
A Novel Approach for Fast Screening of a Complex Cyanobacterial Extract for Immunomodulatory Properties and Antibacterial Activity

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
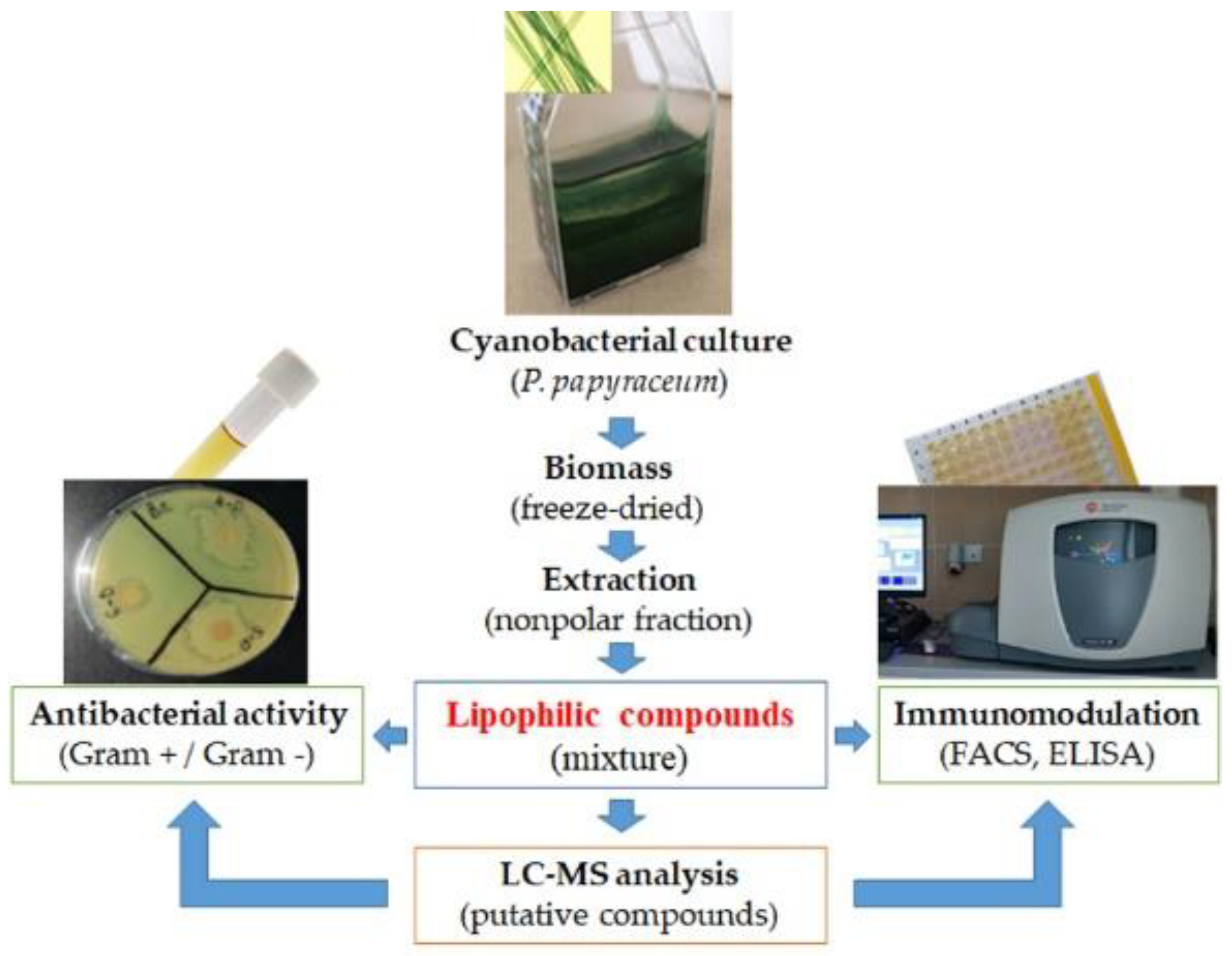
2.2. Cyanobacterial Biomass and Extraction
2.3. Immunomodulatory Properties
2.4. Antibacterial Activity
2.5. LC-MS Analysis
2.5.1. Instrumentation
2.5.2. Reverse Phase (RP) Separation
2.5.3. Mass Spectral Conditions
2.5.4. Data Treatment
2.6. Statistics
3. Results
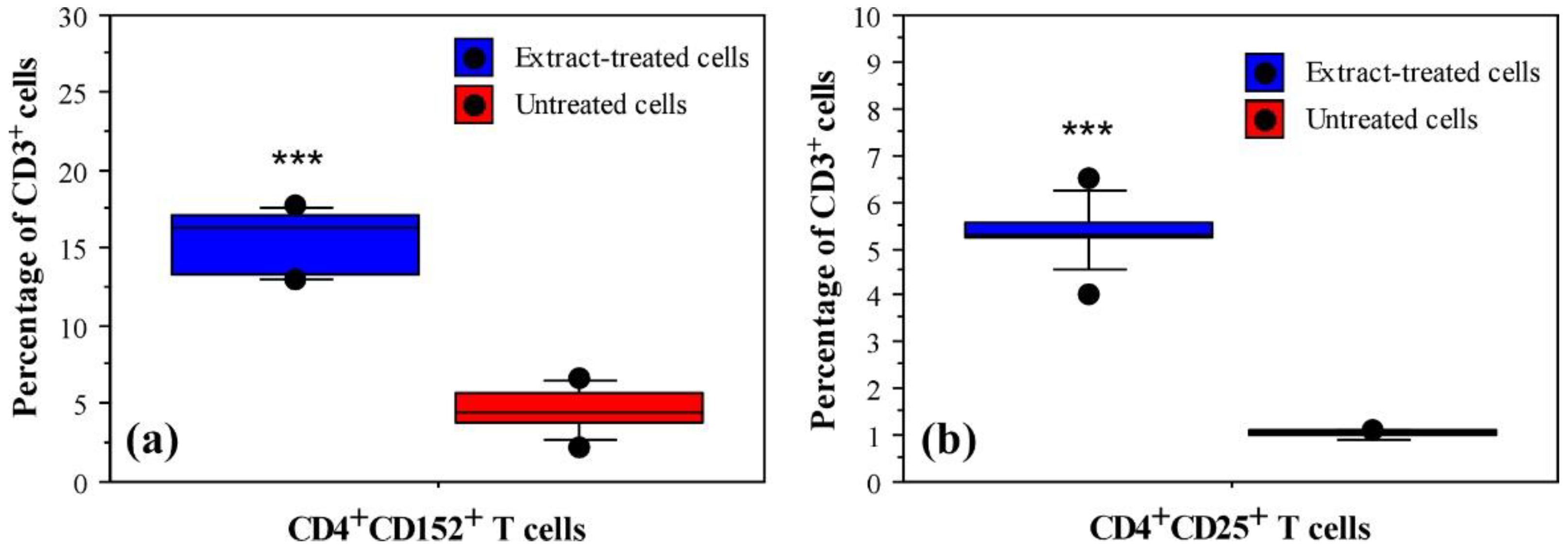
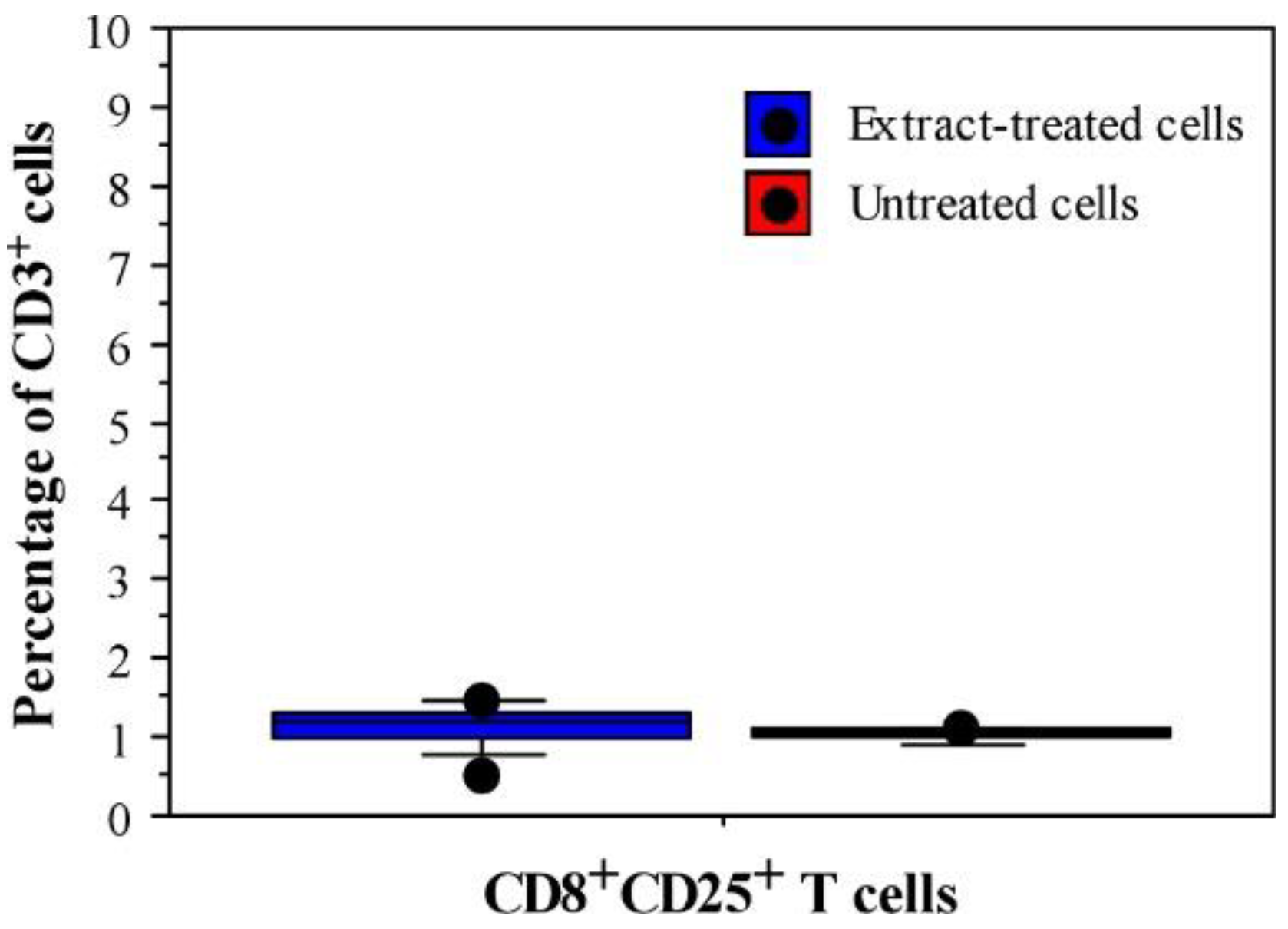
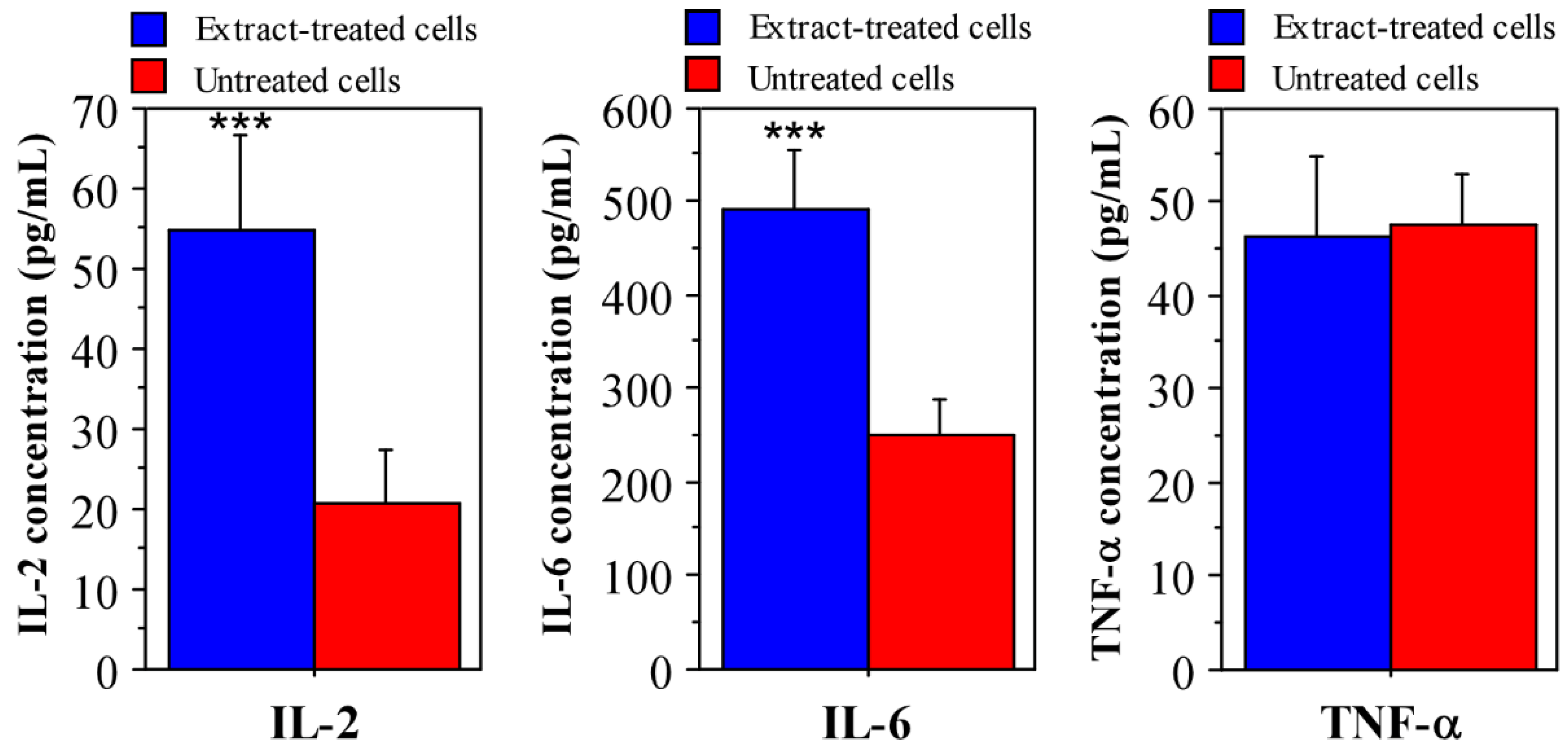
3.1. Immunomodulatory Properties of Phormidium Papyraceum Extract
3.2. Antibacterial Potential of Phormidium Papyraceum Extract
3.3. Chemical Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Morais, J.; Preto, M.; Silva, R.; Urbatzka, R.; Vasconcelos, V.; Reis, M. Uncovering the Bioactive Potential of a Cyanobacterial Natural Products Library Aided by Untargeted Metabolomics. Mar. Drugs 2021, 19, 633. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Grant Burgess, J.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Dzhambazov, B.; Teneva, I.; Mladenov, R.; Popov, N. In vitro cytotoxicity and anticancer properties of two Phormidium molle strains (Cyanoprokaryota). Trav. Sci. Univ. Plovdiv Biol.-Plant. 2006, 39, 3–16. [Google Scholar]
- Dixit, R.B.; Suseela, M.R. Cyanobacteria: Potential candidates for drug discovery. Antonie Van Leeuwenhoek 2013, 103, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T. Anti-infective Natural Products from Cyanobacteria. Planta Med. 2015, 81, 1309–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishido, T.; Humisto, A.; Jokela, J.; Liu, L.; Wahlsten, M.; Tamrakar, A.; Fewer, D.; Permi, P.; Andreote, A.; Fiore, M.; et al. Antifungal Compounds from Cyanobacteria. Mar. Drugs 2015, 13, 2124–2140. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, J.; He, S.; Yan, X. A Review Study on Macrolides Isolated from Cyanobacteria. Mar. Drugs 2017, 15, 126. [Google Scholar] [CrossRef] [Green Version]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2021. [Google Scholar]
- Qamar, H.; Hussain, K.; Soni, A.; Khan, A.; Hussain, T.; Chénais, B. Cyanobacteria as Natural Therapeutics and Pharmaceutical Potential: Role in Antitumor Activity and as Nanovectors. Molecules 2021, 26, 247. [Google Scholar] [CrossRef] [PubMed]
- Papendorf, O.; König, G.M.; Wright, A.D. Hierridin B and 2,4-dimethoxy-6-heptadecyl-phenol, secondary metabolites from the cyanobacterium Phormidium ectocarpi with antiplasmodial activity. Phytochemistry 1998, 49, 2383–2386. [Google Scholar] [CrossRef]
- Sousa, M.L.; Ribeiro, T.; Vasconcelos, V.; Linder, S.; Urbatzka, R. Portoamides A and B are mitochondrial toxins and induce cytotoxicity on the proliferative cell layer of in vitro microtumours. Toxicon 2020, 175, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Marsalek, B.; Sejnohova, L.; von Dohren, H. Detection and identification of oligopeptides in Microcystis (cyanobacteria) colonies: Toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrouzek, P.; Tomek, P.; Lukešová, A.; Urban, J.; Voloshko, L.; Pushparaj, B.; Ventura, S.; Lukavský, J.; Štys, D.; Kopecký, J. Cytotoxicity and secondary metabolites production in terrestrial Nostoc strains, originating from different climatic/geographic regions and habitats: Is their cytotoxicity environmentally dependent? Environ. Toxicol. 2011, 26, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Hrouzek, P.; Kuzma, M.; Cerny, J.; Novak, P.; Fiser, R.; Simek, P.; Lukesova, A.; Kopecky, J. The cyanobacterial cyclic lipopeptides puwainaphycins F/G are inducing necrosis via cell membrane permeabilization and subsequent unusual actin relocalization. Chem. Res. Toxicol. 2012, 25, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Jokela, J.; Oftedal, L.; Herfindal, L.; Permi, P.; Wahlsten, M.; Doskeland, S.O.; Sivonen, K. Anabaenolysins, novel cytolytic lipopeptides from benthic Anabaena cyanobacteria. PLoS ONE 2012, 7, e41222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teneva, I.; Dzhambazov, B.; Koleva, L.; Mladenov, R.; Schirmer, K. Toxic potential of five freshwater Phormidium species (Cyanoprokaryota). Toxicon 2005, 45, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, J.H.; Turnidge, J.D. Manual of Clinical Microbiology, 9th ed.; American Society for Microbiology: Washington, DC, USA, 2007; Volume 1. [Google Scholar]
- Compound Discoverer™ Software. Available online: https://www.thermofisher.com/order/catalog/product/OPTON-31061 (accessed on 5 January 2022).
- Dührkop, K.; Nothias, L.F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C.; et al. Systematic classification of unknown metabolites using high-resolution fragmentation mass spectra. Nat. Biotechnol. 2021, 39, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Lehrstuhl Bioinformatik Jena Home Page. Available online: https://bio.informatik.uni-jena.de/software/sirius/ (accessed on 5 January 2022).
- Science webpage of Dr. Hiroshi Tsugawa for Computational Mass Spectrometry (CompMS). Available online: http://prime.psc.riken.jp/compms/msfinder/main.html (accessed on 5 January 2022).
- PubChem Home Page. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 5 January 2022).
- Caruso, A.; Licenziati, S.; Corulli, M.; Canaris, A.D.; De Francesco, M.A.; Fiorentini, S.; Peroni, L.; Fallacara, F.; Dima, F.; Balsari, A.; et al. Flow cytometric analysis of activation markers on stimulated T cells and their correlation with cell proliferation. Cytometry 1997, 27, 71–76. [Google Scholar] [CrossRef]
- Sermet, S.; Li, J.; Bach, A.; Crawford, R.B.; Kaminski, N.E. Cannabidiol selectively modulates interleukin (IL)-1β and IL-6 production in toll-like receptor activated human peripheral blood monocytes. Toxicology 2021, 464, 153016. [Google Scholar] [CrossRef] [PubMed]
- Dienz, O.; Rincon, M. The effects of IL-6 on CD4 T cell responses. Clin. Immunol. 2009, 130, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewinsohn, D.M.; Ahmed, A.; Adiga, V.; Nayak, S.; Uday Kumar, J.A.J.; Dhar, C.; Sahoo, P.N.; Sundararaj, B.K.; Souza, G.D.; Vyakarnam, A. Circulating HLA-DR+CD4+ effector memory T cells resistant to CCR5 and PD-L1 mediated suppression compromise regulatory T cell function in tuberculosis. PLoS Pathog. 2018, 14, e1007289. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.S.; Asattar, M.A.; Karim, S.; Kutbi, D.; Aljohani, H.; Bakhshwin, D.; Alsieni, M.; Alkreathy, H.M. Pharmacological basis for the potential role of Azithromycin and Doxycycline in management of COVID-19. Arab. J. Chem. 2021, 14, 102983. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.; Prathapan, P. Broad-spectrum therapeutics: A new antimicrobial class. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100011. [Google Scholar] [CrossRef] [PubMed]
- Renda, G.; Gökkaya, İ.; Şöhretoğlu, D. Immunomodulatory properties of triterpenes. Phytochem. Rev. 2021. [Google Scholar] [CrossRef]
- Villarino, N.; Martín-Jiménez, T. Pharmacokinetics of macrolides in foals. J. Vet. Pharmacol. Ther. 2013, 36, 1–13. [Google Scholar] [CrossRef]
- Siddiqui, I.N.; Zahoor, A.; Hussain, H.; Ahmed, I.; Ahmad, V.U.; Padula, D.; Draeger, S.; Schulz, B.; Meier, K.; Steinert, M.; et al. Diversonol and Blennolide Derivatives from the Endophytic Fungus Microdiplodia sp.: Absolute Configuration of Diversonol. J. Nat. Prod. 2011, 74, 365–373. [Google Scholar] [CrossRef]
- Han, S.; Pham, T.-V.; Kim, J.-H.; Lim, Y.-R.; Park, H.-G.; Cha, G.-S.; Yun, C.-H.; Chun, Y.-J.; Kang, L.-W.; Kim, D. Functional characterization of CYP107W1 from Streptomyces avermitilis and biosynthesis of macrolide oligomycin A. Arch. Biochem. Biophys. 2015, 575, 1–7. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “New” fungal carotenoids for industry? Microb. Cell Factories 2018, 17, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Shang, J.; Song, C.; Yang, R.; Shang, X.; Mao, W.; Bao, D.; Tan, Q. Untargeted Metabolite Profiling of Antimicrobial Compounds in the Brown Film of Lentinula edodes Mycelium via LC–MS/MS Analysis. ACS Omega 2020, 5, 7567–7575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loya, S.; Reshef, V.; Mizrachi, E.; Silberstein, C.; Rachamim, Y.; Carmeli, S.; Hizi, A. The inhibition of the reverse transcriptase of HIV-1 by the natural sulfoglycolipids from cyanobacteria: Contribution of different moieties to their high potency. J. Nat. Prod. 1998, 61, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New acylated sulfoglycolipids and digalactolipids and related known glycolipids from cyanobacteria with a potential to inhibit the reverse transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, L.; Zhu, T.; Ba, M.; Li, G.; Gu, Q.; Guo, Y.; Li, D. Phenylspirodrimanes with Anti-HIV Activity from the Sponge-Derived Fungus Stachybotrys chartarum MXH-X73. J. Nat. Prod. 2013, 76, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Li, H.; Zhang, S.; Lu, H.; Chen, Q. A Review on Preparation of Betulinic Acid and Its Biological Activities. Molecules 2021, 26, 5583. [Google Scholar] [CrossRef] [PubMed]
- Hemtasin, C.; Kanokmedhakul, S.; Kanokmedhakul, K.; Hahnvajanawong, C.; Soytong, K.; Prabpai, S.; Kongsaeree, P. Cytotoxic Pentacyclic and Tetracyclic Aromatic Sesquiterpenes from Phomopsis archeri. J. Nat. Prod. 2011, 74, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Rosowsky, A.; Kim, S.H.; Ross, J.; Wick, M.M. Lipophilic 5′-alkyl phosphate esters of 1-.beta.-d-arabinofuranosylcytosine and its N4-acyl and 2,2′-anhydro-3′-O-acyl derivatives as potential prodrugs. J. Med. Chem. 2002, 25, 171–178. [Google Scholar] [CrossRef]
- Hamamura, E.K.; Prystasz, M.; Verheyden, J.P.H.; Moffatt, J.G.; Yamaguchi, K.; Uchida, N.; Sato, K.; Nomura, A.; Shiratori, O. Reactions of 2-acyloxyisobutyryl halides with nucleosides. 8. Synthesis and biological evaluation of some 3′-acyl and 3′,5′-diacyl derivatives of 1-.beta.-d-arabinofuranosylcytosine. J. Med. Chem. 2002, 19, 667–674. [Google Scholar] [CrossRef]
- Reutrakul, V.; Anantachoke, N.; Pohmakotr, M.; Jaipetch, T.; Sophasan, S.; Yoosook, C.; Kasisit, J.; Napaswat, C.; Santisuk, T.; Tuchinda, P. Cytotoxic and Anti-HIV-1 Caged Xanthones from the Resin and Fruits of Garcinia hanburyi. Planta Med. 2006, 73, 33–40. [Google Scholar] [CrossRef]
- Hahnvajanawong, C.; Sahakulboonyarak, T.; Boonmars, T.; Reutrakul, V.; Kerdsin, A.; Boueroy, P. Inhibitory effect of isomorellin on cholangiocarcinoma cells via suppression of NF-κB translocation, the phosphorylated p38 MAPK pathway and MMP-2 and uPA expression. Exp. Ther. Med. 2020, 21, 151. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.; Lin, L.; Li, H. Gambogic Acid as a Candidate for Cancer Therapy: A Review. Int. J. Nanomed. 2020, 15, 10385–10399. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-Inflammatory and Anticancer Effects of Microalgal Carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef] [PubMed]
- Kuramochi, K. Synthetic and Structure-Activity Relationship Studies on Bioactive Natural Products. Biosci. Biotechnol. Biochem. 2014, 77, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Yasui, H.; Ohmori, A.; Tokuda, H.; Suzuki, N.; Osawa, A.; Shindo, K.; Ishibashi, T. Anti-oxidative, anti-tumor-promoting, and anti-carcinogenic activities of adonirubin and adonixanthin. J. Oleo Sci. 2013, 62, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Dalisay, D.S.; Li, F.; Amphlett, J.; Maneerat, W.; Chavez, M.A.G.; Wang, Y.A.; Matainaho, T.; Yu, W.; Brown, P.J.; et al. Nahuoic Acid A Produced by a Streptomyces sp. Isolated From a Marine Sediment Is a Selective SAM-Competitive Inhibitor of the Histone Methyltransferase SETD8. Org. Lett. 2012, 15, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Long, J.Z.; Li, W.; Booker, L.; Burston, J.J.; Kinsey, S.G.; Schlosburg, J.E.; Pavón, F.J.; Serrano, A.M.; Selley, D.E.; Parsons, L.H.; et al. Selective blockade of 2-arachidonoylglycerol hydrolysis produces cannabinoid behavioral effects. Nat. Chem. Biol. 2008, 5, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solanki, H.; Pierdet, M.; Thomas, O.P.; Zubia, M. Insights into the Metabolome of the Cyanobacterium Leibleinia gracilis from the Lagoon of Tahiti and First Inspection of Its Variability. Metabolites 2020, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Symersky, J.; Osowski, D.; Walters, D.E.; Mueller, D.M. Oligomycin frames a common drug-binding site in the ATP synthase. Proc. Natl. Acad. Sci. USA 2012, 109, 13961–13965. [Google Scholar] [CrossRef] [Green Version]
- Capó, X.; Martorell, M.; Tur, J.A.; Sureda, A.; Pons, A. 5-Dodecanolide, a Compound Isolated from Pig Lard, Presents Powerful Anti-Inflammatory Properties. Molecules 2021, 26, 7363. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Bisht, A.; Verma, K.; Negi, S.; Paliwal, S.; Sharma, S. The role of fatty acid amide hydrolase enzyme inhibitors in Alzheimer’s disease. Cell Biochem. Funct. 2021. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Lin, C.-Y.; Chung, Y.-P.; Liu, C.-H.; Huang, K.-T.; Guan, S.-S.; Wu, C.-T.; Liu, S.-H. Protective Effects of Nootkatone on Renal Inflammation, Apoptosis, and Fibrosis in a Unilateral Ureteral Obstructive Mouse Model. Nutrients 2021, 13, 3921. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, S.; Rivetti, S.; Triarico, S.; Romano, A.; Attinà, G.; Maurizi, P.; Ruggiero, A. Mechanisms, Characteristics, and Treatment of Neuropathic Pain and Peripheral Neuropathy Associated with Dinutuximab in Neuroblastoma Patients. Int. J. Mol. Sci. 2021, 22, 12648. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.L.; Cronholm, P.; Hedberg, Y.; Tornberg, M.; De Battice, L.; Svedhem, S.; Wallinder, I.O. Cell membrane damage and protein interaction induced by copper containing nanoparticles—Importance of the metal release process. Toxicology 2013, 313, 59–69. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
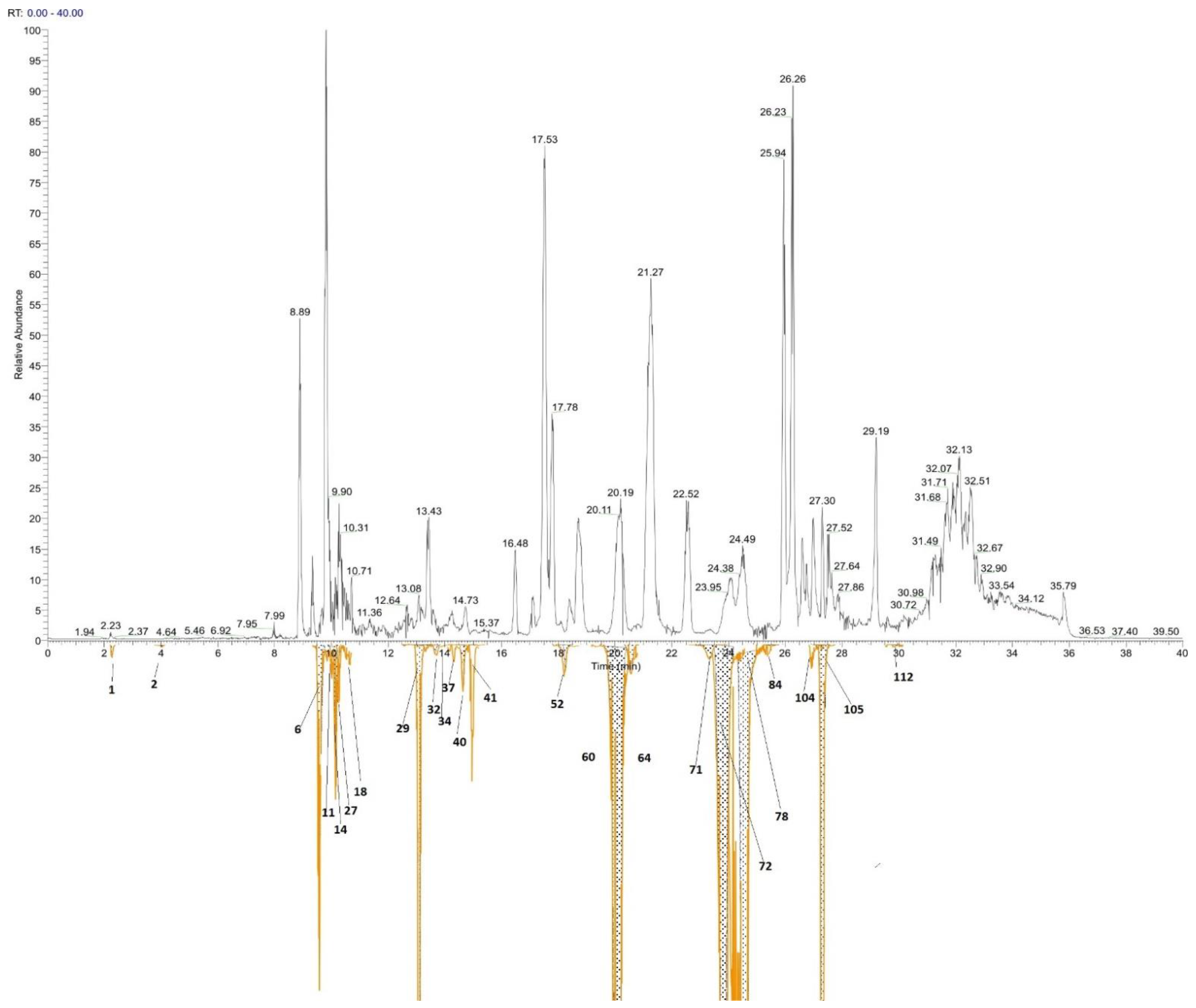{kind=link}
| No. | Time | Solvent A | Solvent B | Solvent C | Flow Rate |
|---|---|---|---|---|---|
| (min) | (%) | (%) | (%) | (μL/min) | |
| 0 | 0.0 | 85 | 15 | 0 | 600 |
| 1 | 0.2 | 70 | 30 | 0 | 250 |
| 2 | 1.0 | 65 | 35 | 0 | 250 |
| 3 | 8.0 | 5 | 95 | 0 | 250 |
| 4 | 19.0 | 0 | 100 | 0 | 250 |
| 5 | 21.0 | 0 | 100 | 0 | 250 |
| 6 | 22.5 | 0 | 60 | 40 | 250 |
| 7 | 30.0 | 0 | 20 | 80 | 250 |
| 8 | 32.5 | 0 | 0 | 100 | 250 |
| 9 | 34.0 | 0 | 0 | 100 | 450 |
| 10 | 35.0 | 85 | 15 | 0 | 600 |
| 11 | 42.0 | 85 | 15 | 0 | 600 |
| CD Markers | Extract-Treated Cells | Untreated Cells (Negative Control) | PHA-L Treated Cells (Positive Control) |
|---|---|---|---|
| CD3 (%) | 22.10 ± 3.92 a | 11.91 ± 4.47 | 28.30 ± 3.13 |
| CD4 (%) | 18.87 ± 2.00 a | 10.57 ± 3.5 | 24.37 ± 5.42 |
| CD8 (%) | 2.10 ± 0.02 | 2.29 ± 1.15 | 10.64 ± 1.07 |
| CD25 (%) | 5.30 ± 1.24 a | 1.02 ± 0.08 | 8.04 ± 1.12 |
| CD152 (%) | 15.70 ± 2.18 a | 4.64 ± 2.18 | 32.60 ± 4.35 |
| CD19 (%) | 3.29 ± 0.19 | 4.55 ± 0.51 | 3.45 ± 0.47 |
| HLA-DR-DP (%) | 11.12 ± 1.79 a | 4.27 ± 0.99 | 12.13 ± 1.80 |
| CD11b (%) | 1.12 ± 0.38 | 1.03 ± 0.02 | 1.38 ± 0.22 |
| CD138 (%) | 2.62 ± 0.39 | 1.42 ± 0.09 | 1.88 ± 0.19 |
| CD80 (%) | 2.26 ± 1.91 | 2.12 ± 0.97 | 1.46 ± 0.13 |
| CD16 (%) | 1.16 ± 0.02 | 1.12 ± 0.04 | 1.16 ± 0.05 |
| CD56 (%) | 1.45 ± 0.90 | 2.79 ± 0.29 | 4.27 ± 0.56 |
| Bacterial Strains | P. Papyraceum Extract | Antibiotics | DMSO/Water (1:1) |
|---|---|---|---|
| Gram-Negative: | |||
| Escherichia coli | 20.28 ± 1.50 | 25.78 ± 1.82 | 0.00 ± 0.00 |
| Pseudomonas aeruginosa | 18.25 ± 0.50 | 27.67 ± 1.53 | 0.00 ± 0.00 |
| Proteus mirabilis | 19.33 ± 2.08 | 26.25 ± 2.06 | 0.00 ± 0.00 |
| Gram-Positive: | |||
| Bacillus cereus | 17.00 ± 0.82 | 24.50 ± 1.00 | 0.00 ± 0.00 |
| Bacillus subtilis | 16.50 ± 1.29 | 25.00 ± 0.58 | 0.00 ± 0.00 |
| Staphylococcus aureus | 10.86 ± 0.85 | 24.58 ± 1.26 | 0.00 ± 0.00 |
| No. | RT [min] | Name | Formula | Molecular Weight | FISH Coverage Score | Structure * |
|---|---|---|---|---|---|---|
| 1 | 2.69 | Gabapentin | C9H17NO2 | 171.13 | 42.31 |  |
| 2 | 4.39 | Istamycin C1 | C19H37N5O6 | 431.27 | 42.86 |  |
| 6 | 9.82 | Autumnolide | C15H20O5 | 280.13 | 38.78 |  |
| 11 | 10.04 | Mueggelone | C18H28O3 | 292.20 | 76.92 |  |
| 14 | 10.40 | Diversonol | C15H18O6 | 294.11 | 41.38 |  |
| 17 | 10.55 | Phomoarcherin B | C23H28O5 | 384.19 | 26.67 |  |
| 18 | 10.66 | Kampanol A | C25H32O6 | 428.22 | 28.57 |  |
| 29 | 12.42 | Nahuoic acid A | C30H50O7 | 522.35 | 26.82 |  |
| 32 | 13.33 | Adonirubin (Phoenicoxanthin) | C18H18 | 580.39 | 41.98 |  |
| 34 | 13.59 | Palmitic amide | C16H33NO | 255.25 | 70.00 |  |
| 37 | 13.92 | Oleamide | C18H35NO | 281.27 | 91.67 |  |
| 40 | 14.91 | 3-hydroxyechinenone | C40H54O2 | 566.41 | 38.33 |  |
| 41 | 14.94 | Canthaxanthin | C40H52O2 | 564.39 | 45.00 |  |
| 52 | 18.40 | Erucamide | C22H43NO | 337.33 | 75.76 |  |
| 60 | 20.33 | Nootkatone | C15H22O | 218.17 | 29.44 |  |
| 64 | 20.98 | Torularhodin | C40H52O2 | 564.39 | 37.97 |  |
| 71 | 23.45 | Tanikolide | C17H32O3 | 284.23 | 67.65 |  |
| 72 | 23.96 | (2S)-3-(beta-d-galactopyrano syloxy)-2-(palmitoyloxy)propyl (9Z,12Z,15Z)-9,12,15-octadecatrienoate | C43H76O10 | 752.54 | 100 |  |
| 78 | 24.65 | Oligomycin C | C45H74O10 | 774.52 | 33.82 |  |
| 84 | 25.52 | Azithromycin | C38H72N2O12 | 748.50 | 28.22 |  |
| 101 | 27.21 | Plakevulin A | C23H42O4 | 382.30 | 25.14 |  |
| 104 | 27.37 | 1,2-dipalmitoyl-3-beta-d-galactosyl-sn-glycerol | C41H78O10 | 730.56 | 35.38 |  |
| 105 | 27.39 | 1-(9Z-octadecenoyl)-2-hexadecanoyl-3-beta-d-galactosyl-sn-glycerol | C43H80O10 | 756.58 | 34.62 |  |
| 112 | 29.07 | Betulin | C30H50O2 | 442.38 | 66.67 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teneva, I.; Batsalova, T.; Bardarov, K.; Moten, D.; Dzhambazov, B. A Novel Approach for Fast Screening of a Complex Cyanobacterial Extract for Immunomodulatory Properties and Antibacterial Activity. Appl. Sci. 2022, 12, 2847. https://doi.org/10.3390/app12062847
Teneva I, Batsalova T, Bardarov K, Moten D, Dzhambazov B. A Novel Approach for Fast Screening of a Complex Cyanobacterial Extract for Immunomodulatory Properties and Antibacterial Activity. Applied Sciences. 2022; 12(6):2847. https://doi.org/10.3390/app12062847
Chicago/Turabian StyleTeneva, Ivanka, Tsvetelina Batsalova, Krum Bardarov, Dzhemal Moten, and Balik Dzhambazov. 2022. "A Novel Approach for Fast Screening of a Complex Cyanobacterial Extract for Immunomodulatory Properties and Antibacterial Activity" Applied Sciences 12, no. 6: 2847. https://doi.org/10.3390/app12062847