

In Vitro Antioxidant and Anti-Inflammatory Activities of Bioactive Proteins and Peptides from Rhodomonas sp.
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain, Growth Conditions, and Protein Accumulation Monitoring
2.2. Biochemical Characterization of Rhodomonas sp. Biomass
2.3. Preparation of Rhodomonas sp. Proteins Extract
2.4. Cell density and Proteins Production Assessments
2.5. Preparation of Rhodomonas sp. Peptides Hydrolysates
2.6. FTIR Spectroscopy
2.7. Antioxidant Activity Evaluation
2.7.1. DPPH Radical-Scavenging Activity
2.7.2. Ferric-Reducing Power Assay
2.8. In vitro Anti-Inflammatory Activity Evaluation
3. Results and Discussion
3.1. Biochemical Characterization of Rhodomonas sp. Biomass
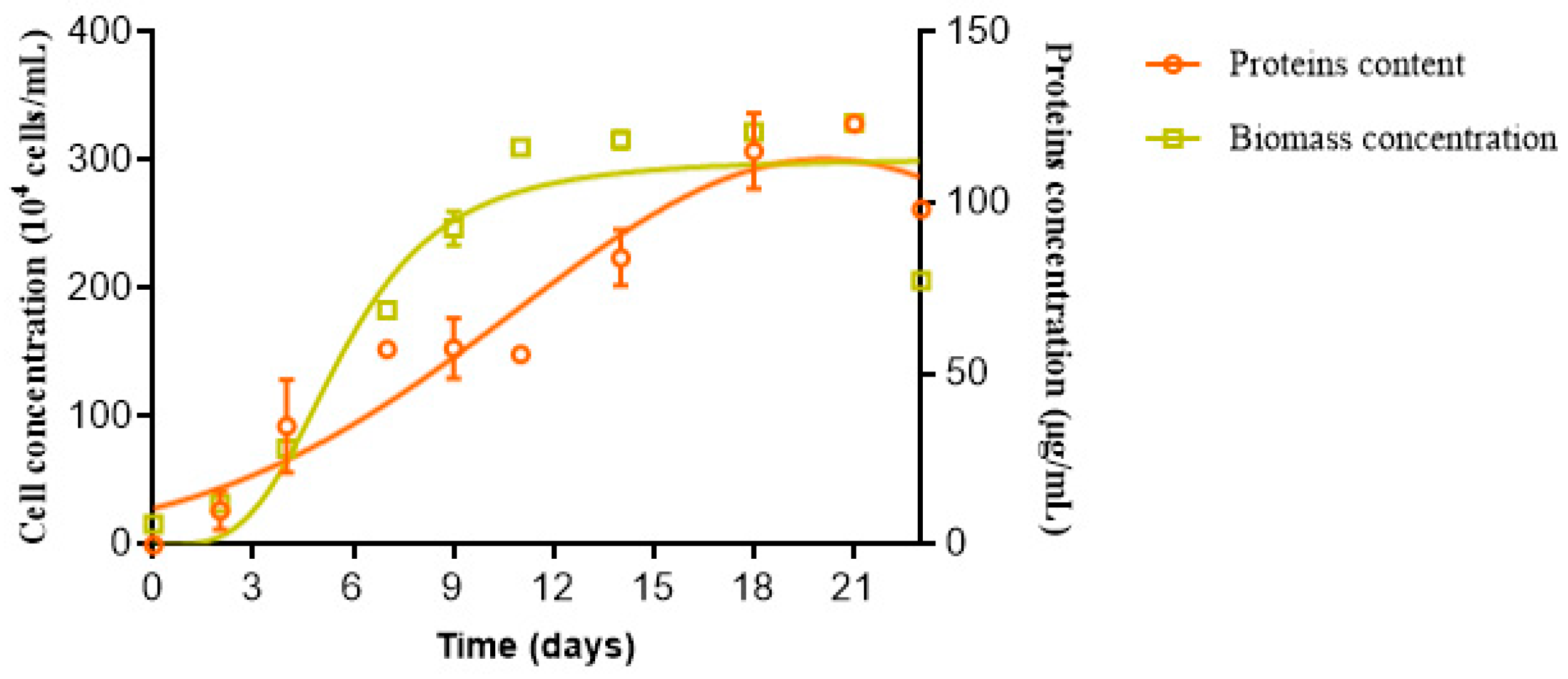
3.2. Cell Density and Protein Production Assessments
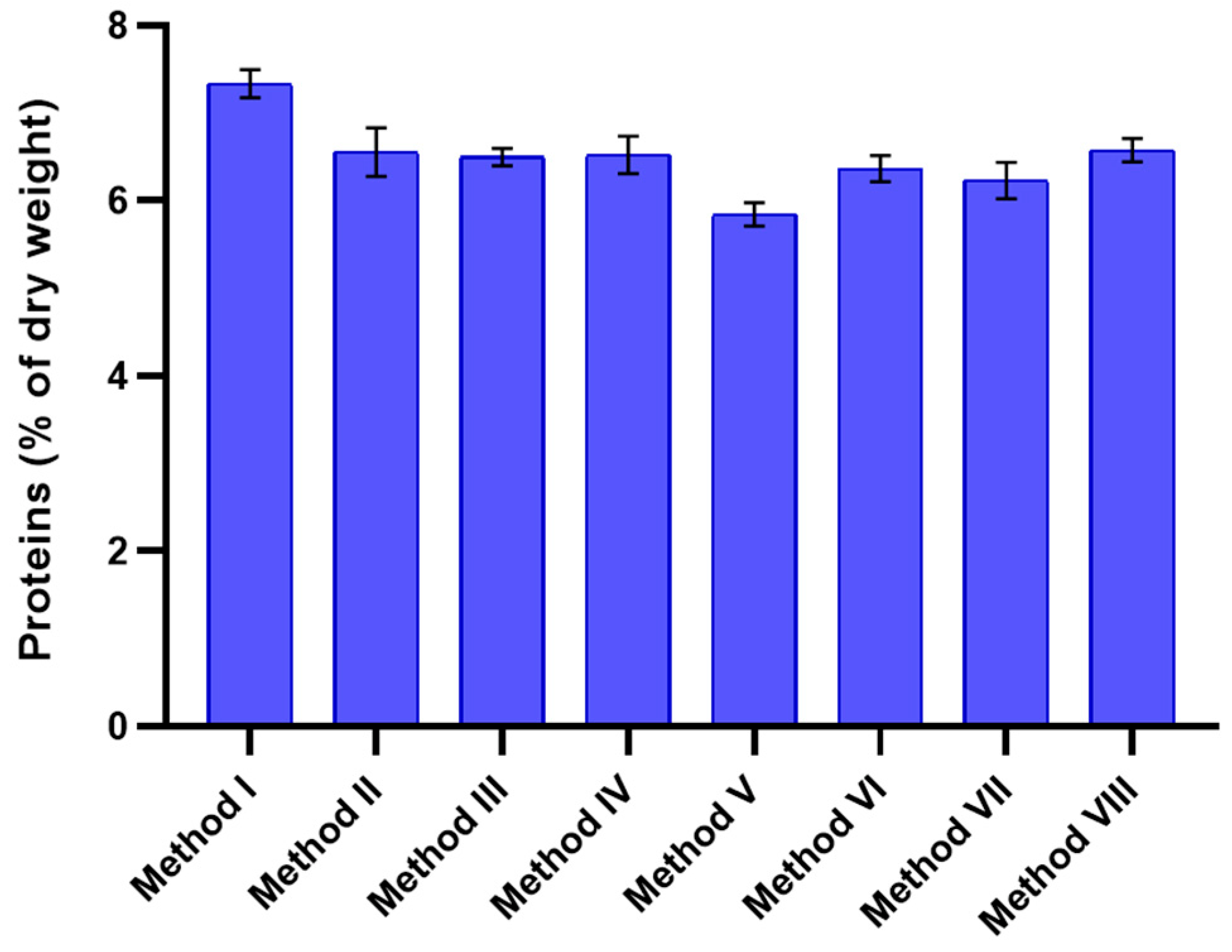
3.3. Total Intracellular Protein Extraction Using Different Lysis Methods
3.4. Peptides Hydrolysed by Alcalase and Pepsin
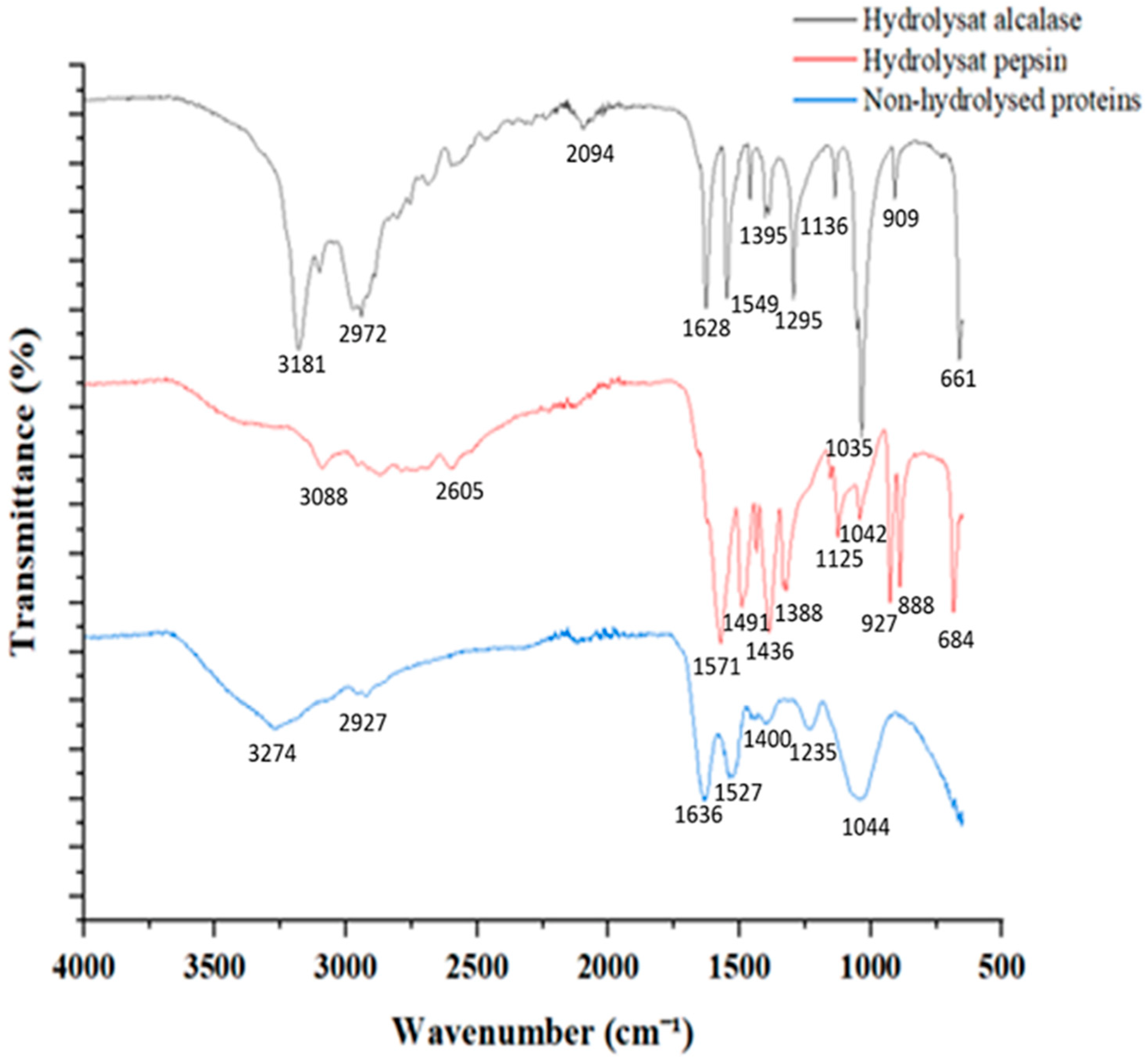
3.5. Proteins and Peptides Characterization
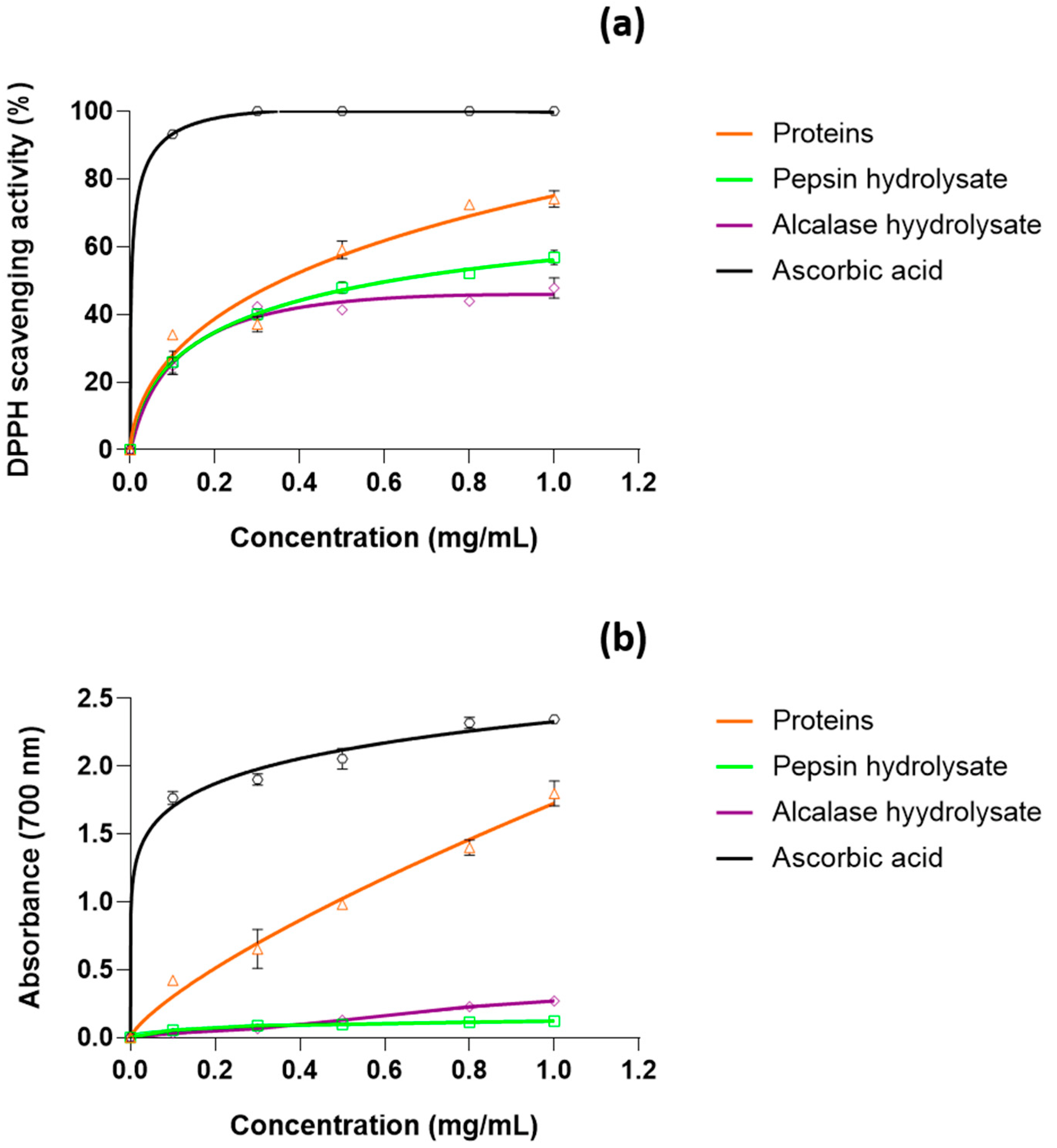
3.6. Antioxidant Activity
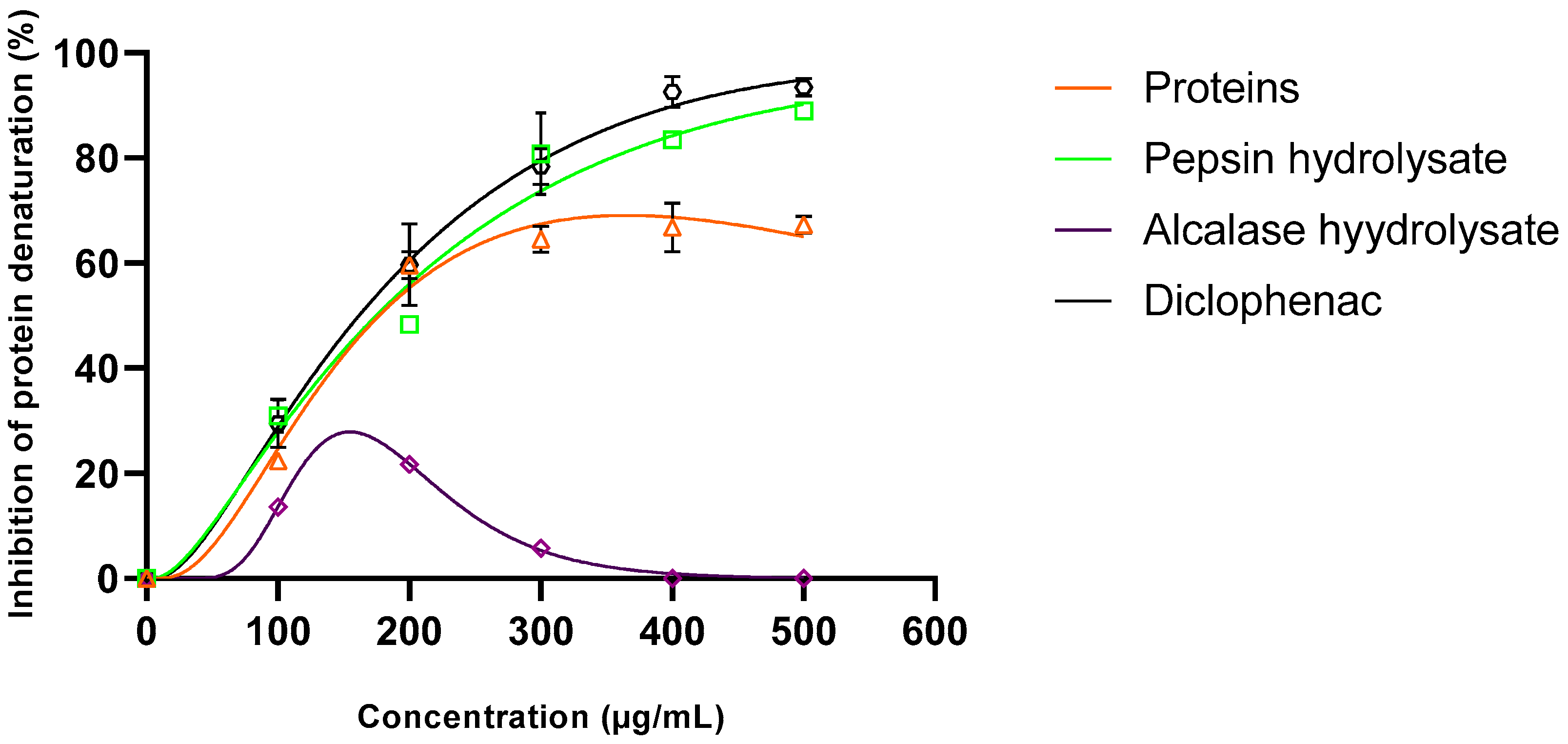
3.7. Anti-Inflammatory Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kar, B.; Kumar, R.S.; Karmakar, I.; Dola, N.; Bala, A.; Mazumder, U.K.; Hadar, P.K. Antioxidant and in vitro anti-inflammatory activities of Mimusops elengi leaves. Asian Pac. J. Trop. Biomed. 2012, 2, 976–980. [Google Scholar] [CrossRef]
- Tierney, M.S.; Croft, A.K.; Hayes, M. A review of antihypertensive and antioxidant activities in macroalgae. Bot. Mar. 2010, 53, 387–408. [Google Scholar] [CrossRef]
- Gargouch, N.; Karkouch, I.; Elleuch, J.; Elkahoua, S.; Michaud, P.; Abdelkafi, S.; Laroche, C.; Fendri, I. Enhanced B-phycoerythrin production by the red microalga Porphyridium marinum: A powerful agent in industrial applications. Int. J. Biol. Macromol. 2018, 120, 2106–2114. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Nelson, D.R.; Mystikou, A.; Daakour, S.; Salehi-Ashtiani, K. Advances in microalgal research and engineering development. Curr. Opin. Biotechnol. 2019, 59, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Miladi, R.; Manghisi, A.; Armeli Mincante, S.; Genovese, G.; Abdelkafi, S.; Morabito, M. A DNA barcoding survey of Ulva (Chlorophyta) in Tunisia and Italy reveals the presence of the overlooked alien Ulva ohnoi. Cryptogam. Algol. 2018, 39, 85–107. [Google Scholar] [CrossRef]
- Uma, V.S.; Usmani, Z.; Sharma, M.; Diwan, D.; Sharma, M.; Guo, M.; Tuohy, M.G.; Makatsoris, C.; Zhao, X.; Thakur, V.K.; et al. Valorisation of algal biomass to value-added metabolites: Emerging trends and opportunities. Phytochem. Rev. 2022. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Hadjkacem, F.; Hentati, F.; Drira, R.; Pierre, G.; Gardarin, C.; Delattre, C.; El Alaoui-Talibi, Z.; Cherkaoui, E.M.; et al. Influence of the sulfate content of the exopolysaccharides from Porphyridium sordidum on their elicitor activities on date palm vitroplants. Plant Physiol. Biochem 2022, 186, 99–106. [Google Scholar] [CrossRef]
- Elleuch, J.; Hadj Kacem, F.; Ben Amor, F.; Hadrich, B.; Michaud, P.; Fendri, I.; Abdelkafi, S. Extracellular neutral protease from Arthrospira platensis: Production, optimization and partial characterization. Int. J. Biol. Macromol. 2021, 167, 1491–1498. [Google Scholar] [CrossRef]
- Hadjkacem, F.; Pierre, G.; Christophe, G.; Elleuch, J.; Fendri, I.; Boual, Z.; Ould El Hadj, M.D.; El Alaoui-Talibi, Z.; El Modafar, C.; Dubessay, P.; et al. Bioconversion of the brown Tunisian seaweed Halopteris scoparia: Application to energy. Energies 2022, 15, 4342. [Google Scholar] [CrossRef]
- Elleuch, F.; Ben Hlima, H.; Barkallah, M.; Baril, P.; Abdelkafi, S.; Pichon, C.; Fendri, I. Carotenoids overproduction in Dunaliella sp.: Transcriptional changes and new insights through lycopene cyclase regulation. Appl. Sci. 2019, 9, 5389. [Google Scholar] [CrossRef] [Green Version]
- Ben Amor, F.; Barkallah, M.; Eleuch, F.; Karkouch, N.; Dammak, M.; Baréa, B.; Villeneuve, P.; Abdelkafi, S.; Fendri, I. Cyanobacteria as sources of marine bioactive compounds: Molecular specific detection based on Δ9 desaturase gene. Int. J. Biol. Macromol. 2017, 105, 1440–1445. [Google Scholar] [CrossRef]
- Tounsi, L.; Hentati, F.; Ben Hlima, H.; Barkallah, M.; Smaoui, S.; Fendri, I.; Philippe, M.; Abdelkafi, S. Microalgae as feedstock for bioactive polysaccharides. Int. J. Biol. Macromol. 2022, 221, 1238–1250. [Google Scholar] [CrossRef]
- Guzmán, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Tabarzad, M.; Atabaki, V.; Hosseinabadi, T. Anti-inflammatory activity of bioactive compounds from microalgae and cyanobacteria by focusing on the mechanisms of action. Mol. Biol. Rep. 2020, 47, 6193–6205. [Google Scholar] [CrossRef]
- Leelaprakash, G.; Dass, S.M. In vitro anti-inflammatory activity of methanol extract of Enicostemma axillare. Int. J. Drug Dev. Res. 2011, 3, 189–196. [Google Scholar]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive peptides: Synthesis, sources, applications, and proposed mechanisms of action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Guzmán, F.; Wong, G.; Román, T.; Cárdenas, C.; Alvárez, C.; Schmitt, P.; Albericio, F.; Rojas, V. Identification of antimicrobial peptides from the microalgae Tetraselmis suecica (Kylin) Butcher and bactericidal activity improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef] [Green Version]
- Vo, T.S.; Ryu, B.; Kim, S.K. Purification of novel anti-inflammatory peptides from enzymatic hydrolysate of the edible microalgal Spirulina maxima. J. Funct. Foods 2013, 5, 1336–1346. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food. Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [Green Version]
- Anh, T.L.Q.; Hoa, N.T.Q.; Nguyen, P.D.T.; Thanh, H.V.; Nguyen, P.B.; Anh, L.T.H.; Dao, D.T.A. Soybean Protein Extraction by Alcalase and Flavourzyme, Combining Thermal Pretreatment for Enteral Feeding Product. Catalysts 2020, 10, 829. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Arguelles, E.; Laurena, A.C.; Martinez-Goss, M.R.; Monsalud, R.G. Antibacterial activity, total phenolic content and antioxidant capacity of a green microalga Desmodesmus sp.(U-AU2) from Los Baños, Laguna (Philippines). J. Nat. Stud. 2017, 16, 1–13. [Google Scholar]
- Kang, K.H.; Qian, Z.J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.K. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in HepG2/CYP2E1 cells induced by ethanol. Phytother Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 13, 3419–3425. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Xia, E.; Zhai, L.; Huang, Z.; Liang, H.; Yang, H.; Song, G.; Li, W.; Tang, H. Optimization and identification of antioxidant peptide from underutilized Dunaliella salina protein: Extraction, in vitro gastrointestinal digestion, and fractionation. Biomed Res. Int. 2019, 2019, 6424651. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Norici, A.; Mollo, L.; Osimani, A.; Aquilanti, L. Fermentation of microalgal biomass for innovative food production. Microorganisms 2022, 10, 2069. [Google Scholar] [CrossRef]
- Oostlander, P.C.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Optimization of Rhodomonas sp. under continuous cultivation for industrial applications in aquaculture. Algal Res. 2020, 47, 101889. [Google Scholar] [CrossRef]
- Latsos, C.; van Houcke, J.; Blommaert, L.; Verbeeke, G.P.; Kromkamp, J.; Timmermans, K.R. Effect of light quality and quantity on productivity and phycoerythrin concentration in the cryptophyte Rhodomonas sp. J. Appl. Phycol. 2021, 33, 729–741. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Pancha, I.; Jain, D.; Mishra, S. Effect of preservatives for food grade C-Phycoerythrin, isolated from marine cyanobacteria Pseudanabaena sp. Int. J. Biol. Macromol. 2010, 47, 597–602. [Google Scholar] [CrossRef]
- Derbel, H.; Elleuch, J.; Tounsi, L.; Nicolo, M.S.; Rizzo, M.G.; Michaud, P.; Fendri, I.; Abdelkafi, S. Improvement of biomass and phycoerythrin production by a strain of Rhodomonas sp. isolated from the Tunisian coast of Sidi Mansour. Biomolecules 2022, 12, 885. [Google Scholar] [CrossRef]
- Elleuch, J.; Ben Amor, F.; Chaaben, Z.; Frikha, F.; Michaud, P.; Fendri, I.; Abdelkafi, S. Zinc biosorption by Dunaliella sp. AL-1: Mechanism and effects on cell metabolism. Sci. Total Environ. 2021, 773, 145024. [Google Scholar] [CrossRef]
- Ben Hlima, H.; Bohli, T.; Kraiem, M.; Ouederni, A.; Mellouli, L.; Michaud, P.; Abdelkafi, S.; Smaoui, S. Combined effect of Spirulina platensis and Punica granatum peel extacts: Phytochemical content and antiphytophatogenic activity. App. Sci. 2019, 9, 5475. [Google Scholar] [CrossRef] [Green Version]
- Ben Mohamed, J.; Elleuch, J.; Drira, M.; Esteban, M.Á.; Michaud, P.; Abdelkafi, S.; Fendri, I. Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast. Appl. Sci. 2021, 11, 5295. [Google Scholar] [CrossRef]
- Montone, C.M.; Capriotti, A.L.; Cavaliere, C.; La Barbera, G.; Piovesana, S.; Zenezini Chiozzi, R.; Laganà, A. Peptidomic strategy for purification and identification of potential ACE-inhibitory and antioxidant peptides in Tetradesmus obliquus microalgae. Anal. Bioanal. Chem. 2018, 410, 3573–3586. [Google Scholar] [CrossRef]
- Qi, W.; Su, R.X.; He, Z.M.; Zhang, Y.B.; Jin, F.M. Pepsin-induced changes in the size and molecular weight distribution of bovine casein during enzymatic hydrolysis. J. Dairy Sci. 2007, 90, 5004–5011. [Google Scholar] [CrossRef] [Green Version]
- Soto-Sierra, L.; Wilken, L.R.; Mallawarachchi, S.; Nikolov, Z.L. Process development of enzymatically-generated algal protein hydrolysates for specialty food applications. Algal Res. 2021, 55, 102248. [Google Scholar] [CrossRef]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrières, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the Tunisian brown seaweed Cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef]
- Yıldırım, A.; Mavi, A.; Kara, A.A. Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef]
- Chakou, F.Z.; Boual, Z.; Hadj, M.D.O.E.; Belkhalfa, H.; Bachari, K.; El Alaoui-Talibi, Z.; El Modafar, C.; Hadjkacem, F.; Fendri, I.; Abdelkafi, S.; et al. Pharmacological investigations in traditional utilization of Alhagi maurorum Medik. in saharan Algeria: In vitro study of anti-inflammatory and antihyperglycemic activities of water-soluble polysaccharides extracted from the seeds. Plants 2021, 10, 2658. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Whitney, C.G.; MacPherso, M.J.; Bhatti, S.; Banskota, A.H.; Stefanova, R.; McGinn, P.J. Biochemical characterization of microalgal biomass from freshwater species isolated in Alberta, Canada for animal feed applications. Algal Res. 2015, 11, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Latsos, C.; Van Houcke, J.; Timmermans, K.R. The effect of nitrogen starvation on biomass yield and biochemical constituents of Rhodomonas sp. Front. Mar. Sci. 2020, 7, 563333. [Google Scholar] [CrossRef]
- Renaud, S.; Thinh, L.; Lambrinidis, G.; Parry, D. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Seixas, P.; Coutinho, P.; Ferreira, M.; Otero, A. Nutritional value of the cryptophyte Rhodomonas lens for Artemia sp. J. Exp. Mar. Biol. 2009, 381, 1–9. [Google Scholar] [CrossRef]
- Dunstan, G.A.; Brown, M.R.; Volkman, J.K. Cryptophyceae and rhodophyceae; chemotaxonomy, phylogeny, and application. Phytochemistry 2005, 66, 2557–2570. [Google Scholar] [CrossRef]
- Fernández-Reiriz, M.J.; Perez-Camacho, A.; Ferreiro, M.J.; Blanco, J.; Planas, M.; Campos, M.J.; Labarta, U. Biomass production and variation in the biochemical profile (total protein, carbohydrates, RNA, lipids and fatty acids) of seven species of marine microalgae. Aquaculture 1989, 83, 17–37. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Sánchez-Bayo, A.; Morales, V.; Rodríguez, R.; Vicente, G.; Bautista, L.F. Cultivation of microalgae and cyanobacteria: Effect of operating conditions on growth and biomass Composition. Molecules 2020, 25, 2834. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of exopolysaccharides production by Porphyridium sordidum and their potential to induce defense responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. [Google Scholar] [CrossRef]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Sui, Y.; Muys, M.; Van de Waal, D.B.; D’Adamo, S.; Vermeir, P.; Fernandes, T.V.; Vlaeminck, S.E. Enhancement of co-production of nutritional protein and carotenoids in Dunaliella salina using a two-phase cultivation assisted by nitrogen level and light intensity. Bioresour. Technol. 2019, 287, 121398. [Google Scholar] [CrossRef] [PubMed]
- Meijer, E.A.; Wijffels, R.H. Development of a fast, reproducible and effective method for the extraction and quantification of proteins of micro-algae. Biotechnol. Technol. 1998, 12, 353–358. [Google Scholar] [CrossRef]
- Corrêa, P.S.; Morais Júnior, W.G.; Martins, A.A.; Caetano, N.S.; Mata, T.M. Microalgae biomolecules: Extraction, separation and purification methods. Processes 2021, 9, 10. [Google Scholar] [CrossRef]
- Allard, B.; Templier, J. Comparison of neutral lipid profile of various trilaminar outer cell wall (TLS)-containing microalgae with emphasis on algaenan occurrence. Phytochemistry 2000, 54, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef]
- Xu, S.; Shen, Y.; Li, Y. Antioxidant activities of sorghum kafirin alcalase hydrolysates and membrane/gel filtrated fractions. Antioxidants 2019, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Kristofersen, K.A.; Amerongen, A.; Böcker, U.; Lindberg, D.; Wubshet, S.G.; de Vogel-van den Bosch, H.; Horn, S.J.; KristianAfseth, N. Fourier-transform infrared spectroscopy for monitoring proteolytic reactions using dryflms treated with trifuoroacetic acid. Sci. Rep. 2020, 10, 7844. [Google Scholar] [CrossRef]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [Green Version]
- Kafle, B.; Böcker, U.; Wubshet, S.G.; Dankel, K.; Måge, I.; O’Farrell, M.; Afseth, N.K. Fourier-transform infrared spectroscopy for characterization of liquid protein solutions: A comparison of two sampling techniques. Vib. Spectrosc. 2023, 124, 103490. [Google Scholar] [CrossRef]
- León-López, A.; Fuentes-Jiménez, L.; Hernández-Fuentes, A.D.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Hydrolysed collagen from sheepskins as a source of functional peptides with antioxidant activity. Int. J. Mol. Sci. 2019, 20, 3931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sala, A.; Spalding, K.E.; Ashton, K.M.; Board, R.; Butler, H.J.; Dawson, T.P.; Harris, D.A.; Hughes, C.S.; Jenkins, C.A.; Jenkinson, M.D.; et al. Fourier-transform infrared spectroscopy of biofluids: A practical approach. Transl. Biophotonics 2020, 3, e202000025. [Google Scholar] [CrossRef]
- Andersen, P.V.; Veiseth-Kent, E.; Wold, J.P. Analyzing pH-induced changes in a myofibril model system with vibrational and fluorescence spectroscopy. Meat Sci. 2017, 125, 1–9. [Google Scholar] [CrossRef]
- Polumbryk, M.; Ivanov, S.; Polumbryk, O. Antioxidants in food systems. Mechanism of action. Ukr. J. Food Sci. 2013, 1, 15–40. [Google Scholar]
- Moaveni, S.; Salami, M.; Khodadadi, M.; McDougall, M.; Emam-Djomeh, Z. Investigation of S. limacinum microalgae digestibility and production of antioxidant bioactive peptides. LWT 2022, 154, 112468. [Google Scholar] [CrossRef]
- Cian, R.E.; Martínez-Augustin, O.; Drago, S.R. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Int. Food Res. J. 2012, 49, 364–372. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gomez, B.; Barba, F.J.; Mora, L.; Perez-Santaescolastica, C.; Toldra, F. Bioactive peptides as natural antioxidants in food products—A review. Trends. Food. Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Alzahrani, M.A.J.; Perera, C.O.; Hemar, Y. Production of bioactive proteins and peptides from the diatom Nitzschia laevis and comparison of their in vitro antioxidant activities with those from Spirulina platensis and Chlorella vulgaris. Int. J. Food Sci. Technol. 2018, 53, 676–682. [Google Scholar] [CrossRef]
- Park, E.Y.; Imazu, H.; Matsumura, Y.; Nakamura, Y.; Sato, K. Effects of peptide fractions with different isoelectric points from wheat gluten hydrolysates on lipid oxidation in pork meat patties. J. Agric. Food Chem. 2012, 60, 7483–7488. [Google Scholar] [CrossRef]
- Sbroggio, M.F.; Montilha, M.S.; Figueiredo, V.R.G.; Georgetti, S.R.; Kurozawa, L.E. Influence of the degree of hydrolysis and type of enzyme on antioxidant activity of okara protein hydrolysates. J. Food Sci. Technol. 2016, 36, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Okeke, E.S.; Enechi, O.C.; Nkwoemeka, N.E. Membrane stabilization, albumin denaturation, protease inhibition, and antioxidant activity as possible mechanisms for the anti-inflammatory effects of flavonoid-rich extract of Peltophorum pterocarpum (DC) K. Heyne (FREPP) Stem Bark. In Proceeding of the First International Electronic Conference on Antioxidants in Health and Disease, Virtual, 1–15 December 2020. [Google Scholar]
- Ruiz-Ruiz, J.C.; Matus-Basto, A.J.; Acereto-Escoffié, P.; Segura-Campos, M.R. Antioxidant and anti-inflammatory activities of phenolic compounds isolated from Melipona beecheii honey. Food Agric. Immunol. 2017, 28, 1424–1437. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Iqbal, M.S.; Ohri, D. In vitro study of anti-inflammatory and antioxidant activity of some medicinal plants and their interrelationship. Asian J. Pharm. Clin. Res. 2018, 11, 2455–3891. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins (% DW) | Lipids (% DW) | Carbohydrates (% DW) | Ash (% DW) | Chlorophylls (% DW) |
|---|---|---|---|---|
| 34.5 ± 0.8 | 13 ± 1 | 14.18 ± 0.16 | 15.73 ± 0.89 | 0.46 ± 0.1 |
| Alcalase Hydrolysate | Pepsin Hydrolysate | Proteins | Wave Number Range (cm−1) | Functional Groups [57,60,61] | |||
|---|---|---|---|---|---|---|---|
| Wave Number (cm−1) | Area | Wave Number (cm−1) | Area | Wave Number (cm−1) | Area | ||
| 3274 | −5.315 | 3269–2920 | N–H stretch O–H stretch | ||||
| 3181 | −365.280 | ||||||
| 3102 | −84.440 | ||||||
| 3088 | −203.607 | ||||||
| 2972 | −93.397 | ||||||
| 2942 | −21.223 | ||||||
| 2927 | −31.459 | ||||||
| 2806 | −37.558 | ||||||
| 2658 | −29.335 | ||||||
| 2605 | −5.525 | ||||||
| 2598 | −39.612 | ||||||
| 2468 | −8.542 | ||||||
| 2094 | −55.081 | ||||||
| 1636 | 188.203 | 1700–1600 | C=O related to Amide I | ||||
| 1628 | −1118.183 | ||||||
| 1549 | −767.192 | 1571 | −1069.794 | 1590–1520 | N–H deformation and C–N stretch at Amide II | ||
| 1527 | −26.986 | ||||||
| 1491 | −611.148 | 1500–1400 | COO– (C-terminal) | ||||
| 1460 | −98.967 | ||||||
| 1436 | −71.050 | ||||||
| 1400 | −14.801 | ||||||
| 1395 | −0.296 | ||||||
| 1388 | −799.001 | ||||||
| 1295 | −909.203 | 1300–1100 | Amide III | ||||
| 1235 | −272.163 | ||||||
| 1136 | −133.919 | ||||||
| 1125 | −308.975 | ||||||
| 1044 | −2188.419 | ~1037 | vibrations of OH groups | ||||
| 1042 | −308.375 | ||||||
| 1035 | −175.511 | ||||||
| 927 | −483.800 | ||||||
| 909 | −162.478 | ||||||
| 888 | −377.804 | ||||||
| 684 | −418.802 | ||||||
| 661 | 103.442 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derbel, H.; Elleuch, J.; Mahfoudh, W.; Michaud, P.; Fendri, I.; Abdelkafi, S. In Vitro Antioxidant and Anti-Inflammatory Activities of Bioactive Proteins and Peptides from Rhodomonas sp. Appl. Sci. 2023, 13, 3202. https://doi.org/10.3390/app13053202
Derbel H, Elleuch J, Mahfoudh W, Michaud P, Fendri I, Abdelkafi S. In Vitro Antioxidant and Anti-Inflammatory Activities of Bioactive Proteins and Peptides from Rhodomonas sp. Applied Sciences. 2023; 13(5):3202. https://doi.org/10.3390/app13053202
Chicago/Turabian StyleDerbel, Hana, Jihen Elleuch, Wael Mahfoudh, Philippe Michaud, Imen Fendri, and Slim Abdelkafi. 2023. "In Vitro Antioxidant and Anti-Inflammatory Activities of Bioactive Proteins and Peptides from Rhodomonas sp." Applied Sciences 13, no. 5: 3202. https://doi.org/10.3390/app13053202