
Influence of Ecological Restoration on Mercury Mobility and Microbial Activities on Former Guyanese Mining Sites

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
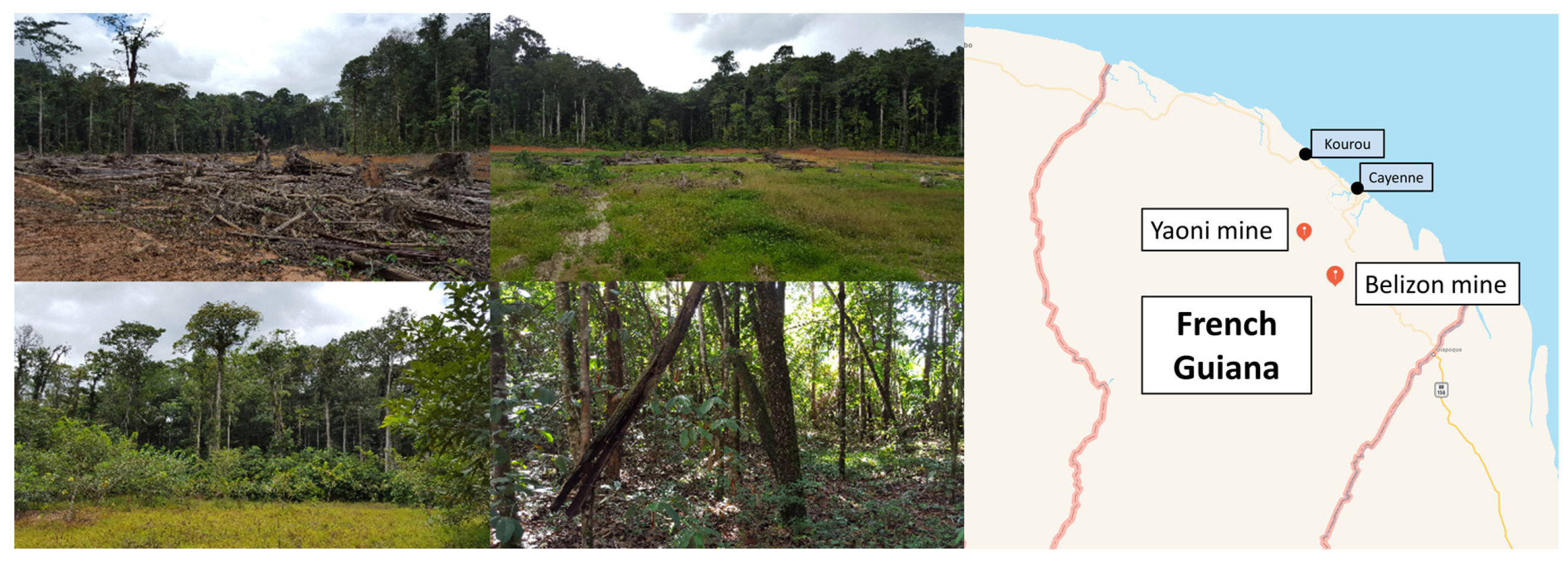
2.1. Site Description and Soil Sampling
2.2. Soil Samples Characterization
2.3. Soil Microbial Biomass and Enzyme Assays
2.4. Mercury Analysis
2.4.1. Total Mercury Content
2.4.2. Environmental Speciation of Mercury
- Extraction of “Highly mobile” mercury species
- Extraction of toxic, semi-mobile and non-mobile mercury
2.5. Statistical Analysis
2.6. Quality Assurance and Control
3. Results
3.1. Soil Physical and Chemical Properties of the 6 Sites
3.2. Soil Microbial Biomass and Enzyme Activities
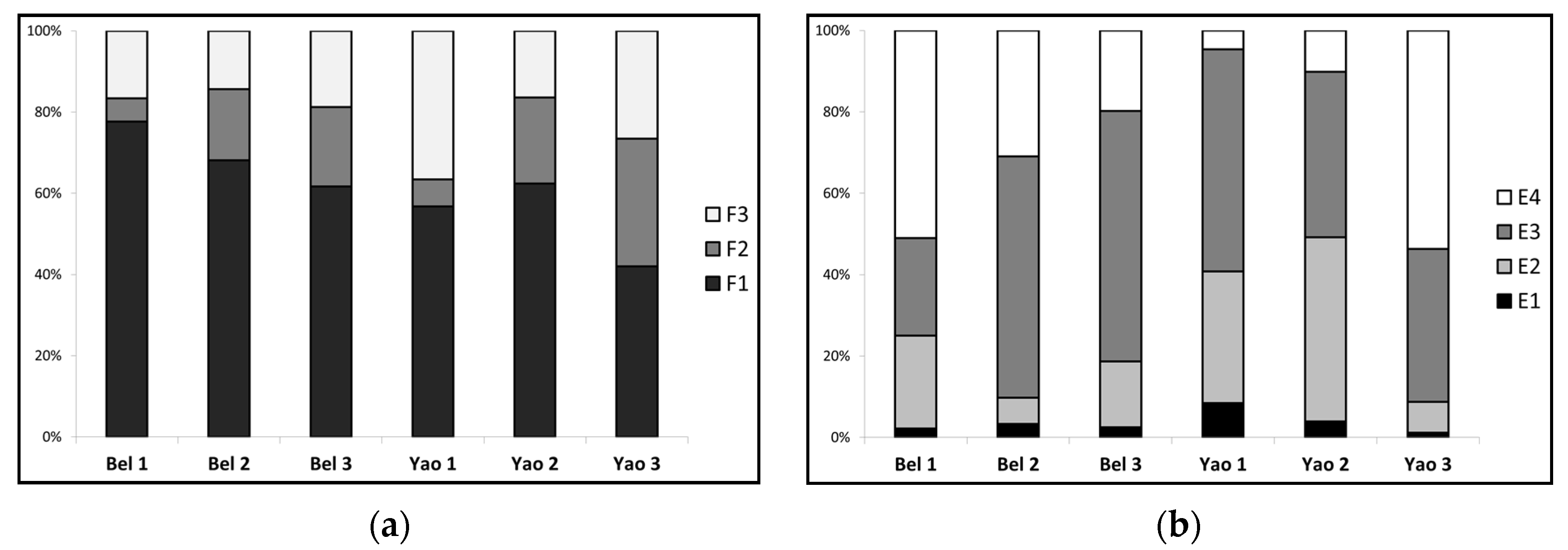
3.3. Mercury Environmental Speciation
- Total mercury
- Total highly mobile and mobile mercury species
- Total semi-mobile mercury species
- Total non-mobile mercury species
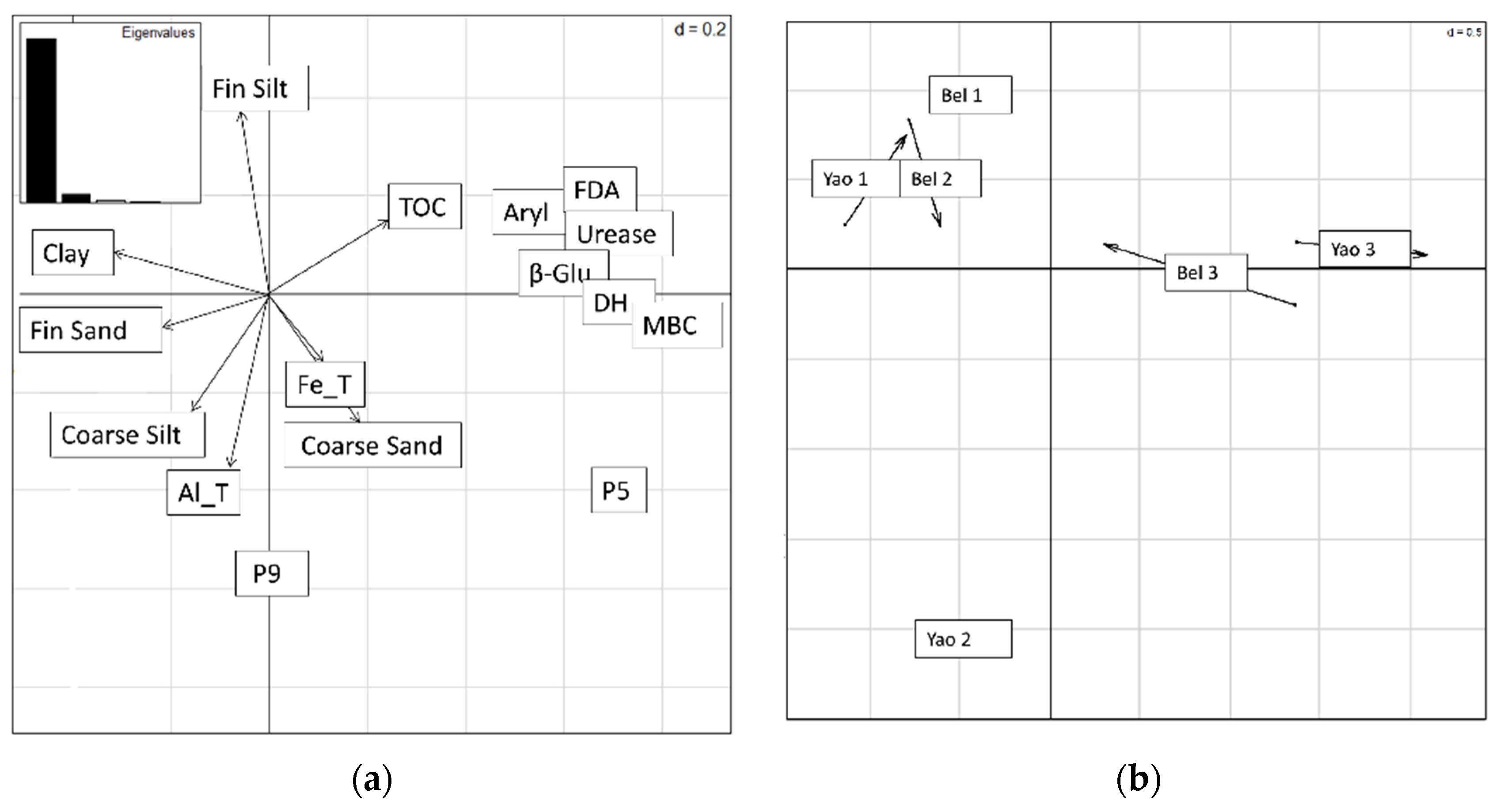
3.4. Co-Inertia between Soil Parameters and Microbial Activities Dataset
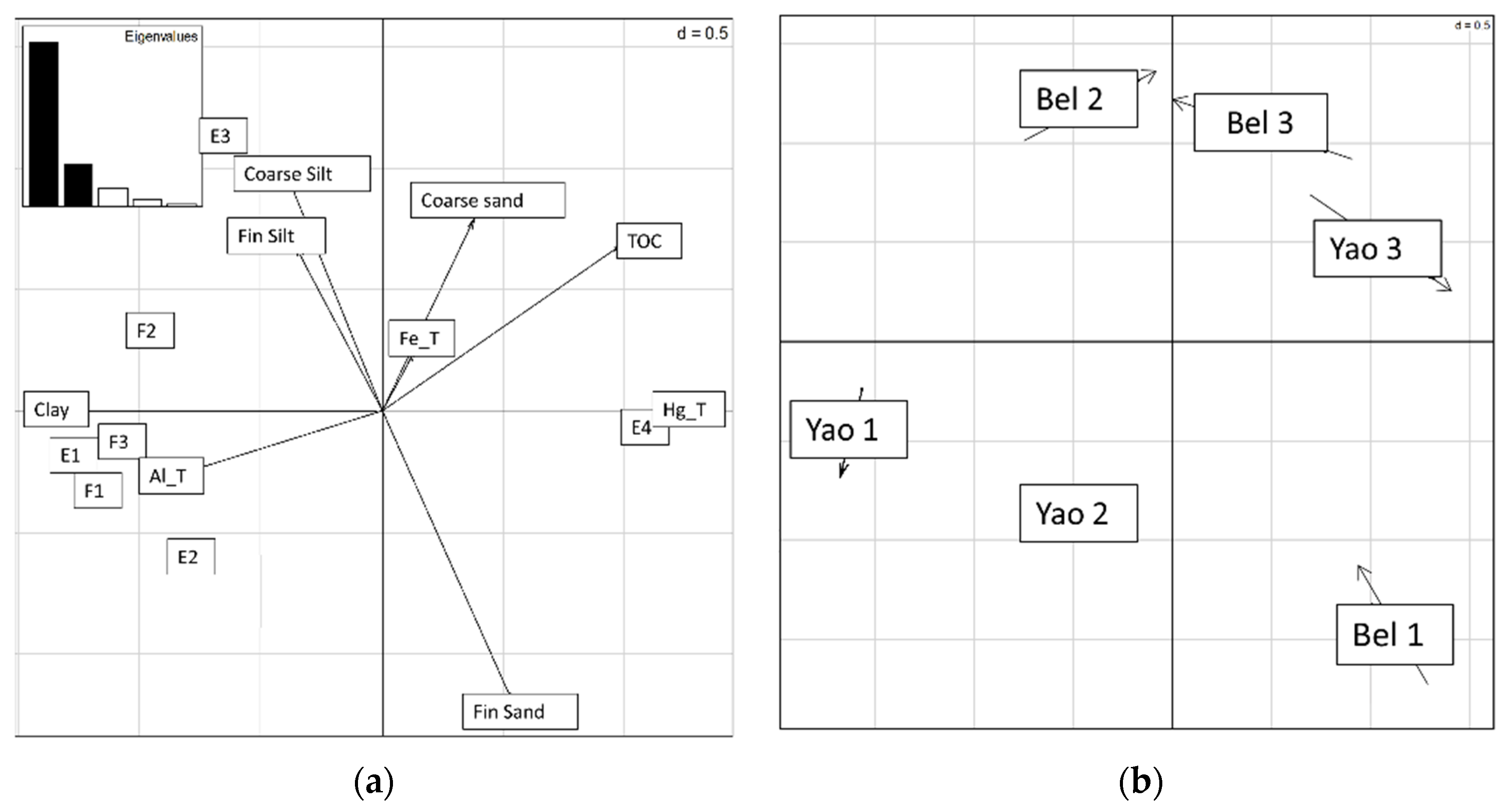
3.5. Co-Inertia between Soil Parameters and Environmental Mercury Speciation
4. Discussion
4.1. Positive Effect of Ecological Restoration on Mercury Mobility
4.2. Effect of Soil and Microbial Properties on Environmental Speciation of Mercury
4.3. Positive Effect of Ecological Restoration on Microbial Activities
4.4. Footprint of Rehabilitation on Soil Functionning and Mercury Mobility
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Van Der Werf, G.R.; Morton, D.C.; Defries, R.S.; Olivier, J.G.J.; Kasibhatla, P.S.; Jackson, R.B.; Collatz, G.J.; Randerson, J.T. CO2 emissions from forest loss. Nat. Geosci. 2009. [Google Scholar] [CrossRef]
- Achard, F.; Beuchle, R.; Mayaux, P.; Stibig, H.J.; Bodart, C.; Brink, A.; Carboni, S.; Desclée, B.; Donnay, F.; Eva, H.D.; et al. Determination of tropical deforestation rates and related carbon losses from 1990 to 2010. Glob. Chang. Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Song, X.P.; Hansen, M.C.; Stehman, S.V.; Potapov, P.V.; Tyukavina, A.; Vermote, E.F.; Townshend, J.R. Global land change from 1982 to 2016. Nature 2018. [Google Scholar] [CrossRef]
- Veiga, M.M.; Maxson, P.A.; Hylander, L.D. Origin and consumption of mercury in small-scale gold mining. J. Clean. Prod. 2006, 14, 436–447. [Google Scholar] [CrossRef]
- Wong, M. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 2003, 50, 775–780. [Google Scholar] [CrossRef]
- Grimaldi, M.; Guédron, S.; Grimaldi, C. Impact of gold mining on mercury contamination and soil degradation in Amazonian ecosystems of French Guiana. In Land-Use Change Impacts on Soil Processes: Tropical and Savannah Ecosystems; CABI International: Wallingford, CT, USA, 2015; pp. 95–107. ISBN 9781780642109. [Google Scholar]
- Telmer, K.; Costa, M.; Angélica, R.S.; Araujo, E.S.; Maurice, Y. The source and fate of sediment and mercury in the Tapajós River, Pará, Brazilian Amazon: Ground- and space-based evidence. J. Environ. Manag. 2006, 81, 101–113. [Google Scholar] [CrossRef]
- Roulet, M.; Lucotte, M.; Farella, N.; Serique, G.; Coelho, H.; Passos, C.S.; Da Silva, E.D.; De Andrade, P.S.; Mergler, D.; Guimarães, J.R.; et al. Effects of Recent Human Colonization on the Presence of Mercury in Amazonian Ecosystems. Water Air. Soil Pollut. 1999, 112, 297–313. [Google Scholar] [CrossRef]
- Wasserman, J.C.; Fundação, S.S.H.; Cruz, O.; Wasserman, M. Biogeochemistry of Mercury in the Amazonian Environment Mercury Exposure in Yanomami Indigenous Communities in the sate of Roraima View project Fate of nanoparticles in the environment View project. Ambio J. Hum. Environ. 2003, 32, 336–342. [Google Scholar] [CrossRef]
- Lacerda, L.D.; De Souza, M.; Ribeiro, M.G. The effects of land use change on mercury distribution in soils of Alta Floresta, Southern Amazon. Environ. Pollut. 2004, 129, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Guedron, S. Impact de l ’Exploitation Minière en Guyane Française Sur Les Flux de Mercure Vers Les Écosystèmes Aquatiques. Ph.D. Thesis, Université Joseph Fourier—Grenoble I, Grenoble, France, 14 March 2008. [Google Scholar]
- Alloway, B. Heavy Metals in Soils:Trace Metals and Metalloids in Soils and Their Bioavailability; Springer: Dordrecht, The Netherlands, 2012; ISBN 9789400744691. [Google Scholar]
- Veiga, M.M.; Baker, R.F. Protocols for Environmental and Health Assessment of Mercury Released by Artisanal and Small-Scale Gold Miners; United Nations: New York, NY, USA, 2004; ISBN 9211064295. [Google Scholar]
- Han, Y.; Kingston, H.M.; Boylan, H.M.; Rahman, G.M.M.; Shah, S.; Richter, R.C.; Link, D.D.; Bhandari, S. Speciation of mercury in soil and sediment by selective solvent and acid extraction. Anal. Bioanal. Chem. 2003, 375, 428–436. [Google Scholar] [CrossRef]
- Navarro, A. Review of characteristics of mercury speciation and mobility from areas of mercury mining in semi-arid environments. Rev. Environ. Sci. Biotechnol. 2008, 7, 287–306. [Google Scholar] [CrossRef]
- Basavaraja, P.K.; Sharma, S.D.; Dhananjaya, B.N.; Badrinath, M.S. Acacia nilotica: A tree species for amelioration of sodic soils in Central dry zone of Karnataka, India. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 73–76. [Google Scholar]
- Balland-Bolou-Bi, C.; Turc, B.; Alphonse, V.; Bousserrhine, N. Impact of microbial communities from tropical soils on the mobilization of trace metals during dissolution of cinnabar ore. J. Environ. Sci. 2017, 56, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, C. La réhabilitation des mines et carrières à ciel ouvert: Restauration des sites miniers. Bois Forêts Trop. 2002, 272, 5–19. [Google Scholar]
- Jaffré, T.; McCoy, S.; Rigault, F.; Dagostini, G. Quelle méthode de végétalisation pour la réhabilitation des anciens sites miniers de Nouvelle-Calédonie. In Écologie des milieux sur roches ultramafiques sur sols métallifères actes la deuxième conférence Int. sur l’écologie des milieux serpentiniques; ORSTOM: Noumea, France, 1997; pp. 285–288. [Google Scholar]
- Loubry, D. Livret Technique Pour la Conduite de la Revégétalisation sur les Surfaces Minières Alluvionanaires de Guyane; Unpublished Technical Report; IRD: Cayenne, Guyane, 2002; 51p. [Google Scholar]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Couic, E.; Grimaldi, M.; Alphonse, V.; Balland-Bolou-Bi, C.; Livet, A.; Giusti-Miller, S.; Sarrazin, M.; Bousserrhine, N. Mercury behaviour and C, N, and P biogeochemical cycles during ecological restoration processes of old mining sites in French Guiana. Environ. Sci. Process. Impacts 2018, 20. [Google Scholar] [CrossRef]
- Guedron, S.; Grangeon, S.; Lanson, B.; Grimaldi, M. Mercury speciation in a tropical soil association; Consequence of gold mining on Hg distribution in French Guiana. Geoderma 2009, 153, 331–346. [Google Scholar] [CrossRef] [Green Version]
- Schimann, H.; Petit-Jean, C.; Guitet, S.; Reis, T.; Domenach, A.M.; Roggy, J.C. Microbial bioindicators of soil functioning after disturbance: The case of gold mining in tropical rainforests of French Guiana. Ecol. Indic. 2012, 20, 34–41. [Google Scholar] [CrossRef]
- Leermakers, M.; Baeyens, W.; Quevauviller, P.; Horvat, M. Mercury in environmental samples: Speciation, artifacts and validation. TrAC Trends Anal. Chem. 2005, 24, 383–393. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Badiane, N.N.Y.; Chotte, J.L.; Pate, E.; Masse, D.; Rouland, C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol. 2001, 18, 229–238. [Google Scholar] [CrossRef]
- Mora, P.; Miambi, E.; Jiménez, J.J.; Decaëns, T.; Rouland, C. Functional complement of biogenic structures produced by earthworms, termites and ants in the neotropical savannas. Soil Biol. Biochem. 2005, 37, 1043–1048. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Assay of urease activity in soils. Soil Biol. Biochem. 1972, 4, 479–487. [Google Scholar] [CrossRef]
- Klein, D.A.; Loh, T.C.; Goulding, R.L. A rapid procedure to evaluate the dehydrogenase activity of soils low in organic matter. Soil Biol. Biochem. 1971, 3, 385–387. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Green, V.S.; Stott, D.E.; Diack, M. Assay for fluorescein diacetate hydrolytic activity: Optimization for soil samples. Soil Biol. Biochem. 2006, 38, 693–701. [Google Scholar] [CrossRef]
- Rasmussen, L.D.; Sørensen, S.J.; Turner, R.R.; Barkay, T. Application of a mer-lux biosensor for estimating bioavailable mercury in soil. Soil Biol. Biochem. 2000, 32, 639–646. [Google Scholar] [CrossRef]
- Silveira, M.L.; Alleoni, L.R.F.; O’Connor, G.A.; Chang, A.C. Heavy metal sequential extraction methods—A modification for tropical soils. Chemosphere 2006, 64, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Harris-Hellal, J.; Vallaeys, T.; Garnier-Zarli, E.; Bousserrhine, N. Effects of mercury on soil microbial communities in tropical soils of French Guyana. Appl. Soil Ecol. 2009, 41, 59–68. [Google Scholar] [CrossRef]
- Harris-Hellal, J.; Grimaldi, M.; Garnier-Zarli, E.; Bousserrhine, N. Mercury mobilization by chemical and microbial iron oxide reduction in soils of French Guyana. Biogeochemistry 2011, 103, 223–234. [Google Scholar] [CrossRef]
- Da Silva, E.; Nahmani, J.; Lapied, E.; Alphonse, V.; Garnier-Zarli, E.; Bousserrhine, N. Toxicity of mercury to the earthworm Pontoscolex corethrurus in a tropical soil of French Guiana. Appl. Soil Ecol. 2016, 104, 79–84. [Google Scholar] [CrossRef]
- Shukla, G.; Varma, A. Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 9783642142246. [Google Scholar]
- Taylor, J.P.; Wilson, B.; Mills, M.S.; Burns, R.G. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biol. Biochem. 2002, 34, 387–401. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Cruz, L.; Sotomayor-Ramírez, D.; Pérez-Alegría, L. Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl. Soil Ecol. 2007. [Google Scholar] [CrossRef]
- Huang, Q.; Liang, W.; Cai, P. Adsorption, desorption and activities of acid phosphatase on various colloidal particles from an Ultisol. Colloids Surf. B Biointerfaces 2005. [Google Scholar] [CrossRef]
- Safari Sinegani, A.A.; Emtiazi, G.; Shariatmadari, H. Sorption and immobilization of cellulase on silicate clay minerals. J. Colloid Interface Sci. 2005. [Google Scholar] [CrossRef]
- Caravaca, F.; Hernández, T.; García, C.; Roldán, A. Improvement of rhizosphere aggregate stability of afforested semiarid plant species subjected to mycorrhizal inoculation and compost addition. Geoderma 2002, 108, 133–144. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Adamson, B.W.; Anderson, S.H. Variations in soil aggregate stability and enzyme activities in a temperate agroforestry practice. Appl. Soil Ecol. 2008. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Harris, J.A. Restoration ecology: Repairing the earth’s ecosystems in the new millennium. Restor. Ecol. 2001, 9, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Viana, R.M.; Ferraz, J.B.S.; Neves, A.F.; Vieira, G.; Pereira, B.F.F. Soil quality indicators for different restoration stages on Amazon rainforest. Soil Tillage Res. 2014, 140, 1–7. [Google Scholar] [CrossRef]
- Elfstrand, S.; Båth, B.; Mårtensson, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Appl. Soil Ecol. 2007, 36, 70–82. [Google Scholar] [CrossRef]
- Butterfield, R.P. Early species selection for tropical reforestation: A consideration of stability. For. Ecol. Manag. 1996, 81, 161–168. [Google Scholar] [CrossRef]
- Norisada, M.; Hitsuma, G.; Kuroda, K.; Yamanoshita, T.; Masumori, M.; Tange, T.; Yagi, H.; Nuyim, T.; Sasaki, S.; Kojima, K. Acacia mangium, a nurse tree candidate for reforestation on degraded sandy soils in the Malay Peninsula. For. Sci. 2005, 51, 498–510. [Google Scholar]
- Fernández-Martínez, R.; Larios, R.; Gómez-Pinilla, I.; Gómez-Mancebo, B.; López-Andrés, S.; Loredo, J.; Ordóñez, A.; Rucandio, I. Mercury accumulation and speciation in plants and soils from abandoned cinnabar mines. Geoderma 2015, 253–254, 30–38. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Murciego, A.; Álvarez-Ayuso, E.; Regina, I.S.; Rodríguez-González, M.A. Mercury in soils and plants in an abandoned cinnabar mining area (SW Spain). J. Hazard. Mater. 2009, 168, 1319–1324. [Google Scholar] [CrossRef]
- Moreno-Jiménez, E.; Gamarra, R.; Carpena-Ruiz, R.O.; Millán, R.; Peñalosa, J.M.; Esteban, E. Mercury bioaccumulation and phytotoxicity in two wild plant species of Almadén area. Chemosphere 2006. [Google Scholar] [CrossRef]
- Santos-Francés, F.; García-Sánchez, A.; Alonso-Rojo, P.; Contreras, F.; Adams, M. Distribution and mobility of mercury in soils of a gold mining region, Cuyuni river basin, Venezuela. J. Environ. Manag. 2011. [Google Scholar] [CrossRef]
- Gavilán-García, I.; Santos-Santos, E.; Tovar-Gálvez, L.R.; Gavilán-García, A.; Suárez, S.; Olmos, J.; La, B.; Ticomán, L.; Delegación, G.A.; Madero, M.D.F. Mercury Speciation in Contaminated Soils from Old Mining Activities in Mexico Using a Chemical Selective Extraction. J. Mex. Chem. Soc. 2008, 52, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Larssen, T.; Vogt, R.D.; Feng, X. Identification of fractions of mercury in water, soil and sediment from a typical Hg mining area in Wanshan, Guizhou province, China. Appl. Geochem. 2010. [Google Scholar] [CrossRef]
- Do Valle, C.M.; Santana, G.P.; Augusti, R.; Egreja Filho, F.B.; Windm??ller, C.C. Speciation and quantification of mercury in Oxisol, Ultisol, and Spodosol from Amazon (Manaus, Brazil). Chemosphere 2005, 58, 779–792. [Google Scholar] [CrossRef]
- Roulet, M.; Lucotte, M. Geochemistry of mercury in pristine and flooded ferralitic soils of a tropical rain forest in French Guiana, South America. Water Air Soil Pollut. 1995, 80, 1079–1088. [Google Scholar] [CrossRef]
- Skyllberg, U.; Bloom, P.R.; Qian, J.; Lin, C.M.; Bleam, W.F. Complexation of mercury(II) in soil organic matter: EXAFS evidence for linear two-coordination with reduced sulfur groups. Environ. Sci. Technol. 2006, 40, 4174–4180. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. (Ed.) Heavy Metals in Soils; Environmental Pollution; Springer: Dordrecht, The Netherlands, 2013; Volume 22, ISBN 978-94-007-4469-1. [Google Scholar]
- Anderson, A. The Biochemistry of Mercury in the Environment. In Mercury in Soils; Elsevier/North-Holland Biomedical Press: Amsterdam, The Netherlands, 1979; pp. 79–112. ISBN 0444801103. [Google Scholar]
- Schuster, E. The behavior of mercury in the soil with special emphasis on complexation and adsorption processes—A review of the literature. Water Air Soil Pollut. 1991, 56, 667–680. [Google Scholar] [CrossRef]
- Campos, J.A.; Esbrí, J.M.; Madrid, M.M.; Naharro, R.; Peco, J.; García-Noguero, E.M.; Amorós, J.A.; Moreno, M.M.; Higueras, P. Does mercury presence in soils promote their microbial activity? The Almadenejos case (Almadén mercury mining district, Spain). Chemosphere 2018, 201, 799–806. [Google Scholar] [CrossRef]
- Jordan, D.; Kremer, R.J.; Bergfield, W.A.; Kim, K.Y.; Cacnio, V.N. Evaluation of microbial methods as potential indicators of soil quality in historical agricultural fields. Biol. Fertil. Soils 1995, 19, 297–302. [Google Scholar] [CrossRef]
- Chaussod, R. La qualité biologique des sols: Évaluation et implications. Etude Gest. des Sols 1996, 3, 261–278. [Google Scholar]
- Niemeyer, J.C.; Lolata, G.B.; de Carvalho, G.M.; Da Silva, E.M.; Sousa, J.P.; Nogueira, M.A. Microbial indicators of soil health as tools for ecological risk assessment of a metal contaminated site in Brazil. Appl. Soil Ecol. 2012. [Google Scholar] [CrossRef]
- Hu, Y.F.; Peng, J.J.; Yuan, S.; Shu, X.Y.; Jiang, S.L.; Pu, Q.; Ma, K.Y.; Yuan, C.M.; Chen, G.D.; Xiao, H.H. Influence of ecological restoration on vegetation and soil microbiological properties in Alpine-cold semi-humid desertified land. Ecol. Eng. 2016, 94, 88–94. [Google Scholar] [CrossRef]
- Yin, R.; Gu, C.; Feng, X.; Hurley, J.P.; Krabbenhoft, D.P.; Lepak, R.F.; Zhu, W.; Zheng, L.; Hu, T. Distribution and geochemical speciation of soil mercury in Wanshan Hg mine: Effects of cultivation. Geoderma 2016, 272, 32–38. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, M.; Xiao, R.; Cui, Y.; Yu, F. Changes in soil microbial biomass and community composition in coastal wetlands affected by restoration projects in a Chinese delta. Geoderma 2017, 289, 124–134. [Google Scholar] [CrossRef]
- Balland-Bolou-Bi, C.; Bolou-Bi, E.B.; Alphonse, V.; Giusti-Miller, S.; Jusselme, M.D.; Livet, A.; Grimaldi, M.; Bousserhine, N. Impact of microbial activity on the mobility of metallic elements (Fe, Al and Hg) in tropical soils. Geoderma 2019, 334, 146–154. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites Characteristics | Bel1 | Bel2 | Bel3 | Yao1 | Yao2 | Yao3 |
|---|---|---|---|---|---|---|
| TOC (g·kg−1) | 2.47 ± 0.22 a | 5.84 ± 0.34 b | 4.99 ± 0.37 b | 3.88 ± 0.03 ab | 3.51 ± 0.23 ab | 15.58 ± 1.4 c |
| C total (g·kg−1) | 3.52 ± 0.36 a | 9.54 ± 0.73 b | 7.01 ± 0.83 b | 6.01 ± 0.32 b | 4.29 ± 0.19 a | 22.88 ± 1.51 c |
| N total (g·kg−1) | 0.14 ± 0.02 a | 0.45 ± 0.01 b | 0.40 ± 0.04 b | 0.40 ± 0.02 b | 0.28 ± 0.03 ab | 1.81 ± 0.04 c |
| C/N | 25.14 | 21.20 | 17.52 | 15.02 | 15.32 | 12.64 |
| P total (g·kg−1) | 0.13 ± 0.01 a | 0.27 ± 0.07 b | 0.17 ± 0.02 ab | 0.21 ± 0.01 ab | 0.18 ±0.01 ab | 0.75 ± 0.03 c |
| pH | 5.3 ± 0.1 a | 4.66 ± 0.08 b | 5.00 ± 0.03 ab | 5.26 ± 0.2 a | 5.02 ± 0.03 ab | 4.66 ± 0.2 b |
| Clay% | 7.05 ± 2.7 a | 22.24 ± 2.4 b | 21.26 ± 1.5 b | 46.44 ± 1.1 c | 24.26 ± 0.5 b | 19.78 ± 1.4 b |
| Fin silt% | 18.62 ± 2.1 b | 44.47 ± 2.6 d | 17.68 ± 3.5 b | 33.92 ± 1.2 c | 10.10 ± 0.3 a | 27.7 ± 0.8 c |
| Coarse silt% | 2.39 ± 0.98 a | 21.39 ± 0.92 d | 7.10 ± 1.1 b | 7.57 ± 0.45 b | 10.40 ± 0.7 c | 7.8 ± 0.5 b |
| Fin sand% | 63.68 ± 1.4 e | 2.67 ± 1.1 a | 4.86 ± 0.6 a | 8.59 ± 0.49 b | 35.26 ± 0.7 d | 14.17 ± 1.35 c |
| Coarse sand% | 7.41 ± 0.33 b | 9.14 ± 0.8 b | 49 ± 5 d | 3.30 ± 0.48 a | 19.30 ± 0.5 c | 22.22 ± 1.17 c |
| Fe (g·kg−1) | 66.85 ± 6.2 b | 44.4 ± 2.8 b | 113.3 ± 15.2 a | 65.04 ± 10 ab | 75.7 ± 12.9 b | 71.4 ± 4.9 b |
| Al (g·kg−1) | 16.54 ± 2.12 a | 41.4 ± 2.2 a | 25.6 ± 2.15 b | 53.6 ± 15.8 b | 76.4 ± 4.8 b | 43.3 ± 10.2 b |
| Microbial Activities | Bel1 | Bel2 | Bel3 | Yao1 | Yao2 | Yao3 |
|---|---|---|---|---|---|---|
| MBC | 312.6 ± 10.0 c | 385.9 ± 21.5 c | 844.4 ± 14.1 b | 223.1 ± 10.8 d | 248.1 ± 13.6 d | 1102 ± 41 a |
| DH | 0.43 ± 0.08 d | 0.34 ± 0.02 d | 2.10 ± 0.19 b | 0.46 ± 0.07 d | 1.03 ± 0.12 c | 5.80 ± 0.44 a |
| Β-Glu | 1.60 ± 0.25 a | 1.46 ± 0.78 a | 5.54 ± 0.38 a | 1.38 ± 0.10 a | 3.48 ± 0.97 a | 18.6 ± 3.4 b |
| Urease | 0.46 ± 0.04 c | 0.70 ± 0.02 bc | 1.52 ± 0.06 b | 0.14 ± 0.01 d | 0.29 ± 0.02 cd | 4.74 ± 0.28 a |
| AcdP | 5.76 ± 0.90 bc | 3.41 ± 0.80 c | 11.78 ± 1.70 b | 2.87 ± 0.98 c | 10.98 ± 0.94 b | 18.09 ± 0.7 a |
| AlkP | 0.36 ± 0.09 c | 4.17 ± 0.70 b | 2.79 ± 0.47 c | 1.02 ± 0.57 c | 12.07 ± 3.01 a | 4.02 ± 0.52 b |
| Aryl | 1.57 ± 0.27 bc | nd | 8.42 ± 1.22 b | 1.04 ± 0.36 c | 1.12 ± 0.50 bc | 56.0 ± 3.5 a |
| FDA | 1.22 ± 0.12 b | 0.89 ± 0.10 b | 1.28 ± 0.09 b | 0.52 ± 0.04 c | 0.49 ± 0.02 c | 3.22 ± 0.21 a |
| Extraction | Bel1 | Bel2 | Bel3 | Yao1 | Yao2 | Yao3 |
|---|---|---|---|---|---|---|
| E1 | 6.28 ± 2.16 a | 7.30 ± 0.48 a | 6.82 ± 0.49 a | 18.32 ± 1.29 b | 8.90 ± 0.88 a | 4.07 ± 0.52 a |
| F1 | 4.87 ± 0.09 b | 4.97 ± 0.12 b | 4.21 ± 0.08 b | 10.39 ± 0.82 a | 5.55 ± 0.45 b | 1.71 ± 0.21 b |
| F2 | 0.36 ± 2.06 a | 1.27 ± 0.47 b | 1.33 ± 0.43 b | 1.23 ± 0.83 b | 1.89 ± 0.54 b | 1.28 ± 0.28 b |
| F3 | 1.04 ± 0.09 a | 1.04 ± 0.19 a | 1.27 ± 0.29 a | 6.69 ± 0.19 b | 1.46 ± 0.27 a | 1.08 ± 0.53 a |
| E2 | 65.8 ± 14.9 ab | 14.4 ± 1.6 bc | 44.1 ± 8.3 b | 70.4 ± 37.7 ab | 103.1 ± 5.3 a | 25.9 ± 4.0 b |
| E3 | 69.2 ± 5.1 c | 132.5 ± 5.9 b | 167.8 ± 9.8 a | 118.6 ± 7.1 bc | 92.8 ± 3.6 bc | 128.6 ± 14.9 b |
| E4 | 147.1 ± 12.2 a | 69.2 ± 4.0 ab | 53.9 ± 15.2 ab | 10.1 ± 3.7 b | 23.1 ± 1.6 b | 184.0 ± 4.8 a |
| Total Hg | 292.8 ± 16.5 ab | 224.1 ± 7.2 b | 273.9 ± 4.6 ab | 228.5 ± 11.5 b | 232.7 ± 3.9 b | 338.8 ± 11.6 a |
| Extraction yield (%) | 98.5 | 99.7 | 99.5 | 103.1 | 97.9 | 101.1 |
| E1 | E2 | E3 | E4 | F1 | F2 | F3 | HgT | |
|---|---|---|---|---|---|---|---|---|
| MBC | −0.90 | −0.84 | −0.26 | 0.84 | −0.93 | −0.60 | −0.83 | 0.92 |
| DH | −0.89 | −0.78 | −0.37 | 0.85 | −0.93 | −0.59 | −0.81 | 0.93 |
| Bglu | −0.89 | −0.79 | −0.39 | 0.86 | −0.93 | −0.59 | −0.81 | 0.93 |
| Urease | −0.90 | −0.84 | −0.35 | 0.88 | −0.93 | −0.62 | −0.82 | 0.93 |
| P5 | −0.91 | −0.64 | −0.42 | 0.79 | −0.94 | −0.51 | −0.87 | 0.91 |
| P9 | 0.09 | 0.45 | 0.14 | −0.37 | 0.09 | 0.72 | −0.05 | −0.31 |
| Aryl | −0.88 | −0.81 | −0.40 | 0.88 | −0.91 | −0.63 | −0.79 | 0.94 |
| FDA | −0.92 | −0.84 | −0.45 | 0.93 | −0.94 | −0.72 | −0.83 | 0.97 |
| Clay | 0.94 | 0.59 | 0.69 | −0.90 | 0.92 | 0.75 | 0.94 | −0.86 |
| TOC | −0.90 | −0.86 | −0.30 | 0.87 | −0.93 | −0.60 | −0.83 | 0.92 |
| Al | 0.68 | 0.70 | 0.43 | −0.79 | 0.66 | 0.92 | 0.60 | −0.75 |
| Fe | −0.46 | −0.12 | −0.10 | 0.22 | −0.48 | −0.22 | −0.44 | 0.47 |
| pH | 0.70 | 0.83 | −0.08 | −0.62 | 0.73 | 0.20 | 0.70 | −0.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Couic, E.; Alphonse, V.; Livet, A.; Giusti-Miller, S.; Bousserrhine, N. Influence of Ecological Restoration on Mercury Mobility and Microbial Activities on Former Guyanese Mining Sites. Appl. Sci. 2021, 11, 2231. https://doi.org/10.3390/app11052231
Couic E, Alphonse V, Livet A, Giusti-Miller S, Bousserrhine N. Influence of Ecological Restoration on Mercury Mobility and Microbial Activities on Former Guyanese Mining Sites. Applied Sciences. 2021; 11(5):2231. https://doi.org/10.3390/app11052231
Chicago/Turabian StyleCouic, Ewan, Vanessa Alphonse, Alexandre Livet, Stéphanie Giusti-Miller, and Noureddine Bousserrhine. 2021. "Influence of Ecological Restoration on Mercury Mobility and Microbial Activities on Former Guyanese Mining Sites" Applied Sciences 11, no. 5: 2231. https://doi.org/10.3390/app11052231