
Skin Antiaging Effects of the Fermented Outer Layers of Leaf Skin of Aloe barbadensis Miller Associated with the Enhancement of Mitochondrial Activities of UVb-Irradiated Human Skin Fibroblasts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Samples
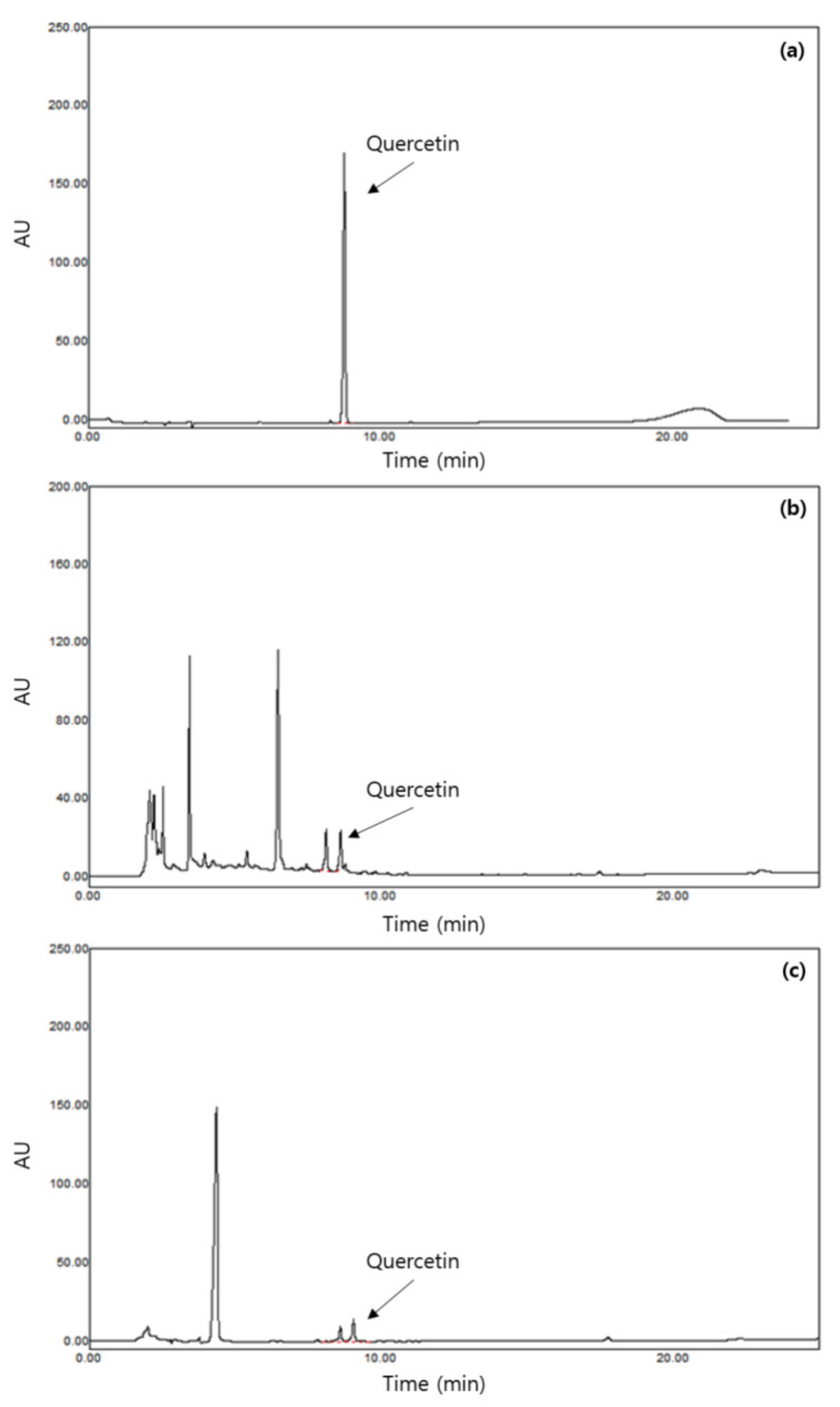
2.2. Measurement of Quercetin Concentrations in the Samples
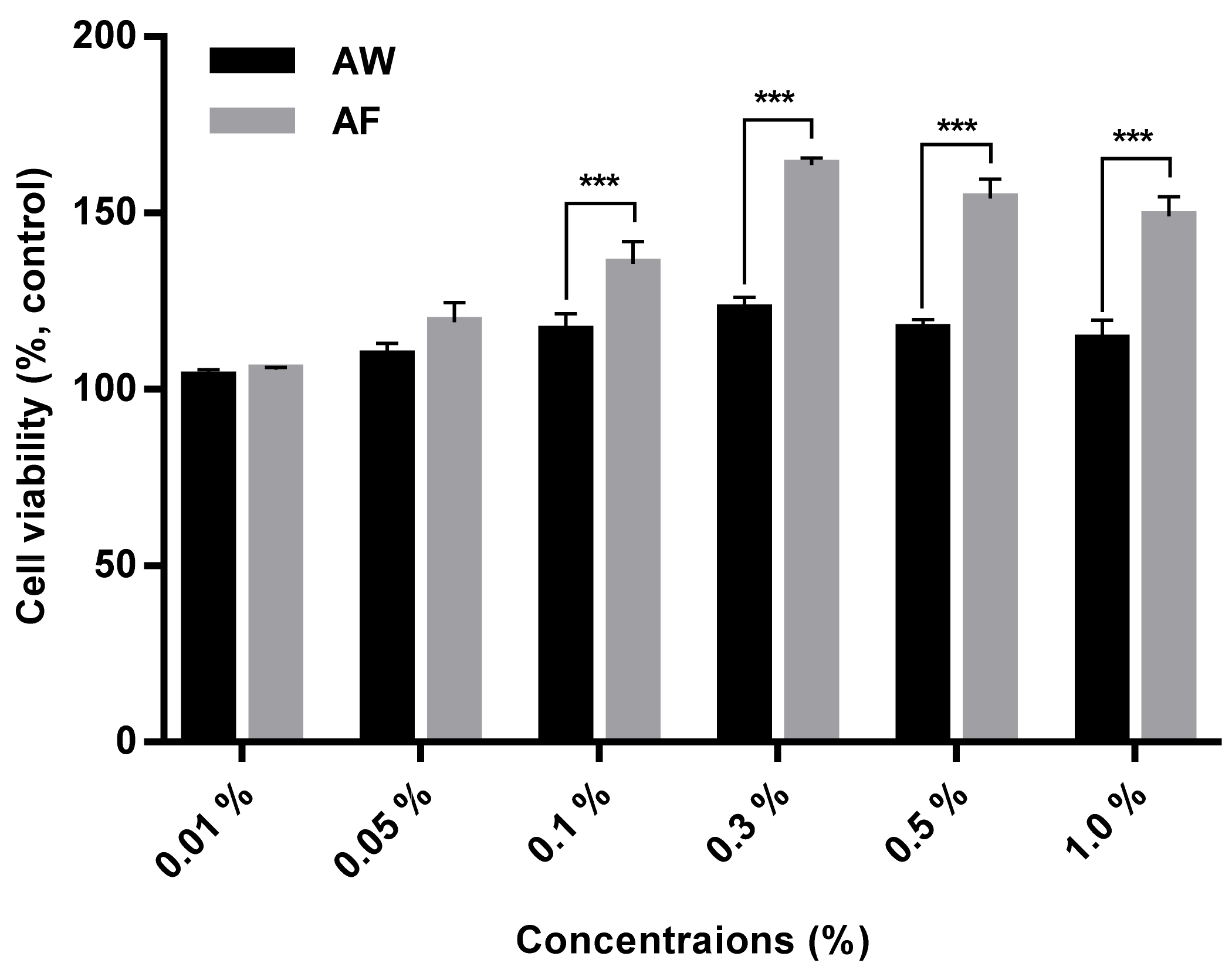
2.3. Measurement of the Cytotoxicity of the Samples
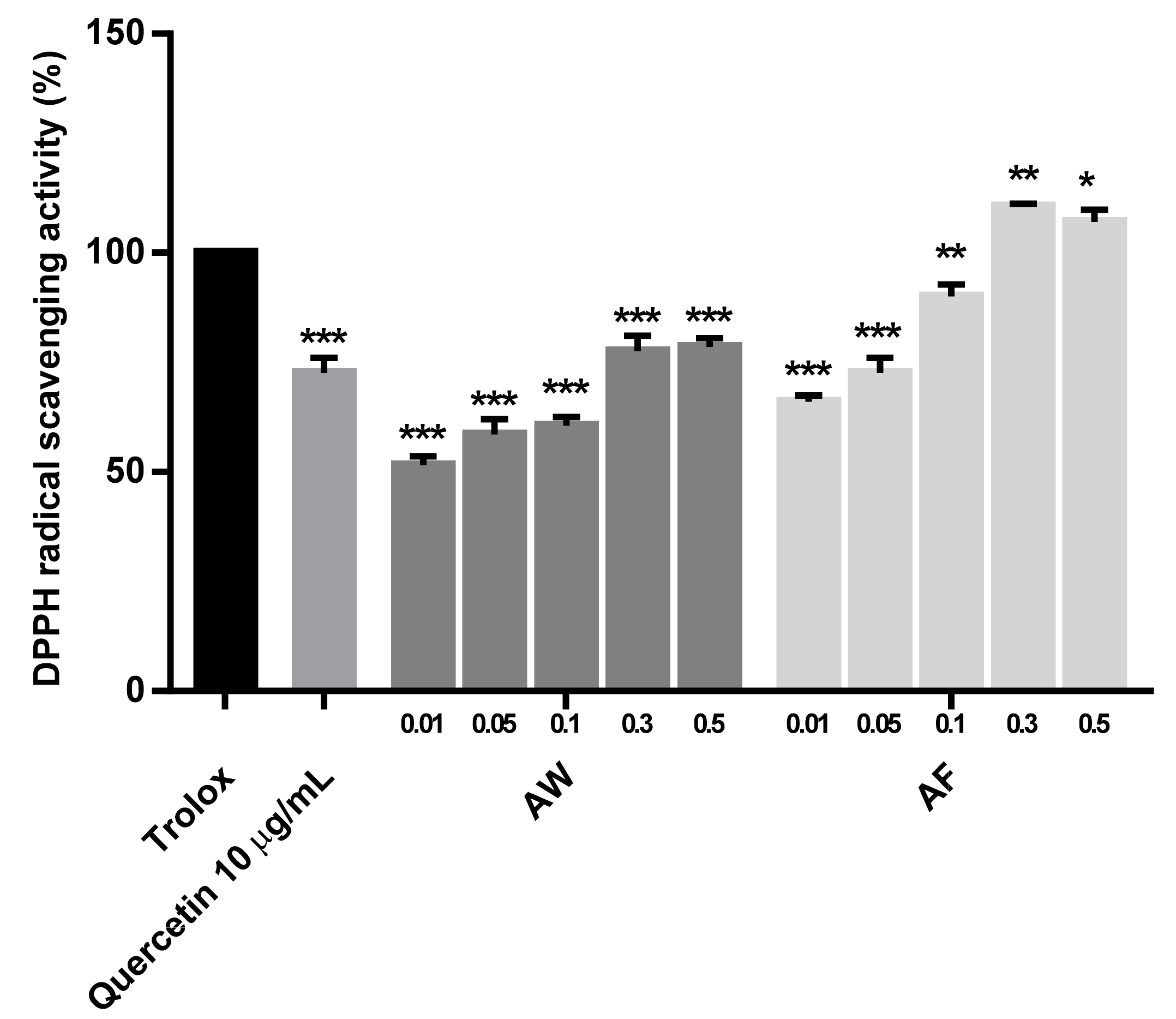
2.4. Measurement of the Antioxidant Activities of the Samples
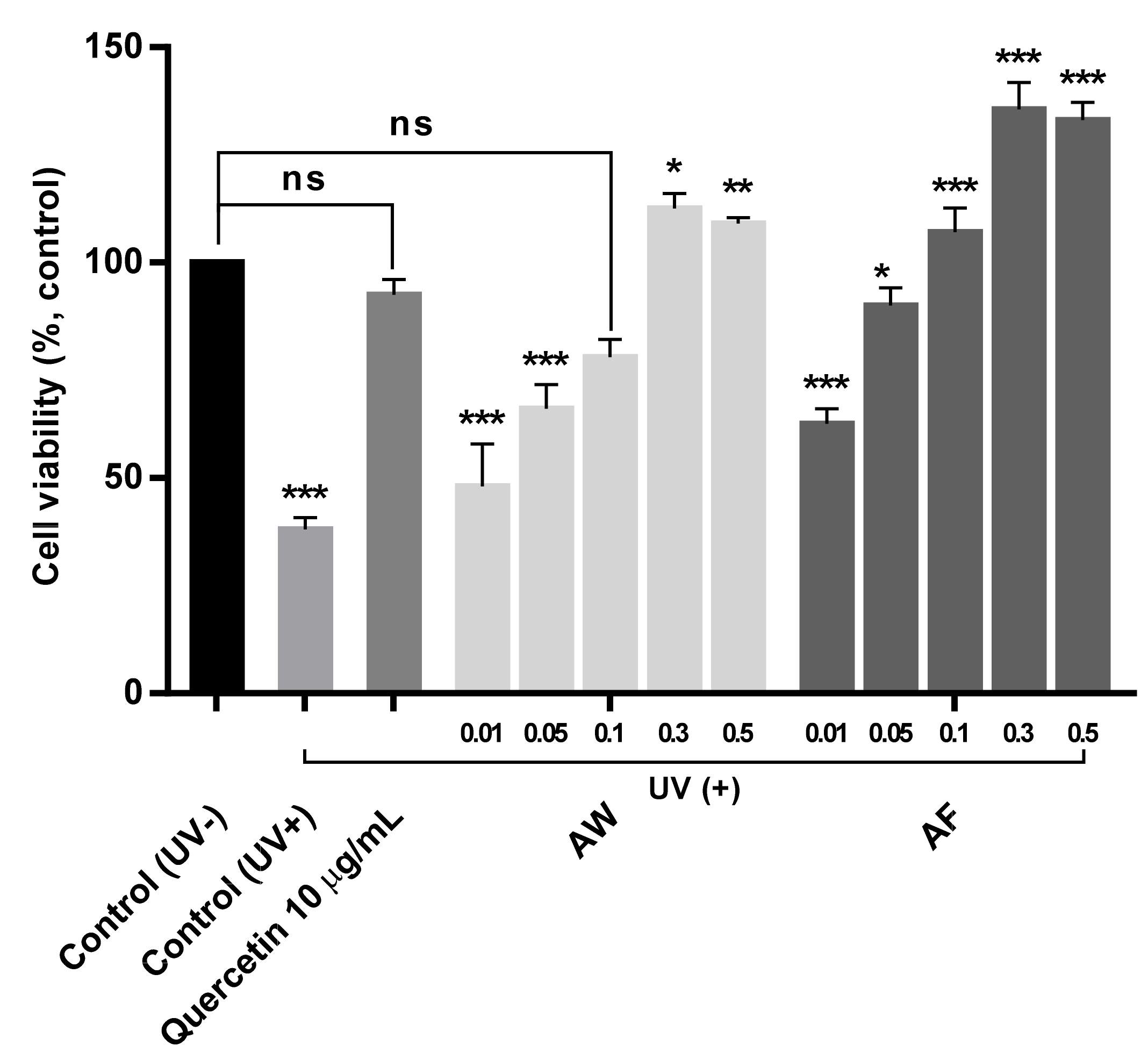
2.5. Estimation of Mitochondrial Activities in UVb-Irradiated Human Fibroblasts
2.6. Measurement of Collagen Production by UVb-Irradiated Human Fibroblasts Treated with the Samples
2.7. Measurement of the Inhibition of Matrix Metalloproteinase-1 (MMP-1) Synthesis by the Samples
2.8. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.9. Statistical Analysis
3. Results and Discussion
3.1. Concentrations of Quercetin in the Samples
3.2. Cytotoxicity and the Prevention of Mitochondrial Dysfunction by the Samples
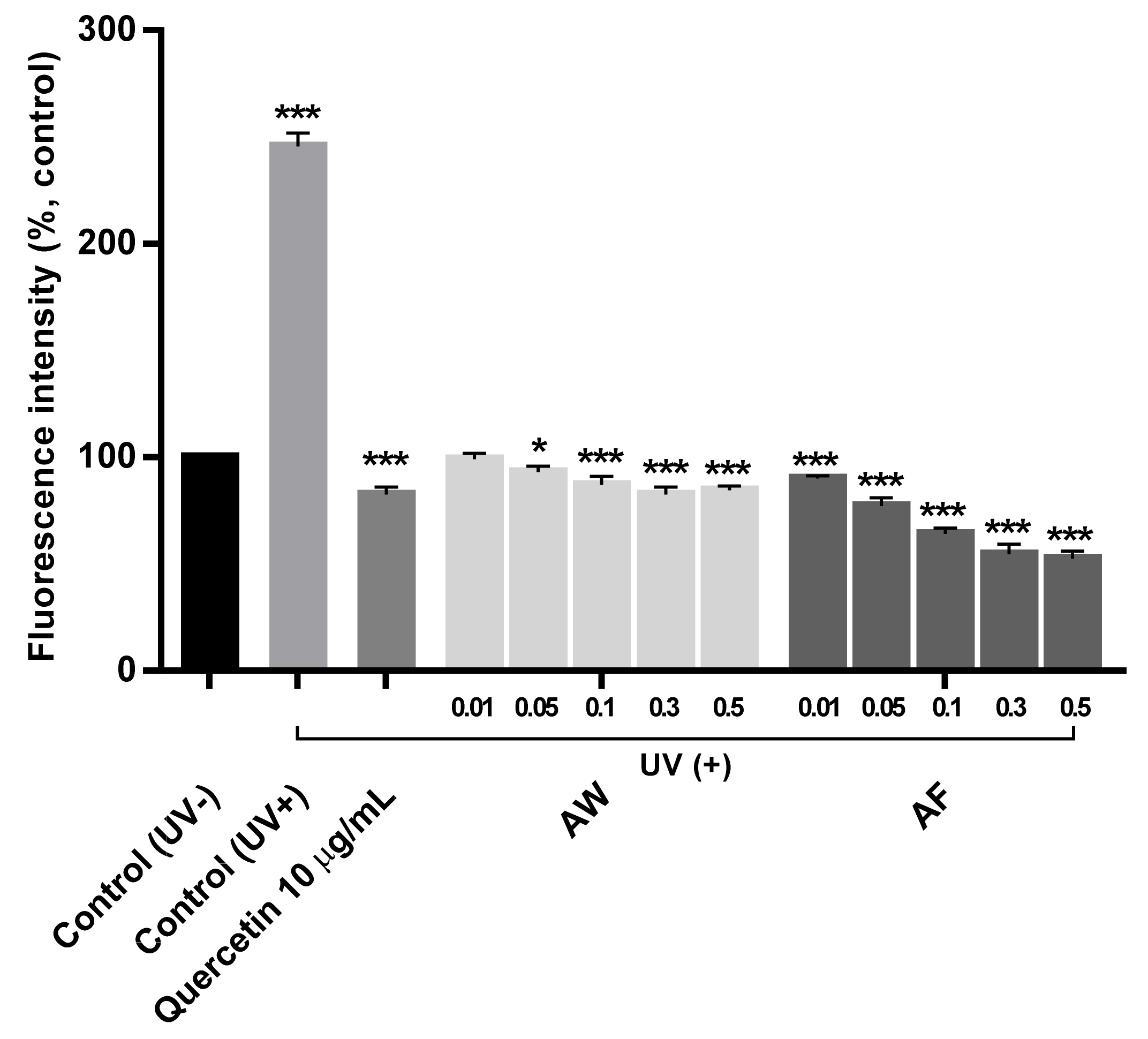
3.3. DPPH Free Radical Scavenging Effect and the Reduction of ROS Production by the Samples
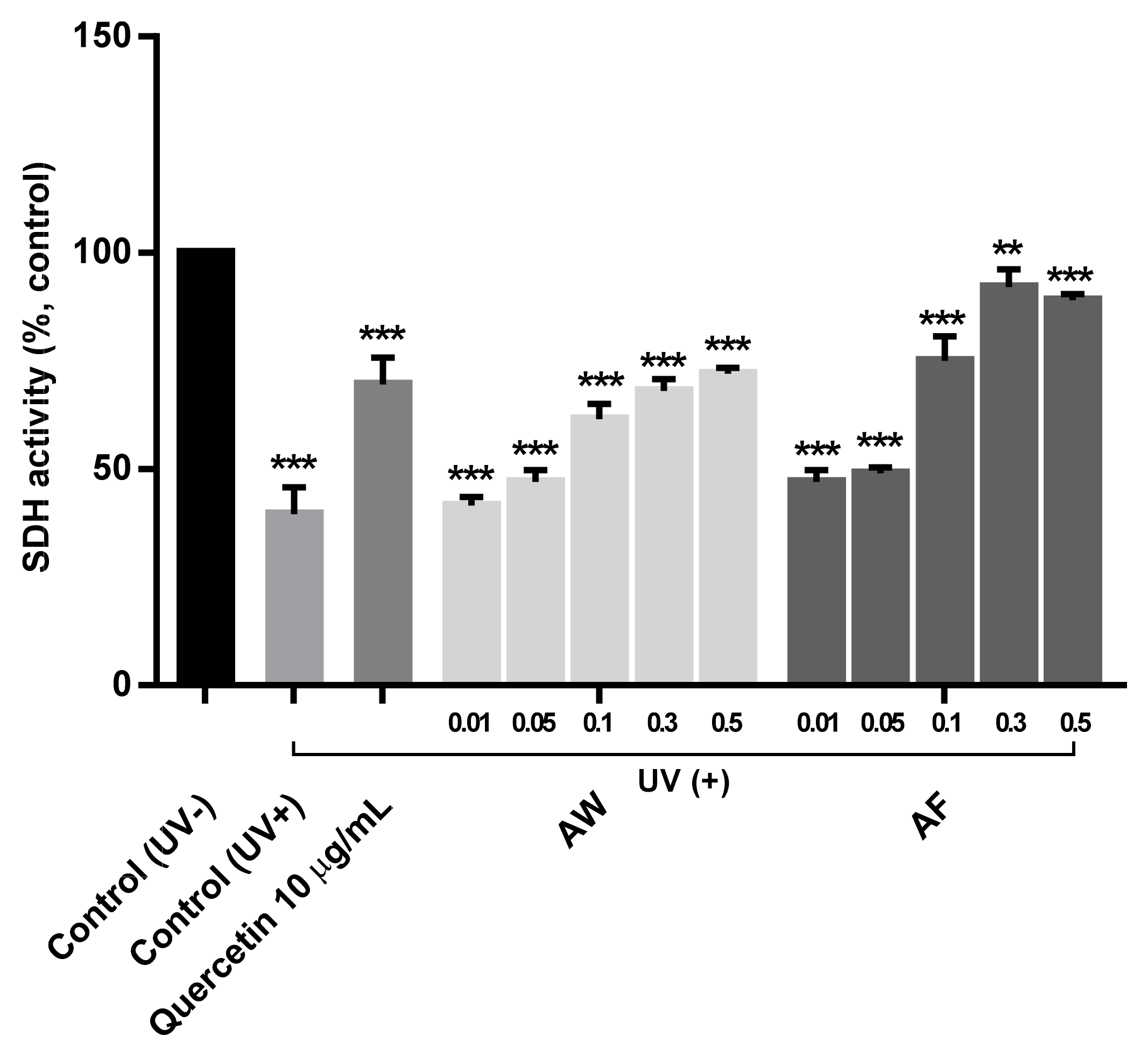
3.4. The Restoration of Mitochondrial Cytochrome II Activities in UVb-Irradiated Human Fibroblasts
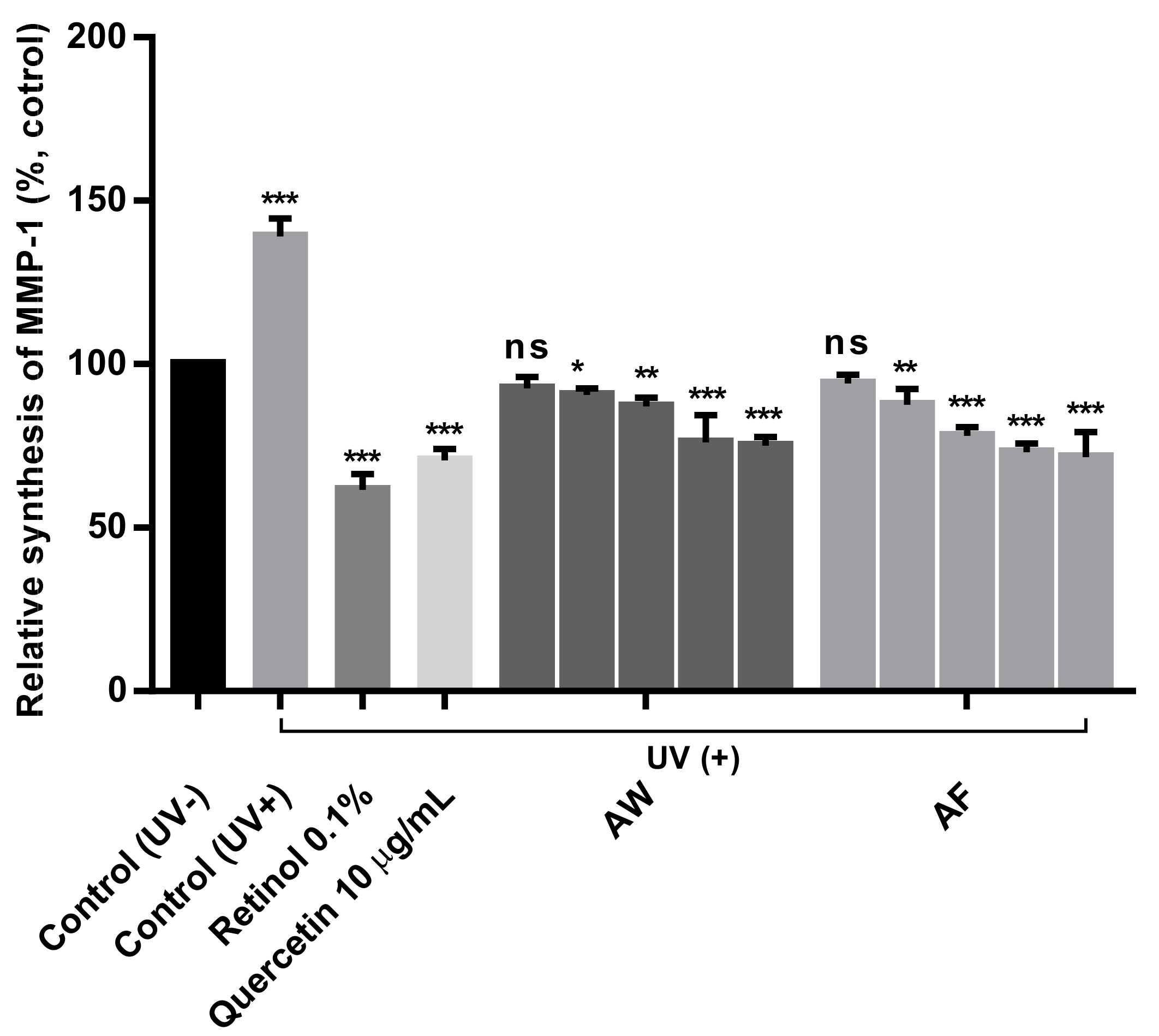
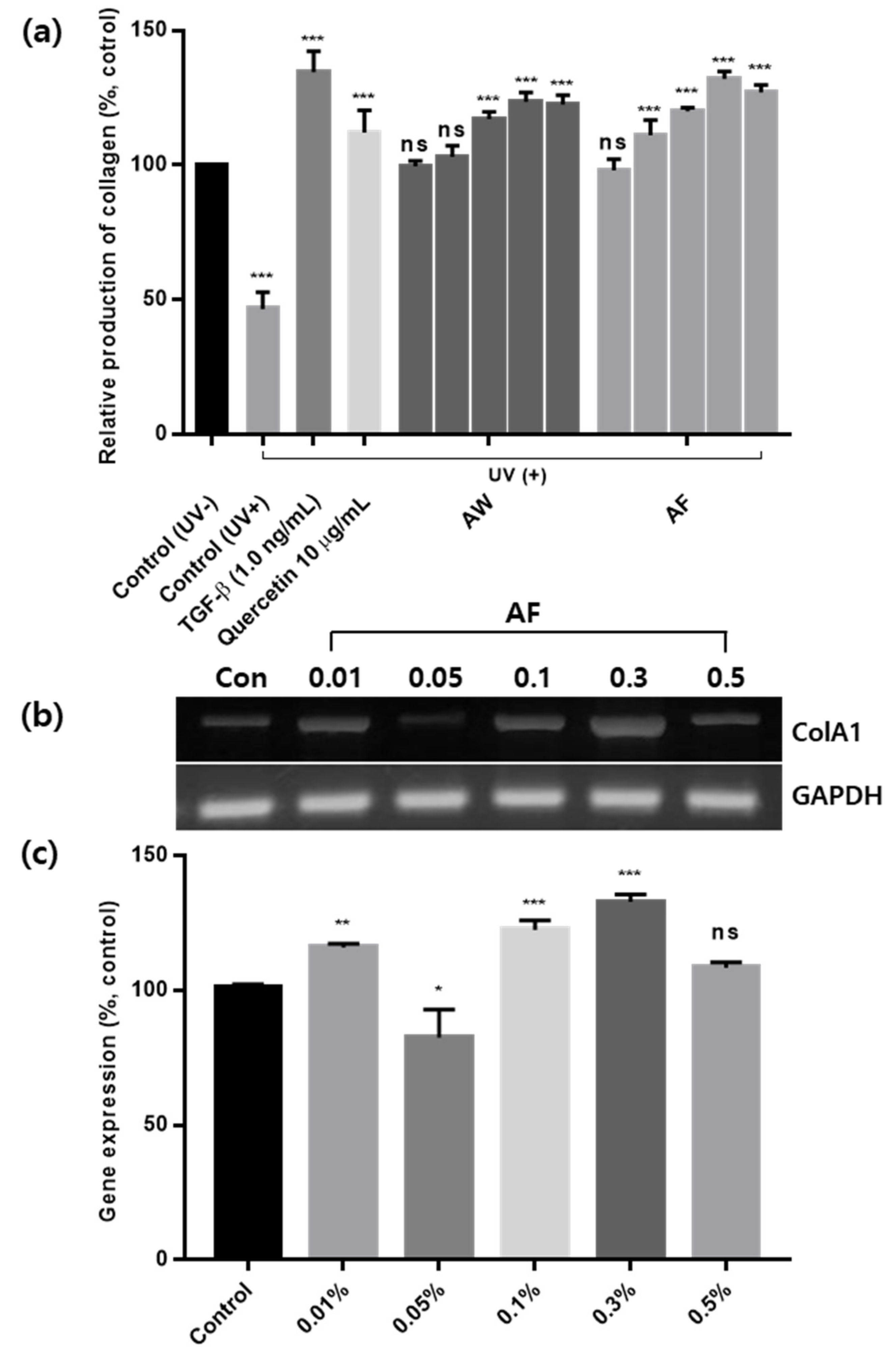
3.5. Skin Antiaging Effects of the Samples on UVb-Irradiated Human Fibroblasts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moon, H.J.; Lee, S.R.; Shim, S.N.; Jeong, S.H.; Stonik, V.A.; Rasskazov, V.A.; Zvyagintseva, T.; Lee, Y.H. Fucoidan inhibits UVB-induced MMP-1 expression in human skin fibroblasts. Biol. Pharm. Bull. 2008, 31, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.A.; Jung, M.; Lee, S.M.; Lee, T.H.; Kim, J. Protective effect of Disporum sessile D.Don extract against UVB-induced photoaging via suppressing MMP-1 expression and collagen degradation in human skin cells. J. Photochem. Photobiol. B Biol. 2014, 133, 73–79. [Google Scholar] [CrossRef]
- Jenkins, G. Molecular mechanisms of skin ageing. Mech. Ageing Dev. 2002, 123, 801–810. [Google Scholar] [CrossRef]
- Agnieszka, G.; Pedro, D.; Elżbieta, S. Proteins involved in the antioxidant and inflammatory response in rutin-treated human skin fibroblasts exposed to UVA or UVB irradiation. J. Dermatol. Sci. 2018, 90, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Piao, M.J.; Kim, K.C.; Zheng, J.; Yao, C.W.; Cha, J.W.; Boo, S.J. The ethyl acetate fraction of Sargassum muticum attenuates ultraviolet B radiation-induced apoptotic cell death via regulation of MAPK- and caspase-dependent signaling pathways in human HaCaT keratinocytes. Pharm. Biol. 2014, 52, 1110–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Choi, S.I.; Jung, T.D.; Cho, B.Y.; Lee, J.H.; Kim, S.H.; Yoon, S.A.; Ham, Y.M.; Yoon, W.J.; Cho, J.H.; et al. Anti-Photoaging Effect of Jeju Putgyul (Unripe Citrus) Extracts on Human Dermal Fibroblasts and Ultraviolet B-induced Hairless Mouse Skin. Int. J. Mol. Sci. 2017, 18, 2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadet, J.; Davies, K.J.A.; Medeiros, M.H.G.; Mascio, P.D.; Wagner, J.R. Formation and repair of oxidatively generated damage in cellular DNA. Free Radic. Biol. Med. 2017, 107, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Yusoff, A.A.M.; Khair, S.Z.N.M.; Radzak, S.M.A.; Idris, Z.; Lee, H.C. Prevalence of mitochondrial DNA common deletion in patients with gliomas and meningiomas: A first report from a Malaysian study group. J. Chin. Med. Assoc. 2020, 83, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 25, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and mitochondria: Activation of cytoprotective pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef] [PubMed]
- Ratree, M.; Nathorn, C.; Surachet, N.; Chuenjid, K. The efficacy of Aloe vera used for burn wound healing: A systematic review. Burns 2007, 33, 713–718. [Google Scholar] [CrossRef]
- Supreet, J.; Nirav, R.; Ravleen, N.; Jaideep, S.; Afshan, L.; Naveen, G.; Priyanka, A.; Swati, P. Antibacterial effect of Aloe vera gel against oral pathogens: An in-vitro study. J. Clin. Diagn. Res. 2016, 10, 41–44. [Google Scholar] [CrossRef]
- Langmead, L.; Makins, R.J.; Rampton, D.S. Anti-inflammatory effect of Aloe vera gel in human colorectal mucosa in vitro. Aliment. Pharmacol. Ther. 2004, 19, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Chandegara, V.K.; Varshney, A.K. Aloe vera L. processing and products: A review. Int. J. Med. Aromat. Plants 2013, 3, 492–506. [Google Scholar]
- Chang, X.L.; Chen, B.Y.; Feng, Y.M. Water-soluble polysaccharides isolated from skin juice, gel juice and flower of Aloe vera miller. J. Taiwan Inst. Chem. Eng. 2011, 42, 197–203. [Google Scholar] [CrossRef]
- Minjares-Fuentes, R.; Femenia, A.; Comas-Serra, F.; Rodríguez-González, V.M. Compositional and structural features of the main bioactive polysaccharides present in the Aloe vera plant. J. AOAC Int. 2018, 101, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Hidehiko, B.; Kan, S.; Takeshi, C.; Takaaki, K.; Ikuko, T.; Sachiyo, Y.; Sayaka, O.; Hiroshi, K.; Shigeru, S. Antidiabetic effects of dietary administration of Aloe arborescens Miller components on multiple low-dose streptozotocin-induced diabetes in mice: Investigation on hypoglycemic action and systemic absorption dynamics of aloe components. J. Ethnopharmacol. 2006, 103, 468–477. [Google Scholar] [CrossRef]
- Kong, B.M.; Park, B.M.; Min, M.J.; Kim, S.Y.; Yang, D.C. Physico-chemicial characteristics of white, fermented and red ginseng extracts. J. Ginseng Res. 2008, 32, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Perdigón, G.; Fuller, R.; Raya, R. Lactic acid bacteria and their effect on the immune system. Curr. Issues Intest. Microbiol. 2001, 12, 27–42. [Google Scholar]
- Yang, M.C.; Jeong, S.W.; Ma, J.Y. Analysis of constituents in Sipjundaebo-tangs fermented by lactic acid bacteria. Microbiol. Biotechnol. Lett. 2011, 39, 350–358. [Google Scholar]
- Lee, H.Y. Improvement of skin barrier dysfunction by Scutellaria baicalensis GEOGI extracts through lactic acid fermentation. J. Cosmet. Dermatol. 2019, 18, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Lee, D.E.; Park, S.D.; Kim, Y.T.; Jeong, J.W.; Chung, D.K.; Lee, J.H. Oral administration of Lactobacillus plantarum HY7714 protects hairless mouse against ultraviolet B-induced photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef]
- Lee, X.M.; Lee, H.A.; Kwon, M.; Park, E.S.; Park, K.Y. Probiotic Effects of Lactobacillus plantarum strains isolated from Kimchi. J. Korean Soc. Food Sci. Nutr. 2016, 45, 1717–1724. [Google Scholar] [CrossRef]
- Hu, Z.; Wang, X.; Yang, L.; Tan, X.; Bao, T. Determination of quercetin in Yi Gan Qing capsule by HPLC. J. Anhui Agric. Sci. 2008, 36, 9347–9348. [Google Scholar]
- Park, H.J.; Cho, J.H.; Hong, S.H.; Kim, D.H.; Jung, H.Y.; Kang, I.K.; Cho, Y.J. Whitening and anti-wrinkle activities of ferulic acid isolated from Tetragonia tetragonioides in B16F10 melanoma and CCD-986sk fibroblast cells. J. Nat. Med. 2018, 72, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Frezzini, M.A.; Castellani, F.; Francesco, N.; Ristorini, M.; Canepari, S. Application of DPPH assay for assessment of particulate matter reducing properties. Atmosphere 2019, 10, 816. [Google Scholar] [CrossRef] [Green Version]
- Kobashigawa, L.C.; Xu, Y.C.; Padbury, J.F.; Tseng, Y.T.; Yano, N. Metformin protects cardiomyocyte from doxorubicin induced cytotoxicity through an AMP-activated protein kinase dependent signaling pathway: An in vitro study. PLoS ONE 2014, 15, e104888–e104899. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Wang, X.; Zhao, Y.; Yang, Q.; Ding, H.; Dong, Q.; Chen, X.; Cui, M. Ketogenic diet improves brain ischemic tolerance and inhibits NLRP3 inflammasome activation by preventing Drp1-mediated mitochondrial fission and endoplasmic reticulum stress. Front. Mol. Neurosci. 2018, 20, 29662437–29662452. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.Y.; Lee, K.H. Effects of Draconis resina on the collagenase activities and the procollagen synthesis in Hs68 human fibroblasts, and tyrosinase activity. Korea J. Herbol. 2015, 6, 1–6. [Google Scholar] [CrossRef]
- Lee, K.E.; Bharadwaj, S.; Yadava, U.; Kang, S.G. Computational and in vitro investigation of (-)-Epicatechin and Proanthocyanidin B2 as inhibitors of human matrix metalloproteinase 1. Biomolecules 2020, 10, 1379. [Google Scholar] [CrossRef]
- Chen, B.; Li, Y.; Yan, N.; Chen, G.; Qian, W.; Jiang, H.L.; Ji, C.; Bi, Z.G. Astragaloside IV controls collagen reduction in photoaging skin by improving transforming growth factor-β/Smad signaling suppression and inhibiting matrix metalloproteinase-1. Mol. Med. Rep. 2015, 11, 3344–3348. [Google Scholar] [CrossRef] [Green Version]
- Sultana, B.; Anwar, F. Flavonols (kaempeferol, quercetin, myricetin) contents of selected fruits, vegetables and medicinal plants. Food Chem. 2008, 108, 879–884. [Google Scholar] [CrossRef]
- Silva, A.M.; Trevisan, G.; ZeniKlafke, J.; Rossato, M.F.; Isabel, C.; Walker, B.; Oliveira, S.M.; Silva, C.R.; Boligon, A.; Flores, F.C.; et al. Antinociceptive and anti-inflammatory effects of Aloe saponaria Haw on thermal injury in rats. J. Ethnopharmacol. 2013, 146, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Q.; Wang, H.; Guo, X.; Abbasi, A.M.; Wang, T.; Li, T.; Fu, X.; Li, J.; Liu, R.H. Comparison of phytochemical profiles, antioxidant and cellular antioxidant activities of seven cultivars of Aloe. Int. J. Food Sci. Technol. 2016, 51, 1489–1494. [Google Scholar] [CrossRef]
- López, A.; De Tangil, M.S.; Vega-Orellana, O.; Ramírez, A.S.; Rico, M. Phenolic constituents, antioxidant and preliminary antimycoplasmic activities of leaf skin and flowers of Aloe vera (L.) Burm. f. (syn. A. barbadensis Mill.) from the canary islands (Spain). Molecules 2013, 18, 4942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuttila, A.M.; Puupponen-Pimia, R.; Aarni, M.; Oksman-Caldentey, K.M. Comparison of antioxidant activities of onion and garlic extracts by inhibition of lipid peroxidation and radical scavenging activity. Food Chem. 2003, 81, 485–493. [Google Scholar] [CrossRef]
- Polerà, N.; Badolato, M.; Perri, F.; Carullo, G.; Aiello, F. Quercetin and its natural sources in wound healing management. Curr. Med. Chem. 2019, 26, 5825–5848. [Google Scholar] [CrossRef]
- Shah, Z.A.; Li, R.; Ahmad, A.S.; Kensler, T.W.; Yamamoto, M.; Biswal, S.; Doré, S. The flavanol (-)-epicatechin prevents stroke damage through the Nrf2/HO1 pathway. J. Cereb. Blood Flow Metab. 2010, 30, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.A.; Wei, Y.; Guo, M. Iron-binding and anti-Fenton properties of baicalein and baicalin. J. Inorg. Biochem. 2009, 103, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Arredondo, F.; Echeverry, C.; Abin-Carriquiry, J.; Blasina, F.; Antúnez, K.; Jones, D.; Go, Y.; Liang, Y.; Dajas, F. After cellular internalization, quercetin causes Nrf2 nuclear translocation, increases glutathione levels, and prevents neuronal death against an oxidative insult. Free Radic. Biol. Med. 2010, 49, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Seyed, T.A.; Nabavi, F. Quercetin and the mitochondria: A mechanistic view. Biotechnol. Adv. 2016, 34, 532–549. [Google Scholar] [CrossRef]
- Hong, M.S.; Yoon, Y.M.; An, S.K.; An, I.S.; Byun, B.H. Protective effects of the natural ingredient quercetin in human dermal fibroblasts. Asian J. Beauty Cosmetol. 2012, 10, 571–579. [Google Scholar]
- Muthukumaran, P.; Divya, R.; Indhumathi, E.; Keerthika, C. Total phenolic and flavonoid content of membrane processed Aloe vera extract: A comparative study. Int. Food Res. J. 2018, 25, 1450–1456. [Google Scholar]
- Kim, S.H.; Nam, G.W.; Gang, B.Y.; Lee, H.K.; Moon, S.J.; Chang, I.S. The effect of kaempferol and quercetin on hyaluronan-synthesis in human keratinocytes (HaCaT). J. Soc. Cosmet. Sci. Korea 2005, 37, 97–102. [Google Scholar]
- Li, Y.F.; Ouyang, S.H.; Tu, L.F.; Wang, X.; Yuan, W.L.; Wang, G.E.; Wu, Y.P.; Duan, W.J.; Yu, H.M.; Fang, Z.Z.; et al. Caffeine protects skin from oxidative stress-induced senescence through the activation of autophagy. Theranostics 2018, 8, 5713–5730. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.R.; Nasir, B.; Haq, I.; Kim, S.J. Oxidative stress, consequences and ROS mediated cellular signaling in rheumatoid arthritis. Chem. Biol. 2018, 281, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of mitochondrial DNA damage in ROS-mediated pathogenesis of age-related macular degeneration (AMD). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef] [Green Version]
- Vera, A.V. Production of reactive oxygen species in brain mitochondria: Contribution by electron transport chain and non–electron transport chain sources. Antioxid. Redox Signal 2005, 7, 1140–1149. [Google Scholar] [CrossRef]
- Bowman, A.; Birch-Machin, M.A. Age-dependent decrease of mitochondrial complex II activity in human skin fibroblasts. J. Investig. Dermatol. 2016, 136, 912–919. [Google Scholar] [CrossRef]
- Dudylina, A.; Ivanova, M.; Shumaev, K.; Ruuge, E. Superoxide formation in cardiac mitochondria and effect of phenolic antioxidants. Cell Biochem. Biophys. 2019, 77, 99–107. [Google Scholar] [CrossRef]
- Varani, J.; Warner, R.L.; Gharaee-Kermani, M.; Phan, S.H.; Kang, S.; Chung, J.H.; Wang, Z.Q.; Datta, S.C.; Fisher, G.j.; Voorhees, J.J. Vitamin A antagonizes decreased cell growth and elevated collagen-degrading matrix metalloproteinases and stimulates collagen accumulation in naturally aged human skin. J. Investig. Dermatol. 2000, 114, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Liao, P.L.; Li, C.H.; Chang, C.Y.; Lu, S.R.; Lin, C.H.; Tse, L.S.; Cheng, Y.W. Anti-ageing effects of alpha-naphthoflavone on normal and UVB-irradiated human skin fibroblasts. Exp. Dermatol. 2012, 21, 535–561. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.D.; Kaiser, P.; Satti, N.K.; Zargar, A.M.; Vishwakarma, R.A.; Tasduq, S.A. Effect of Emblica officinalis (fruit) against UVB-induced photo-aging in humanskin fibroblasts. J. Ethnopharmacol. 2010, 132, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Wittenauer, J.; Mackle, S.; Sussmann, D.; Schweiggert-Weisz, U.; Carle, R. Inhibitory effects of polyphenols from grape pomace extract on collagenase and elastase activity. Fitoterapia 2015, 101, 179–187. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Range (μg/mL) | Equation for Regression Line | R2 | SD Intercept | SD Slope |
|---|---|---|---|---|---|
| AW | 298.5–3825.1 | Y = 1.534X + 59.0595 | 0.9997 | 38.35 | 0.19 |
| AF | 273.4–3489.2 | Y = 1.207X + 35.8621 | 0.9996 | 36.41 | 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Choi, W.; Ro, H.; Kim, G.; Lee, H. Skin Antiaging Effects of the Fermented Outer Layers of Leaf Skin of Aloe barbadensis Miller Associated with the Enhancement of Mitochondrial Activities of UVb-Irradiated Human Skin Fibroblasts. Appl. Sci. 2021, 11, 5660. https://doi.org/10.3390/app11125660
Lee H, Choi W, Ro H, Kim G, Lee H. Skin Antiaging Effects of the Fermented Outer Layers of Leaf Skin of Aloe barbadensis Miller Associated with the Enhancement of Mitochondrial Activities of UVb-Irradiated Human Skin Fibroblasts. Applied Sciences. 2021; 11(12):5660. https://doi.org/10.3390/app11125660
Chicago/Turabian StyleLee, Hyeonwoo, Woonyong Choi, Hyangseon Ro, Gyurae Kim, and Hyeonyong Lee. 2021. "Skin Antiaging Effects of the Fermented Outer Layers of Leaf Skin of Aloe barbadensis Miller Associated with the Enhancement of Mitochondrial Activities of UVb-Irradiated Human Skin Fibroblasts" Applied Sciences 11, no. 12: 5660. https://doi.org/10.3390/app11125660