
Improvement of Immune and Hematopoietic Functions by Litsea japonica Fruit Extract in Cyclophosphamide-Treated BALB/c Mice
 , , , and
, , , and 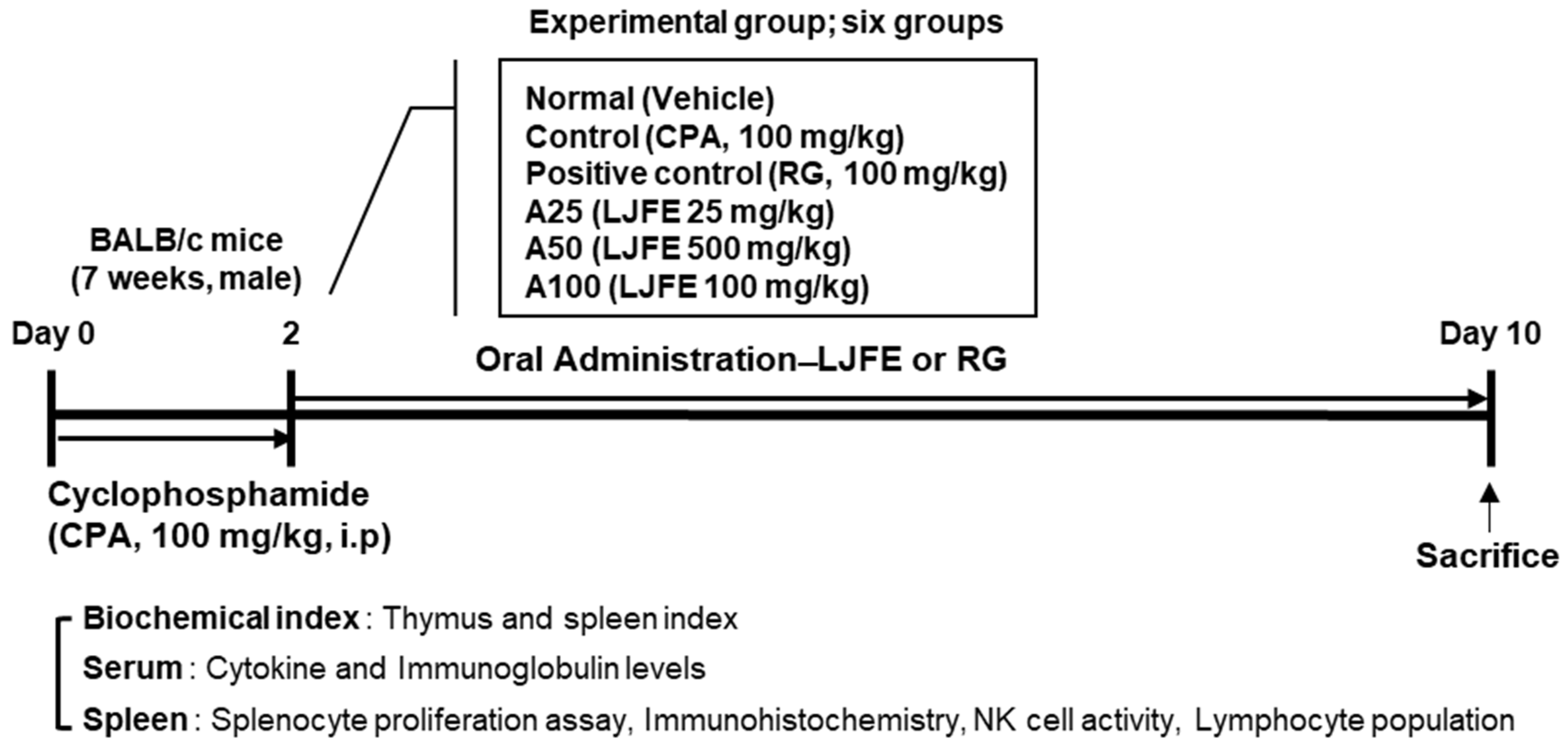{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of LJFE
2.2. Animal Treatment and Experimental Design
2.3. Collection of Spleen and Blood Samples
2.4. Hematological Analysis
2.5. Immune Marker Analysis
2.6. Isolation of Splenocytes
2.7. Measurement of Natural-Killer (NK)-Cell Activity
2.8. Measurement of Splenocyte Proliferation
2.9. Lymphocyte Subpopulation Analysis
2.10. Hematoxylin and Eosin (H&E) Staining
2.11. Statistical Analysis
3. Results
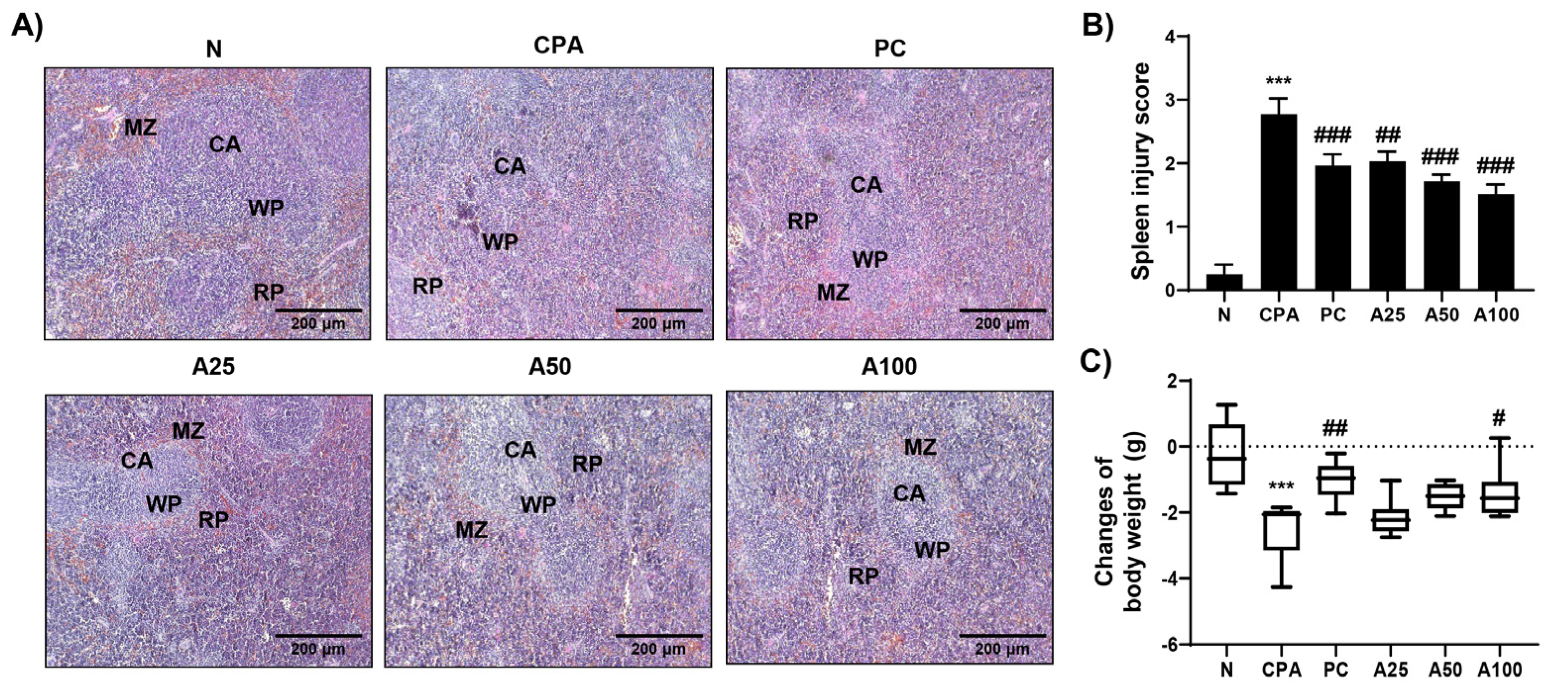
3.1. Effect of LJFE on Spleen and Body Weight
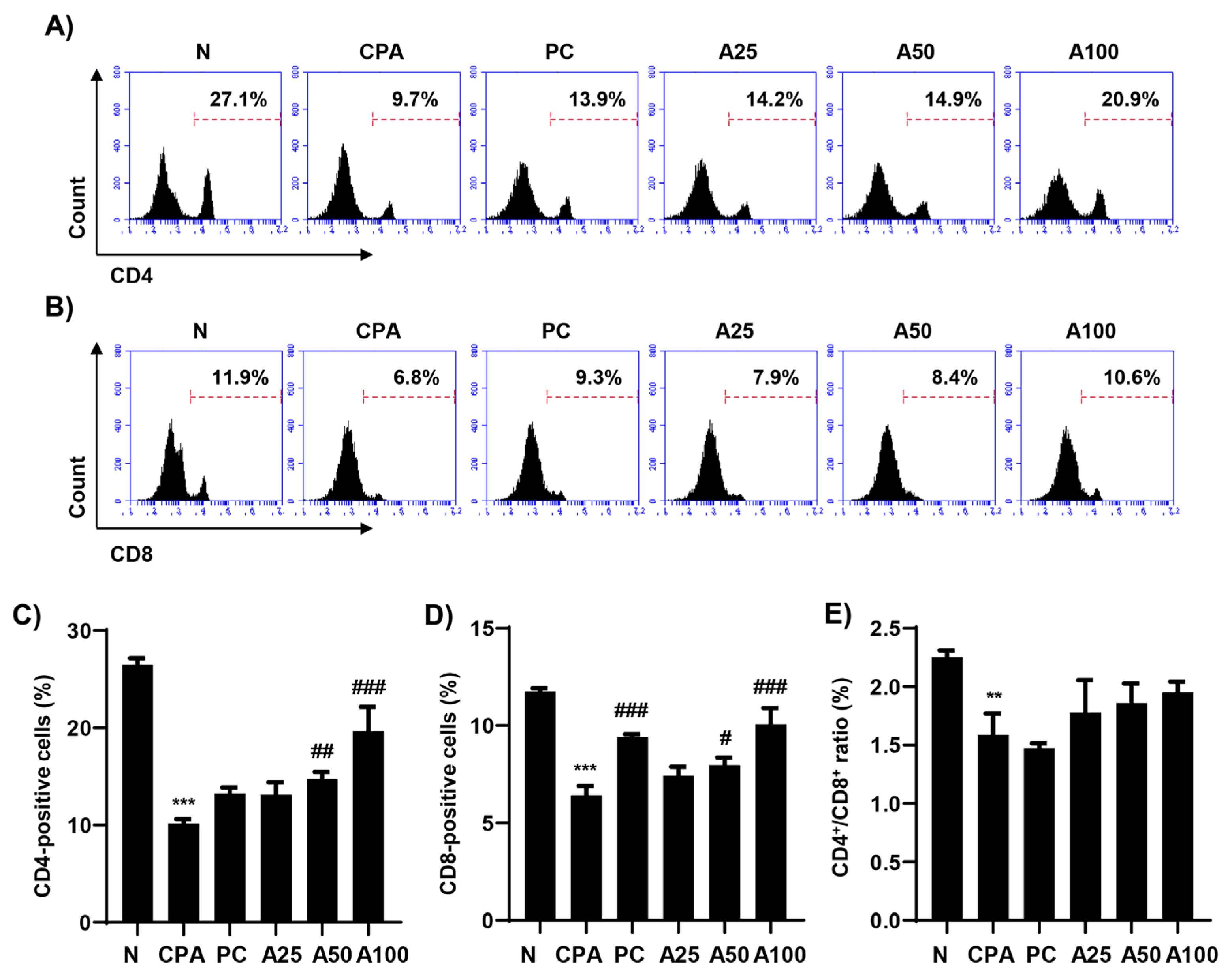
3.2. LJFE Enhanced the Numbers of CD4+ and CD8+ Lymphocytes
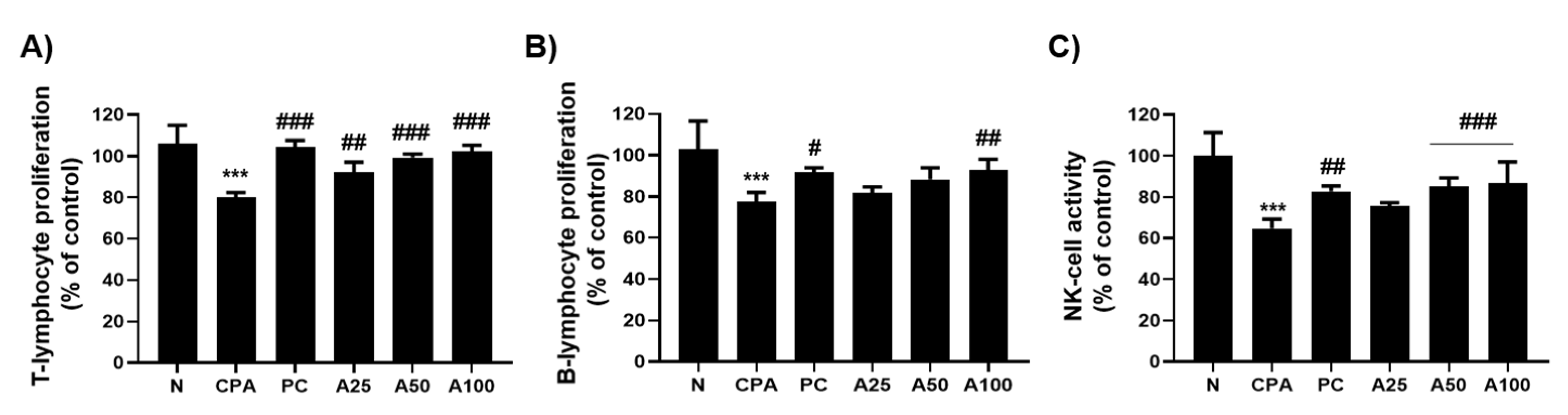
3.3. LJFE Increased Lymphocyte Proliferation and NK-Cell Activity
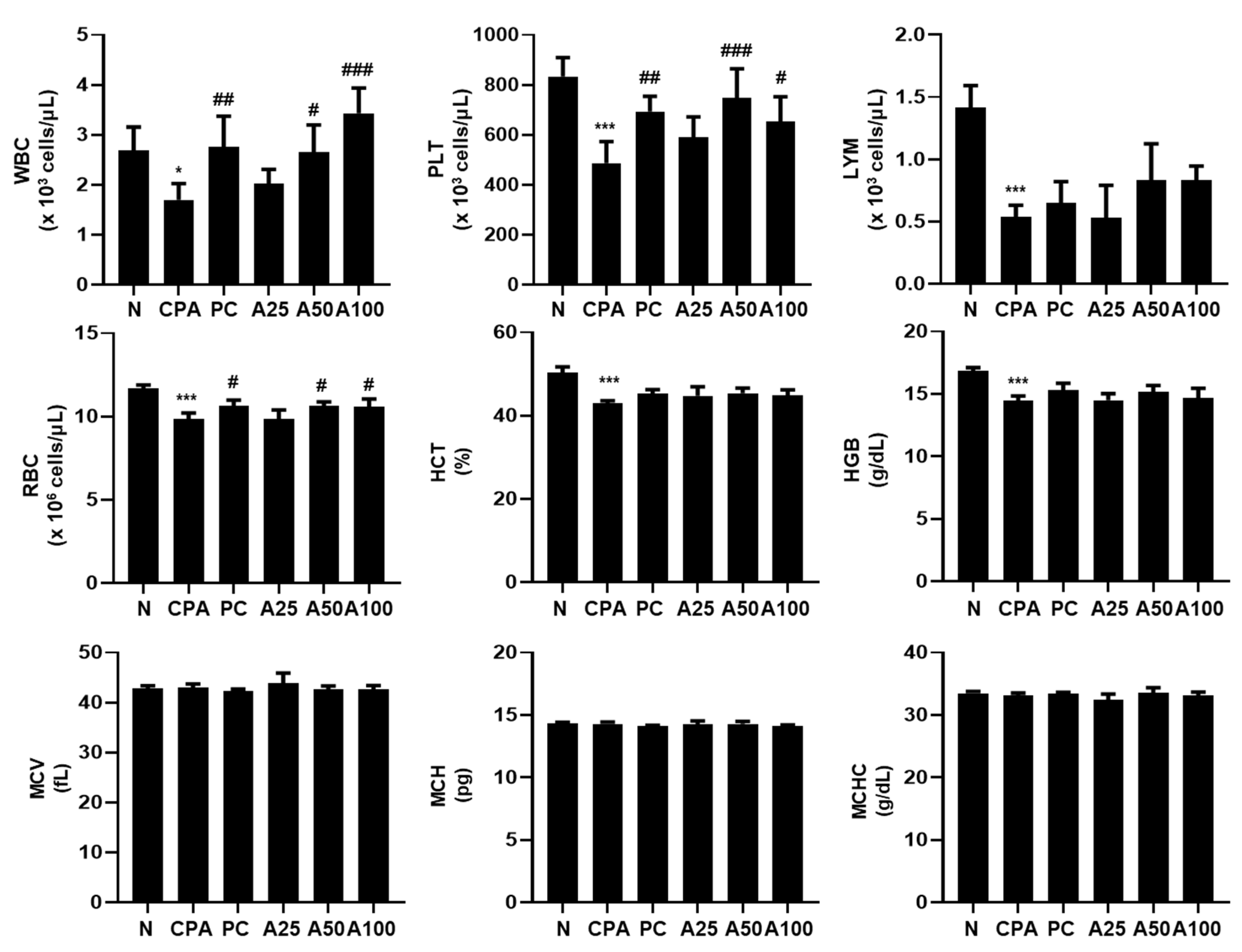
3.4. LJFE Increased the Number of Immunological Peripheral Blood Cells
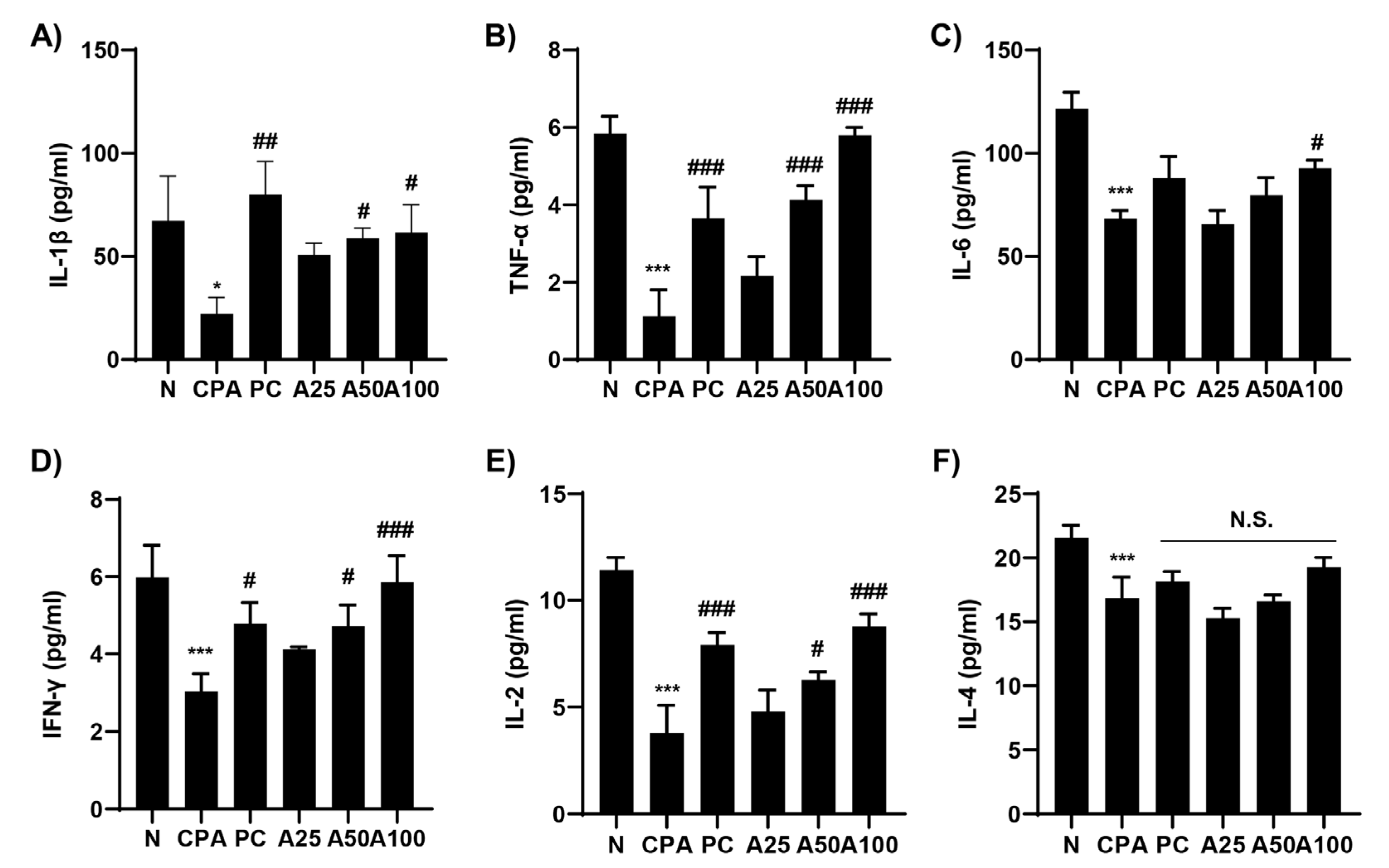
3.5. LJFE Promoted the Production of Immune-Related Cytokines
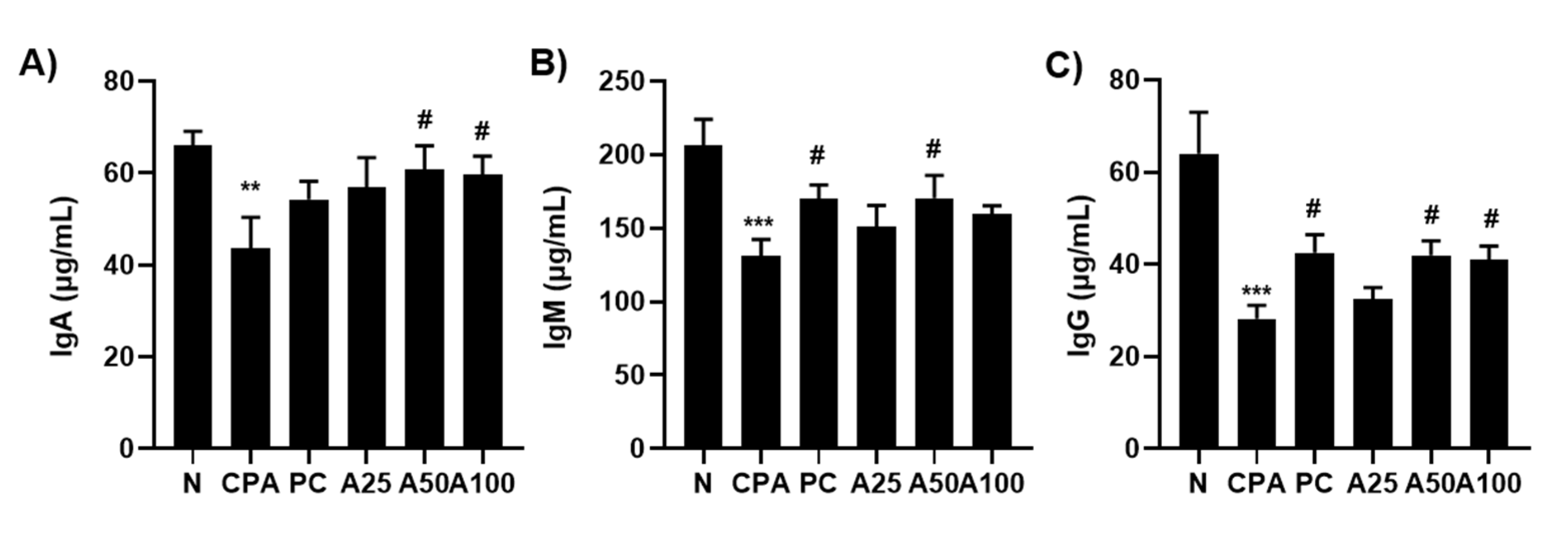
3.6. LJFE Increased the Amount of Immunological Igs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ngo, Q.T.; Cao, T.Q.; Tran, P.L.; Kim, J.A.; Seo, S.T.; Kim, J.C.; Woo, M.H.; Lee, J.H.; Min, B.S. Lactones from the pericarps of Litsea japonica and their anti-inflammatory activities. Bioorg. Med. Chem. Lett. 2018, 28, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Kim, Y.J.; Jung, S.H.; Kim, J.H.; Kim, J.S. Flavonoids from Litsea japonica inhibit AGEs formation and rat lense aldose reductase in vitro and vessel dilation in zebrafish. Planta Med. 2017, 83, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Lee, S.Y.; Kim, J.H.; Kwon, O.K.; Park, B.Y.; An, R.B.; Lee, J.K.; Moon, H.I.; Kim, T.J.; Kim, Y.H.; et al. Lactones from the leaves of Litsea japonica and their anti-complement activity. J. Nat. Prod. 2003, 66, 1388–1390. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Choi, H.J.; Yang, W.K.; Lee, J.E.; Cho, J.H.; Park, I.J.; Park, S.; Park, B.K.; Jin, M. Effect of the n-hexane extract of Litsea japonica fruit flesh on monosodium-iodoacetate-induced osteoarthritis in rats. Evid. Based Complement. Alternat. Med. 2017, 2017, 1791403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.J.; Kim, I.; Cho, J.H.; Park, D.W.; Kwon, J.E.; Jung, M.W.; Meng, X.; Jo, S.M.; Song, H.S.; Cho, Y.M.; et al. Anti-osteoarthritic effects of the Litsea japonica fruit in a rat model of osteoarthritis induced by monosodium iodoacetate. PLoS ONE 2015, 10, e0134856. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, C.S.; Lee, I.S.; Lee, Y.M.; Sohn, E.; Jo, K.; Kim, J.H.; Kim, J.S. Extract of Litsea japonica ameliorates blood-retinal barrier breakdown in db/db mice. Endocrine 2014, 46, 462–469. [Google Scholar] [CrossRef]
- Kim, J.; Kim, C.S.; Lee, Y.M.; Sohn, E.; Jo, K.; Kim, J.S. Litsea japonica extract inhibits neuronal apoptosis and the accumulation of advanced glycation end products in the diabetic mouse retina. Mol. Med. Rep. 2015, 12, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, C.S.; Sohn, E.; Lee, Y.M.; Jo, K.; Kim, J.S. Litsea japonica extract inhibits aldose reductase activity and hyperglycemia-induced lenticular sorbitol accumulation in db/db Mice. Evid. Based Complement Alternat Med. 2015, 2015, 747830. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Min, B.S.; Kim, J.H.; Lee, J.; Kim, T.J.; Kim, C.S.; Kim, Y.H.; Lee, H.K. Flavonoids from the leaves of Litsea japonica and their anti-complement activity. Phytother. Res. 2005, 19, 273–276. [Google Scholar] [CrossRef]
- Park, S.J.; Kang, J.C.; Lee, D.Y.; Cho, J.H.; Yoon, M.H. Immunostimulatory and bone health-promoting activities of Litsea japonica fruit extract. J. Food Hyg. Saf. 2020, 35, 284–289. [Google Scholar] [CrossRef]
- Yun, I.G.; Ahn, S.H.; Yoon, W.J.; Kim, C.S.; Lim, Y.K.; Kook, J.K.; Jung, S.; Choi, C.H.; Lee, T.H. Litsea japonica leaf extract suppresses proinflammatory cytokine production in periodontal ligament fibroblasts stimulated with oral pathogenic bacteria or interleukin-1β. Int. J. Mol. Sci. 2018, 19, 2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.J.; Yoon, W.J.; Sohn, E.H.; Ham, Y.M.; Jang, S.A.; Kwon, J.E.; Jeong, Y.J.; Kwak, J.H.; Sohn, E.; Park, S.Y.; et al. The analgesic and anti-inflammatory effects of Litsea japonica fruit are mediated via suppression of NF-κB and JNK/p38 MAPK activation. Int. Immunopharmacol. 2014, 22, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Bou Zerdan, M.; Moussa, S.; Atoui, A.; Assi, H.I. Mechanisms of immunotoxicity: Stressors and evaluators. Int. J. Mol. Sci. 2021, 22, 8242. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.R.; Efroni, S. The immune system computes the state of the body: Crowd wisdom, machine learning, and immune cell reference repertoires help manage inflammation. Front. Immunol. 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Gray, K.J.; Gibbs, J.E. Adaptive immunity, chronic inflammation and the clock. Semin. Immunopathol. 2022, 44, 209–224. [Google Scholar] [CrossRef]
- Aamir, K.; Sugumar, V.; Khan, H.U.; Looi, C.Y.; Juneja, R.; Waqas, M.; Arya, A. Non-toxic nature of chebulinic acid on biochemical, hematological and histopathological analysis in normal Sprague Dawley rats. Toxicol. Res. 2022, 38, 159–174. [Google Scholar] [CrossRef]
- Yun, W.B.; Kim, J.E.; Lee, M.L.; Choi, J.Y.; Park, J.J.; Song, B.R.; Kang, B.C.; Nam, K.T.; Lee, H.W.; Hwang, D.Y. Sensitivity to tumor development by TALEN-mediated Trp53 mutant genes in the susceptible FVB/N mice and the resistance C57BL/6 mice. Lab. Anim. Res. 2021, 37, 32. [Google Scholar] [CrossRef]
- Borhan, B.A.; Ah, J.Y. The role of andrographolide on skin inflammations and modulation of skin barrier functions in human keratinocyte. Biotechnol. Bioprocess Eng. 2021, 26, 804–813. [Google Scholar] [CrossRef]
- Santana, J.P.P.; Marcato, P.D.; Massaro, T.N.C.; Godoy, N.L.; Anibal, F.F.; Borra, R.C. Efficacy of instillation of MB49 cells and thermoreversible polymeric gel in urothelial bladder carcinoma immunization. Lab. Anim. Res. 2022, 38, 11. [Google Scholar] [CrossRef]
- Park, J.; Heo, Y.J.; Kwon, S. Interaction between hepatocytes and proximal tubular epithelial cells in hypoxia-induced lipotoxicity. Biotechnol. Bioprocess Eng. 2022, 27, 30–39. [Google Scholar] [CrossRef]
- Jeong, M.K.; Kim, B.H. Grading criteria of histopathological evaluation in BCOP assay by various staining methods. Toxicol. Res. 2021, 38, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ahrends, T.; Busselaar, J.; Severson, T.M.; Bąbała, N.; de Vries, E.; Bovens, A.; Wessels, L.; van Leeuwen, F.; Borst, J. CD4+ T cell help creates memory CD8+ T cells with innate and help-independent recall capacities. Nat Commun. 2019, 10, 5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, J.A.; Striker, R. Imbalance in the game of T cells: What can the CD4/CD8 T-cell ratio tell us about HIV and health? PLoS Pathog. 2017, 13, e1006624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Rodríguez, V.; Herrero-Fernández, I.; Castro, M.J.; Castillo, A.; Rosado-Sánchez, I.; Galvá, M.I.; Ramos, R.; Olivas-Martínez, I.; Bulnes-Ramos, Á.; Cañizares, J.; et al. Immunological features beyond CD4/CD8 ratio values in older individuals. Aging 2021, 13, 13443–13459. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Kashimura, M. The human spleen as the center of the blood defense system. Int. J. Hematol. 2020, 112, 147–158. [Google Scholar] [CrossRef]
- Hilligan, K.L.; Ronchese, F. Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cell Mol. Immunol. 2020, 17, 587–599. [Google Scholar] [CrossRef]
- Jansen, J.M.; Gerlach, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J. Clin. Virol. 2019, 119, 44–52. [Google Scholar] [CrossRef]
- Brummelman, J.; Pilipow, K.; Lugli, E. The single-cell phenotypic identity of human CD8+ and CD4+ T cells. Int. Rev. Cell Mol. Biol. 2018, 341, 63–124. [Google Scholar] [CrossRef]
- Heinzel, S.; Marchingo, J.M.; Horton, M.B.; Hodgkin, P.D. The regulation of lymphocyte activation and proliferation. Curr. Opin. Immunol. 2018, 51, 32–38. [Google Scholar] [CrossRef]
- Hu, S.; Wang, X. The role of helper innate lymphoid cells in cancer. Immunotherapy 2019, 11, 1067–1081. [Google Scholar] [CrossRef] [PubMed]
- Kucuksezer, U.C.; Aktas Cetin, E.; Esen, F.; Tahrali, I.; Akdeniz, N.; Gelmez, M.Y.; Deniz, G. The role of natural killer cells in autoimmune diseases. Front. Immunol. 2021, 12, 622306. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Kucia, M. Hematopoiesis and innate immunity: An inseparable couple for good and bad times, bound together by an hormetic relationship. Leukemia 2022, 36, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Andreani, M.; Gaspari, S.; Locatelli, F. Human leucocyte antigen diversity: A biological gift to escape infections, no longer a barrier for haploidentical hemopoietic stem cell transplantation. Int. J. Immunogenet. 2020, 47, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Hippen, K.L.; Hefazi, M.; Larson, J.H.; Blazar, B.R. Emerging translational strategies and challenges for enhancing regulatory T cell therapy for graft-versus-host disease. Front. Immunol. 2022, 13, 926550. [Google Scholar] [CrossRef] [PubMed]
- King, W.; Toler, K.; Woodell-May, J. Role of white blood cells in blood- and bone marrow-based autologous therapies. Biomed Res. Int. 2018, 10, 6510842. [Google Scholar] [CrossRef] [PubMed]
- Cytokines in the balance. Nat. Immunol. 2019, 12, 1557. [CrossRef]
- Sonmez, O.; Sonmez, M. Role of platelets in immune system and inflammation. Porto. Biomed J. 2017, 2, 311–314. [Google Scholar] [CrossRef]
- Anderson, H.L.; Brodsky, I.E.; Mangalmurti, N.S. The evolving erythrocyte: Red blood cells as modulators of innate immunity. J. Immunol. 2018, 201, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Megha, K.B.; Mohanan, P.V. Role of immunoglobulin and antibodies in disease management. Int. J. Biol. Macromol. 2021, 169, 28–38. [Google Scholar] [CrossRef]
- Gorovits, B. Current considerations for immunoglobulin isotype characterization of antibody response against biotherapeutics. AAPS J. 2020, 22, 144. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Chang, C.; Gershwin, M.E. IgA deficiency and autoimmunity. Autoimmun. Rev. 2014, 13, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Zaragoza, M.; Hernández-Ávila, R.; Viedma-Rodríguez, R.; Arenas-Aranda, D.; Ostoa-Saloma, P. Natural and adaptive IgM antibodies in the recognition of tumor-associated antigens of breast cancer (Review). Oncol. Rep. 2015, 34, 1106–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott-Taylor, T.H.; Axinia, S.C.; Amin, S.; Pettengell, R.; Immunoglobulin, G. Structure and functional implications of different subclass modifications in initiation and resolution of allergy. Immun. Inflamm. Dis. 2018, 6, 13–33. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, S.Y.; Bang, E.; Hwangbo, H.; Kim, M.Y.; Kim, D.H.; Koo, Y.T.; Kim, J.S.; Lee, K.W.; Park, S.Y.; Kwon, C.-Y.; et al. Improvement of Immune and Hematopoietic Functions by Litsea japonica Fruit Extract in Cyclophosphamide-Treated BALB/c Mice. Appl. Sci. 2023, 13, 145. https://doi.org/10.3390/app13010145
Ji SY, Bang E, Hwangbo H, Kim MY, Kim DH, Koo YT, Kim JS, Lee KW, Park SY, Kwon C-Y, et al. Improvement of Immune and Hematopoietic Functions by Litsea japonica Fruit Extract in Cyclophosphamide-Treated BALB/c Mice. Applied Sciences. 2023; 13(1):145. https://doi.org/10.3390/app13010145
Chicago/Turabian StyleJi, Seon Yeong, EunJin Bang, Hyun Hwangbo, Min Yeong Kim, Da Hye Kim, Young Tae Koo, Jin Soo Kim, Ki Won Lee, Sun Young Park, Chan-Young Kwon, and et al. 2023. "Improvement of Immune and Hematopoietic Functions by Litsea japonica Fruit Extract in Cyclophosphamide-Treated BALB/c Mice" Applied Sciences 13, no. 1: 145. https://doi.org/10.3390/app13010145