × 10−6 | −0.2618 | 5.56 | 50.48 | 39.13 | 54.67 |
| DEG-Set | Total DEG | Up-Regulated | Down-Regulated |
|---|---|---|---|
| WS2_VS_MS2 | 18,246 | 10,801 | 7445 |
| MS1_VS_MS2 | 15,180 | 8246 | 6934 |
| WS1_VS_MS1 | 7030 | 5010 | 2020 |
| WS1_VS_WS2 | 11,744 | 5145 | 6599 |
| Sample Pair | Class | Annotation | GO ID | p-Value |
|---|---|---|---|---|
| WS1_VS_WS2 | Biological process | maltose metabolic process | GO:0000023 | 1.19 × 10−7 |
| tertiary alcohol metabolic process | GO:1902644 | 3.1 × 10−7 | ||
| Cellular component | plastoglobule | GO:0010287 | 5.92 × 10−7 | |
| cytoskeleton | GO:0005856 | 1.13 × 10−6 | ||
| Molecular function | aromatic amino acid transmembrane transporter activity | GO:0015173 | 1.19 × 10−7 | |
| fructosyltransferase activity | GO:0050738 | 2.78 × 10−7 | ||
| MS2_VS_WS2 | Biological process | photosystem II oxygen evolving complex assembly | GO:0010270 | 4.55 × 10−8 |
| xanthophyll biosynthetic process | GO:0016123 | 1.5 × 10−7 | ||
| Cellular component | cell surface | GO:0009986 | 6.12 × 10−8 | |
| chloroplast thylakoid membrane protein complex | GO:0098807 | 9.89 × 10−8 | ||
| Molecular function | magnesium chelatase activity | GO:0016851 | 4.55 × 10−8 | |
| malate transmembrane transporter activity | GO:0015140 | 4.55 × 10−8 | ||
| MS1_VS_WS1 | Biological process | S-adenosylhomocysteine metabolic process | GO:0046498 | 4.58 × 10−8 |
| S-adenosylhomocysteine catabolic process | GO:0019510 | 4.58 × 10−8 | ||
| Cellular component | COPI-coated vesicle membrane | GO:0030663 | 7.61 × 10−8 | |
| oligosaccharyltransferase complex | GO:0008250 | 1.63 × 10−7 | ||
| Molecular function | asparaginase activity | GO:0004067 | 7.61 × 10−8 | |
| protein disulfide isomerase activity | GO:0003756 | 3.15 × 10−7 | ||
| MS1_VS_MS2 | Biological process | xanthophyll metabolic process | GO:0016122 | 1.45 × 10−7 |
| fructose metabolic process | GO:0006000 | 2.16 × 10−7 | ||
| Cellular component | chloroplast thylakoid membrane protein complex | GO:0098807 | 1.62 × 10−7 | |
| photosystem I reaction center | GO:0009538 | 1.66 × 10−7 | ||
| Molecular function | fructosyltransferase activity | GO:0050738 | 7.88 × 10−8 | |
| fructose 1,6-bisphosphate 1-phosphatase activity | GO:0042132 | 1.62 × 10−7 |
| Sample Pair | Pathway | Ko ID | p-Value |
|---|---|---|---|
| WS1_VS_WS2 | Starch and sucrose metabolism | map00500 | 8.02 × 10−11 |
| Glycerophospholipid metabolism | map00564 | 5.49 × 10−11 | |
| Carotenoid biosynthesis | map00906 | 1.9 × 10−10 | |
| MAPK signaling pathway—plant | map04016 | 1.12 × 10−8 | |
| Glycine, serine and threonine metabolism | map00260 | 6.23 × 10−8 | |
| MS2_VS_WS2 | Photosynthesis—antenna proteins | map00196 | 9.69 × 10−45 |
| Porphyrin and chlorophyll metabolism | map00860 | 4.92 × 10−24 | |
| Glyoxylate and dicarboxylate metabolism | map00630 | 2.83 × 10−14 | |
| Carbon fixation in photosynthetic organisms | map00710 | 8.26 × 10−14 | |
| Carotenoid biosynthesis | map00906 | 3.22 × 10−11 | |
| MS1_VS_WS1 | Protein processing in endoplasmic reticulum | map04141 | 3.25 × 10−25 |
| Endocytosis | map04144 | 8.51 × 10−14 | |
| Amino sugar and nucleotide sugar metabolism | map00520 | 2 × 10−13 | |
| N-Glycan biosynthesis | map00510 | 3.3 × 10−11 | |
| Various types of N-glycan biosynthesis | map00513 | 1.09 × 10−10 | |
| MS1_VS_MS2 | Photosynthesis—antenna proteins | map00196 | 2.23 × 10−44 |
| Carbon fixation in photosynthetic organisms | map00710 | 1.9 × 10−25 | |
| Starch and sucrose metabolism | map00500 | 3.75 × 10−25 | |
| Glyoxylate and dicarboxylate metabolism | map00630 | 3.15 × 10−20 | |
| Porphyrin and chlorophyll metabolism | map00860 | 5.48 × 10−19 |
| Sample Pair | UP TF Number | DOWN TF Number | ALL TF Number |
|---|---|---|---|
| WS1_VS_WS2 | 96 | 262 | 358 |
| MS2_VS_WS2 | 358 | 162 | 520 |
| MS1_VS_WS1 | 111 | 70 | 181 |
| MS1_VS_MS2 | 256 | 238 | 494 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Sun, L.; Jin, X.; Islam, M.A.; Guo, F.; Tang, X.; Zhao, K.; Hao, H.; Li, N.; Zhang, W.; et al. Analysis of Physiological and Transcriptomic Differences between a Premature Senescence Mutant (GSm) and Its Wild-Type in Common Wheat (Triticum aestivum L.). Biology 2022, 11, 904. https://doi.org/10.3390/biology11060904
Lu J, Sun L, Jin X, Islam MA, Guo F, Tang X, Zhao K, Hao H, Li N, Zhang W, et al. Analysis of Physiological and Transcriptomic Differences between a Premature Senescence Mutant (GSm) and Its Wild-Type in Common Wheat (Triticum aestivum L.). Biology. 2022; 11(6):904. https://doi.org/10.3390/biology11060904
Chicago/Turabian StyleLu, Juan, Lili Sun, Xiujuan Jin, Md Ashraful Islam, Feng Guo, Xiaosha Tang, Kai Zhao, Huifang Hao, Ning Li, Wenjun Zhang, and et al. 2022. "Analysis of Physiological and Transcriptomic Differences between a Premature Senescence Mutant (GSm) and Its Wild-Type in Common Wheat (Triticum aestivum L.)" Biology 11, no. 6: 904. https://doi.org/10.3390/biology11060904