Brassica oleracea Prevents HCl/Ethanol-Induced Gastric Damages in Mice
by
 ,
,
Mi-Ryung Kim
1,†, 
Tae-Il Kim
1,†,
Beom-Rak Choi
2,
Moon Bong Kim
3,
Il Je Cho
4,*,
Keun-Woo Lee
1,* and
Sae Kwang Ku
4,* 1
College of Veterinary Medicine, Kyungpook National University, Daegu 41566, Korea
2
Nutracore Co., Ltd., Suwon, Gyeonggi-do 16514, Korea
3
Echo Trading Co., Seongnam, Gyeonggi-do 13596, Korea
4
College of Korean Medicine, Daegu Haany University, Gyeongsan, Gyeongsangbuk-do 38610, Korea
*
Authors to whom correspondence should be addressed.
†
M.-R.K. and T.-I.K. contributed equally to this work.
Appl. Sci. 2021, 11(1), 16; https://doi.org/10.3390/app11010016
Submission received: 25 November 2020
/
Revised: 18 December 2020
/
Accepted: 20 December 2020
/
Published: 22 December 2020
(This article belongs to the Special Issue Functional Food and Chronic Disease)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Brassica oleracea var. capitata L. (cabbage) is a popular vegetable with a wide range of pharmacological activities that help to promote human health. The present study investigated the beneficial effects of B. oleracea var. capitata L. extract (BOE) on HCl/ethanol (H/E)-induced gastric damages in mice. Pre-administration of BOE (25–100 mg/kg) for 7 consecutive days significantly decreased macroscopically visible lesion on the gastric mucosa induced by H/E. In addition, results from hematoxylin and eosin-stained gastric tissue showed that BOE inhibited invaded percentage of lesion and prevented the reduction in mucosal thickness in peri-ulcerative region. BOE significantly alleviated the H/E-mediated decreases in Alcian blue binding, total hexose, sialic acid, and collagen in the gastric tissue, suggesting BOE attenuates the gastric damage via preserving the integrity of gastric mucus. Moreover, BOE significantly decreased histamine level in the plasma and reduced mRNA levels associated with secreting gastric acid. Furthermore, BOE inhibited myeloperoxidase activity and suppressed nuclear factor-κB mRNA and its dependent inflammatory genes expression induced by H/E. BOE also strengthened antioxidant enzyme activity, with a mitigating H/E-mediated increase in malondialdehyde level of the gastric tissue. Thus, these results suggest that BOE has the potential to protect the gastric tissue via inhibiting gastric acid secretion, inflammation, and oxidative stress.
1. Introduction
Gastric ulcer is a multifactorial common disorder induced by dysregulation of acid-peptic activity in gastric juice, and lifetime prevalence of the disease has been estimated to be approximately 5 to 10% of general population [1]. Several aggressive factors, such as alcohol, smoking, non-steroidal anti-inflammatory drugs, and Helicobacter pylori infection has been considered to cause gastric ulcer by inhibiting mucosal defensive mechanisms (e.g., tight junctions between epithelial cells, microvascular blood circulation, and bicarbonate buffering system) [2,3]. Especially, it has been reported that alcohol rapidly disrupts mucosal layer and exposes the gastric mucosa to acidic and enzymatic degradation. In addition, alcohol-mediated decrease in blood flow provokes microvascular injuries and accumulates production of reactive oxygen species and pro-inflammatory cytokines [4]. Furthermore, the alcohol-mediated gastric damages can be more aggravated in the presence of hydrogen chloride (HCl) [5]. Although cessation of alcohol would be essential for treating an alcohol-mediated gastric ulcer, it is difficult for heavy drinkers to maintain the abstinence. Moreover, standard pharmacological agents for treating gastric ulcer (e.g., histamine receptor blockers and proton pump inhibitors) still confer mild to severe side effects.
A number of plants, vegetables, and those-derived natural compounds have recently received great attention for their ability to prevent gastric ulcer [6,7,8,9,10]. Among diverse natural resources, Brassica oleracea var. capitata L. (common name, cabbage; family, Brassicaceae) grows over 90 countries including South Korea and is rich in minerals (e.g., calcium and potassium), micronutrients (e.g., vitamin C and K), and phytochemicals (e.g., vitamin U, glucosinolates, and polyphenols) [11,12]. Growing evidence suggests that B. oleracea has a wide range of prophylactic potential for preventing cancer, contact dermatitis, dyslipidemia, hepatitis, and hyperglycemia [11,13,14,15,16]. Especially, early clinical investigation has reported that cabbage juice can heal peptic ulcer in human [12,17]. However, little attention has been paid to proving the gastroprotective effect of cabbage in experimental animals. Only two experimental studies have shown that post-administration of cabbage aqueous extract shortens the length of gastric ulcer, increases pH of gastric juice, and alleviates the histopathological ulcerative changes caused by administration of nonsteroidal anti-inflammatory drugs (e.g., aspirin) [18,19]. Although B. oleracea has the potential for preventing gastric ulcer, detailed mechanisms for B. oleracea-mediated gastroprotection have not been fully understood.
Of diverse toxicological, physiological, and surgical models for gastric ulcer, the experimental model administered by HCl/ethanol (H/E) resembles pathophysiological features of human acute gastric ulcer [10]. Thus, H/E-induced gastric ulcers have been frequently used to study the pathogenesis of gastric ulcer and to evaluate the anti-ulcerative effect of valuable candidates [6,7,8,9]. The present study aims to assess the protective effect of B. oleracea var. capitata L. extract (BOE) against H/E-induced gastric damage in mice and to investigate the underlying mechanisms. Furthermore, as the first process for developing the potent alternative gastroprotective ingredient for functional food, gastroprotective effects of BOE are compared to those of ranitidine hydrochloride (RA), a representative histamine H2 receptor (H2R) antagonist.
2. Materials and Methods
2.1. Reagents
BOE powder was supplied by grow® company (Ridgefield, NJ, USA) via Echo Trading Co. Ltd. (Seoul, Korea), and physical, nutritional and safety properties of BOE were authenticated by grow® company (Ridgefield, NJ, USA; data not shown). TRIzol and a high-capacity cDNA reverse transcription kit were obtained from Thermo Fisher Scientific (Rockford, IL, USA). Enzyme-linked immunosorbent assay (ELISA) kits for mouse histamine, tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and IL-18 were provided by Mybiosource (San Diego, CA, USA). Vitamin U, RA, HCl, ethanol, hematoxylin, eosin, Alcian blue, hydrogen peroxide, o-dianisidine dihydrochloride, thiobarbituric acid, nitroblue tetrazolium, and other chemicals with analytical grade were purchased from Sigma-Aldrich (St. Louise, MO, USA).
2.2. Quantification of Vitamin U in BOE
Vitamin U and BOE were analyzed using an agilent liquid chromatography system (Waters Inc.; Milford, MA, USA) equipped with Capcellpak UG120 C18 column (pore size, 5 μm; column size, 4.6 × 250 mm) and photodiode array detector. Vitamin U or BOE was eluted in a gradient solution comprising 40 mM NaH2PO4 (pH 7.8) and acetonitrile: methanol: water (45: 45: 10). Vitamin U was detected at a wavelength of 338 nm. The standard curve for vitamin U (peak area versus concentration) was obtained after various concentrations of vitamin U were injected to liquid chromatography. The concentration of vitamin U in BOE was calculated by interpolating the peak area representing the same retention time into the standard curve.
2.3. Animals and Treatment
All laboratory animals were maintained according to the national regulations of the usage and welfare of laboratory animals, and experimental protocol was approved by the Institutional Animal Care and Use Committee of Daegu Haany University (Approval No. DHU2019-089). Sixty ICR mice (gender, male; age, six weeks old upon receipt) were supplied by OrientBio (Seungnam, Korea), acclimatized for 9 days, and divided into six groups (N = 10 per group): vehicle, H/E, H/E + RA, H/E + 100 mg/kg BOE, H/E + 50 mg/kg BOE, H/E + 25 mg/kg BOE. Test materials were prepared by dissolving appropriate amount of BOE or RA in distilled water, and HCl/ethanol (H/E) was prepared by mixing 98% ethanol with 150 mM HCl [7]. Mice in H/E + BOE groups were orally administered 100, 50, or 25 mg/kg BOE once daily for 7 days. On the contrary, mice in the H/E + RA group were orally received 100 mg/kg RA on day 7 after 6 days of water administration. Mice in vehicle and H/E groups were only treated with equal volume of water for 7 days. At 1 h after the last drug treatment, gastric damage was induced by oral administration of H/E (5 mL/kg). Mice in the vehicle group were received the same amount of water instead of H/E. All mice were euthanized at 1 h after H/E administration. Blood and gastric tissue were collected to assess the effects of BOE.
2.4. Quantification of Gross Lesions
Gastric tissue was opened along the greater curvature and then washed mildly with water to remove the gastric contents and blood clots. Total area showing the ulcerous lesions was quantified as mm2 of gastric mucosa using an image analyzer (iSolution FL ver 9.1, IMT i-solution Inc.; Burnaby, BC, Canada).
2.5. Histopathology of Gastric Tissue
Preparation of paraffin-embedded gastric tissue, hematoxylin and eosin staining, and histomorphometric analyses were conducted, as described previously [7]. Briefly, tissue section from the fundus/corpus areas was observed under light microscopy (Eclipse 80i, Nikon; Tokyo, Japan). Using an image analyzer (iSolution FL ver 9.1), invaded percentage of lesion was expressed as a percentage of lesion length in fundus/corpus wall, and mucosa thickness was measured from luminal mucosal surface to muscularis mucosa in the peri-ulcerative region. Histological score was semi-quantitatively estimated according to four grades: 0—normal intact mucosa; 1—slight surface erosive damage; 2—moderate mucosa damage; 3—severe total mucosa damage.
2.6. Preparation of Plasma and Gastric Tissue Homogenates
Plasma was prepared from the 50 unit/mL heparin-treated blood by centrifugation at 12,500× g for 10 min. Gastric tissue in 10 volumes of ice-cold phosphate-buffered saline was homogenized using bead beater (TacoTM Prep, GeneReach Biotechology; Taichung, Taiwan) and ultrasonic cell disruptor (KS-750, Madell Technology; Ontario, CA, USA). The homogenized tissue was then centrifuged at 11,000× g for 15 min, and the resulting supernatant was used as gastric tissue homogenates.
2.7. Measurement of Gastric Mucus, Glycoprotein, and Collagen
Gastric mucus was quantified by the amount of Alcian blue binding to the gastric tissue [9]. The gastric tissue was directly immersed in assay solution (0.02% Alcian blue, 0.16 M sucrose, and 0.05 M sodium acetate (pH 5.8)) for 24 h and centrifuged at 3000× g for 10 min. After absorbance of the supernatant was measured at 620 nm using a spectrophotometer (OPTIZEN POP, Mecasys; Daejeon, Korea), the amount of the Alcian blue binding (mg) was normalized by tissue weight. Total hexose, sialic acid, and collagen contents in the gastric tissue were quantified, as described previously [20]. Briefly, total hexose of mucosal glycoproteins was quantified by measuring the intensity at 425 nm of wavelength after tissue homogenates were boiled with sulfuric acid and 5-methyl orcinol. To assess sialic acid bound to mucosal glycoprotein, proteins in the homogenates were precipitated by adding ethanol. The precipitate was dissolved in sulfuric acid and oxidized by periodic acid. After oxidation reaction terminated by adding sodium arsenate, the mixture was further reacted with thiobarbituric acid and measured absorbance at 550 nm. For quantifying collagen, tissue homogenate was hydrolyzed by HCl, subsequently reacted with chloramine-T and Ehrlich’s reagents, and measured absorbance at 565 nm. The amounts of mucosal components were normalized by tissue weight.
2.8. ELISA
Histamine in plasma and pro-inflammatory cytokines (e.g., TNF-α, IL-1β, IL-6, and IL-18) in gastric tissue homogenate were quantified using commercial ELISA kits (Mybiosource), according to the manufacturer’s instructions. The level of histamine or pro-inflammatory cytokines was determined at 450 nm of wavelength using a microplate reader (Sunrise, Tecan; Männedorf, Switzerland), and presented as pg/mL (for histamine) or pg/mg (for pro-inflammatory cytokines).
2.9. Real-Time Polymerase Chain Reaction (PCR)
Total RNA isolation from the gastric tissue using a TRIzol reagent, reverse transcription reaction for obtaining cDNA, and real-time PCR were conducted, as described previously [21]. To amplify specific genes, the following oligonucleotides were synthesized: H2R (GenBank accession #, NM001010973.2), 5′-CTGGCTGTCAGCTTGAATCG-3′ (sense) and 5′-GCTGCCAGGGACACAATGA-3′ (anti-sense); H+/K+-ATPase (GenBank accession #, NM009724.2), 5′-CTACACCCCAGACTACCAGGAC-3′ (sense), and 5′-TAAACTCCACCTTCCCTTCGTA-3′ (anti-sense); cholecystokinin-2 receptor (CCK2R; GenBank accession #, NM007627.5), 5′-GATGGCTGCTACGTGCAACT-3′ (sense) and 5′- CGCACCACCCGCTTCTTAG-3′ (anti-sense); nuclear factor-κB (NF-κB; GenBank accession #, M61909.1), 5′-CAATGGCTACACAGGACCA-3′ (sense) and 5′-CACTGTCACCTGGAAGCAGA-3′ (anti-sense); cyclooxygenase-2 (COX-2; GenBank accession #, NM011198.4), 5′-CACTACATCCTGACCCACTT-3′ (sense) and 5′-ATGCTCCTGCTTGAGTATGT-3′ (anti-sense); inducible nitric oxide synthase (iNOS; GenBank accession #, M92649.1), 5′-CCCTTCCGAAGTTTCTGGCAGCAG-3′ (sense) and 5′-GGCTGTCAGAGCCTCGTGGCTTTGG-3′ (anti-sense); glyceraldehyde 3-phosphate dehydrogenase (GAPDH; GenBank accession #, GU214026.1), 5′-CATCTTCCAGGAGCGAGACC-3′ (sense) and 5′-TCCACCACCCTGTTGCTGT -3′ (anti-sense). GAPDH was used as an endogenous housekeeping gene for quantifying relative level of specific gene, and the expression level of the gene was calculated according to the 2-ΔΔCT method [22].
2.10. Measurement of Myeloperoxidase (MPO) Activity
MPO activity in the gastric tissue was determined, as described previously [23]. Briefly, the gastric tissue in 50 mM potassium phosphate buffer (pH 6.0) and 0.5% (w/v) cetrimonium bromide was homogenized and clarified at 12,000× g. After discarding the supernatant, the pellet was resuspended in the phosphate buffer containing 0.5% (w/v) cetrimonium bromide and 10 mM ethylenediaminetetraacetic acid, followed by incubating with 0.0005% hydrogen peroxide and 0.167 mg/mL o-dianisidine dihydrochloride. MPO activity was measured at 460 nm and normalized by tissue weight.
2.11. Determination of Malondialdehyde and Antioxidant Activities in the Gastric Tissue
The level of malondialdehyde in the gastric tissue was measured, as described previously [7]. To determine malondialdehyde level, reaction mixture comprising gastric tissue homogenates, sodium dodecyl sulfate, acetic acid, and 2-thiobarbiturate were incubated at 98 °C for 1 h. After n-butanol: pyridine (15: 1) were added to the reaction mixture and centrifuged at 4000× g for 30 min, absorbance of the resulting supernatant was determined at 512 nm. Molar concentration of malondialdehyde was calculated by interpolating absorbance intensity to standard curve and normalized by tissue weight. Enzyme assay for superoxide dismutase and catalase activity in the gastric tissue were conducted using nitroblue tetrazolium and hydrogen peroxide, as described previously [23,24]. Enzyme activity was normalized by tissue weight.
2.12. Statistical Analysis
Numerical results were presented as the mean ± standard deviation (SD). To compare means, Tukey honestly significant difference or Dunnett’s T3 test was conducted after one-way analysis of variance or Welch test. p value less than 0.05 was considered as significant.
3. Results
3.1. Quantification of Vitamin U in BOE
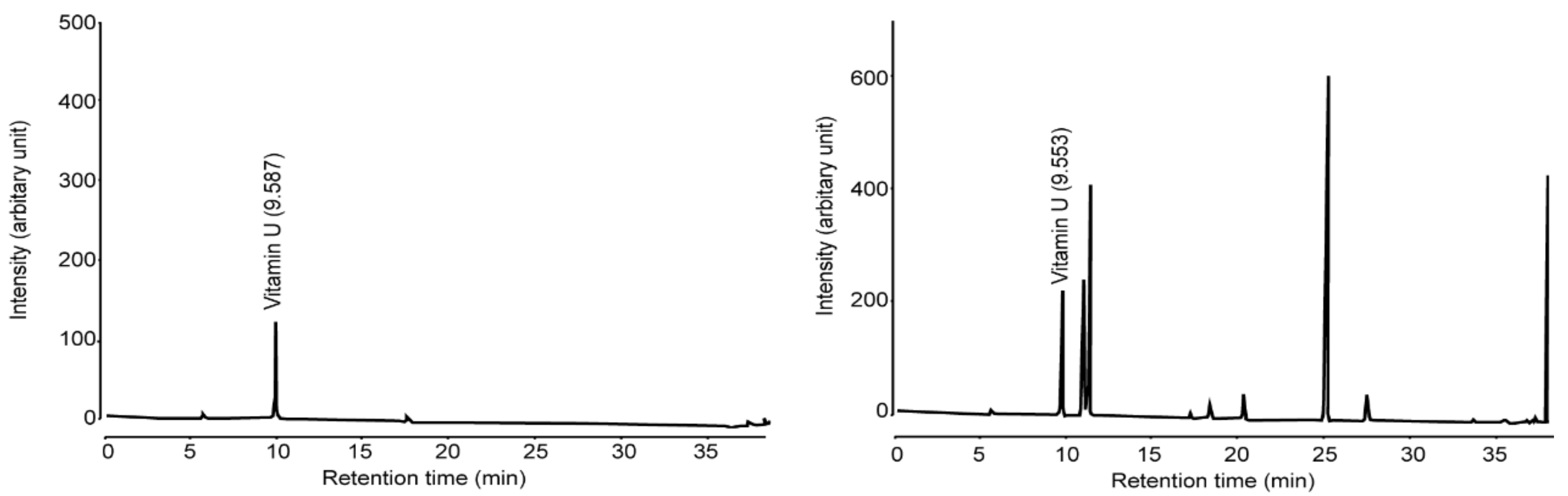
Because vitamin U has been known as a representative gastroprotective natural product found in B. oleracea [25,26], we firstly quantified the level of vitamin U in BOE. After establishing the detection condition of vitamin U by high-performance liquid chromatography (Figure 1, left), we found that BOE used in the present study contained 50.26 mg/g of vitamin U (Figure 1, right).
3.2. BOE Reduces Gastic Damages Induced by H/E
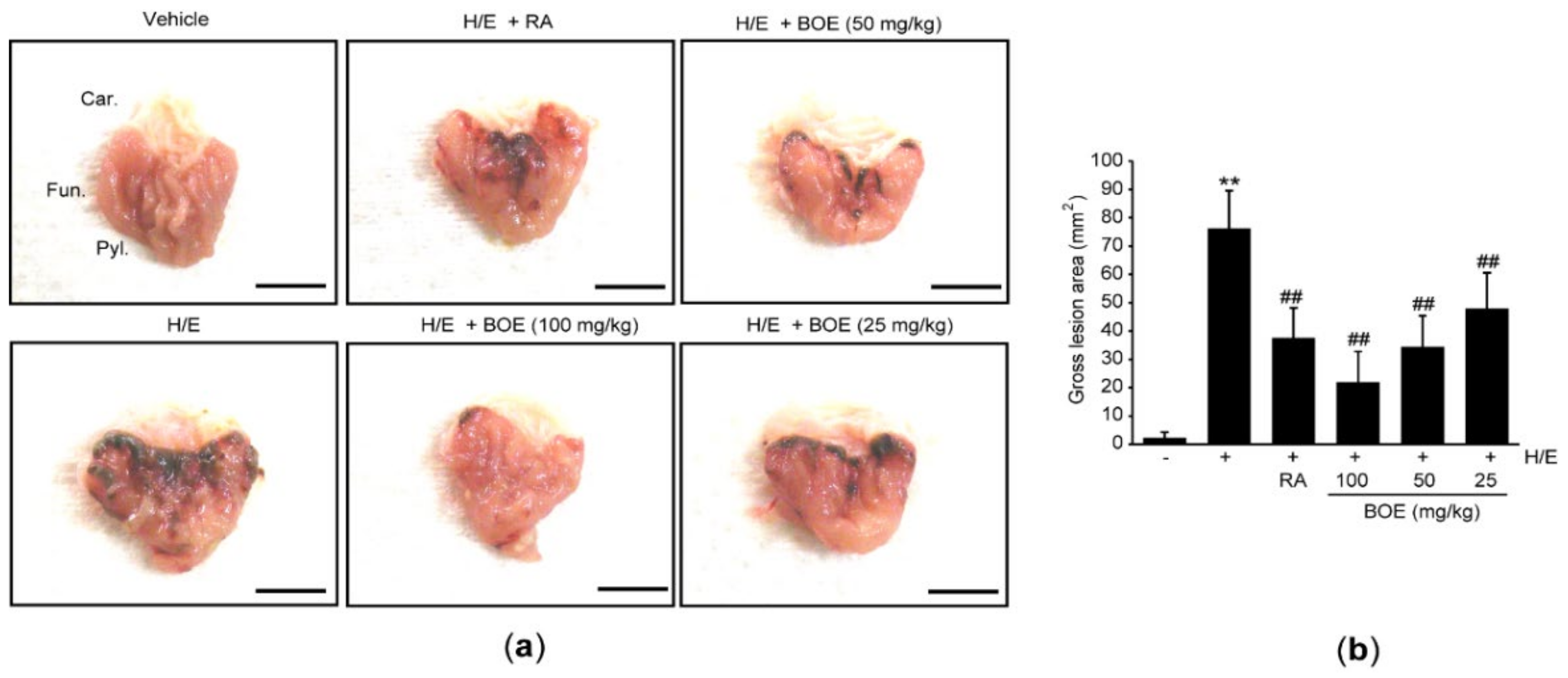
ICR mice were given 25–100 mg/kg BOE for seven consecutive days and induced gastric damages for 1 h by administration of H/E. To compare the gastroprotective effect of BOE, RA (100 mg/kg) was administered once into the mice. During the experimental procedure, there were no differences in body weight among the experimental groups (data not shown). When gastric tissues were grossly observed at 1 h after H/E administration, focal hemorrhagic ulcerative damages were found in H/E group. However, administration of three doses of BOE for 7 days as well as single administration of RA significantly reduced the H/E-mediated macroscopic visible lesions in a dosage-dependent manner (Figure 2). There were no statistical differences in gross gastric damage between the group treated with H/E + BOE and H/E + RA.
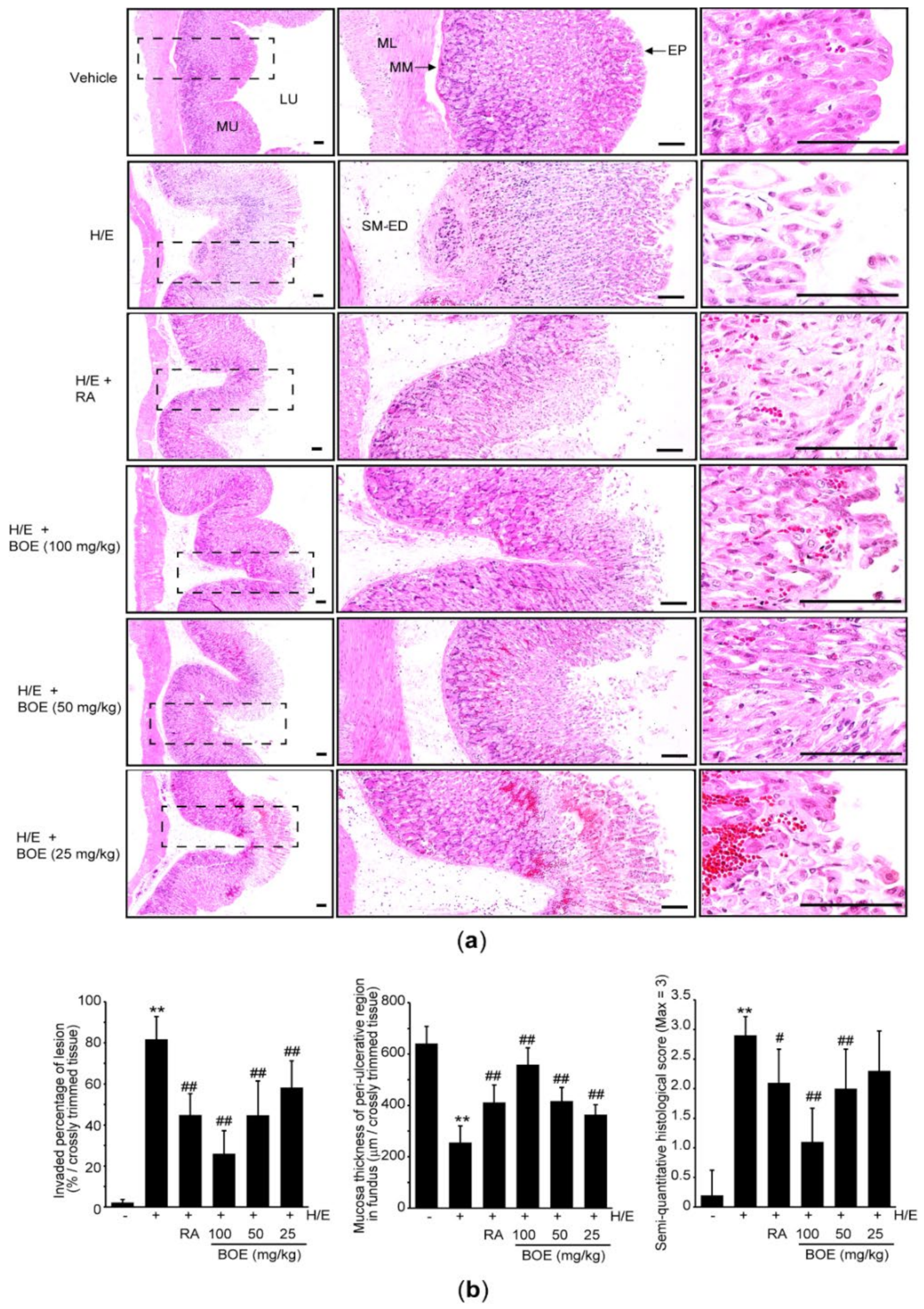
Next, the gastric tissue was stained with hematoxylin and eosin to explore the pathological changes more in depth. Results from hematoxylin and eosin staining of the gastric tissue revealed that H/E provoked severe ulcerative lesions, which included focal extensive superficial epithelial damage, desquamation of focal epithelium, edema of submucosa, congestion and hemorrhage, infiltration of inflammatory cells, and necrosis of gastric glands. Pre-administration of BOE or RA significantly attenuated those microscopic ulcerative changes induced by H/E (Figure 3a). Especially, histomorphometric analysis indicated that H/E administration significantly increased the semi-quantitative histological score (Figure 3b, right) as a result of an increase in invaded percentage of lesion in the fundus/corpus regions (Figure 3b, left) as well as reduction in mucosal thickness in the peri-ulcerative region (Figure 3b, middle). These H/E-mediated histopathological changes were significantly alleviated by the administration of 25–100 mg/kg BOE, except for semi-quantitative histological score, which did not differ between the H/E + BOE (25 mg/kg) group and the H/E group. RA administration also significantly prevented the H/E-mediated histopathological changes in the gastric tissue. When the preventive effect of BOE was compared to RA, 100 mg/kg BOE yielded a greater reduction in H/E-mediated histomorphometric changes relative to RA (p < 0.0.5 in invaded percentage of lesion; p < 0.01 in mucosa thickness; p < 0.01 in semi-quantitative histological score). However, the preventive effects by 25 and 50 mg/kg BOE did not differ from those by RA.
3.3. BOE Attenuates Mucus Loss Induced by H/E
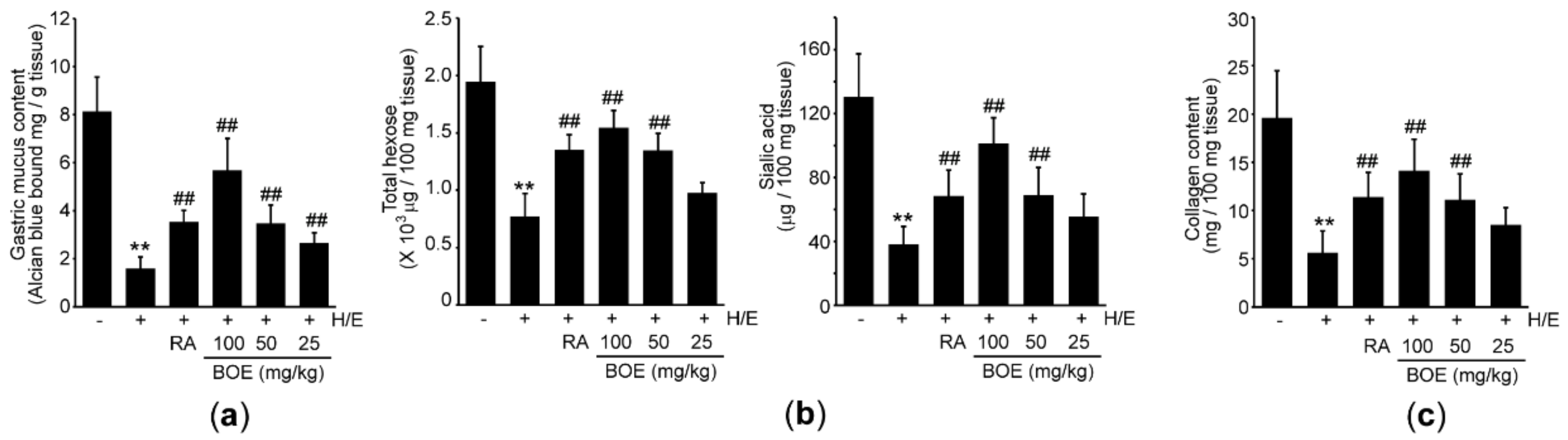
To explore the effect of BOE on integrity of the gastric mucus, we determined the mucus content via Alcian blue binding assay. In parallel with results from histopathology, pre-administration of BOE significantly attenuated H/E-mediated loss of gastric mucus in a dosage-dependent manner (Figure 4a). RA also significantly prevented the mucus loss, and its effect did not differ from that seen in response to 50 mg/kg BOE. Next, we quantified the level of carbohydrates in the gastric mucus. Compared to the vehicle, H/E significantly decreased the levels of total hexose and sialic acid, suggesting that H/E damages glycoprotein in the gastric mucus. By contrast, 50–100 mg/kg BOE as well as RA significantly blocked the decreases in the level of total hexose and sialic acid. However, the carbohydrate levels by 25 mg/kg BOE did not differ from those by H/E (Figure 4b). Compared between H/E + BOE and H/E + RA group, the magnitude of the sialic acid’s restoration by 100 mg/kg BOE was only greater than that by RA (p < 0.01). In addition, 50–100 mg/kg BOE and RA significantly prevented the H/E-mediated decrease in collagen (Figure 4c), which is a representative structural protein found in the extracellular matrix [27]. There were no statistical differences in collagen content between the group treated with three different doses of BOE and RA.
3.4. BOE Decreases Histamine in the Plasma and Reduces mRNAs Expression Associated with Secreting Gastric Acid
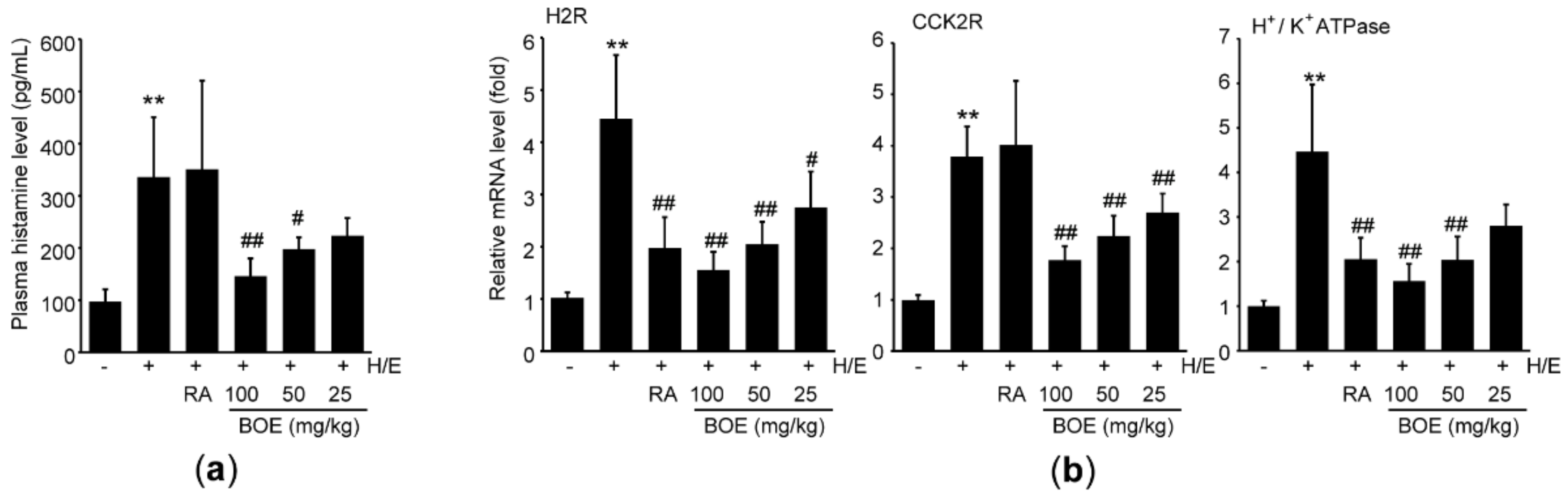
Because it has been reported that ethanol decreases gastric pH in experimental animals [28], we further explored the effect of BOE on the major regulating factors for secreting gastric acid. Compared to vehicle, H/E significantly increased plasma level of histamine. Although pre-administration of RA did not attenuate H/E-mediated increase in plasma histamine, 50–100 mg/kg BOE significantly decreased plasma histamine level, relative to H/E group (Figure 5a), and the magnitude of the reduction in histamine level by 100 mg/kg BOE was greater than that by RA (p < 0.05). Real-time PCR analysis was conducted to assess the effects of BOE on major responsible genes for secreting gastric acid. In parallel with the result from plasma histamine, H/E significantly induced mRNA expression levels of H2R, CCK2R, and H+/K+ ATPase. However, H/E-mediated increases in those mRNAs were significantly inhibited by the three different doses of BOE, except for H+/K+ ATPase mRNA, which did not differ significantly between H/E and H/E + BOE (25 mg/kg) group. In addition, mRNA levels of H2R and CCK2R, but not H+/K+ ATPase, were also reduced by pre-administration of RA, and decreases in H2R and CCK2R mRNAs by RA did not differ from those by 50–100 mg/kg BOE (Figure 5b).
3.5. BOE Inhibits H/E-Mediated Inflammation in the Gastric Tissue
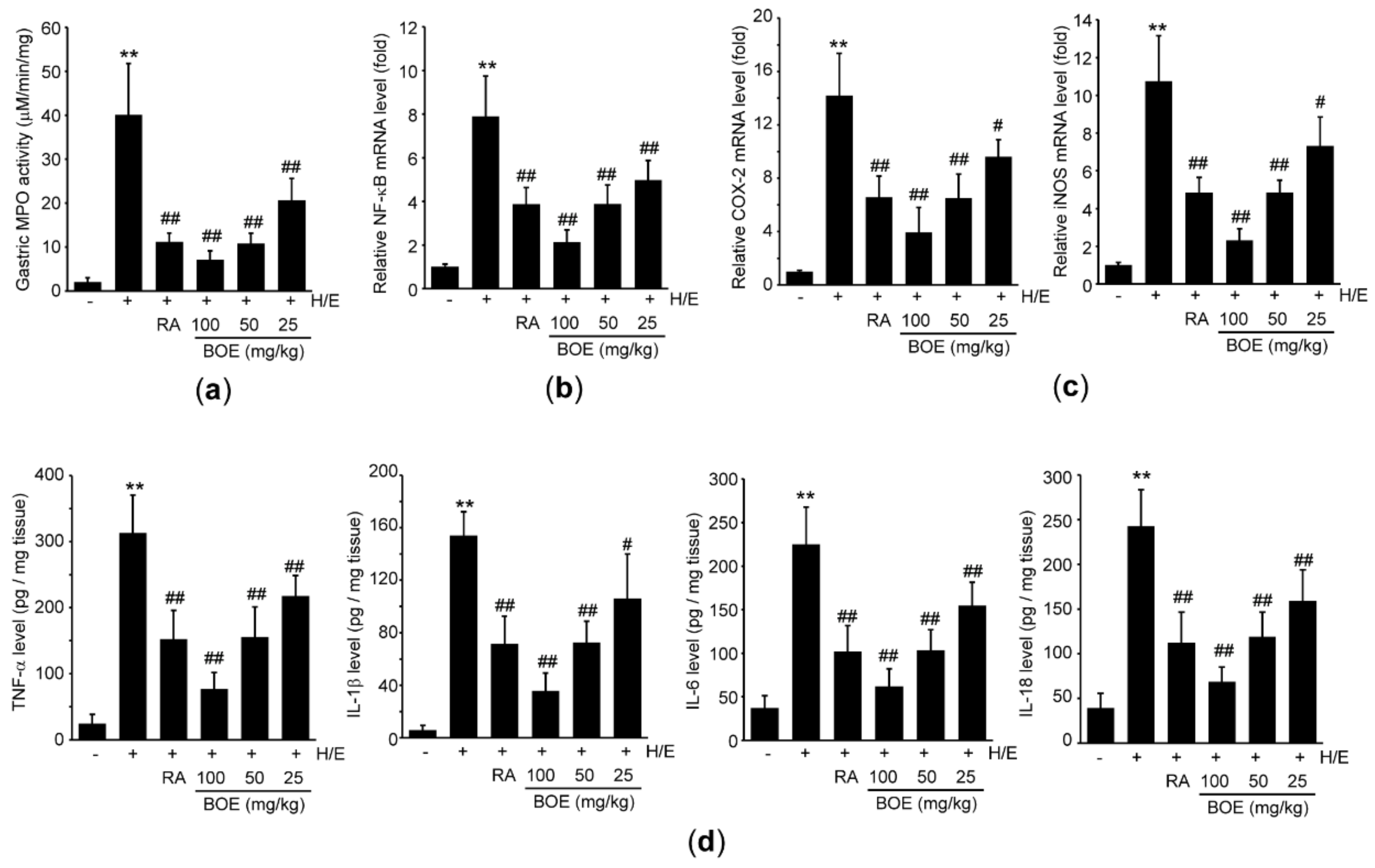
To explore whether BOE protects the gastric tissues by reducing H/E-mediated acute inflammation, we measured MPO activity in the gastric tissue. Compared to vehicle group, H/E significantly increased the MPO activity. However, pre-administration of three different doses of BOE as well as RA significantly attenuated an H/E-mediated increase in MPO activity (Figure 6a). Next, mRNA and protein levels of pro-inflammatory mediators were determined using real-time PCR and ELISA, respectively. As expected, H/E significantly increased the mRNA and protein levels of NF-κB (Figure 6b), COX-2, iNOS (Figure 6c), TNF-α, IL-1β, IL-6, and IL-18 (Figure 6d) compared to vehicle. Pre-administration of three different doses of BOE as well as RA significantly inhibited the increases in mRNA and protein levels of pro-inflammatory mediators induced by H/E (Figure 6b–d). Although the magnitude of the reduction in pro-inflammatory mediators by 100 mg/kg BOE was greater than that by RA (p < 0.01), the inhibitory effect by 50 mg/kg BOE did not differ from that seen in response to RA.
3.6. BOE Alleviates H/E-Mediated Lipid Peroxidation and Enhances Antioxidant Activity
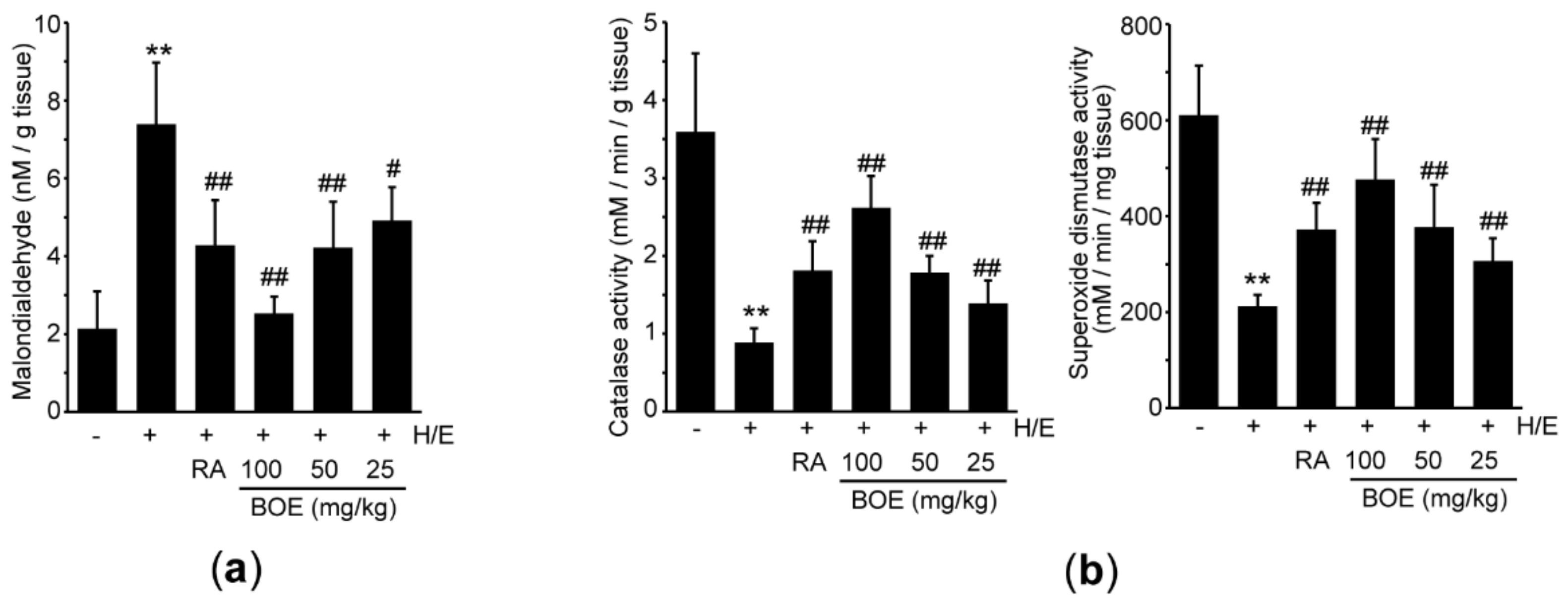
To investigate whether BOE prevents H/E-mediated tissue damage by reducing oxidative stress, malondialdehyde (a marker of lipid peroxidation) level was measured using the gastric tissue homogenate. Compared to vehicle, H/E significantly increased the level of malondialdehyde. All three different doses of BOE significantly decreased the malondialdehyde level, relative to the H/E group (Figure 7a), and the reduction in malondialdehyde by 100 mg/kg BOE was more potent than that seen in response to RA (p < 0.05). Next, we explored the effects of BOE on antioxidant activity in H/E-treated mice. H/E significantly inhibited catalase and superoxide dismutase activities. All three doses of BOE as well as RA significantly alleviated the reduction in catalase and superoxide dismutase activities (Figure 7b). There were no differences between the group treated with H/E + BOE and H/E + RA, except for catalase activity, which the magnitude of restorative effect by 100 mg/kg BOE was greater than that seen by RA (p < 0.01).
4. Discussion
Dr. Cheney referred vitamin U as an uncharacterized gastroprotective factor in cabbage juice [12], and this factor has been denominated S-methylmethionine sulfonium chloride [29]. Although vitamin U is not classified as a real vitamin, deficiency of vitamin U is thought to induce gastric ulcers [26]. This heat-labile factor recovers mucosal integrity by enhancing secretion of mucin [26,30], and sulfhydryl group after metabolic transformation is considered a pivotal chemical moiety for vitamin U-mediated gastroprotection [26]. In addition, it has been also reported that vitamin U can inhibit differentiation of pre-adipocytes by activating AMP-activated protein kinase [31]. Moreover, accumulated evidence has suggested that vitamin U reveals diverse beneficial effects, which include hypolipidemic, hepatoprotective, anti-inflammatory, and cytoprotective activities [32,33,34,35]. Because plant species belonging to the Brassicaceae family contain the highest amount of vitamin U [25], in the present study, vitamin U was quantified as a representative biomarker of BOE. Furthermore, quantifying vitamin U will help assess the quality of BOE in the process of developing BOE as a functional food.
Although we reported that BOE contained 50.26 mg/g vitamin U, the concentration of vitamin U was too low to explain maximal gastroprotective effect of BOE. Of diverse phytochemicals found in cabbage, it has been reported that allyl isothiocyanate alleviates the erosion of gastric mucosa through growth inhibition of H. pylori [36]. In addition, iberin and sulforaphane inhibit the activity of H. pylori urease [37]. Moreover, indol-3-carbinol alone or in combination with omeprazole shows profound protection to the gastric mucosa in aspirin-induced gastric ulcer [38]. Furthermore, several polyphenols such as quercetin, kaempferol, and apigenin, have also been reported to exhibit beneficial effects on gastric ulcer in experimental animals [39,40,41]. Thus, vitamin U as well as other phytochemicals in BOE may contribute collaboratively to prevent gastric ulcer caused by H/E. Further studies are warranted to identify other valuable biomarkers in BOE.
Results from macroscopic/microscopic histopathological assessment indicated that BOE decreased hemorrhagic gross lesions and restored mucosal thickness in the peri-ulcerative region after H/E treatment. Mucus secreted from mucous cells in the gastric pits is regarded as the first barrier to diminish pathogen penetration and to protect the tissue from ulcerogenic stimuli [42]. Mucus is primarily composed to mucin, a highly viscous glycosylated proteins, and the carbohydrate side chains account for more than 80% of the weight of glycoprotein in the stomach [43]. In addition, collagen is an essential extracellular matrix protein for providing the strength and sustaining (sub)mucosal and muscular integrity of the stomach [44]. Thus, the increase in Alcian blue binding of the gastric tissue as well as increases in carbohydrate and collagen of the tissue homogenates provide a direct evidence that BOE can protect from H/E-induced gastric damages by restoring mucus and sustaining mucosal integrity.
It has been reported that enhanced secretion of gastric acids facilitates gastric damage by excess intake of ethanol or acids [28,45]. Among diverse neurohumoral signaling molecules, histamine released from enterochromaffin-like cells and/or neuroinflammatory cells stimulates the secretion of gastric acid in the parietal cells via H2R activation. In addition, gastrin directly activates CCK2R in the parietal cells and indirectly stimulates histamine release from the enterochromaffin-like cells [46,47]. Activation of H2R and CCK2R leads to pumping out H+ ion to the gastric lumen via H+/K+-ATPase [46]. In the present study, a significant increase in plasma histamine level was shown in H/E group, along with related increases in H2R, CCK2R, and H+/K+-ATPase mRNAs of the gastric tissue. However, H/E-mediated those changes were significantly blocked by pre-administration of BOE for 7 days, suggesting that BOE-dependent gastric protection from H/E can be attributed to the modulation of gastric acid.
Histamine secreted from the mast cells and basophils contributes to increase permeability of injured tissue and to recruit diverse inflammatory cells into the damaged sites. MPO is a representative enzyme stored in the azurophilic granules of the neutrophils [48]. In conjunction with the reduction in plasma histamine, present results showed that pre-administration of three different doses of BOE significantly decreased the MPO activity in the gastric tissue, suggesting that BOE can attenuate the infiltration of neutrophil to the gastric tissue. In parallel with previous report [6], the present results also showed that BOE significantly reduced the level of gastric NF-κB (p65 subunit) mRNA induced by H/E. NF-κB is a pivotal transcription factor, which accelerates pro-inflammatory response (e.g., cytokine and adhesion molecule) in a wide range of tissue including the stomach [49]. Damage-associated molecular patterns released from injured cells stimulate degradation of inhibitory κB, which sequestrate NF-κB in the cytoplasm, allow the NF-κB to translocate into the nucleus, and lead to an increase in the transcription of target genes including COX-2, iNOS, TNF-α, IL-1β, IL-6, and IL-18 [50,51]. Although inhibition of NF-κB signaling pathway by BOE may be further established by other experimental approaches (e.g., subcellular expression of NF-κB and post-translational modification of NF-κB signaling molecules), present results suggest that not only recruitment of inflammatory cells into the lesions but also expression of inflammatory mediators can be suppressed by three different doses of BOE through the modulation of NF-κB-dependent signaling molecules.
One of the most important executors in alcohol-mediated tissue damage is an oxidative stress, which is accumulated by dysregulation between endogenous antioxidants defense system and reactive oxygen (nitrogen) species [52]. Results from the present study showed that ethanol elevated the level of malondialdehyde (a representative marker of lipid peroxidation) and decreased the activities of catalase and superoxide dismutase in the gastric tissue, which is parallel with previous reports that alcohol provokes gastric damage through the induction of oxidative stress [7,8]. However, H/E-mediated lipid peroxidation and reduction in antioxidant enzymes activities were significantly attenuated by three different doses of BOE. Therefore, BOE-mediated upregulation of antioxidant defense system also contributes to protecting the gastric tissue from H/E.
5. Conclusions
The present results showed that BOE effectively alleviated H/E-mediated gastric damages by restoring mucosal integrity and inhibiting gastric acid secretion, acute inflammation, and oxidative stress. In addition, we compared the gastroprotective effect of BOE to those of RA. Although some gastroprotective effects by pre-administration of 100 mg/kg BOE were more potent than those by RA, most of the statistical results seen in response to 50–100 mg/kg BOE did not differ from RA. Because inflammation and oxidative stress are also considered the pivotal etiology in the progression of chronic gastric disease, BOE may have enough potential to develop as functional food for the prevention or treatment of acute and chronic gastric damages.
Author Contributions
Conceptualization, K.-W.L. and S.K.K.; methodology, M.-R.K., T.-I.K., and S.K.K.; formal analysis, M.-R.K., T.-I.K., M.B.K., I.J.C., and S.K.K.; investigation, M.-R.K., T.-I.K., I.J.C., and S.K.K.; resources, B.-R.C. and M.B.K.; data curation, I.J.C. and S.K.K.; writing—original draft preparation, M.-R.K., T.-I.K., I.J.C., and S.K.K.; writing—review and editing, I.J.C., K.-W.L., and S.K.K.; visualization, I.J.C. and S.K.K.; supervision, K.-W.L. and S.K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All laboratory animals were maintained according to the national regulations of the usage and welfare of laboratory animals, and experimental protocol was approved by the Institutional Animal Care and Use Committee of Daegu Haany University (Approval No. DHU2019-089).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lanas, A.; Chan, F.K.L. Peptic ulcer disease. Lancet 2017, 390, 613–624. [Google Scholar] [CrossRef]
- Kolgazi, M.; Ozdemir-Kumral, Z.N.; Cantali-Ozturk, C.; Demirci, E.K.; Yuksel, M.; Sirvanci, S.; Yegen, B.C. Anti-inflammatory effects of nesfatin-1 on acetic acid-induced gastric ulcer in rats: Involvement of cyclo-oxygenase pathway. J. Physiol. Pharmacol. 2017, 68, 765–777. [Google Scholar] [PubMed]
- Morsy, M.; El-Sheikh, A. Prevention of Gastric Ulcers. In Peptic Ulcer Disease; InTech Europe: London, UK, 2011. [Google Scholar]
- Franke, A.; Teyssen, S.; Singer, M.V. Alcohol-related diseases of the esophagus and stomach. Dig. Dis. 2005, 23, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Barreto, J.C.; Smith, G.S.; Russel, D.H.; Miller, T.A. Gastric damage caused by acidified ethanol: Role of molecular HCl. Am. J. Physiol. 1993, 265, G133–G137. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.H.; Park, E.J.; Kim, S.H.; Lee, H.J. Gastroprotective effect of fermented lotus root against ethanol/HCl-induced gastric mucosal acute toxicity in rats. Nutrients 2020, 12, 808. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.M.; Song, C.H.; Park, S.J.; Park, D.C.; Cho, H.R.; Jung, G.W.; Bashir, K.M.I.; Ku, S.K.; Choi, J.S. Protective effects of a triple-fermented barley extract (FBe) against HCl/EtOH-induced gastric mucosa damage in mice. Food Sci. Nutr. 2018, 6, 2036–2046. [Google Scholar] [CrossRef]
- Li, J.H.; Ju, G.X.; Jiang, J.L.; Li, N.S.; Peng, J.; Luo, X.J. Lipoic acid protects gastric mucosa from ethanol-induced injury in rat through a mechanism involving aldehyde dehydrogenase 2 activation. Alcohol 2016, 56, 21–28. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; do Nascimento Valença, J.D.; da Silva Santos, J.; Boeing, T.; da Silva, L.M.; de Andrade, S.F.; Albuquerque-Júnior, R.L.; Thomazzi, S.M. The effects of baicalein on gastric mucosal ulcerations in mice: Protective pathways and anti-secretory mechanisms. Chem. Biol. Interact. 2016, 260, 33–41. [Google Scholar] [CrossRef]
- Hamedi, S.; Arian, A.A.; Farzaei, M.M. Gastroprotective effect of aqueous stem bark extract of Ziziphus jujuba L. against HCl/Ethanol-induced gastric mucosal injury in rats. J. Tradit. Chin. Med. 2015, 35, 666–670. [Google Scholar] [CrossRef]
- Thangam, R.; Suresh, V.; Rajkumar, M.; Vincent, J.D.; Gunasekaran, P.; Anbazhagan, C.; Kaveri, K.; Kannan, S. Antioxidant and in vitro anticancer effect of 2-pyrrolidinone rich fraction of Brassica oleracea var. capitata through induction of apoptosis in human cancer cells. Phytother. Res. 2013, 27, 1664–1670. [Google Scholar] [CrossRef]
- Cheney, G. Anti-peptic ulcer dietary factor (vitamin “U”) in the treatment of peptic ulcer. J. Am. Diet. Assoc. 1950, 26, 668–672. [Google Scholar] [PubMed]
- Lee, Y.; Kim, S.; Yang, B.; Lim, C.; Kim, J.H.; Kim, H.; Cho, S. Anti-inflammatory effects of Brassica oleracea Var. capitata L. (Cabbage) methanol extracts in mice with contact dermatitis. Pharmacogn. Mag. 2018, 14, 174–179. [Google Scholar] [PubMed]
- Morales-López, J.; Centeno-Álvarez, M.; Nieto-Camacho, A.; López, M.G.; Pérez-Hernández, E.; Pérez-Hernández, N.; Fernández-Martínez, E. Evaluation of antioxidant and hepatoprotective effects of white cabbage essential oil. Pharm. Biol. 2017, 55, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S. Red cabbage (Brassica oleracea L.) mediates redox-sensitive amelioration of dyslipidemia and hepatic injury induced by exogenous cholesterol administration. Am. J. Chin. Med. 2014, 42, 189–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assad, T.; Khan, R.A.; Feroz, Z. Evaluation of hypoglycemic and hypolipidemic activity of methanol extract of Brassica oleracea. Chin. J. Nat. Med. 2014, 12, 648–653. [Google Scholar] [CrossRef]
- Doll, R.; Pygott, F. Clinical trial of robaden and of cabbage juice in the treatment of gastric ulcer. Lancet 1954, 267, 1200–1204. [Google Scholar] [CrossRef]
- Hadda, T.B.; ElSawy, N.A.; Header, E.A.M.; Mabkhot, Y.N.; Mubarak, M.S. Effect of garlic and cabbage on healing of gastric ulcer in experimental rats. Med. Chem. Res. 2014, 23, 5110–5119. [Google Scholar] [CrossRef]
- Carvalho, C.A.; Fernandes, K.M.; Matta, S.L.P.; Silva, M.B.; Oliveira, L.L.; Fonseca, C.C. Evaluation of antiulcerogenic activity of aqueous extract of Brassica oleracea var. capitata (cabbage) on wistar rat gastric ulceration. Arq. Gastroenterol. 2011, 48, 276–282. [Google Scholar]
- Cho, S.Y.; Song, C.H.; Lee, J.E.; Choi, S.H.; Ku, S.K.; Park, S.J. Effects of platycodin D on reflux esophagitis due to modulation of antioxidant defense systems. Evid. Complement. Altern. Med. 2018, 2018, 7918034. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.R.; Cho, I.J.; Jung, S.J.; Kim, J.K.; Park, S.M.; Lee, D.G.; Ku, S.K.; Park, K.M. Lemon balm and dandelion leaf extract synergistically alleviate ethanol-induced hepatotoxicity by enhancing antioxidant and anti-inflammatory activity. J. Food Biochem. 2020, 44, e13232. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, M.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.M.; Song, C.H.; Park, S.J.; Park, D.C.; Jung, G.W.; Cho, H.R.; Bashir, K.M.I.; Ku, S.K.; Choi, J.S. Protective effects of a triple-fermented barley extract (FBe) on indomethacin-induced gastric mucosal damage in rats. BMC Complement. Altern. Med. 2019, 19, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.R.; Chun, Y.S.; Kim, J.K.; Cho, I.J.; Ku, S.K. Ginseng berry aqueous extract prevents scopolamine-induced memory impairment in mice. Exp. Ther. Med. 2019, 18, 4388–4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rácz, I.; Paldi, E.; Szalai, G.; Janta, T.; Pal, M.; Lasztity, D. S-methylmethionine reduces cell membrane damage in higher plants exposed to low-temperature stress. J. Plant Physiol. 2008, 165, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Ohara, S.; Ichikawa, T.; Sigenji, K.; Hotta, K. Mechanisms for cytoprotection by vitamin U from ethanol-induced gastric mucosal damage in rats. Dig. Dis. Sci. 1996, 41, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Hendel, L. Hydroxyproline in the oesophageal mucosa of patients with progressive systemic sclerosis during omeprazole-induced healing of reflux oesophagitis. Aliment. Pharmacol. Ther. 1991, 5, 471–480. [Google Scholar] [CrossRef]
- Rahman, Z.; Dwivedi, D.K.; Jena, G.B. Ethanol-induced gastric ulcer in rats and intervention of tert-butylhydroquinone: Involvement of Nrf2/HO-1 signaling pathway. Hum. Exp. Toxicol. 2020, 39, 547–562. [Google Scholar] [CrossRef]
- Patel, A.D.; Prajapati, N.K. Review on biochemical importance of vitamin-U. J. Chem. Pharm. Res. 2012, 4, 209–215. [Google Scholar]
- Ichikawa, T.; Ito, Y.; Saegusa, Y.; Iwai, T.; Goso, Y.; Ikezawa, T.; Ishihara, K. Effects of combination treatment with famotidine and methylmethionine sulfonium chloride on the mucus barrier of rat gastric mucosa. J. Gastroenterol. Hepatol. 2009, 24, 488–492. [Google Scholar] [CrossRef]
- Lee, N.Y.; Park, K.Y.; Min, H.J.; Song, K.Y.; Lim, Y.Y.; Park, J.; Kim, B.J.; Kim, N.M. Inhibitory effect of vitamin U (S-methylmethionine sulfonium chloride) on differentiation of 3T3-L1 pre-adipocyte cell lines. Ann. Dermatol. 2012, 24, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Seo, H.M.; Kim, W.K.; Choi, J.S.; Kim, I.; Sung, J.H. The photoprotective effect of S-methylmethionine sulfonium in skin. Int. J. Mol. Sci. 2015, 16, 17088–17100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokmen, B.B.; Tunali, S.; Yanardag, R. Effects of vitamin U (S-methyl methionine sulphonium chloride) on valproic acid induced liver injury in rats. Food Chem. Toxicol. 2012, 50, 3562–3566. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Uzawa, H.; Kanazawa, K.; Tamai, Y.; Tashiro, Y.; Koide, M. Hypolipidemic effect of L-form S-methylmethionine sulfonium chloride in man. Arzneimittelforschung 1981, 31, 725–729. [Google Scholar] [PubMed]
- Urazaeva, L.G. Anti-inflammatory effect of methylmethionine sulfonium chloride (vitamin U). Farmakol. Toksikol. 1976, 39, 316–319. [Google Scholar] [PubMed]
- Masuda, S.; Masuda, H.; Shimamura, Y.; Sugiyama, C.; Takabayashi, F. Improvement effects of wasabi (Wasabia japonica) leaves and allyl isothiocyanate on stomach lesions of mongolian gerbils infected with Helicobacter pylori. Nat. Prod. Commun. 2017, 12, 595–598. [Google Scholar] [PubMed]
- Fahey, J.W.; Stephenson, K.K.; Wade, K.L.; Talalay, P. Urease from Helicobacter pylori is inactivated by sulforaphane and other isothiocyanates. Biochem. Biophys. Res. Commun. 2013, 435, 1–7. [Google Scholar] [CrossRef] [Green Version]
- El-Shinnawy, N.A.; Abd-Elmageid, S.A.; Alshailabi, E.M. Evaluation of antiulcer activity of indole-3-carbinol and/or omeprazole on aspirin-induced gastric ulcer in rats. Toxicol. Ind. Health 2014, 30, 357–375. [Google Scholar] [CrossRef]
- Li, Q.; Hu, X.; Xuan, Y.; Ying, J.; Fei, Y.; Rong, J.; Zhang, Y.; Zhang, J.; Liu, C.; Liu, Z. Kaempferol protects ethanol-induced gastric ulcers in mice via pro-inflammatory cytokines and NO. Acta Biochim. Biophys. Sin. 2018, 50, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Alkushi, A.G.R.; Elsawy, N.A.M. Quercetin attenuates, indomethacin-induced acute gastric ulcer in rats. Folia Morphol. Warsz 2017, 76, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Min, Y.S.; Yim, S.H.; Bai, K.L.; Choi, H.J.; Jeong, J.H.; Song, H.J.; Park, S.Y.; Ham, I.; Whang, W.K.; Sohn, U.D. The effects of apigenin-7-O-beta-D-glucuronopyranoside on reflux oesophagitis and gastritis in rats. Auton. Autacoid Pharmacol. 2005, 25, 85–91. [Google Scholar] [CrossRef]
- Wallace, J.L. Prostaglandins, NSAIDs, and gastric mucosal protection: Why doesn’t the stomach digest itself? Physiol. Rev. 2008, 88, 1547–1565. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, S.; Anandan, R.; Sini, T.K.; Mathew, P.T. Protective effect of glucosamine against ibuprofen-induced peptic ulcer in rats. J. Gastroenterol. Hepatol. 2007, 22, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.V.; Ganguly, K.; Paul, S.; Maulik, N.; Swarnakar, S. Curcumin heals indomethacin-induced gastric ulceration by stimulation of angiogenesis and reconstitution of collagen fiber via VEGF and MMP-2 mediated signaling. Antioxid. Redox Signal. 2012, 16, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, D.A.; Burns, G.P. The effect of acetic acid on gastric secretion stimulated by endogenous gastrin in vagally denervated canine fundic pouches. Am. J. Dig. Dis. 1973, 18, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Engevik, A.C.; Kaji, I.; Goldenring, J.R. The physiology of the gastric parietal cell. Physiol. Rev. 2020, 100, 573–602. [Google Scholar] [CrossRef]
- Barocelli, E.; Ballabeni, V. Histamine in the control of gastric acid secretion: A topic review. Pharmacol. Res. 2003, 47, 299–304. [Google Scholar] [CrossRef]
- Carlson, M.; Raab, Y.; Seveus, L.; Xu, S.; Hallgren, R.; Venge, P. Human neutrophil lipocalin is a unique marker of neutrophil inflammation in ulcerative colitis and proctitis. Gut 2002, 50, 501–506. [Google Scholar] [CrossRef]
- Sokolova, O.; Naumann, M. NF-κB Signaling in Gastric Cancer. Toxins 2017, 9, 119. [Google Scholar] [CrossRef]
- Kim, J.K.; Lee, J.E.; Jung, E.H.; Jung, J.Y.; Jung, D.H.; Ku, S.K.; Cho, I.J.; Kim, S.C. Hemistepsin A ameliorates acute inflammation in macrophages via inhibition of nuclear factor-κB and activation of nuclear factor erythroid 2-related factor 2. Food Chem. Toxicol. 2018, 111, 176–188. [Google Scholar] [CrossRef]
- Grandjean-Laquerriere, A.; Antonicelli, F.; Gangloff, S.C.; Guenounou, M.; Le Naour, R. UVB-induced IL-18 production in human keratinocyte cell line NCTC 2544 through NF-kappaB activation. Cytokine 2007, 37, 76–83. [Google Scholar] [CrossRef]
- Suzuki, H.; Nishizawa, T.; Tsugawa, H.; Mogami, S.; Hibi, T. Roles of oxidative stress in stomach disorders. J. Clin. Biochem. Nutr. 2012, 50, 35–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Representative chromatogram after eluting vitamin U (left) and BOE (right). BOE, B. oleracea var. capitata L. extract.
Figure 1.
Representative chromatogram after eluting vitamin U (left) and BOE (right). BOE, B. oleracea var. capitata L. extract.

Figure 2.
Macroscopic effect of BOE on H/E-mediated gastric damage. (a) Representative gross stomach images. Scale bars indicate 6.5 mm. (b) Macroscopically visible lesion on the gastric mucosa was quantified as a gross lesion area (mm2). Numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; Car., cardiac region of the stomach; Fun., fundus/corpus region of the stomach; H/E, HCl/ethanol; Pyl., pylorus region of the stomach; RA, ranitidine hydrochloride.
Figure 2.
Macroscopic effect of BOE on H/E-mediated gastric damage. (a) Representative gross stomach images. Scale bars indicate 6.5 mm. (b) Macroscopically visible lesion on the gastric mucosa was quantified as a gross lesion area (mm2). Numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; Car., cardiac region of the stomach; Fun., fundus/corpus region of the stomach; H/E, HCl/ethanol; Pyl., pylorus region of the stomach; RA, ranitidine hydrochloride.

Figure 3.
Microscopic effect of BOE on H/E-mediated gastric damage. (a) Representative histological image of the fundus/corpus mucosa after staining with hematoxylin and eosin. Dashed squares of left column are enlarged in the middle and right column, and scale bars indicate 80 μm. (b) Histomorphometric analyses. Invaded percentage of lesion, mucosa thickness of peri-ulcerative region, and semi-quantitative histological score were calculated as described in the method section. Numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; ED, edematous change; EP, surface epithelium; H/E, HCl/ethanol; LU, lumen; ML, muscular layer; MM, muscularis mucosa; MU, mucosa layer; RA, ranitidine hydrochloride; SM, submucosa.
Figure 3.
Microscopic effect of BOE on H/E-mediated gastric damage. (a) Representative histological image of the fundus/corpus mucosa after staining with hematoxylin and eosin. Dashed squares of left column are enlarged in the middle and right column, and scale bars indicate 80 μm. (b) Histomorphometric analyses. Invaded percentage of lesion, mucosa thickness of peri-ulcerative region, and semi-quantitative histological score were calculated as described in the method section. Numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; ED, edematous change; EP, surface epithelium; H/E, HCl/ethanol; LU, lumen; ML, muscular layer; MM, muscularis mucosa; MU, mucosa layer; RA, ranitidine hydrochloride; SM, submucosa.

Figure 4.
Effect of BOE on H/E-mediated mucus loss. (a) Alcian blue binding assay was used to determine mucus content in the gastric tissue. The levels of total hexose, sialic acid (b), and collagen (c) were measured using a gastric tissue homogenate. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; H/E, HCl/ethanol; RA, ranitidine hydrochloride.
Figure 4.
Effect of BOE on H/E-mediated mucus loss. (a) Alcian blue binding assay was used to determine mucus content in the gastric tissue. The levels of total hexose, sialic acid (b), and collagen (c) were measured using a gastric tissue homogenate. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; H/E, HCl/ethanol; RA, ranitidine hydrochloride.

Figure 5.
Effect of BOE on histamine and mRNAs expression associated with secreting gastric acid. (a) Plasma histamine was quantified using a commercial ELISA kit. (b) mRNA expression associated with secreting gastric acid was quantified via real-time PCR. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; CCK2R, cholecystokinin-2 receptor; H/E, HCl/ethanol; H2R, histamine H2 receptor; RA, ranitidine hydrochloride.
Figure 5.
Effect of BOE on histamine and mRNAs expression associated with secreting gastric acid. (a) Plasma histamine was quantified using a commercial ELISA kit. (b) mRNA expression associated with secreting gastric acid was quantified via real-time PCR. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; CCK2R, cholecystokinin-2 receptor; H/E, HCl/ethanol; H2R, histamine H2 receptor; RA, ranitidine hydrochloride.

Figure 6.
Effect of BOE on H/E-mediated inflammation in the gastric tissue. (a) Hydrogen peroxide and o-dianisidine dihydrochloride were used as substrates for detecting MPO activity in the gastric tissue. mRNA levels of NF-κB and pro-inflammatory enzymes (b) and (c) and protein levels of pro-inflammatory cytokines (d) were determined by real-time PCR and ELISA, respectively. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; COX-2, cyclooxygenase-2; H/E, HCl/ethanol; IL, interleukin; iNOS, inducible nitric oxide synthase; MPO, myeloperoxidase; NF-κB, nuclear factor-κB; RA, ranitidine hydrochloride; TNF, tumor necrosis factor.
Figure 6.
Effect of BOE on H/E-mediated inflammation in the gastric tissue. (a) Hydrogen peroxide and o-dianisidine dihydrochloride were used as substrates for detecting MPO activity in the gastric tissue. mRNA levels of NF-κB and pro-inflammatory enzymes (b) and (c) and protein levels of pro-inflammatory cytokines (d) were determined by real-time PCR and ELISA, respectively. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; COX-2, cyclooxygenase-2; H/E, HCl/ethanol; IL, interleukin; iNOS, inducible nitric oxide synthase; MPO, myeloperoxidase; NF-κB, nuclear factor-κB; RA, ranitidine hydrochloride; TNF, tumor necrosis factor.

Figure 7.
Effect of BOE on H/E-mediated oxidative stress in the gastric tissue. (a) The level of malondialdehyde in the gastric tissue was measured as a marker of lipid peroxidation. (b) Activities for catalase and superoxide dismutase were determined using hydrogen peroxide and nitroblue tetrazolium, respectively. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; H/E, HCl/ethanol; RA, ranitidine hydrochloride.
Figure 7.
Effect of BOE on H/E-mediated oxidative stress in the gastric tissue. (a) The level of malondialdehyde in the gastric tissue was measured as a marker of lipid peroxidation. (b) Activities for catalase and superoxide dismutase were determined using hydrogen peroxide and nitroblue tetrazolium, respectively. All numerical results are presented as mean ± SD of ten mice (** p < 0.01 between the vehicle and H/E groups; ## p < 0.01, # p < 0.05 versus H/E-administered group). BOE, B. oleracea var. capitata L. extract; H/E, HCl/ethanol; RA, ranitidine hydrochloride.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, M.-R.; Kim, T.-I.; Choi, B.-R.; Kim, M.B.; Cho, I.J.; Lee, K.-W.; Ku, S.K. Brassica oleracea Prevents HCl/Ethanol-Induced Gastric Damages in Mice. Appl. Sci. 2021, 11, 16. https://doi.org/10.3390/app11010016
AMA Style
Kim M-R, Kim T-I, Choi B-R, Kim MB, Cho IJ, Lee K-W, Ku SK. Brassica oleracea Prevents HCl/Ethanol-Induced Gastric Damages in Mice. Applied Sciences. 2021; 11(1):16. https://doi.org/10.3390/app11010016
Chicago/Turabian StyleKim, Mi-Ryung, Tae-Il Kim, Beom-Rak Choi, Moon Bong Kim, Il Je Cho, Keun-Woo Lee, and Sae Kwang Ku. 2021. "Brassica oleracea Prevents HCl/Ethanol-Induced Gastric Damages in Mice" Applied Sciences 11, no. 1: 16. https://doi.org/10.3390/app11010016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.