
Comprehensive Review of the Components in Cat’s Claw (Uncaria tomentosa) and Their Antibacterial Activity


Abstract
:1. Introduction
2. Methodology
3. Alkaloids
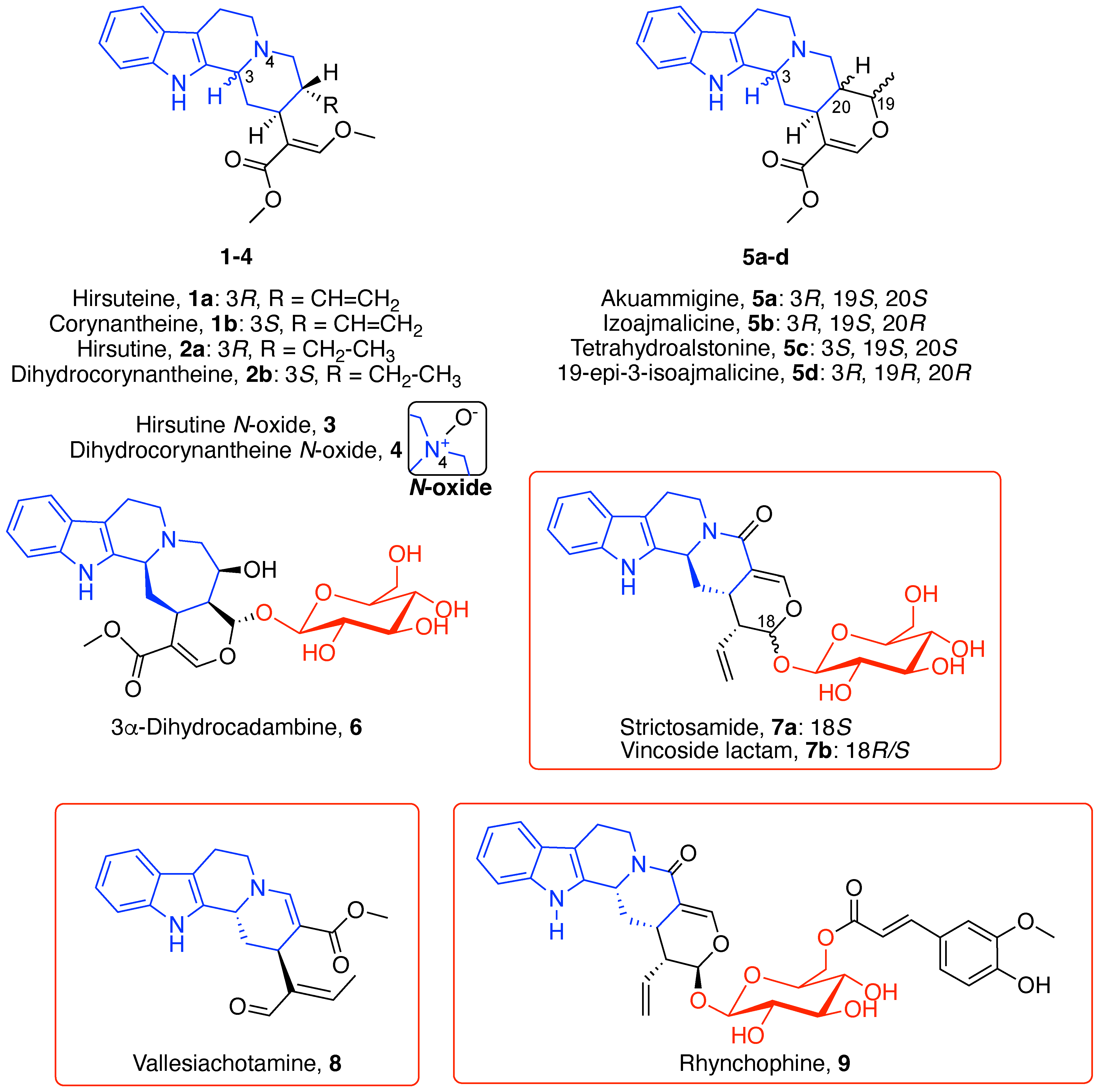
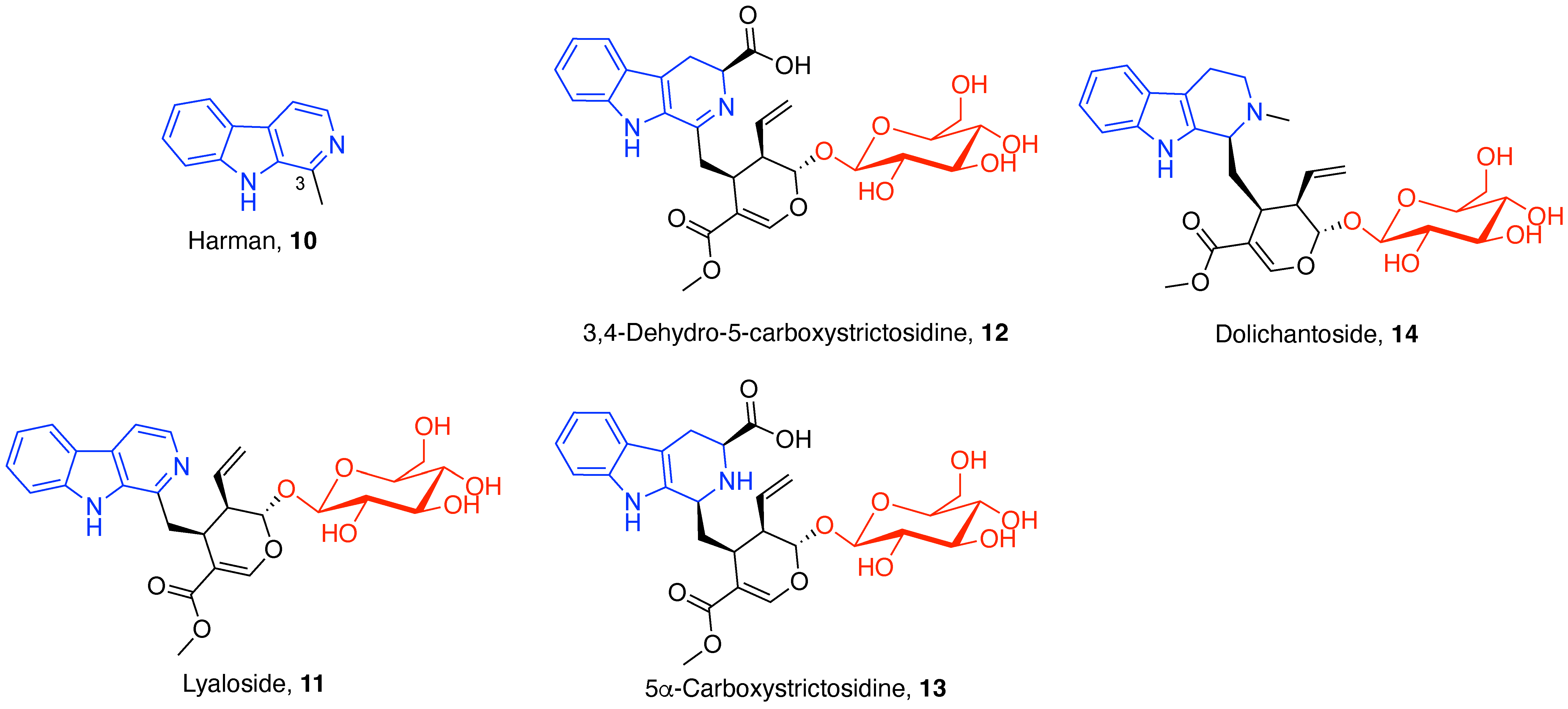
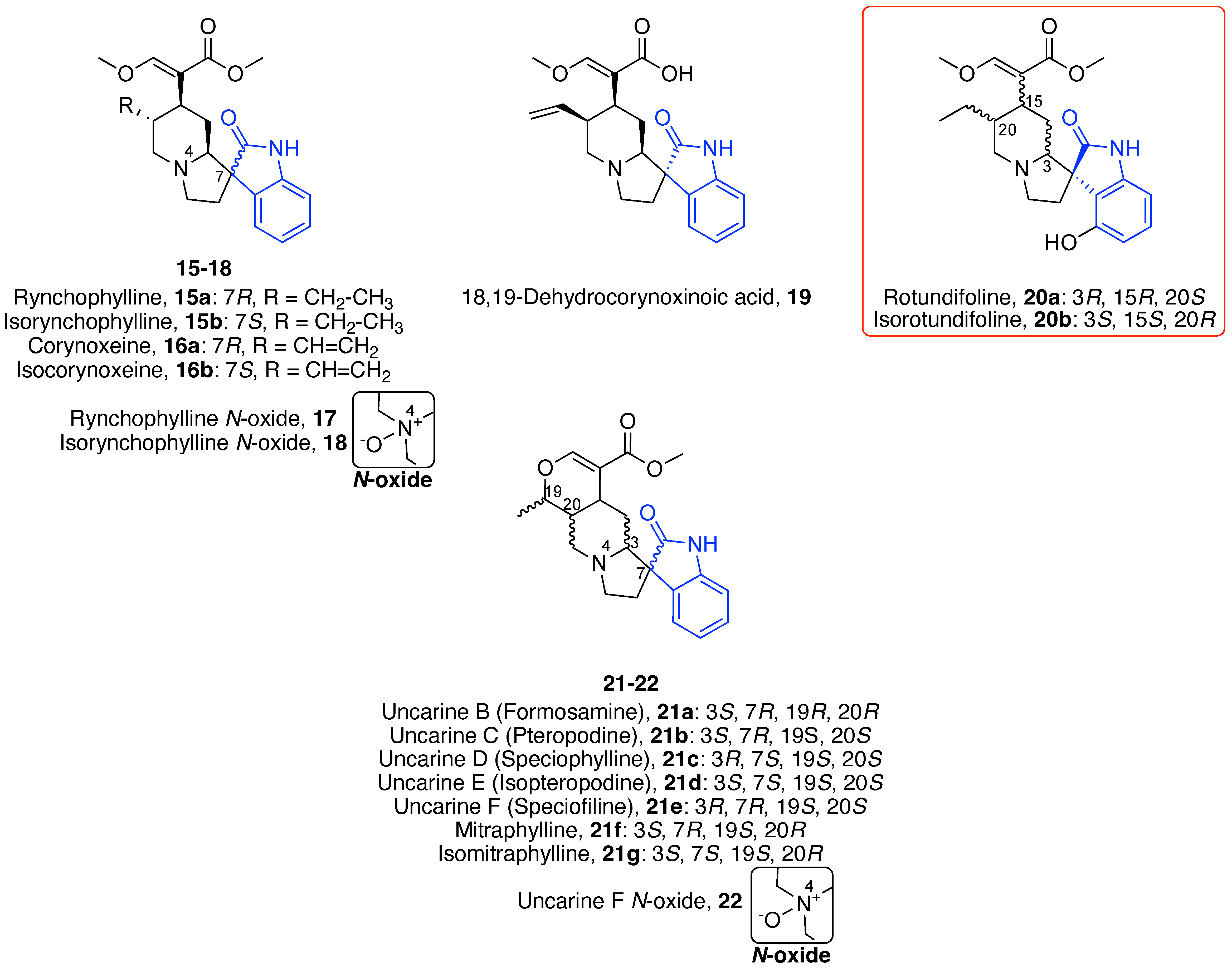
3.1. Indole Alkaloids
3.1.1. Tetracyclic Indoloquinolizidine Alkaloids
3.1.2. Pentacyclic Indoloquinolizidine Alkaloids
3.1.3. β-Carboline Alkaloids
3.2. Oxindole Alkaloids
3.2.1. Tetracyclic Oxindole Alkaloids
3.2.2. Pentacyclic Oxindole Alkaloids
4. Polyphenols
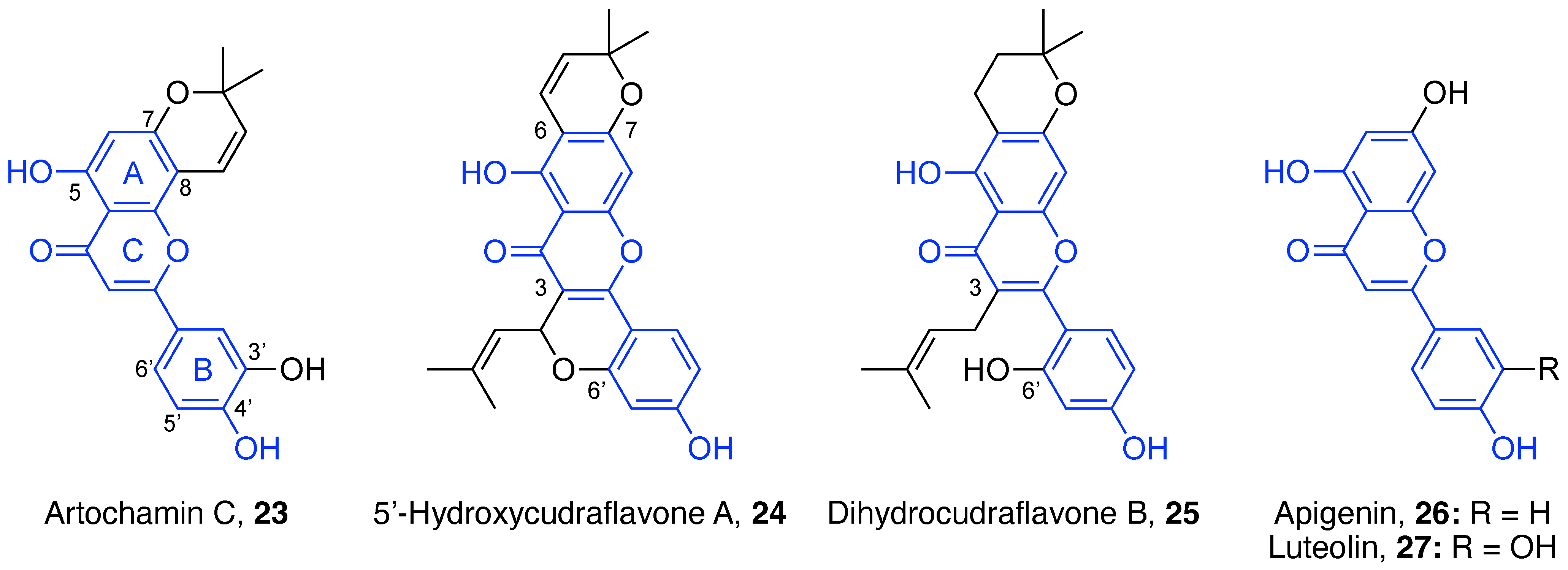
4.1. Flavonoids
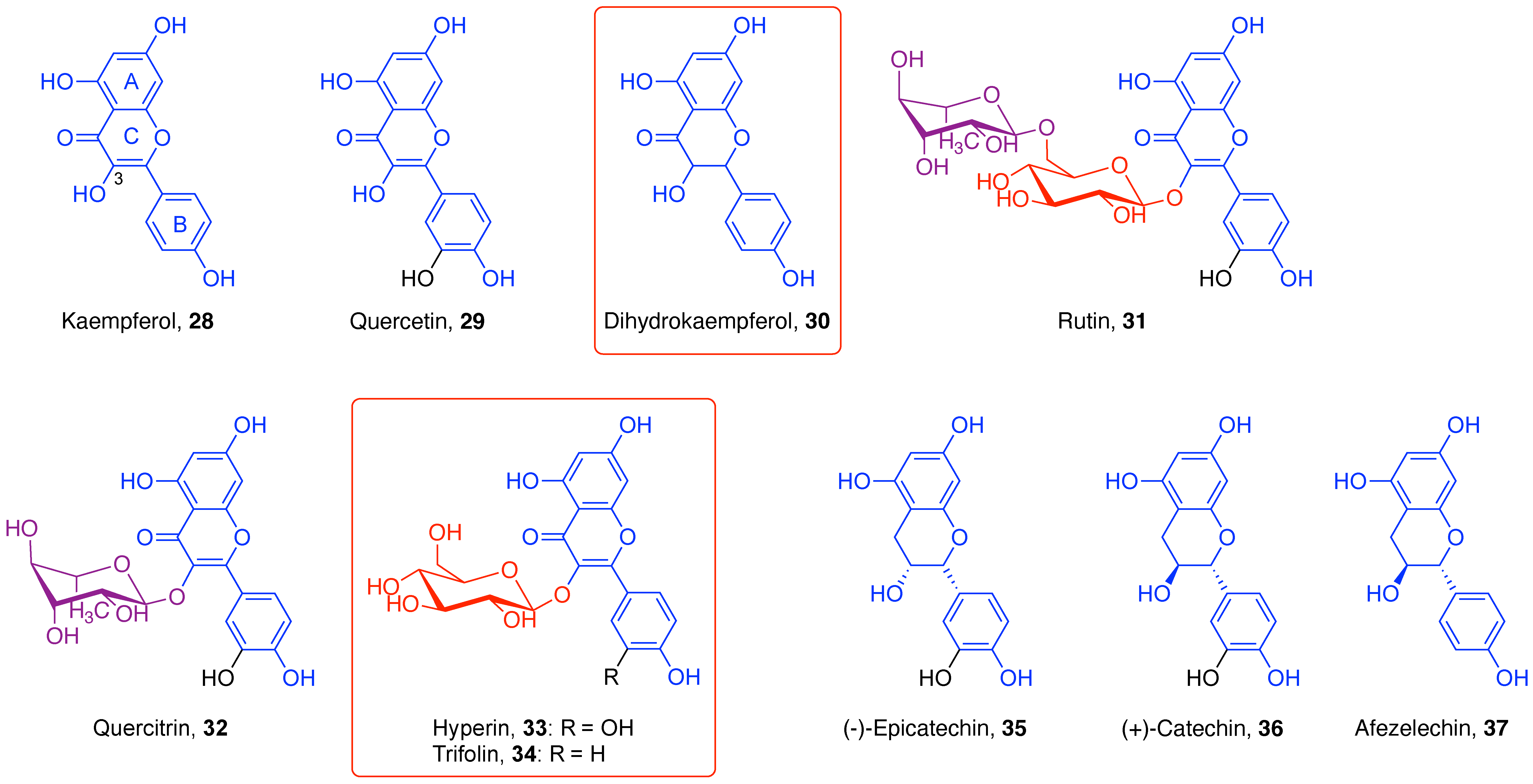
4.1.1. Flavones
4.1.2. Flavonols
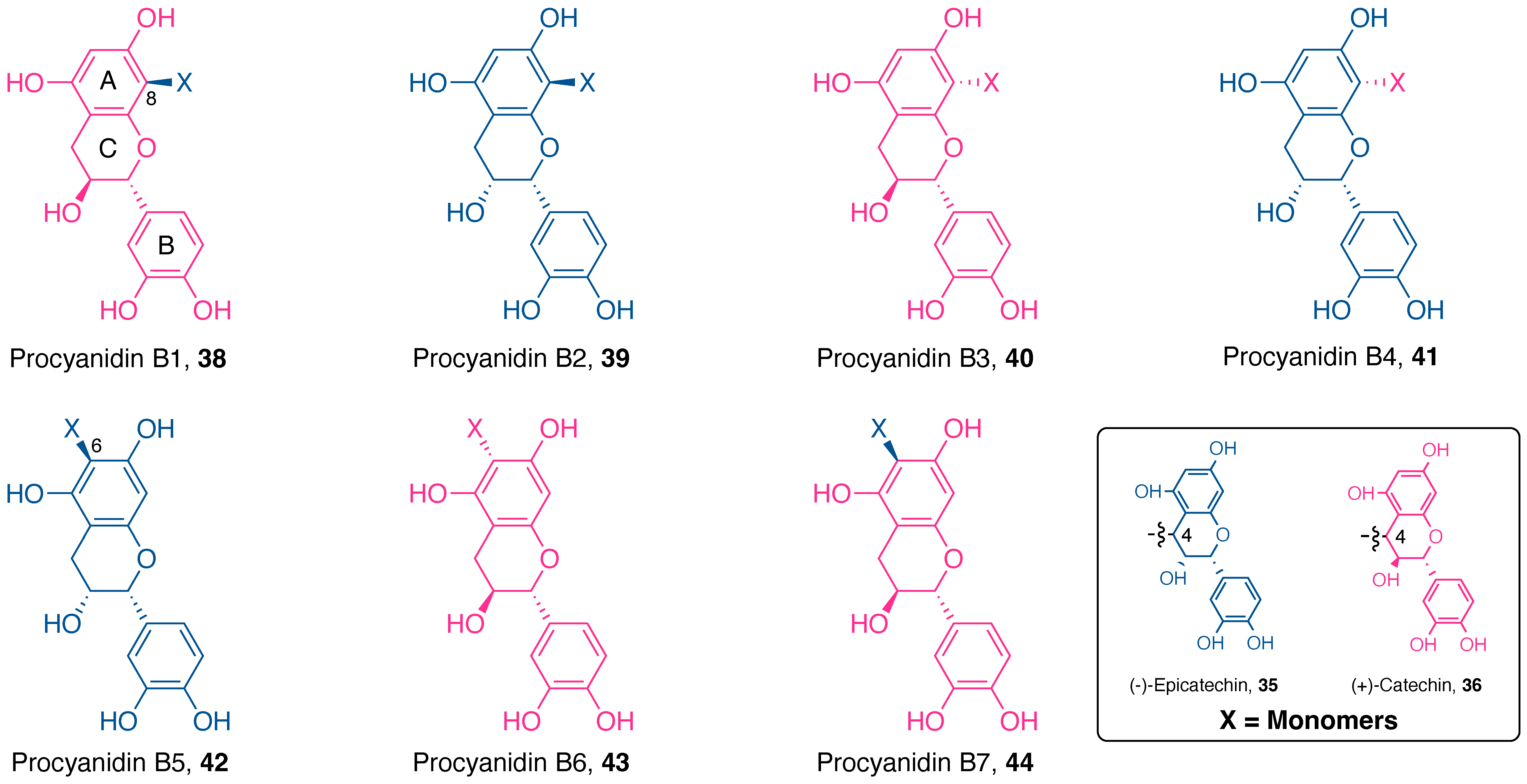
4.1.3. Flavan-3-ols
Procyanidin Type
Procyanidin Dimers
Procyanidin Trimers
Propelargonidin Type
Proanthocyanidins Polymeric Structures (DP > 3)
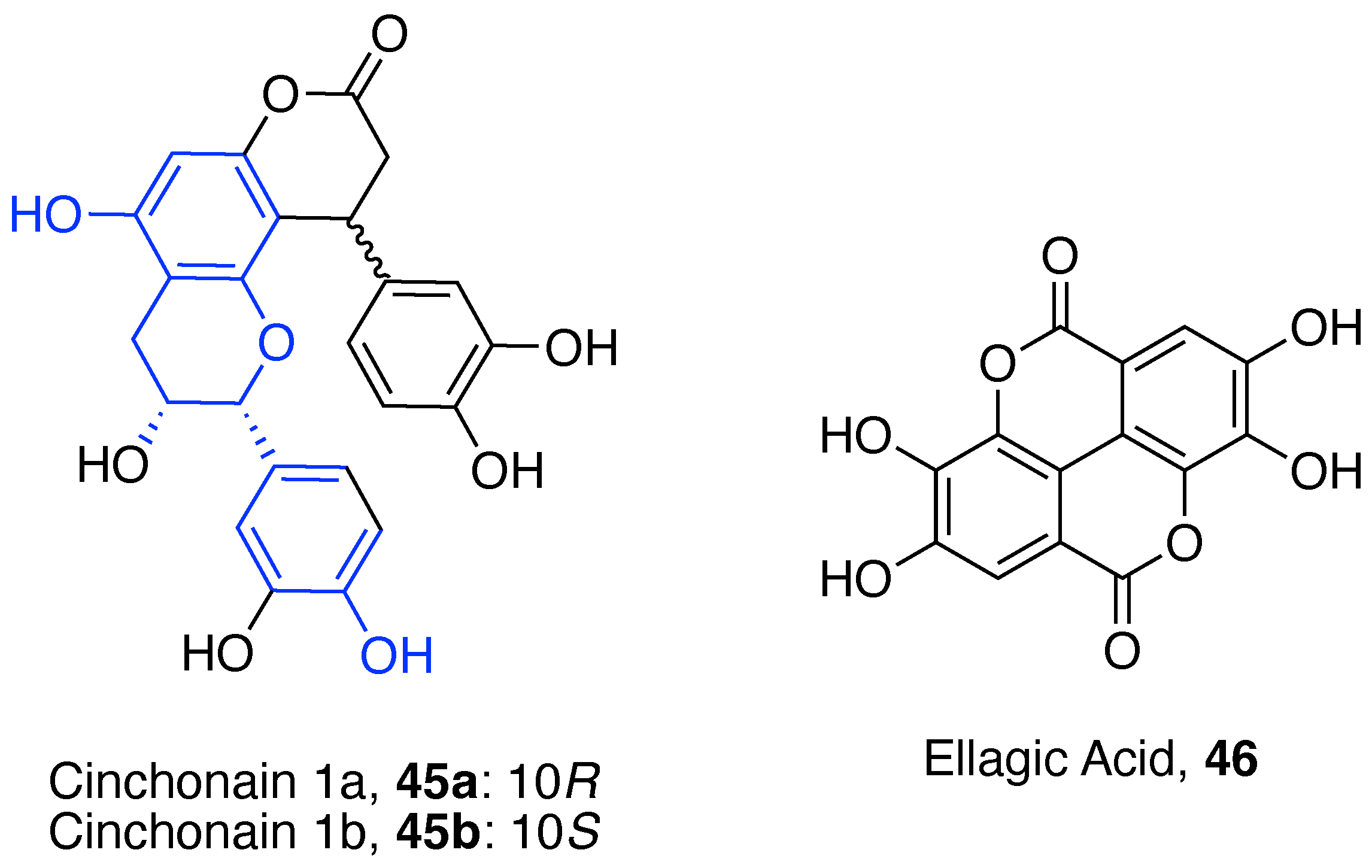
4.2. Miscellaneous Polyphenols
4.3. Phenolic Acids
4.3.1. Hydroxybenzoic Acids
4.3.2. Hydroxycinnamic Acids
5. Terpenes
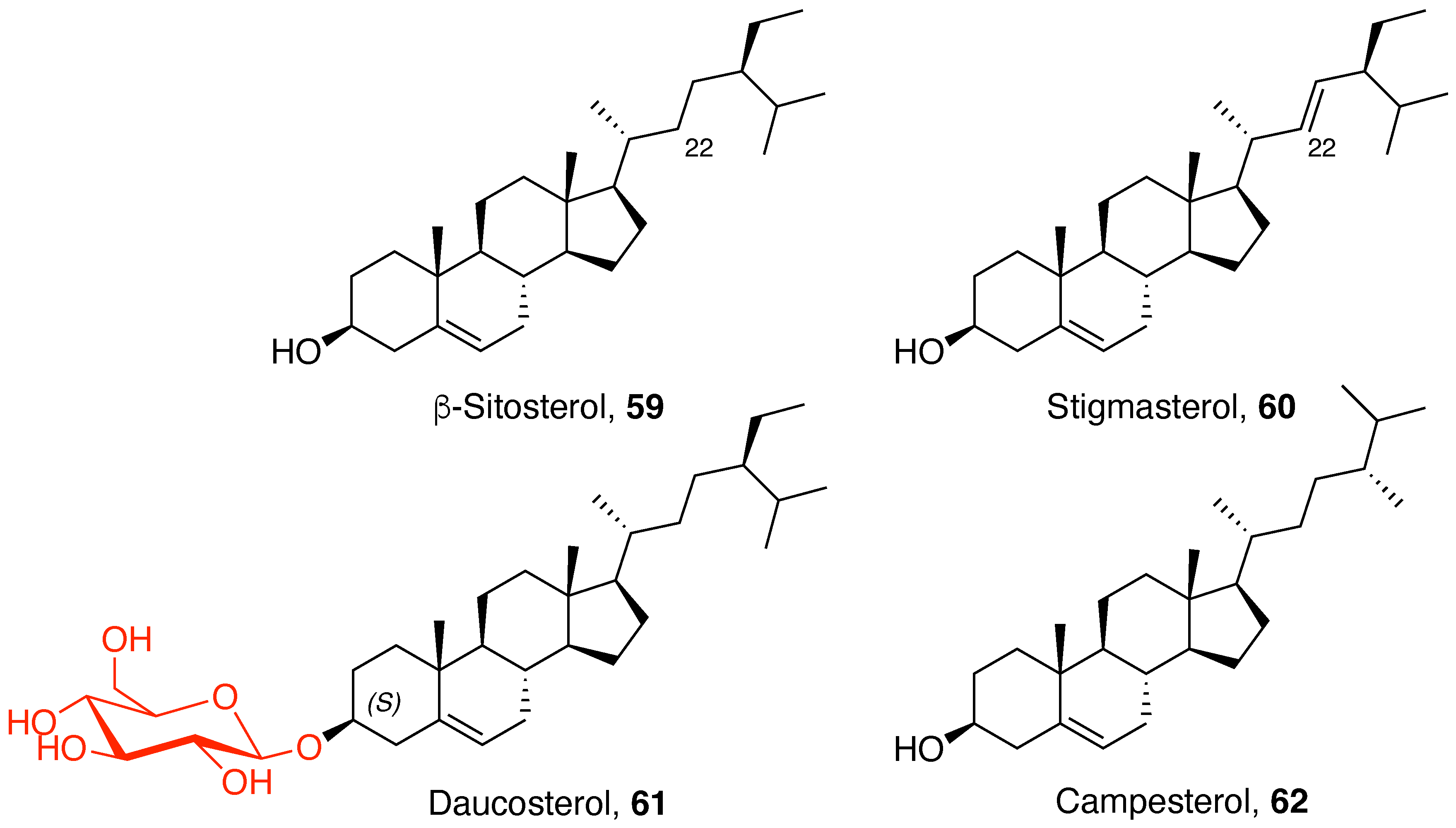
5.1. Phytosterols
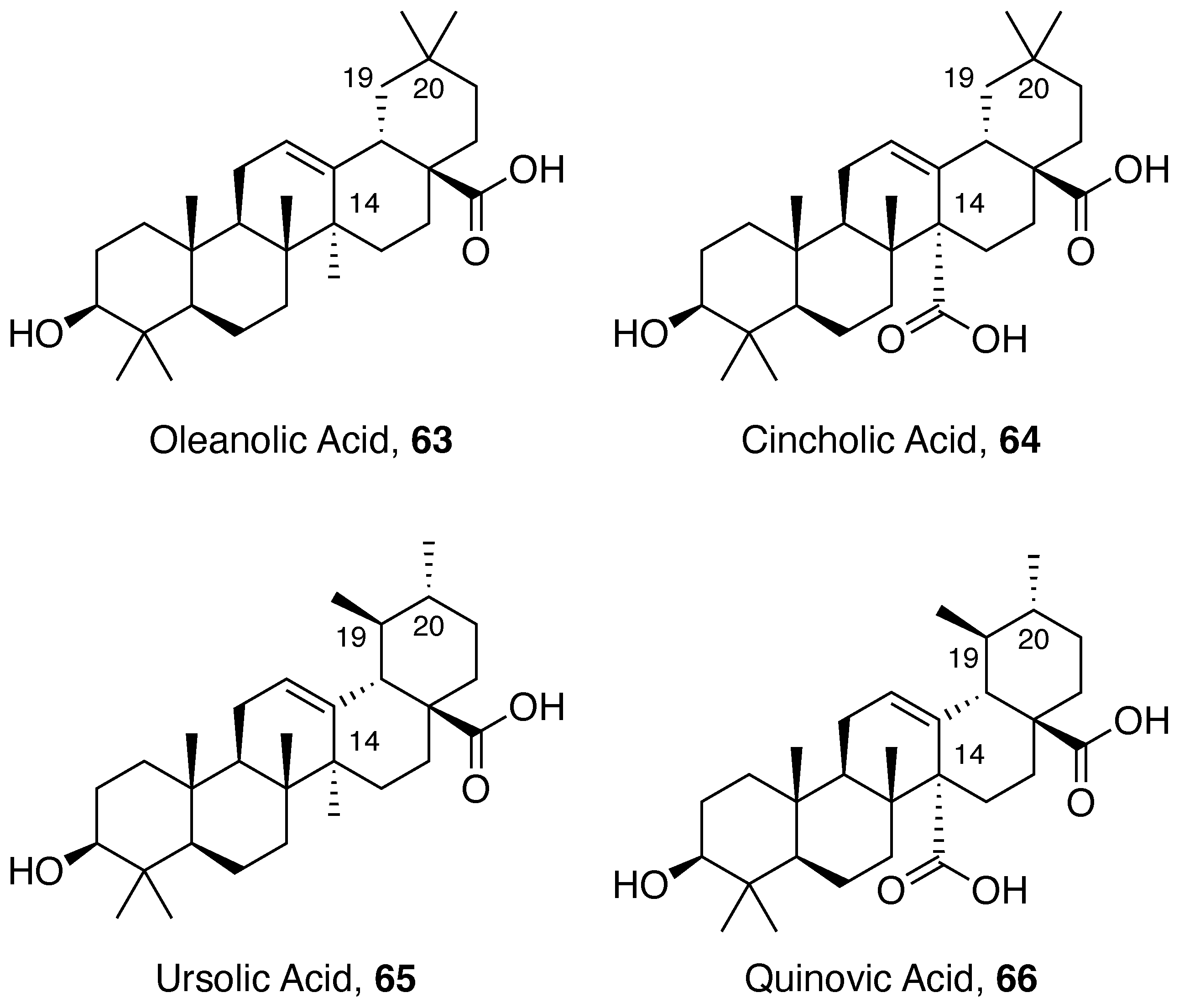
5.2. Pentacyclic Triterpene Acids
5.2.1. Ursolic Acid Derivatives
5.2.2. Glycosylated Pentacyclic Triterpene Acids
Cincholic Acid Derivatives
Quinovic Acid Derivatives
Pyroquinovic and Pyrocincholic Acid Derivatives
5.3. Miscellaneous Terpenoid
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Navarro Hoyos, M.; Sánchez-Patán, F.; Murillo Masis, R.; Martín-Álvarez, P.; Zamora Ramirez, W.; Monagas, M.; Bartolomé, B. Phenolic Assesment of Uncaria tomentosa L. (Cat’s Claw): Leaves, Stem, Bark and Wood Extracts. Molecules 2015, 20, 22703–22717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Hoyos, M.; Lebrón-Aguilar, R.; Quintanilla-López, J.E.; Cueva, C.; Hevia, D.; Quesada, S.; Azofeifa, G.; Moreno-Arribas, M.; Monagas, M.; Bartolomé, B. Proanthocyanidin characterization and bioactivity of extracts from different parts of Uncaria tomentosa L. (cat’s claw). Antioxidants 2017, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, M.; Yokoya, M.; Takayama, H.; Aimi, N. Co-occurrence of Harman and β-Carboline-type Monoterpenoid Glucoindole Alkaloids in Una de Gato (Uncaria tomentosa). Nat. Med. 2001, 55, 308–310. [Google Scholar]
- Kitajima, M.; Hashimoto, K.-I.; Yokoya, M.; Takayama, H.; Aimi, N.; Sakai, S.-I. A New Gluco Indole Alkaloid, 3, 4-Dehydro-5-carboxystrictosidine, from Peruvian Una de Gato (Uncaria tomentosa). Chem. Pharm. Bull. 2000, 48, 1410–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, M.; Hashimoto, K.-I.; Sandoval, M.; Aimi, N.; Takayama, H. New Oleanan-Type Triterpene and Cincholic Acid Glycosides from Peruvian “Uña de Gato” (Uncaria tomentosa). Chem. Pharm. Bull. 2004, 52, 1258–1261. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, M.; Yokoya, M.; Takayama, H.; Aimi, N. Synthesis and Absolute Configuration of a New 3,4-Dihydro-β-carboline-Type Alkaloid, 3,4-Dehydro-5(S)-5-carboxystrictosidine, Isolated from Peruvian Una de Gato (Uncaria tomentosa). Chem. Pharm. Bull. 2002, 50, 1376–1378. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, M.; Hashimoto, K.-i.; Yokoya, M.; Takayama, H.; Aimi, N. Two New 19-Hydroxyursolic Acid-type Triterpenes from Peruvian ‘Uña de Gato’ (Uncaria tomentosa). Tetrahedron 2000, 56, 547–552. [Google Scholar] [CrossRef]
- Kitajima, M.; Hashimoto, K.-I.; Yokoya, M.; Takayama, H.; Sandoval, M.; Aimi, N. Two New Nor-triterpene Glycosides from Peruvian “Uña de Gato” (Uncaria tomentosa). J. Nat. Prod. 2003, 66, 320–323. [Google Scholar] [CrossRef]
- Laus, G.; Brössner, D.; Keplinger, K. Alkaloids of peruvian Uncaria tomentosa. Phytochemistry 1997, 45, 855–860. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, J.J.; Xu, J.; Feng, F.; Qu, W. Medicinal uses, phytochemistry and pharmacology of the genus Uncaria. J. Ethnopharmacol. 2015, 173, 48–80. [Google Scholar] [CrossRef] [PubMed]
- Heitzman, M.E.; Neto, C.C.; Winiarz, E.; Vaisberg, A.J.; Hammond, G.B. Ethnobotany, phytochemistry and pharmacology of Uncaria (Rubiaceae). Phytochemistry 2005, 66, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Magdy Beshbishy, A.; Wasef, L.; Elewa, Y.H.; El-Hack, A.; Mohamed, E.; Taha, A.E.; Al-Sagheer, A.A.; Devkota, H.P.; Tufarelli, V. Uncaria tomentosa (Willd. ex Schult.) DC.: A Review on Chemical Constituents and Biological Activities. Appl. Sci. 2020, 10, 2668. [Google Scholar] [CrossRef] [Green Version]
- Urdanibia, I.; Taylor, P. Uncaria tomentosa (Willd. ex Schult.) DC. and Uncaria guianensis (Aubl.) JF Gmell. In Medicinal and Aromatic Plants of South America; Springer: Berlin/Heidelberg, Germany, 2018; pp. 453–463. [Google Scholar]
- Herrera, D.R.; Tay, L.Y.; Rezende, E.C.; Kozlowski, V.A., Jr.; dos Santos, E.B. In vitro antimicrobial activity of phytotherapic Uncaria tomentosa against endodontic pathogens. J. Oral Sci. 2010, 52, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Herrera, D.R.; Durand-Ramirez, J.E.; Falcao, A.; SILVA, E.J.L.N.d.; SANTOS, E.B.d.; GOMES, B.P.F.d.A. Antimicrobial activity and substantivity of Uncaria tomentosa in infected root canal dentin. Braz. Oral Res. 2016, 30, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ccahuana-Vasquez, R.A.; Santos, S.S.F.d.; Koga-Ito, C.Y.; Jorge, A.O.C. Antimicrobial activity of Uncaria tomentosa against oral human pathogens. Braz. Oral Res. 2007, 21, 46–50. [Google Scholar] [CrossRef]
- Silva, D.; Ribeiro, G.E.; Rufino, L.R.A.; Oliveira, N.M.S.; Fiorini, J.E. Atividade Antimicrobiana da Uncaria tomentosa (Willd) DC. J. Basic Appl. Pharm. Sci. 2014, 35, 53–57. [Google Scholar]
- Kloucek, P.; Polesny, Z.; Svobodova, B.; Vlkova, E.; Kokoska, L. Antibacterial screening of some Peruvian medicinal plants used in Calleria District. J. Ethnopharmacol. 2005, 99, 309–312. [Google Scholar] [CrossRef]
- Datar, A.N.; Kaur, N.; Patel, S.; Luecke, D.F.; Sapi, E. In Vitro Effectiveness of Samento and Banderol Herbal Extracts on Different Morphological Forms of Borrelia Burgdorferi; University of New Haven: West Haven, CT, USA, 2010. [Google Scholar]
- Feng, J.; Leone, J.; Schweig, S.; Zhang, Y. Evaluation of Natural and Botanical Medicines for Activity against Growing and Non-growing Forms of B. burgdorferi. Front. Med. 2020, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Sadilek, A.; Hswen, Y.; Bavadekar, S.; Shekel, T.; Brownstein, J.S.; Gabrilovich, E. Lymelight: Forecasting Lyme disease risk using web search data. NPJ Digit. Med. 2020, 3, 16. [Google Scholar] [CrossRef]
- Kuehn, B.M. CDC estimates 300 000 US cases of Lyme disease annually. JAMA 2013, 310, 1110. [Google Scholar] [CrossRef] [PubMed]
- Keplinger, K.; Laus, G.; Wurm, M.; Dierich, M.P.; Teppner, H. Uncaria tomentosa (Willd.) DC.—Ethnomedicinal use and new pharmacological, toxicological and botanical results. J. Ethnopharmacol. 1998, 64, 23–34. [Google Scholar] [CrossRef]
- Reinhard, K.-H. Uncaria tomentosa (Willd.) D.C.: Cat’s Claw, Uña de Gato, or Savéntaro. J. Altern. Complement. Med. 1999, 5, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkiewicz, B.; Lukasiak, J. Vilcacora (Uncaria tomentosa (Willd.) DC. and Uncaria guianensis (Aublet) Gmell.)—A review of published scientific literature. Case Rep. Clin. Pract. Rev. 2001, 2, 305–316. [Google Scholar]
- Laus, G. Advances in chemistry and bioactivity of the genus Uncaria. Phytother. Res. 2004, 18, 259–274. [Google Scholar] [CrossRef]
- Potawale, S.E.; Mehta, U.K.; Sadiq, M.W.M. Phytopharmacology of Uncaria tomentosa: A review. Pharmacologyonline 2008, 2, 197–214. [Google Scholar]
- Andre, N.; Wang, X.; He, Y.; Pan, G.; Kojo, A.; Liu, Y. A review of the occurrence of non-alkaloid constituents in Uncaria species and their structure-activity relationships. Am. J. Biomed. Life Sci. 2013, 1, 79–98. [Google Scholar] [CrossRef]
- Ravipati, A.S.; Reddy, N.; Koyyalamudi, S.R. Chapter 13—Biologically Active Compounds from the Genus Uncaria (Rubiaceae). In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 43, pp. 381–408. [Google Scholar]
- Liang, J.-H.; Wang, C.; Huo, X.-K.; Tian, X.-G.; Zhao, W.-Y.; Wang, X.; Sun, C.-P.; Ma, X.-C. The genus Uncaria: A review on phytochemical metabolites and biological aspects. Fitoterapia 2020, 147, 104772. [Google Scholar] [CrossRef]
- Rosales, P.F.; Bordin, G.S.; Gower, A.E.; Moura, S. Indole alkaloids: 2012 until now, highlighting the new chemical structures and biological activities. Fitoterapia 2020, 143, 104558. [Google Scholar] [CrossRef]
- Cushnie, T.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef]
- Aniszewski, T. Alkaloids: Chemistry, Biology, Ecology, and Applications; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hesse, M. Alkaloids: Nature’s Curse or Blessing? John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- O’Connor, S.E.; Maresh, J.J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat. Prod. Rep. 2006, 23, 532–547. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Singh, O.M. Recent progress in biological activities of indole and indole alkaloids. Mini Rev. Med. Chem. 2018, 18, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.E.; Abdul-Hameed, Z.H.; Alotaibi, M.O.; Bawakid, N.O.; Sobahi, T.R.; Abdel-Lateff, A.; Alarif, W.M. Chemical Diversity and Bioactivities of Monoterpene Indole Alkaloids (MIAs) from Six Apocynaceae Genera. Molecules 2021, 26, 488. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, J.; Hemingway, S. Chromatographic and spectroscopic methods for the identification of alkaloids from herbarium samples of the genus Uncaria. J. Chromatogr. A 1975, 105, 163–178. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Gloriozova, T.A.; Poroikov, V.V. Naturally occurring plant isoquinoline N-oxide alkaloids: Their pharmacological and SAR activities. Phytomedicine 2015, 22, 183–202. [Google Scholar] [CrossRef]
- Phillipson, J.D.; Hemingway, S.R.; Ridsdale, C.E. Alkaloids of Uncaria. V. Their Occurrence and Chemotaxonomy; University of the Philippines: Quezon City, Philippines, 1978. [Google Scholar]
- Mfuh, A.M.; Larionov, O.V. Heterocyclic N-Oxides—An Emerging Class of Therapeutic Agents. Curr. Med. Chem. 2015, 22, 2819–2857. [Google Scholar] [CrossRef]
- Ihsan, A.; Wang, X.; Tu, H.G.; Zhang, W.; Dai, M.H.; Peng, D.P.; Wang, Y.L.; Huang, L.L.; Chen, D.M.; Mannan, S.; et al. Genotoxicity evaluation of Mequindox in different short-term tests. Food Chem. Toxicol. 2013, 51, 330–336. [Google Scholar] [CrossRef]
- Keri, R.A.-O.; Pandule, S.S.; Budagumpi, S.; Nagaraja, B.M. Quinoxaline and quinoxaline-1,4-di-N-oxides: An emerging class of antimycobacterials. Arch. Pharm. 2018, 351, 1700325. [Google Scholar] [CrossRef]
- Cheng, G.; Li, B.; Wang, C.; Zhang, H.; Liang, G.; Weng, Z.; Hao, H.; Wang, X.; Liu, Z.; Dai, M.; et al. Systematic and Molecular Basis of the Antibacterial Action of Quinoxaline 1,4-Di-N-Oxides against Escherichia coli. PLoS ONE 2015, 10, e0136450. [Google Scholar] [CrossRef] [Green Version]
- Shamma, M.; Richey, J.M. The stereochemistry of the heteroyohimbine alkaloids. J. Am. Chem. Soc. 1963, 85, 2507–2512. [Google Scholar] [CrossRef]
- Qin, N.; Lu, X.; Liu, Y.; Qiao, Y.; Qu, W.; Feng, F.; Sun, H. Recent research progress of Uncaria spp. based on alkaloids: Phytochemistry, pharmacology and structural chemistry. Eur. J. Med. Chem. 2021, 210, 112960. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Heredia, A.A.; Marín-López, R.; Ponce-Noyola, T.; Cerda-García-Rojas, C.M.; Trejo-Tapia, G.; Ramos-Valdivia, A.C. Oxidative stress induces alkaloid production in Uncaria tomentosa root and cell cultures in bioreactors. Eng. Life Sci. 2009, 9, 211–218. [Google Scholar] [CrossRef]
- Vera-Reyes, I.; Huerta-Heredia, A.A.; Ponce-Noyola, T.; Flores-Sanchez, I.J.; Esparza-García, F.; Cerda-García-Rojas, C.M.; Trejo-Tapia, G.; Ramos-Valdivia, A.C. Strictosidine-related enzymes involved in the alkaloid biosynthesis of Uncaria tomentosa root cultures grown under oxidative stress. Biotechnol. Prog. 2013, 29, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Luna-Palencia, G.; Huerta-Heredia, A.; Cerda-García-Rojas, C.; Ramos-Valdivia, A. Differential alkaloid profile in Uncaria tomentosa micropropagated plantlets and root cultures. Biotechnol. Lett. 2013, 35, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Aimi, N.; Shito, T.; Fukushima, K.; Itai, Y.; Aoyama, C.; Kunisawa, K.; Sakai, S.; Haginiwa, J.; Yamasaki, K. Studies on Plants containing Indole Alkaloids. VIII Indole Alkaloid Glycosides and Other Constituents of the Leaves of Uncaria rhynchophylla MIQ. Chem. Pharm. Bull. 1982, 30, 4046–4051. [Google Scholar] [CrossRef] [Green Version]
- Nenaah, G. Antibacterial and antifungal activities of (beta)-carboline alkaloids of Peganum harmala (L) seeds and their combination effects. Fitoterapia 2010, 81, 779–782. [Google Scholar] [CrossRef]
- Dai, J.; Dan, W.; Ren, S.; Shang, C.; Wang, J. Design, synthesis and biological evaluations of quaternization harman analogues as potential antibacterial agents. Eur. J. Med. Chem. 2018, 160, 23–36. [Google Scholar] [CrossRef]
- de Carvalho Junior, A.R.; Oliveira Ferreira, R.; de Souza Passos, M.; da Silva Boeno, S.I.; Glória das Virgens, L.D.L.; Ventura, T.L.B.; Calixto, S.D.; Lassounskaia, E.; de Carvalho, M.G.; Braz-Filho, R.; et al. Antimycobacterial and Nitric Oxide Production Inhibitory Activities of Triterpenes and Alkaloids from Psychotria nuda (Cham. & Schltdl.) Wawra. Molecules 2019, 24, 1026. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Singh, M.; Chadha, N.; Silakari, O. Oxindole: A chemical prism carrying plethora of therapeutic benefits. Eur. J. Med. Chem. 2016, 123, 858–894. [Google Scholar] [CrossRef]
- Ahmad, R.; Salim, F. Oxindole Alkaloids of Uncaria (Rubiaceae, Subfamily Cinchonoideae): A Review on Its Structure, Properties, and Bioactivities. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2015; Volume 45, pp. 485–525. [Google Scholar]
- Lopes, A.A.; Chioca, B.; Musquiari, B.; Crevelin, E.J.; França, S.D.C.; Silva, M.F.D.G.F.D.; Pereira, A.M.S. Unnatural spirocyclic oxindole alkaloids biosynthesis in Uncaria guianensis. Sci. Rep. 2019, 9, 11349. [Google Scholar] [CrossRef] [Green Version]
- Khetmalis, Y.M.; Shivani, M.; Murugesan, S.; Chandra Sekhar, K.V.G. Oxindole and its derivatives: A review on recent progress in biological activities. Biomed. Pharmacother. 2021, 141, 111842. [Google Scholar] [CrossRef] [PubMed]
- Cerchiaro, G.; Ferreira, A.M.D.C. Oxindoles and copper complexes with oxindole-derivatives as potential pharmacological agents. J. Braz. Chem. Soc. 2006, 17, 1473–1485. [Google Scholar] [CrossRef] [Green Version]
- Yurttaş, L.; Ertaş, M.; Cankılıç Yılmaz, M.; Demirayak, Ş. Synthesis and Antimycobacterial Activity Evaluation of Isatin-derived 3-[(4-aryl-2-thiazolyl])hydrazone]-1H-indol-2,3-diones. Acta Pharm. Sci. 2017, 55, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Bertol, G.; Franco, L.; Oliveira, B.H. HPLC Analysis of Oxindole Alkaloids in Uncaria tomentosa: Sample Preparation and Analysis Optimisation by Factorial Design. Phytochem. Anal. 2012, 23, 143–151. [Google Scholar] [CrossRef]
- Luna-Palencia, G.; Cerda-Garcıa-Rojas, C.; Romo-Cruz, I.; Albarrán, J.; Ramos-Valdivia, A. Advances in the biosynthesis of oxindole alkaloids in Uncaria tomentosa cultures. In Proceedings of the Inter Symp Chem Pharmacol and Biosynthesis of Alkaloids, Belek, Antalya, Turkey, April 2006. [Google Scholar]
- Luna-Palencia, G.R.; Cerda-García-Rojas, C.M.; Rodríguez-Monroy, M.; Ramos-Valdivia, A.C. Influence of Auxins and Sucrose in Monoterpenoid Oxindole Alkaloid Production by Uncaria tomentosa Cell Suspension Cultures. Biotechnol. Prog. 2005, 21, 198–204. [Google Scholar] [CrossRef]
- Yuan, D.; Ma, B.; Wu, C.; Yang, J.; Zhang, L.; Liu, S.; Wu, L.; Kano, Y. Alkaloids from the Leaves of Uncaria rhynchophylla and Their Inhibitory Activity on NO Production in Lipopolysaccharide-Activated Microglia. J. Nat. Prod. 2008, 71, 1271–1274. [Google Scholar] [CrossRef]
- Hemingway, S.R.; Phillipson, J.D. Alkaloids from S. American species of Uncaria (Rubiaceae). J. Pharm. Pharmacol. 2011, 26, 113P. [Google Scholar] [CrossRef]
- Seki, H.; Takayama, H.; Aimi, N.; Sakai, S.-I.; Ponglux, D. A Nuclear Magnetic Resonance Study on the Eleven Stereoisomers of Heteroyohimbine-Type Oxindole Alkaloids. Chem. Pharm. Bull. 1993, 41, 2077–2086. [Google Scholar] [CrossRef] [Green Version]
- Lounasmaa, M.; Kan, S.-K. A 400 mhz 1H NMR study of the eight basic heteroyohimbine alkaloids. Tetrahedron 1980, 36, 1607–1611. [Google Scholar] [CrossRef]
- Paniagua-Pérez, R.; Madrigal-Bujaidar, E.; Molina-Jasso, D.; Reyes-Cadena, S.; Álvarez-González, I.; Sánchez-Chapul, L.; Pérez-Gallaga, J. Antigenotoxic, Antioxidant and Lymphocyte Induction Effects Produced by Pteropodine. Basic Clin. Pharmacol. Toxicol. 2009, 104, 222–227. [Google Scholar] [CrossRef]
- Dreifuss, A.A.; Bastos-Pereira, A.L.; Fabossi, I.A.; Lívero, F.A.D.R.; Stolf, A.M.; de Souza, C.E.A.; Gomes, L.D.O.; Constantin, R.P.; Furman, A.E.F.; Strapasson, R.L.B.; et al. Uncaria tomentosa Exerts Extensive Anti-Neoplastic Effects against the Walker-256 Tumour by Modulating Oxidative Stress and Not by Alkaloid Activity. PLoS ONE 2013, 8, e54618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, B.C.; Morel, L.J.F.; Carmona, F.; Cunha, T.M.; Contini, S.H.T.; Delprete, P.G.; Ramalho, F.S.; Crevelin, E.; Bertoni, B.W.; França, S.C. Aqueous extracts from Uncaria tomentosa (Willd. ex Schult.) DC. reduce bronchial hyperresponsiveness and inflammation in a murine model of asthma. J. Ethnopharmacol. 2018, 218, 76–89. [Google Scholar] [CrossRef]
- Rojas-Duran, R.; González-Aspajo, G.; Ruiz-Martel, C.; Bourdy, G.; Doroteo-Ortega, V.H.; Alban-Castillo, J.; Robert, G.; Auberger, P.; Deharo, E. Anti-inflammatory activity of Mitraphylline isolated from Uncaria tomentosa bark. J. Ethnopharmacol. 2012, 143, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Kośmider, A.; Czepielewska, E.; Kuraś, M.; Gulewicz, K.; Pietrzak, W.; Nowak, R.; Nowicka, G. Uncaria tomentosa Leaves Decoction Modulates Differently ROS Production in Cancer and Normal Cells, and Effects Cisplatin Cytotoxicity. Molecules 2017, 22, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erasto, P.; Marciale, C.; Omolo, J.J.; Hamilton, C.J. Potentiation of Isoniazid Efficacy against Isoniazid-resistant Mycobacteria Strains. Res. J. Med. Plant 2014, 8, 32–40. [Google Scholar] [CrossRef]
- García, R.; Cayunao, C.; Bocic, R.; Backhouse, N.; Delporte, C.; Zaldivar, M.; Erazo, S. Antimicrobial activity of isopteropodine. Z. Nat. C 2005, 60, 385–388. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; Zarrelli, A.; Pinto, G.; Pollio, A. Plant polyphenols and their anti-cariogenic properties: A review. Molecules 2011, 16, 1486–1507. [Google Scholar] [CrossRef] [Green Version]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Jovanović, A.; Petrović, P.; Đorđević, V.; Zdunić, G.; Šavikin, K.; Bugarski, B. Polyphenols extraction from plant sources. Lek. Sirovine 2017, 37, 45–49. [Google Scholar] [CrossRef]
- Panche, A.; Diwan, A.; Chandra, S. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Júnior, O.T.; Kuhn, F.; Padilha, P.J.M.; Vicente, L.R.M.; da Costa, S.W.; da Silva, B.C.; Schleder, D.D.; Nunes Nesi, C.; Magro, J.D.; de Lamo-Castellví, S. Survival of White Spot Syndrome Virus–Infected Litopenaeus vannamei Fed with Ethanol Extract of Uncaria tomentosa. J. World Aquac. Soc. 2018, 49, 165–174. [Google Scholar] [CrossRef]
- White, G.; Bourbonnais-Spear, N.; Garner, F. Antibacterial constituents from Uncaria tomentosa. Phytopharmacology 2011, 1, 16–19. [Google Scholar]
- Peñaloza, E.M.C.; Kaiser, S.; Resende, P.E.D.; Pittol, V.; Carvalho, Â.R.; Ortega, G.G. Chemical composition variability in the Uncaria tomentosa (cat s claw) wild population. Química Nova 2015, 38, 378–386. [Google Scholar]
- Pavei, C.; Kaiser, S.; Borré, G.L.; Ortega, G.G. Validation of a LC method for polyphenols assay in cat’s claw (Uncaria tomentosa). J. Liq. Chromatogr. Relat. Technol. 2010, 33, 1551–1561. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.-J.; Gao, K. Chemical Constituents and Biological Activities of Dicranopteris linearis. Chem. Nat. Compd. 2014, 49, 1129–1131. [Google Scholar] [CrossRef]
- Albuquerque, U.P.; Patil, U.; Máthé, Á. Medicinal and Aromatic Plants of South America: Brazil; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Yang, H.; Tuo, X.; Wang, L.; Tundis, R.; Portillo, M.P.; Simal-Gandara, J.; Yu, Y.; Zou, L.; Xiao, J.; Deng, J. Bioactive procyanidins from dietary sources: The relationship between bioactivity and polymerization degree. Trends Food Sci. Technol. 2021, 111, 114–127. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Monagas Juan, M.J.; Sánchez-Patán, F.; Quintanilla-López, J.E.; Lebrón-Aguilar, R.; Bartolomé, B.; Navarro Hoyos, M. Phenolic Extracts of Uncaria tomentosa L. (cats claw) Containing Procyanidins, Propelargonidins and Flavonolignans, Method for the Production Thereof, and Applicarions of Same. WO2014096488A1, 26 June 2014. [Google Scholar]
- Navarro-Hoyos, M.; Alvarado-Corella, D.; Moreira-Gonzalez, I.; Arnaez-Serrano, E.; Monagas-Juan, M. Polyphenolic Composition and Antioxidant Activity of Aqueous and Ethanolic Extracts from Uncaria tomentosa Bark and Leaves. Antioxidants 2018, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; He, C.; Song, L.; Li, T.; Cui, S.; Zhang, L.; Jia, Y. Antimicrobial activity and mechanism of Larch bark procyanidins against Staphylococcus aureus. Acta Biochim. Biophys. Sin. 2017, 49, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, T.; Dib, R.; Al-Youssef, H.; Amina, M. Chemical composition and antimicrobial and cytotoxic activities of Antidesm abunius L. Pak. J. Pharm. Sci. 2019, 32, 153–163. [Google Scholar] [PubMed]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta (BBA) Biomembr. 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Karioti, A.; Sokovic, M.; Ciric, A.; Koukoulitsa, C.; Bilia, A.R.; Skaltsa, H. Antimicrobial Properties of Quercus ilex L. Proanthocyanidin Dimers and Simple Phenolics: Evaluation of Their Synergistic Activity with Conventional Antimicrobials and Prediction of Their Pharmacokinetic Profile. J. Agric. Food Chem. 2011, 59, 6412–6422. [Google Scholar] [CrossRef] [PubMed]
- Wirth, C.; Wagner, H. Pharmacologically active procyanidines from the bark of Uncaria tomentosa. Phytomedicine 1997, 4, 265–266. [Google Scholar] [CrossRef]
- Maas, J.L.; Galletta, G.J.; Stoner, G.D. Ellagic acid, an anticarcinogen in fruits, especially in strawberries: A review. HortScience 1991, 26, 10–14. [Google Scholar] [CrossRef]
- Vattem, D.; Shetty, K. Biological functionality of ellagic acid: A review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Joshi, J.R.; Burdman, S.; Lipsky, A.; Yariv, S.; Yedidia, I. Plant phenolic acids affect the virulence of Pectobacterium aroidearum and P. carotovorum ssp. brasiliense via quorum sensing regulation. Mol. Plant Pathol. 2016, 17, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure–function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Rivas, C.L.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Godlewska-Żyłkiewicz, B.; Świsłocka, R.; Kalinowska, M.; Golonko, A.; Świderski, G.; Arciszewska, Ż.; Nalewajko-Sieliwoniuk, E.; Naumowicz, M.; Lewandowski, W. Biologically Active Compounds of Plants: Structure-Related Antioxidant, Microbiological and Cytotoxic Activity of Selected Carboxylic Acids. Materials 2020, 13, 4454. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Huang, S.-T.; Sun, F.-M.; Chiang, Y.-L.; Chiang, C.-J.; Tsai, C.-M.; Weng, C.-J. Transformation of cinnamic acid from trans-to cis-form raises a notable bactericidal and synergistic activity against multiple-drug resistant Mycobacterium tuberculosis. Eur. J. Pharm. Sci. 2011, 43, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Wu, X.; Shi, J.; Yang, Q.; Zhang, Y. Phenolic compounds from the edible seeds extract of Chinese Mei (Prunus mume Sieb. et Zucc) and their antimicrobial activity. Food Sci. Technol. 2011, 44, 347–349. [Google Scholar] [CrossRef]
- Kępa, M.; Miklasińska-Majdanik, M.; Wojtyczka, R.D.; Idzik, D.; Korzeniowski, K.; Smoleń-Dzirba, J.; Wąsik, T.J. Antimicrobial potential of caffeic acid against Staphylococcus aureus clinical strains. BioMed Res. Int. 2018, 2018, 7413504. [Google Scholar] [CrossRef] [Green Version]
- Hemaiswarya, S.; Doble, M. Synergistic interaction of phenylpropanoids with antibiotics against bacteria. J. Med. Microbiol. 2010, 59, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. In Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2019; pp. 333–359. [Google Scholar] [CrossRef]
- Patočka, J. Biologically active pentacyclic triterpenes and their current medicine signification. J. Appl. Biomed. 2003, 1, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef]
- Copp, B.R. Antimycobacterial natural products. Nat. Prod. Rep. 2003, 20, 535–557. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, E.; Lin, F.Y. Terpene biosynthesis: Modularity rules. Angew. Chem. Int. Ed. 2012, 51, 1124–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, P.D.; Almeida, A.; Bak, S. Evolution of Structural Diversity of Triterpenoids. Front. Plant Sci. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Benveniste, P. Biosynthesis and accumulation of sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. [Google Scholar] [CrossRef]
- Miras-Moreno, B.; Sabater-Jara, A.B.; Pedreño, M.A.; Almagro, L. Bioactivity of Phytosterols and Their Production in Plant in Vitro Cultures. J. Agric. Food Chem. 2016, 64, 7049–7058. [Google Scholar] [CrossRef]
- Senatore, A.; Cataldo, A.; Iaccarino, F.P.; Elberti, M.G. Phytochemical and biological study of Uncaria tomentosa. Boll. della Soc. Ital. Biol. Sper. 1989, 65, 517–520. [Google Scholar]
- Muhammad, I.; Dunbar, D.C.; Khan, R.A.; Ganzera, M.; Khan, I.A. Investigation of Uña De Gato I. 7-Deoxyloganic acid and 15N NMR spectroscopic studies on pentacyclic oxindole alkaloids from Uncaria tomentosa. Phytochemistry 2001, 57, 781–785. [Google Scholar] [CrossRef]
- Sen, A.; Dhavan, P.; Shukla, K.K.; Singh, S.; Tejovathi, G. Analysis of IR, NMR and antimicrobial activity of β-sitosterol isolated from Momordica charantia. Sci. Secur. J. Biotechnol. 2012, 1, 9–13. [Google Scholar]
- Hoskeri, J.; Krishna, H.; Jignesh, V.; Roshan, S.; Vijay, S. In-silico drug designing using β-sitosterol isolated from Flaveria trinervia against peptide deformylase protein to hypothesize bactericidal effect. Int. J. Pharm. Pharm. Sci. 2012, 4, 192–196. [Google Scholar]
- Mailafiya, M.M.; Yusuf, A.J.; Abdullahi, M.I.; Aleku, G.A.; Ibrahim, I.A.; Yahaya, M.; Abubakar, H.; Sanusi, A.; Adamu, H.W.; Alebiosu, C.O. Antimicrobial activity of stigmasterol from the stem bark of Neocarya macrophylla. J. Med. Plants Econ. Dev. 2018, 2, 1–5. [Google Scholar]
- Kuete, V.; Eyong, K.; Folefoc, G.; Beng, V.; Hussain, H.; Krohn, K.; Nkengfack, A. Antimicrobial activity of the methanolic extract and of the chemical constituents isolated from Newbouldia laevis. Pharmazie 2007, 62, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Sycz, Z.; Tichaczek-Goska, D.; Wojnicz, D. Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals. Biomolecules 2022, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Catteau, L.; Zhu, L.; Françoise Van, B.; Quetin-Leclercq, J. Natural and hemi-synthetic pentacyclic triterpenes as antimicrobials and resistance modifying agents against Staphylococcus aureus: A review. Phytochem. Rev. 2018, 17, 1129–1163. [Google Scholar] [CrossRef]
- Ren, D.; Zuo, R.; Barrios, A.F.G.; Bedzyk, L.A.; Eldridge, G.R.; Pasmore, M.E.; Wood, T.K. Differential Gene Expression for Investigation of Escherichia coli Biofilm Inhibition by Plant Extract Ursolic Acid. Appl. Environ. Microbiol. 2005, 71, 4022–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Tan, X.; Jiao, Y.; Liu, L.; Zhao, W.; Yang, S.; Jia, A. RNA-Seq-based transcriptome analysis of methicillin-resistant Staphylococcus aureus biofilm inhibition by ursolic acid and resveratrol. Sci. Rep. 2014, 4, 5467. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Gao, Y.; Wang, A.; Zhou, X.; Zheng, Y.; Zhou, J. Evolution in medicinal chemistry of ursolic acid derivatives as anticancer agents. Eur. J. Med. Chem. 2015, 92, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.-W.; Dai, Y.-C.; Xue, J.-P.; Wang, J.-C.; Lin, F.-P.; Guo, Y.-H. In vitro and in vivo anticancer activity evaluation of ursolic acid derivatives. Eur. J. Med. Chem. 2011, 46, 2652–2661. [Google Scholar] [CrossRef]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic acid and its derivatives as bioactive agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [Green Version]
- Fontanay, S.; Grare, M.; Mayer, J.; Finance, C.; Duval, R.E. Ursolic, oleanolic and betulinic acids: Antibacterial spectra and selectivity indexes. J. Ethnopharmacol. 2008, 120, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Wolska, K.I.; Grudniak, A.M.; Fiecek, B.; Kraczkiewicz-Dowjat, A.; Kurek, A. Antibacterial activity of oleanolic and ursolic acids and their derivatives. Cent. Eur. J. Biol. 2010, 5, 543–553. [Google Scholar] [CrossRef]
- Kurek, A.; Nadkowska, P.; Pliszka, S.; Wolska, K.I. Modulation of antibiotic resistance in bacterial pathogens by oleanolic acid and ursolic acid. Phytomedicine 2012, 19, 515–519. [Google Scholar] [CrossRef]
- Aquino, R.; De Tommasi, N.; De Simone, F.; Pizza, C. Triterpenes and quinovic acid glycosides from Uncaria tomentosa. Phytochemistry 1997, 45, 1035–1040. [Google Scholar] [CrossRef]
- Yepes-Pérez, A.F.; Herrera-Calderon, O.; Quintero-Saumeth, J. Uncaria tomentosa (cat’s claw): A promising herbal medicine against SARS-CoV-2/ACE-2 junction and SARS-CoV-2 spike protein based on molecular modeling. J. Biomol. Struct. Dyn. 2020, 1–17. [Google Scholar] [CrossRef]
- Aquino, R.; De Simone, F.; Vincieri, F.F.; Pizza, C.; Gaćs-Baitz, E. New polyhydroxylated triterpenes from Uncaria tomentosa. J. Nat. Prod. 1990, 53, 559–564. [Google Scholar] [CrossRef]
- Aquino, R.; De Feo, V.; De Simone, F.; Pizza, C.; Cirino, G. Plant metabolites. New compounds and anti-inflammatory activity of Uncaria tomentosa. J. Nat. Prod. 1991, 54, 453–459. [Google Scholar] [CrossRef]
- Sakakibara, J.; Kaiya, T.; Fukuda, H. Triterpenoids from enkianthus campanulatus. Phytochemistry 1984, 23, 627–630. [Google Scholar] [CrossRef]
- Amir, F.; Wong, K.C.; Eldeen, I.; Asmawi, M.Z.; Osman, H. Evaluation of biological activities of extracts and chemical constituents of Mimusops elengi. Trop. J. Pharm. Res. 2013, 12, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Cerri, R.; Aquino, R.; de Simone, F.; Pizza, C. New Quinovic Acid Glycosides from Uncaria tomentosa. J. Nat. Prod. 1988, 51, 257–261. [Google Scholar] [CrossRef]
- Aquino, R.; De Simone, F.; Pizza, C.; Conti, C.; Stein, M. Plant Metabolites. Structure and In Vitro Antiviral Activity of Quinovic Acid Glycosides from Uncaria tomentosa and Guettarda platyipoda. J. Nat. Prod. 1989, 52, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Pavei, C.; Kaiser, S.; Verza, S.G.; Borre, G.L.; Ortega, G.G. HPLC-PDA method for quinovic acid glycosides assay in Cat’s claw (Uncaria tomentosa) associated with UPLC/Q-TOF–MS analysis. J. Pharm. Biomed. Anal. 2012, 62, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S.A.; et al. Phytochemical Profiling, In Vitro and In Silico Anti-Microbial and Anti-Cancer Activity Evaluations and Staph GyraseB and h-TOP-IIβ Receptor-Docking Studies of Major Constituents of Zygophyllum coccineum L. Aqueous-Ethanolic Extract and Its Subsequent Fractions: An Approach to Validate Traditional Phytomedicinal Knowledge. Molecules 2021, 26, 577. [Google Scholar] [CrossRef]
- Kezetas Bankeu, J.J.; Kenou Kagho, D.U.; Fotsing Fongang, Y.S.p.; Kouipou Toghueo, R.M.; Mba’ning, B.M.r.; Tchouya Feuya, G.R.; Boyom Fekam, F.; Tchouankeu, J.C.; Ngouela, S.r.A.; Sewald, N.; et al. Constituents from Nauclea latifolia with Anti-Haemophilus influenzae Type b Inhibitory Activities. J. Nat. Prod. 2019, 82, 2580–2585. [Google Scholar] [CrossRef]
- Asada, K.; Salim, V.; Masada-Atsumi, S.; Edmunds, E.; Nagatoshi, M.; Terasaka, K.; Mizukami, H.; De Luca, V. A 7-Deoxyloganetic Acid Glucosyltransferase Contributes a Key Step in Secologanin Biosynthesis in Madagascar Periwinkle. Plant Cell 2013, 25, 4123–4134. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||
|---|---|---|---|---|---|
| Compound | R1 | R2 | R3 | R4 | R5 |
| Benzoic, 47 | H | H | H | H | H |
| Salicylic acid, 48 | OH | H | H | H | H |
| 4-Hydroxbenzoic, 49 | H | H | OH | H | H |
| Protocatechuic acid, 50 | H | OH | OH | H | H |
| Gallic acid, 51 | H | OH | OH | OH | H |
| Syringic, 52 | H | OMe | OH | OMe | H |
| Vanillic acid, 53 | H | OMe | OH | H | H |
 |  | ||
| Compound | R1 | R2 | |
| p-Coumaric acid, 54 | H | OH | |
| Caffeic acid, 55 | OH | OH | |
| Ferulic acid, 56 | OMe | OH | |
| Isoferulic, 57 | OH | OMe | |
 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | R10 |
| (3β)-3-Hydroxy-7-oxo-urs-12-ene-27,28-dioic acid, 67 | OH | Me | Me | H | =O | CO2H | H | Me | H | H |
| (3β,16α)-16-Hydroxy-3-methoxy-ursa-12,19(29)-diene-27,28-dioic acid, 68 | MeO | Me | Me | H | H | CO2H | OH | = | H | |
| (3β,4α)-3,19-Dihydroxy-6,23-dioxo-urs-12-en-28-oic acid, 69 | OH | Me | CHO | =O | H | Me | H | Me | OH | H |
| (3β,4α)-3,19,23-trihydroxy-6-oxo-urs-12-en-28-oic acid, 70 | OH | Me | CH2OH | =O | H | Me | H | Me | OH | H |
| (6β)-6,19-dihydroxy-3-oxo-urs-12-en-28-oic acid, 71 | =O | Me | Me | OH | H | Me | H | Me | OH | H |
| (3β,6β)-3,6,19-Trihydroxyurs-12-en-28-oic acid (uncaric acid), 72 | OH | Me | Me | OH | H | Me | H | Me | OH | H |
| (3β,4α,6β)-3,6,19-trihydroxy-23-oxo-norurs-12-en-28-oic acid, 73 | OH | =O | OH | H | Me | H | Me | OH | H | |
| (3β,4α,6β)-3,6,19-trihydroxy-23-oxo-urs-12-en-28-oic acid, 74 | OH | Me | CHO | OH | H | Me | H | Me | OH | H |
| (3β,6β)-3,6,19-Trihydroxy-24-norursa-4(23),12-dien-28-oic acid (floridic acid), 75 | OH | = | OH | H | Me | H | Me | OH | H | |
| (3β,4α,6β)-dimethyl ester-3,6,19-trihydroxy-urs-12-ene-23,28-dioic acid, 76 | OH | Me | CO2Me | OH | H | Me | H | Me | OH | Me |
| (3β,4α,6β)-3,6,19,23-tetrahydroxy-urs-12-en-28-oic acid, 77 | OH | Me | CH2OH | OH | H | Me | H | Me | OH | H |
 | |
|---|---|
| No. | Compound Name |
| 78 | Cincholic acid 27-O-β-D-fucopyranosyl-28-O-β-D-glucopyranoside |
| 79 | Cincholic acid 27-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranoside |
| 80 | Cincholic acid 27-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranosyl-28-O-β-D-glucopyranoside |
| 81 | Pyrocincholic acid 27-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranosyl-28-O-β-D-glucopyranoside (Tomentoside B) |
 | |
|---|---|
| No. | Compound |
| 82 | Quinovic acid 3-O-β-D-quinovopyranosyl-27-O-β-D-glucopyranoside |
| 83 | Quinovic acid 3-O-β-D-quinovopyranosyl-28-O-β-D-glucopyranoside |
| 84 | Quinovic acid 3-O-α-L-rhamnoside |
| 85 | Quinovic acid 3-O-β-D-fucopyranosyl-27-O-β-D-glucopyranoside |
| 86 | Quinovic acid 3-O-β-D-fucopyranosyl-28-O-β-D-glucopyranoside |
| 87 | Quinovic acid 27-O-β-D-glucopyranoside |
| 88 | Quinovic acid 3-O-β-D-glucopyranosyl-(1→3)-β-D-quinovopyranoside |
| 89 | Quinovic acid 3-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranoside |
| 90 | Quinovic acid 3-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranosyl-27-O-β-D-glucopyranoside |
| 91 | Quinovic acid 3-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranosyl-28-O-β-D-glucopyranoside |
| 92 | Quinovic acid 3-O-α-L-rhamnopyranosyl-(3→1)-β-D-glucopyranoside |
| 93 | Quinovic acid 3-O-α-L-rhamnopyranosyl-(3→1)-β-D-glucopyranosyl-27-O-β-D-glucopyranoside |
| 94 | Quinovic acid 3-O-β-D-galactopyranosyl-(1→3)-β-D-quinovopyranoside |
| 95 | Pyroquinovic acid 3-O-β-D-glucopyranosyl-(1→3)-β-D-fucopyranosyl-28-O-β-D-glucopyranoside (Tomentoside A) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanck, J.J.; Huebner, T.M.; Rolls, A.M.; Cornell, J.S.; Hwang, C.S. Comprehensive Review of the Components in Cat’s Claw (Uncaria tomentosa) and Their Antibacterial Activity. AppliedChem 2022, 2, 1-29. https://doi.org/10.3390/appliedchem2010001
Blanck JJ, Huebner TM, Rolls AM, Cornell JS, Hwang CS. Comprehensive Review of the Components in Cat’s Claw (Uncaria tomentosa) and Their Antibacterial Activity. AppliedChem. 2022; 2(1):1-29. https://doi.org/10.3390/appliedchem2010001
Chicago/Turabian StyleBlanck, Jason J., Thomas M. Huebner, Alyssa M. Rolls, Josh S. Cornell, and Candy S. Hwang. 2022. "Comprehensive Review of the Components in Cat’s Claw (Uncaria tomentosa) and Their Antibacterial Activity" AppliedChem 2, no. 1: 1-29. https://doi.org/10.3390/appliedchem2010001