Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Why an Endothelial Model?
3. Umbilical Cord and HUVEC
3.1. Characteristics of Umbilical Cord
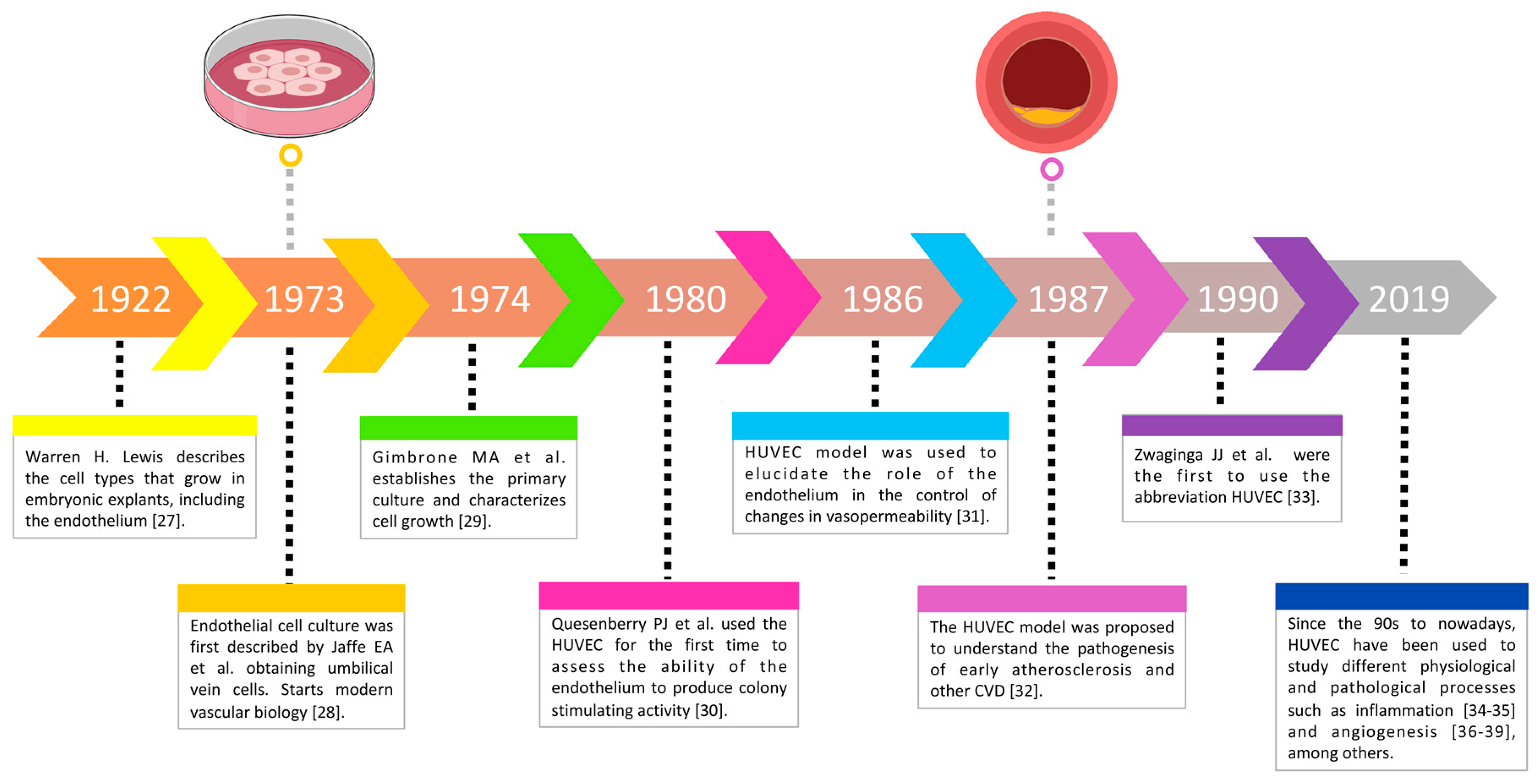
3.2. The HUVEC Model through Time
3.3. Isolation and Culture of HUVEC
3.3.1. First HUVEC Isolation Techniques
3.3.2. HUVEC Cultures
3.3.3. Isolation and Culture of HUVEC Conditions
Umbilical Cord Collection and Conservation
HUVEC Culture Considerations
- a)
- Heat the sterile transport buffer in a water bath at 37 °C. This will prevent thermal shock disabling cell growth.
- b)
- Prepare a sterile area in a laminar flow hood. This will prevent bacterial and fungal contamination of both reagents and cultures.
- c)
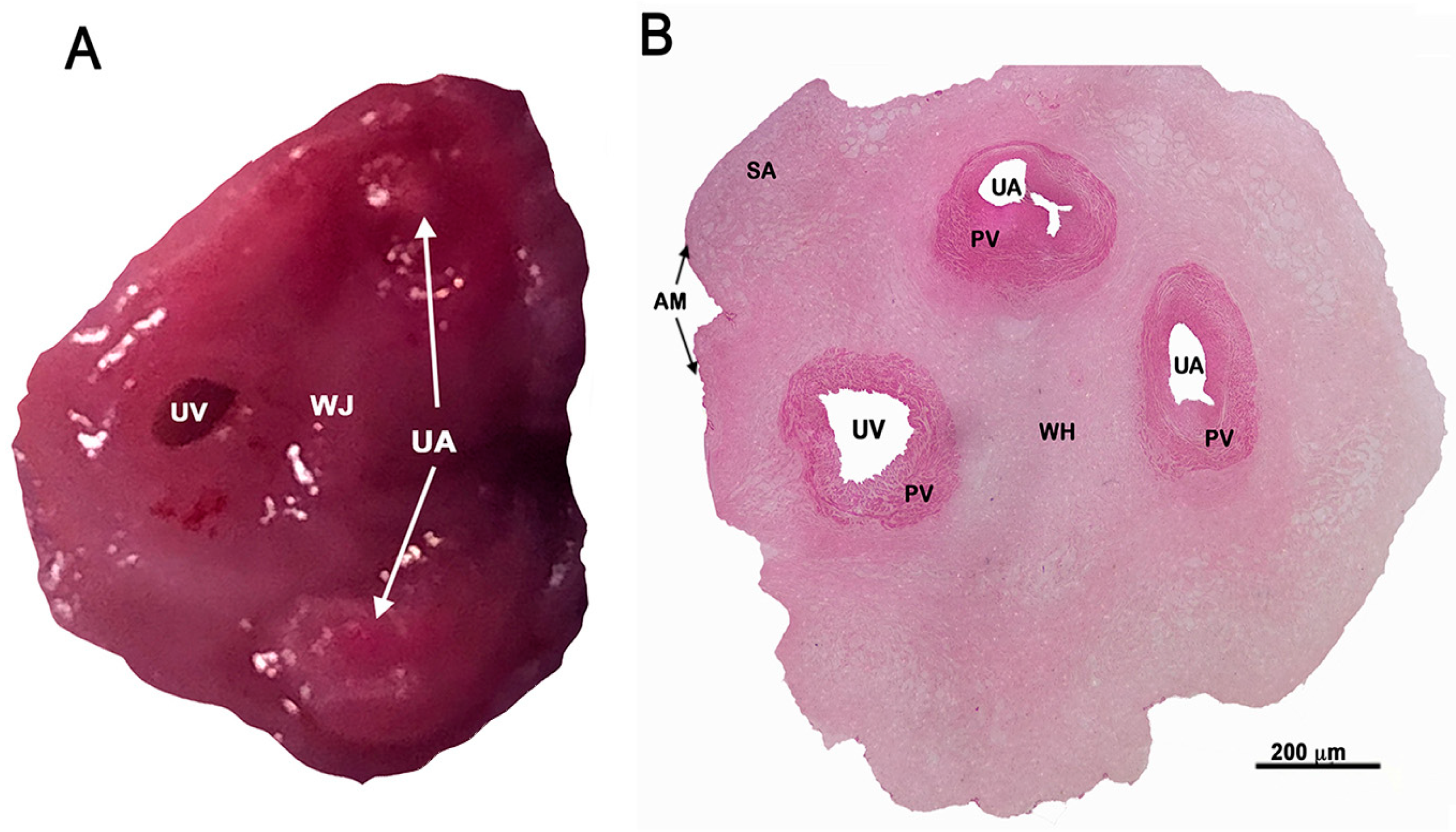
- Before starting the isolation procedure, it is desirable to identify the human umbilical vein. Umbilical arteries are more rigid and difficult to dilate (Figure 1A,B).
- d)
- It is recommended to store collagenase in 10–12 mL aliquots at -20 °C to avoid denaturation.
- e)
- The culture flasks must be previously covered with gelatin or fibronectin to ensure cell adhesion.
3.4. Advantages and Disadvantages of the HUVEC Model
4. HUVEC Identification
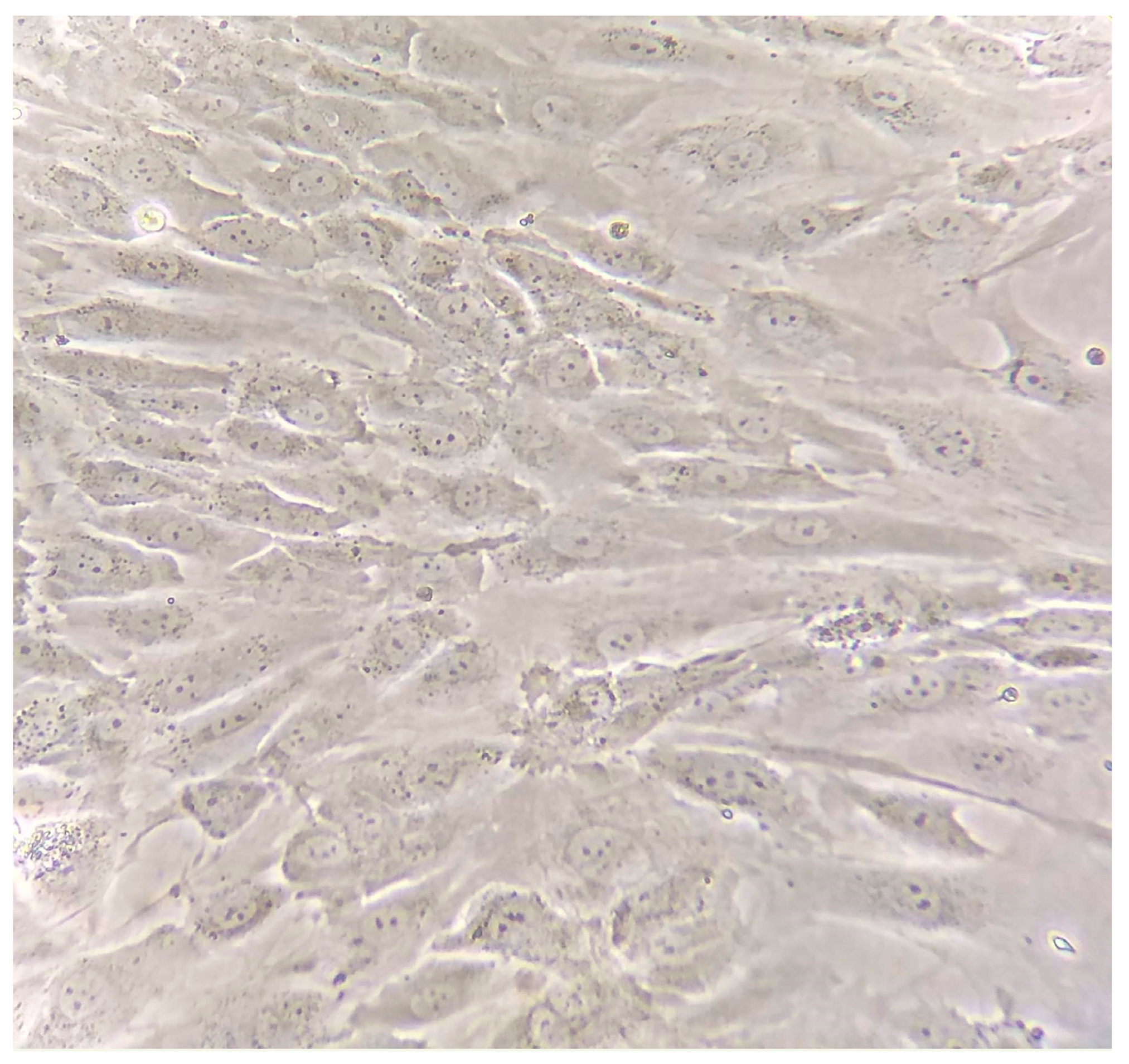
4.1. Morphological Description
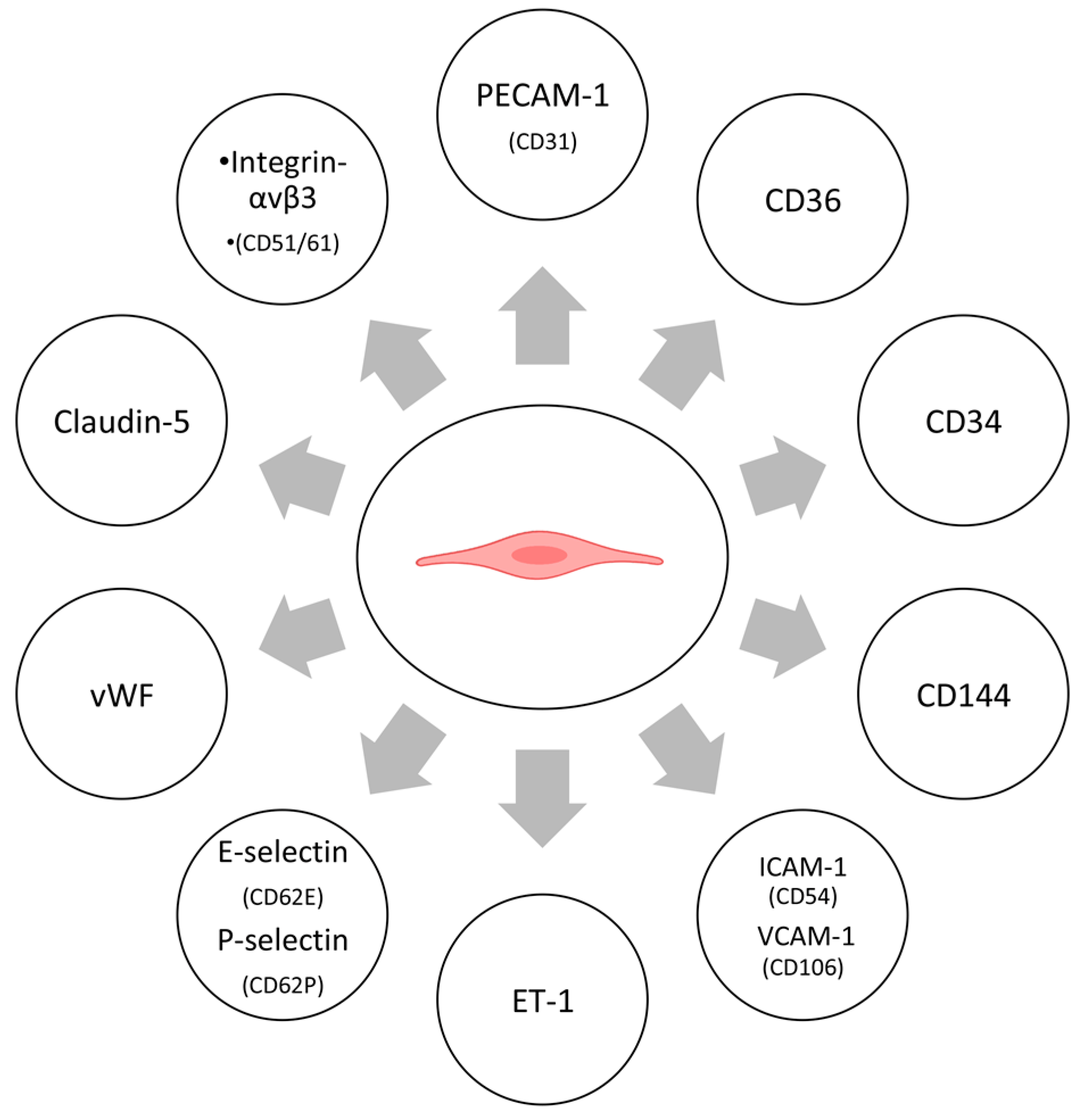
4.2. Cellular Markers
4.2.1. Protein Markers
4.2.2. Enzymes
4.2.3. Other Markers
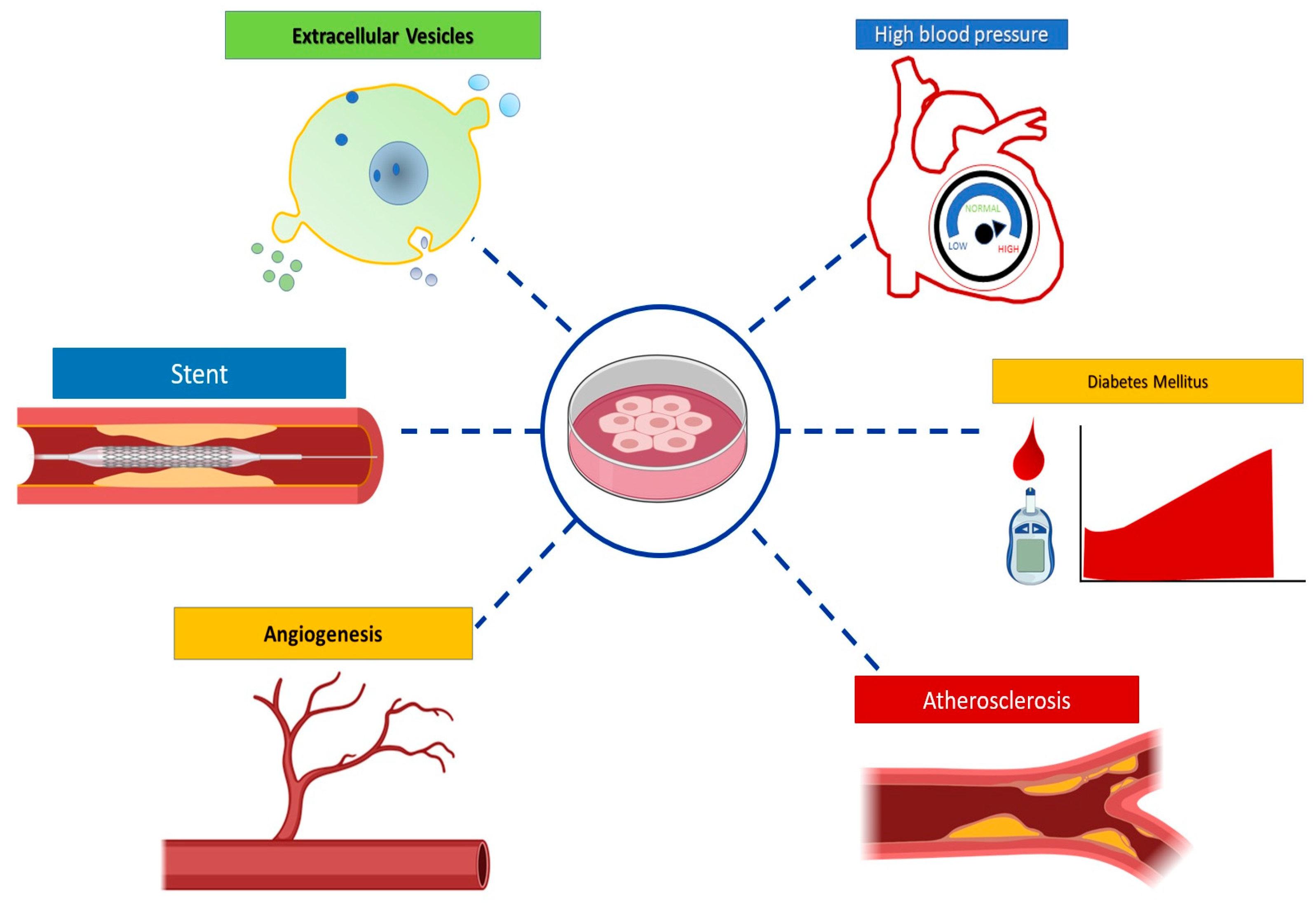
5. HUVEC Model Applications
5.1. Cardiovascular and Metabolic Diseases
5.1.1. Hypertension
5.1.2. Atherosclerosis
5.1.3. Hypercholesterolemia
5.1.4. Diabetes Mellitus
5.2. Angiogenesis
5.3. Other applications
5.3.1. Extracellular Vesicles
5.3.2. Stent Use
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Soler-Botija, C.; Galvez-Monton, C.; Bayes-Genis, A. Epigenetic Biomarkers in Cardiovascular Diseases. Front. Genet. 2019, 10, 950. [Google Scholar] [CrossRef] [PubMed]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting Early Atherosclerosis: A Focus on Oxidative Stress and Inflammation. Oxid. Med. Cell Longev. 2019, 2019, 8563845. [Google Scholar] [CrossRef]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgozoglu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Maamoun, H.; Abdelsalam, S.S.; Zeidan, A.; Korashy, H.M.; Agouni, A. Endoplasmic Reticulum Stress: A Critical Molecular Driver of Endothelial Dysfunction and Cardiovascular Disturbances Associated with Diabetes. Int. J. Mol. Sci. 2019, 20, 1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vion, A.C.; Ramkhelawon, B.; Loyer, X.; Chironi, G.; Devue, C.; Loirand, G.; Tedgui, A.; Lehoux, S.; Boulanger, C.M. Shear stress regulates endothelial microparticle release. Circ. Res. 2013, 112, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Hou, X.; Yang, S. microRNA-338-3p promotes ox-LDL-induced endothelial cell injury through targeting BAMBI and activating TGF-β/Smad pathway. J. Cell. Physiol. 2019, 234, 11577–11586. [Google Scholar] [CrossRef] [PubMed]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm J. 2015, 23, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Barre-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Future Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef] [Green Version]
- MacGowan, A.; Rogers, C.; Bowker, K. In vitro models, in vivo models, and pharmacokinetics: What can we learn from in vitro models? Clin. Infect. Dis. 2001, 33, S214–S220. [Google Scholar] [CrossRef]
- Roudsari, L.C.; West, J.L. Studying the influence of angiogenesis in in vitro cancer model systems. Adv. Drug Deliv. Rev. 2016, 97, 250–259. [Google Scholar] [CrossRef]
- Baudin, B.; Bruneel, A.; Bosselut, N.; Vaubourdolle, M. A protocol for isolation and culture of human umbilical vein endothelial cells. Nat. Protoc. 2007, 2, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.S.; Tsai, W.P.; Chen, Z.Q.; Chen, Z.; Van Waes, C.; Burger, A.M.; Lautenberger, J.A. A human vascular endothelial cell model to study angiogenesis and tumorigenesis. Carcinogenesis 1998, 19, 673–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onat, D.; Brillon, D.; Colombo, P.C.; Schmidt, A.M. Human vascular endothelial cells: A model system for studying vascular inflammation in diabetes and atherosclerosis. Curr. Diab. Rep. 2011, 11, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleger, C.; Platz, S.J.; Deschl, U. Development of an in vitro model for vascular injury with human endothelial cells. ALTEX 2004, 21 (Suppl 3), 12–19. [Google Scholar]
- Fearon, I.M.; Gaca, M.D.; Nordskog, B.K. In vitro models for assessing the potential cardiovascular disease risk associated with cigarette smoking. Toxicol. In Vitro 2013, 27, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.M. Establishment of the Basic Embryonic Body Plan. In Human Embryology and Developmental Biology, 5th ed.; Carlson, B.M., Ed.; Elsevier: Philadelphia, PA, USA, 2014; pp. 111–115. [Google Scholar]
- Can, A.; Karahuseyinoglu, S. Concise review: Human umbilical cord stroma with regard to the source of fetus-derived stem cells. Stem Cells 2007, 25, 2886–2895. [Google Scholar] [CrossRef]
- Kim, D.W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.D.; Borlongan, C.V. Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. Int. J. Mol. Sci 2013, 14, 11692–11712. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Kubota, T.; Aso, T. Study on myofibroblast differentiation in the stromal cells of Wharton’s jelly: Expression and localization of alpha-smooth muscle actin. Early Hum. Dev. 1998, 51, 223–233. [Google Scholar] [CrossRef]
- Weiss, M.L.; Medicetty, S.; Bledsoe, A.R.; Rachakatla, R.S.; Choi, M.; Merchav, S.; Luo, Y.; Rao, M.S.; Velagaleti, G.; Troyer, D. Human umbilical cord matrix stem cells: Preliminary characterization and effect of transplantation in a rodent model of Parkinson’s disease. Stem Cells 2006, 24, 781–792. [Google Scholar] [CrossRef]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, M.; Suga, Y.; Sanmano, B.; Ikeda, S.; Ogawa, H. Organotypic culture and surface plantation using umbilical cord epithelial cells: Morphogenesis and expression of differentiation markers mimicking cutaneous epidermis. J. Dermatol. Sci. 2004, 35, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem cell characteristics of amniotic epithelial cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzon, I.; Alfonso-Rodriguez, C.A.; Martinez-Gomez, C.; Carriel, V.; Martin-Piedra, M.A.; Fernandez-Valades, R.; Sanchez-Quevedo, M.C.; Alaminos, M. Expression of epithelial markers by human umbilical cord stem cells. A topographical analysis. Placenta 2014, 35, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, R.R.; Cetrulo, K.J.; Cetrulo, C.L. Wharton’s Jelly stem cells: Future clinical applications. Placenta 2011, 32 (Suppl 4), S311–S315. [Google Scholar] [CrossRef]
- Schultheiss, D.; Bloom, D.A.; Wefer, J.; Jonas, U. Tissue engineering from Adam to the zygote: Historical reflections. World J. Urol. 2000, 18, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, W.H. Endothelium in tissue cultures. Am. J. Anat. 1922, 30, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E. Culture of human endothelial cells derived from umbilical veins. J. Clin. Invest. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; Cotran, R.S.; Folkman, J. Human vascular endothelial cells in culture. Growth and DNA synthesis. J. Cell Biol. 1974, 60, 673–684. [Google Scholar] [CrossRef]
- Quesenberry, P.J.; Gimbrone, M.J. Vascular endothelium as a regulator of granulopoiesis: Production of colony-stimulating activity by cultured human endothelial cells. Blood 1980, 56, 1060–1067. [Google Scholar] [CrossRef]
- Killackey, J.J.; Johnston, M.G.; Movat, H.Z. Increased permeability of microcarrier-cultured endothelial monolayers in response to histamine and thrombin. A model for the in vitro study of increased vasopermeability. Am. J. Pathol 1986, 122, 50–61. [Google Scholar]
- Vaccaro, P.S.; Joseph, L.B.; Titterington, L.; Stephens, R.E. Methods for the initiation and maintenance of human endothelial cell culture. Vasc. Surg. 1987, 21, 391–400. [Google Scholar] [CrossRef]
- Zwaginga, J.; Sixma, J.; de Groot, P.G. Activation of endothelial cells induces platelet thrombus formation on their matrix. Studies of new in vitro thrombosis model with low molecular weight heparin as anticoagulant. Arterioscler. An. Off. J. Am. Heart Assoc. Inc. 1990, 10, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Koenig, A.; Yakisan, E.; Reuter, M.; Huang, M.; Sykora, K.W.; Corbacioglu, S.; Welte, K. Differential regulation of stem cell factor mRNA expression in human endothelial cells by bacterial pathogens: An in vitro model of inflammation. Blood 1994, 83, 2836–2843. [Google Scholar] [CrossRef] [Green Version]
- Waldman, W.J.; Knight, D.A.; Huang, E.H. An in vitro model of T cell activation by autologous cytomegalovirus (CMV)-infected human adult endothelial cells: Contribution of CMV-enhanced endothelial ICAM-1. J. Immunol. 1998, 160, 3143–3151. [Google Scholar] [PubMed]
- Lee, O.H.; Bae, S.K.; Bae, M.H.; Lee, Y.M.; Moon, E.J.; Cha, H.J.; Kwon, Y.G.; Kim, K.W. Identification of angiogenic properties of insulin-like growth factor II in in vitro angiogenesis models. Br. J. Cancer 2000, 82, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vailhe, B.; Vittet, D.; Feige, J.J. In vitro models of vasculogenesis and angiogenesis. Lab. Invest. 2001, 81, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Starke, R.D.; Ferraro, F.; Paschalaki, K.E.; Dryden, N.H.; McKinnon, T.A.; Sutton, R.E.; Payne, E.M.; Haskard, D.O.; Hughes, A.D.; Cutler, D.F.; et al. Endothelial von Willebrand factor regulates angiogenesis. Blood 2011, 117, 1071–1080. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, B.S.; Lee, J. Evaluation of the angiogenic potency of a novel exopolysaccharide produced by the MK1 bacterial strain. Arch. Pharm. Res. 2016, 39, 1223–1231. [Google Scholar] [CrossRef]
- Maruyama, Y. The human endothelial cell in tissue culture. Z Zellforsch Mikrosk Anat 1963, 60, 69–79. [Google Scholar] [CrossRef]
- Henriksen, T.; Evensen, S.A.; Elgjo, R.F.; Vefling, A. Human fetal endothelial cells in culture. Scand. J. Haematol. 1975, 14, 233–241. [Google Scholar] [CrossRef]
- Marin, V.; Kaplanski, G.; Gres, S.; Farnarier, C.; Bongrand, P. Endothelial cell culture: Protocol to obtain and cultivate human umbilical endothelial cells. J. Immunol. Methods 2001, 254, 183–190. [Google Scholar] [CrossRef]
- Cheung, A.L. Isolation and culture of human umbilical vein endothelial cells (HUVEC). Curr. Protoc. Microbiol 2007, 4, A.4B.1–A.4B.8. [Google Scholar] [CrossRef]
- Lopez-Macay, A.; Ruiz-Medina, E.J.; Ventura-Gallegos, J.L.; Arechavaleta-Velasco, F.; Gomez-Quiroz, L.E.; Konigsberg-Fainstein, M.; Zentella-Dehesa, A. Characterization of hemodynamic ex vivo model to study endothelial activation by TNF-alpha in prefunded human veins. Gac Med. Mex 2015, 151, 206–215. [Google Scholar] [PubMed]
- Weibel, E.R.; Palade, G.E. New Cytoplasmic Components in Arterial Endothelia. J. Cell Biol. 1964, 23, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K. Human endothelial cells in culture. Arch. Histol. Jpn. 1979, 42, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haudenschild, C.C.; Cotran, R.S.; Gimbrone, M.A., Jr.; Folkman, J. Fine structure of vascular endothelium in culture. J. Ultrastruct. Res. 1975, 50, 22–32. [Google Scholar] [CrossRef]
- Elgjo, R.F.; Henriksen, T.; Evensen, S.A. Ultrastructural identification of umbilical cord vein endothelium in situ and in culture. Cell Tissue Res. 1975, 162, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Bachetti, T.; Morbidelli, L. Endothelial cells in culture: A model for studying vascular functions. Pharmacol. Res. 2000, 42, 9–19. [Google Scholar] [CrossRef]
- Larrivée, B.; Karsan, A. Isolation and culture of primary endothelial cells. In Basic Cell Culture Protocols; Helgason, C.D., Miller, C.L., Eds.; Springer: Basel, Switzerland, 2005; pp. 315–329. [Google Scholar]
- Shao, C.; Chen, J.; Chen, P.; Zhu, M.; Yao, Q.; Gu, P.; Fu, Y.; Fan, X. Targeted transplantation of human umbilical cord blood endothelial progenitor cells with immunomagnetic nanoparticles to repair corneal endothelium defect. Stem Cells Dev. 2015, 24, 756–767. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, H.; Wang, F.; Li, L.; Deng, M.; Huang, L.; Zhao, X. Transplantation of cryopreserved human umbilical cord blood-derived endothelial progenitor cells induces recovery of carotid artery injury in nude rats. Stem Cell Res. Ther. 2015, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Qiu, Q.; Zeng, S.; Xiao, Y.; Shi, M.; Zou, Y.; Ye, Y.; Liang, L.; Yang, X.; Xu, H. Niclosamide inhibits the inflammatory and angiogenic activation of human umbilical vein endothelial cells. Inflamm. Res. 2015, 64, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Borges, L.E.; Bloise, E.; Dela Cruz, C.; Galleri, L.; Apa, R.; Petraglia, F.; Reis, F.M. Urocortin 1 expression and secretion by human umbilical vein endothelial cells: In vitro effects of interleukin 8, interferon gamma, lipopolysaccharide, endothelin 1, prostaglandin F-2alpha, estradiol, progesterone and dexamethasone. Peptides 2015, 74, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Rapp, B.M.; Saadatzedeh, M.R.; Ofstein, R.H.; Bhavsar, J.R.; Tempel, Z.S.; Moreno, O.; Morone, P.; Booth, D.A.; Traktuev, D.O.; Dalsing, M.C. Resident endothelial progenitor cells from human placenta have greater vasculogenic potential than circulating endothelial progenitor cells from umbilical cord blood. Cell Med. 2011, 2, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Lie, P.; Miao, T.; Yu, M.; Lu, Q.; Feng, T.; Li, J.; Zu, T.; Liu, X.; Li, H. Conditioned medium from umbilical cord mesenchymal stem cells induces migration and angiogenesis. Mol. Med. Rep. 2015, 12, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahim, N.A.; Leach, L. Temporal studies into attachment, VE-cadherin perturbation, and paracellular migration of human umbilical mesenchymal stem cells across umbilical vein endothelial monolayers. Stem Cells Dev. 2014, 24, 426–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Chapple, S.J.; Patel, B.; Puszyk, W.; Sugden, D.; Yin, X.; Mayr, M.; Siow, R.C.; Mann, G.E. Gestational diabetes mellitus impairs Nrf2-mediated adaptive antioxidant defenses and redox signaling in fetal endothelial cells in utero. Diabetes 2013, 62, 4088–4097. [Google Scholar] [CrossRef] [Green Version]
- Namiki, A.; Brogi, E.; Kearney, M.; Kim, E.A.; Wu, T.; Couffinhal, T.; Varticovski, L.; Isner, J.M. Hypoxia induces vascular endothelial growth factor in cultured human endothelial cells. J. Biol. Chem. 1995, 270, 31189–31195. [Google Scholar] [CrossRef] [Green Version]
- Wisgrill, L.; Muck, M.; Wessely, I.; Berger, A.; Spittler, A.; Forster-Waldl, E.; Sadeghi, K. Endothelial cells of extremely premature infants display impaired immune response after proinflammatory stimulation. Pediatr. Res. 2018, 83, 128–134. [Google Scholar] [CrossRef]
- Nussbaum, C.; Gloning, A.; Pruenster, M.; Frommhold, D.; Bierschenk, S.; Genzel-Boroviczeny, O.; von Andrian, U.H.; Quackenbush, E.; Sperandio, M. Neutrophil and endothelial adhesive function during human fetal ontogeny. J. Leukoc. Biol. 2013, 93, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Illsinger, S.; Janzen, N.; Sander, S.; Bode, J.; Mallunat, L.; Thomasmeyer, R.; Hagebolling, F.; Schmidt, K.H.; Bednarczyk, J.; Vaske, B.; et al. Energy metabolism in umbilical endothelial cells from preterm and term neonates. J. Perinat. Med. 2011, 39, 587–593. [Google Scholar] [CrossRef]
- Lorant, D.E.; Li, W.; Tabatabaei, N.; Garver, M.K.; Albertine, K.H. P-selectin expression by endothelial cells is decreased in neonatal rats and human premature infants. Blood 1999, 94, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, J.; Olave, N.; Ballinger, S.W.; Ambalavanan, N. Vascular Endothelial Mitochondrial Function Predicts Death or Pulmonary Outcomes in Preterm Infants. Am. J. Respir. Crit. Care Med. 2017, 196, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, N.; Krouwer, V.J.; Post, J.A. A new, rapid and reproducible method to obtain high quality endothelium in vitro. Cytotechnology 2013, 65, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirce, E.C. Tissue-culture evaluation of the viability of blood vessels stored by refrigeration. Ann. Surg. 1949, 129, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manconi, F.; Markham, R.; Fraser, I.S. Culturing endothelial cells of microvascular origin. Methods Cell Sci. 2000, 22, 89–99. [Google Scholar] [CrossRef]
- Thornton, S.C.; Mueller, S.N.; Levine, E.M. Human endothelial cells: Use of heparin in cloning and long-term serial cultivation. Science 1983, 222, 623–625. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Metzner, C.; Bhonde, R.R.; Malsch, G.; Schiermeyer, B.; Vetter, H. Simultaneous isolation of endothelial and smooth muscle cells from human umbilical artery or vein and their growth response to low-density lipoproteins. In Vitro Cell. Dev. Biol. Anim. 2002, 38, 265–272. [Google Scholar] [CrossRef]
- Freshney, R.I. Culture of specific cell types. In Culture of Animal Cells: A Manual of Basic Technique; Freshney, R.I., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2005. [Google Scholar]
- Jarrell, B.; Levine, E.; Shapiro, S.; Williams, S.; Carabasi, R.A.; Mueller, S.; Thornton, S. Human adult endothelial cell growth in culture. J. Vasc. Surg. 1984, 1, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Maciag, T.; Hoover, G.A.; Stemerman, M.B.; Weinstein, R. Serial propagation of human endothelial cells in vitro. J. Cell Biol. 1981, 91, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Garbern, J.C.; Mummery, C.L.; Lee, R.T. Model systems for cardiovascular regenerative biology. Cold Spring Harb Perspect Med. 2013, 3, a014019. [Google Scholar] [CrossRef] [Green Version]
- Monti, M.; Terzuoli, E.; Ziche, M.; Morbidelli, L. H2S dependent and independent anti-inflammatory activity of zofenoprilat in cells of the vascular wall. Pharmacol. Res. 2016, 113, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Bouis, D.; Hospers, G.A.; Meijer, C.; Molema, G.; Mulder, N.H. Endothelium in vitro: A review of human vascular endothelial cell lines for blood vessel-related research. Angiogenesis 2001, 4, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Addis, R.; Campesi, I.; Fois, M.; Capobianco, G.; Dessole, S.; Fenu, G.; Montella, A.; Cattaneo, M.G.; Vicentini, L.M.; Franconi, F. Human umbilical endothelial cells (HUVECs) have a sex: Characterisation of the phenotype of male and female cells. Biol. Sex. Differ. 2014, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M.; Blaschke, B.; Benn, A.; Hammer, E.; Witt, E.; Kirwan, J.; Fritsche-Guenther, R.; Gloaguen, Y.; Bartsch, C.; Vietzke, A.; et al. Sex-specific metabolic and functional differences in human umbilical vein endothelial cells from twin pairs. Atherosclerosis 2019, 291, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M.; Koschate, J.; Kaufmann, K.; Kreye, C.; Mertens, M.; Kuebler, W.M.; Baumann, G.; Gossing, G.; Marki, A.; Zakrzewicz, A.; et al. Does cellular sex matter? Dimorphic transcriptional differences between female and male endothelial cells. Atherosclerosis 2015, 240, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lingappan, K. Differential sex-specific effects of oxygen toxicity in human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun. 2017, 486, 431–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staton, C.A.; Reed, M.W.; Brown, N.J. A critical analysis of current in vitro and in vivo angiogenesis assays. Int. J. Exp. Pathol. 2009, 90, 195–221. [Google Scholar] [CrossRef]
- Haudenschild, C.C.; Zahniser, D.; Folkman, J.; Klagsbrun, M. Human vascular endothelial cells in culture. Lack of response to serum growth factors. Exp. Cell Res. 1976, 98, 175–183. [Google Scholar] [CrossRef]
- Folkman, J.; Haudenschild, C.C.; Zetter, B.R. Long-term culture of capillary endothelial cells. Proc. Natl. Acad. Sci. USA 1979, 76, 5217–5221. [Google Scholar] [CrossRef] [Green Version]
- Franke, R.P.; Grafe, M.; Schnittler, H.; Seiffge, D.; Mittermayer, C.; Drenckhahn, D. Induction of human vascular endothelial stress fibres by fluid shear stress. Nature 1984, 307, 648–649. [Google Scholar] [CrossRef]
- Pu, F.R.; Williams, R.L.; Markkula, T.K.; Hunt, J.A. Expression of leukocyte-endothelial cell adhesion molecules on monocyte adhesion to human endothelial cells on plasma treated PET and PTFE in vitro. Biomaterials 2002, 23, 4705–4718. [Google Scholar] [CrossRef]
- Ho, M.; Yang, E.; Matcuk, G.; Deng, D.; Sampas, N.; Tsalenko, A.; Tabibiazar, R.; Zhang, Y.; Chen, M.; Talbi, S.; et al. Identification of endothelial cell genes by combined database mining and microarray analysis. Physiol. Genomics 2003, 13, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Floris, I.; Descamps, B.; Vardeu, A.; Mitic, T.; Posadino, A.M.; Shantikumar, S.; Sala-Newby, G.; Capobianco, G.; Mangialardi, G.; Howard, L.; et al. Gestational Diabetes Mellitus Impairs Fetal Endothelial Cell Functions Through a Mechanism Involving MicroRNA-101 and Histone Methyltransferase Enhancer of Zester Homolog-2. Arterioscl. Throm. Vas. 2015, 35, 664–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albelda, S.M.; Muller, W.A.; Buck, C.A.; Newman, P.J. Molecular and cellular properties of PECAM-1 (endoCAM/CD31): A novel vascular cell-cell adhesion molecule. J. Cell Biol. 1991, 114, 1059–1068. [Google Scholar] [CrossRef]
- Dawson, D.W.; Pearce, S.F.; Zhong, R.; Silverstein, R.L.; Frazier, W.A.; Bouck, N.P. CD36 mediates the In vitro inhibitory effects of thrombospondin-1 on endothelial cells. J. Cell Biol. 1997, 138, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Greenwalt, D.E.; Lipsky, R.H.; Ockenhouse, C.F.; Ikeda, H.; Tandon, N.N.; Jamieson, G.A. Membrane glycoprotein CD36: A review of its roles in adherence, signal transduction, and transfusion medicine. Blood 1992, 80, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Nickoloff, B.J. The human progenitor cell antigen (CD34) is localized on endothelial cells, dermal dendritic cells, and perifollicular cells in formalin-fixed normal skin, and on proliferating endothelial cells and stromal spindle-shaped cells in Kaposi’s sarcoma. Arch. Dermatol. 1991, 127, 523–529. [Google Scholar] [CrossRef]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical expression of endothelial markers CD31, CD34, von Willebrand factor, and Fli-1 in normal human tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Keiper, T.; Santoso, S.; Nawroth, P.; Orlova, V.; Chavakis, T. The role of junctional adhesion molecules in cell-cell interactions. Histol. Histopathol. 2005, 20, 197–203. [Google Scholar]
- Doan, C.C.; Le, T.L.; Hoang, N.S.; Doan, N.T.; Le, V.D.; Do, M.S. Differentiation of umbilical cord lining membrane-derived mesenchymal stem cells into endothelial-like cells. Iran. Biomed. J. 2014, 18, 67. [Google Scholar]
- Steffen, B.J.; Butcher, E.C.; Engelhardt, B. Evidence for involvement of ICAM-1 and VCAM-1 in lymphocyte interaction with endothelium in experimental autoimmune encephalomyelitis in the central nervous system in the SJL/J mouse. Am. J. Pathol 1994, 145, 189–201. [Google Scholar] [PubMed]
- Davies, M.J.; Gordon, J.L.; Gearing, A.J.; Pigott, R.; Woolf, N.; Katz, D.; Kyriakopoulos, A. The expression of the adhesion molecules ICAM-1, VCAM-1, PECAM, and E-selectin in human atherosclerosis. J. Pathol. 1993, 171, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Wagner, O.F.; Christ, G.; Wojta, J.; Vierhapper, H.; Parzer, S.; Nowotny, P.J.; Schneider, B.; Waldhausl, W.; Binder, B.R. Polar secretion of endothelin-1 by cultured endothelial cells. J. Biol. Chem. 1992, 267, 16066–16068. [Google Scholar] [PubMed]
- Dong, Z.M.; Chapman, S.M.; Brown, A.A.; Frenette, P.S.; Hynes, R.O.; Wagner, D.D. The combined role of P- and E-selectins in atherosclerosis. J. Clin. Invest. 1998, 102, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Asa, D.; Raycroft, L.; Ma, L.; Aeed, P.A.; Kaytes, P.S.; Elhammer, A.P.; Geng, J.-G. The P-selectin glycoprotein ligand functions as a common human leukocyte ligand for P-and E-selectins. J. Biol. Chem. 1995, 270, 11662–11670. [Google Scholar] [CrossRef] [Green Version]
- Rice, J.W.; Davis, J.E.; Crowl, R.M.; Johnston, P.A. Development of a high volume screen to identify inhibitors of endothelial cell activation. Anal. Biochem 1996, 241, 254–259. [Google Scholar] [CrossRef]
- Kluger, M.S.; Clark, P.R.; Tellides, G.; Gerke, V.; Pober, J.S. Claudin-5 controls intercellular barriers of human dermal microvascular but not human umbilical vein endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial claudin: Claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J. Cell Biol. 1999, 147, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Voyta, J.C.; Via, D.P.; Butterfield, C.E.; Zetter, B.R. Identification and isolation of endothelial cells based on their increased uptake of acetylated-low density lipoprotein. J. Cell Biol. 1984, 99, 2034–2040. [Google Scholar] [CrossRef] [Green Version]
- Pober, J.S.; Sessa, W.C. Evolving functions of endothelial cells in inflammation. Nat. Rev. Immunol. 2007, 7, 803–815. [Google Scholar] [CrossRef]
- Birukova, A.A.; Zagranichnaya, T.; Fu, P.; Alekseeva, E.; Chen, W.; Jacobson, J.R.; Birukov, K.G. Prostaglandins PGE2 and PGI2 promote endothelial barrier enhancement via PKA-and Epac1/Rap1-dependent Rac activation. Exp. Cell Res. 2007, 313, 2504–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Graham, J.; Kahn, J.W.; Schwartz, E.A.; Gerritsen, M.E. Functional roles for PECAM-1 (CD31) and VE-cadherin (CD144) in tube assembly and lumen formation in three-dimensional collagen gels. Am. J. Pathol. 1999, 155, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Jain, S.; Kelemen, S.E.; Autieri, M.V. AIF-1 expression regulates endothelial cell activation, signal transduction, and vasculogenesis. Am. J. Physiol. Cell Physiol. 2009, 296, C256–C266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takase, H.; Matsumoto, K.; Yamadera, R.; Kubota, Y.; Otsu, A.; Suzuki, R.; Ishitobi, H.; Mochizuki, H.; Kojima, T.; Takano, S.; et al. Genome-wide identification of endothelial cell-enriched genes in the mouse embryo. Blood 2012, 120, 914–923. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Song, H.; Zhang, Y.; Zhang, Y.; Mu, Q.; Jiang, M.; Wang, F.; Zhang, W.; Li, L.; Li, H.; et al. Irisin Induces Angiogenesis in Human Umbilical Vein Endothelial Cells In Vitro and in Zebrafish Embryos In Vivo via Activation of the ERK Signaling Pathway. PLoS ONE 2015, 10, e0134662. [Google Scholar] [CrossRef] [Green Version]
- Fiordelisi, A.; Iaccarino, G.; Morisco, C.; Coscioni, E.; Sorriento, D. NFkappaB is a Key Player in the Crosstalk between Inflammation and Cardiovascular Diseases. Int. J. Mol. Sci. 2019, 20, 1599. [Google Scholar] [CrossRef] [Green Version]
- Hoare, D.; Bussooa, A.; Neale, S.; Mirzai, N.; Mercer, J. The Future of Cardiovascular Stents: Bioresorbable and Integrated Biosensor Technology. Adv. Sci. (Weinh) 2019, 6, 1900856. [Google Scholar] [CrossRef] [Green Version]
- Kruger-Genge, A.; Blocki, A.; Franke, R.P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.G.; Yi, F.F.; Cheng, W.W.; Qu, X.; Wang, C.T. Molecular mechanisms in vascular injury induced by hypertension: Expression and role of microRNA-34a. Exp. Ther. Med. 2017, 14, 5497–5502. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, W.; Guo, Z.; Zhang, J.; Yu, H.; Liu, B. Mechanisms of lncRNA/microRNA interactions in angiogenesis. Life Sci. 2019, 116900. [Google Scholar] [CrossRef]
- Churov, A.; Summerhill, V.; Grechko, A.; Orekhova, V.; Orekhov, A. MicroRNAs as Potential Biomarkers in Atherosclerosis. Int J. Mol. Sci. 2019, 20, 5547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriegel, A.J.; Baker, M.A.; Liu, Y.; Liu, P.; Cowley, A.W., Jr.; Liang, M. Endogenous microRNAs in human microvascular endothelial cells regulate mRNAs encoded by hypertension-related genes. Hypertension 2015, 66, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzinari, T.C.; Oishi, J.C.; De Moraes, T.F.; Vatanabe, I.P.; Selistre-de-Araujo, H.S.; Pestana, C.R.; Rodrigues, G.J. Treatment with sodium nitroprusside improves the endothelial function in aortic rings with endothelial dysfunction. Eur. J. Pharm. Sci. 2017, 105, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; He, Y.; Yang, M.; Sun, H.; Zhang, S.; Wang, C. Safflor yellow B suppresses angiotensin II-mediated human umbilical vein cell injury via regulation of Bcl-2/p22phox expression. Toxicol. Appl. Pharmacol. 2013, 273, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cruz, E.; Cerezo, A.B.; Cantos-Villar, E.; Richard, T.; Troncoso, A.M.; Garcia-Parrilla, M.C. Inhibition of VEGFR-2 Phosphorylation and Effects on Downstream Signaling Pathways in Cultivated Human Endothelial Cells by Stilbenes from Vitis Spp. J. Agric. Food Chem. 2019, 67, 3909–3918. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Fernandez, B.; Gomez-Bris, R.; Somovilla-Crespo, B.; Gonzalez-Granado, J.M. Immunobiology of Atherosclerosis: A Complex Net of Interactions. Int. J. Mol. Sci. 2019, 20, 5293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.Y.; Chiu, J.J. Atherosclerosis and flow: Roles of epigenetic modulation in vascular endothelium. J. Biomed. Sci 2019, 26, 56. [Google Scholar] [CrossRef]
- Ge, C.; Song, J.; Chen, L.; Wang, L.; Chen, Y.; Liu, X.; Zhang, Y.; Zhang, L.; Zhang, M. Atheroprotective pulsatile flow induces ubiquitin-proteasome-mediated degradation of programmed cell death 4 in endothelial cells. PLoS ONE 2014, 9, e91564. [Google Scholar] [CrossRef]
- Souilhol, C.; Gauci, I.; Feng, S.; Tardajos Ayllon, B.; Mahmoud, M.; Canham, L.; Fragiadaki, M.; Serbanovic-Canic, J.; Ridger, V.; Evans, P.C. Homeobox B9 integrates bone morphogenic protein 4 with inflammation at atheroprone sites. Cardiovasc. Res. 2019, cvz235. [Google Scholar] [CrossRef]
- Krause, M.D.; Huang, R.T.; Wu, D.; Shentu, T.P.; Harrison, D.L.; Whalen, M.B.; Stolze, L.K.; Di Rienzo, A.; Moskowitz, I.P.; Civelek, M.; et al. Genetic variant at coronary artery disease and ischemic stroke locus 1p32.2 regulates endothelial responses to hemodynamics. Proc. Natl. Acad. Sci. USA 2018, 115, E11349–E11358. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, S.; Gale, Z.; Rainger, G.E.; Nash, G.B. Methods for exposing multiple cultures of endothelial cells to different fluid shear stresses and to cytokines, for subsequent analysis of inflammatory function. J. Immunol. Methods 2004, 288, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.H.; Zhang, Y.Y.; Zhang, Y.R.A.; Miao, X.F.; Li, S.; Yang, H.; Ling, Q.J.; Hoffinann, P.R.; Huang, Z. Use of a Mouse Model and Human Umbilical Vein Endothelial Cells to Investigate the Effect of Arsenic Exposure on Vascular Endothelial Function and the Associated Role of Calpains. Environ. Health Persp. 2019, 127, 077003. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, J.; Qi, G. Mechanism of the effect of saikosaponin on atherosclerosis in vitro is based on the MAPK signaling pathway. Mol. Med. Rep. 2017, 16, 8868–8874. [Google Scholar] [CrossRef] [PubMed]
- Gliozzi, M.; Scicchitano, M.; Bosco, F.; Musolino, V.; Carresi, C.; Scarano, F.; Maiuolo, J.; Nucera, S.; Maretta, A.; Paone, S.; et al. Modulation of Nitric Oxide Synthases by Oxidized LDLs: Role in Vascular Inflammation and Atherosclerosis Development. Int. J. Mol. Sci 2019, 20, 3294. [Google Scholar] [CrossRef] [Green Version]
- Da, J.J.; Zhuo, M.; Qian, M.Z. MCPIP is induced by cholesterol and participated in cholesterol-caused DNA damage in HUVEC. Int. J. Clin. Exp. Patho 2015, 8, 10625–10634. [Google Scholar]
- Zhou, B.; Ren, C.; Zu, L.; Zheng, L.; Guo, L.; Gao, W. Elevated plasma migration inhibitory factor in hypertension-hyperlipidemia patients correlates with impaired endothelial function. Medicine (Baltimore) 2016, 95, e5207. [Google Scholar] [CrossRef]
- Araujo, P.; Belghit, I.; Aarsaether, N.; Espe, M.; Lucena, E.; Holen, E. The Effect of Omega-3 and Omega-6 Polyunsaturated Fatty Acids on the Production of Cyclooxygenase and Lipoxygenase Metabolites by Human Umbilical Vein Endothelial Cells. Nutrients 2019, 11, 966. [Google Scholar] [CrossRef] [Green Version]
- Abudawood, M. Diabetes and cancer: A comprehensive review. J. Res. Med. Sci. 2019, 24, 94. [Google Scholar] [CrossRef]
- Dal Canto, E.; Ceriello, A.; Rydén, L.; Ferrini, M.; Hansen, T.B.; Schnell, O.; Standl, E.; Beulens, J.W. Diabetes as a cardiovascular risk factor: An overview of global trends of macro and micro vascular complications. Eur. J. Prev. Cardiol. 2019, 26 (Suppl 2), 25–32. [Google Scholar] [CrossRef] [Green Version]
- Katsiki, N.; Banach, M.; Mikhailidis, D.P. Is type 2 diabetes mellitus a coronary heart disease equivalent or not? Do not just enjoy the debate and forget the patient! Arch. Med. Sci. 2019, 15, 1357–1364. [Google Scholar] [CrossRef]
- Ryden, L.; Ferrannini, G.; Mellbin, L. Risk factor reduction in type 2 diabetes demands a multifactorial approach. Eur. J. Prev. Cardiol. 2019, 26, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujadas, G.; De Nigris, V.; Prattichizzo, F.; La Sala, L.; Testa, R.; Ceriello, A. The dipeptidyl peptidase-4 (DPP-4) inhibitor teneligliptin functions as antioxidant on human endothelial cells exposed to chronic hyperglycemia and metabolic high-glucose memory. Endocrine 2017, 56, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Duong, M.N.; Psaltis, P.J.; Bursill, C.A.; Nicholls, S.J. High-density lipoproteins attenuate high glucose-impaired endothelial cell signaling and functions: Potential implications for improved vascular repair in diabetes. Cardiovasc. Diabetol. 2017, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, Z.X.; Tang, Q.B. Reduced expression of microRNA-199a-3p is associated with vascular endothelial cell injury induced by type 2 diabetes mellitus. Exp. Ther. Med. 2018, 16, 3639–3645. [Google Scholar] [CrossRef]
- Yan, H.Y.; Bu, S.Z.; Zhou, W.B.; Mai, Y.F. TUG1 promotes diabetic atherosclerosis by regulating proliferation of endothelial cells via Wnt pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6922–6929. [Google Scholar] [CrossRef]
- Qin, R.; Zhang, L.; Lin, D.; Xiao, F.; Guo, L. Sirt1 inhibits HG-induced endothelial injury: Role of Mff-based mitochondrial fission and Factin homeostasis-mediated cellular migration. Int J. Mol. Med. 2019, 44, 89–102. [Google Scholar] [CrossRef]
- Shen, J.; Liu, M.; Xu, J.; Sun, B.; Xu, H.; Zhang, W. ARL15 overexpression attenuates high glucose-induced impairment of insulin signaling and oxidative stress in human umbilical vein endothelial cells. Life Sci. 2019, 220, 127–135. [Google Scholar] [CrossRef]
- Banarjee, R.; Sharma, A.; Bai, S.; Deshmukh, A.; Kulkarni, M. Proteomic study of endothelial dysfunction induced by AGEs and its possible role in diabetic cardiovascular complications. J. Proteomics 2018, 187, 69–79. [Google Scholar] [CrossRef]
- Maamoun, H.; Zachariah, M.; McVey, J.H.; Green, F.R.; Agouni, A. Heme oxygenase (HO)-1 induction prevents Endoplasmic Reticulum stress-mediated endothelial cell death and impaired angiogenic capacity. Biochem. Pharmacol. 2017, 127, 46–59. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Wang, X.F.; Li, L.; Zhang, L.; Shen, D.L.; Li, D.H.; Jin, Q.S.; Zhang, J.Y. Effects of high glucose on human umbilical vein endothelial cell permeability and myosin light chain phosphorylation. Diabetol. Metab. Syndr. 2015, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Li, G.; Li, G.; Liang, S. The role of osthole on the human umbilical vein endothelial cells in chronic high glucose. In Proceedings of the 2011 International Conference on Human Health and Biomedical Engineering (HHBE 2011), Jilin, China, 19–22 August 2011; pp. 224–226. [Google Scholar]
- Chu, P.; Han, G.; Ahsan, A.; Sun, Z.; Liu, S.; Zhang, Z.; Sun, B.; Song, Y.; Lin, Y.; Peng, J.; et al. Phosphocreatine protects endothelial cells from Methylglyoxal induced oxidative stress and apoptosis via the regulation of PI3K/Akt/eNOS and NF-kappaB pathway. Vascul. Pharmacol. 2017, 91, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safaeian, L.; Vaseghi, G.; Jabari, H.; Dana, N. Evolocumab, a proprotein convertase subtilisin/kexin type 9 inhibitor, promotes angiogenesis in vitro. Can. J. Physiol. Pharmacol. 2019, 97, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Fang, M.; Zhu, Q.; Liu, Y.; Liu, L.; Li, X. MicroRNA-939 governs vascular integrity and angiogenesis through targeting γ-catenin in endothelial cells. Biochem. Biophys. Res. Commun. 2017, 484, 27–33. [Google Scholar] [CrossRef]
- Zhang, H.M.; Liu, M.Y.; Lu, J.X.; Zhu, M.L.; Jin, Q.; Ping, S.; Li, P.; Jian, X.; Han, Y.L.; Wang, S.X.; et al. Intracellular acidosis via activation of Akt-Girdin signaling promotes post ischemic angiogenesis during hyperglycemia. Int. J. Cardiol. 2019, 277, 205–211. [Google Scholar] [CrossRef]
- Bartoszewski, R.; Serocki, M.; Janaszak-Jasiecka, A.; Bartoszewska, S.; Kochan-Jamrozy, K.; Piotrowski, A.; Kroliczewski, J.; Collawn, J.F. miR-200b downregulates Kruppel Like Factor 2 (KLF2) during acute hypoxia in human endothelial cells. Eur. J. Cell Biol. 2017, 96, 758–766. [Google Scholar] [CrossRef]
- Howe, G.A.; Kazda, K.; Addison, C.L. MicroRNA-30b controls endothelial cell capillary morphogenesis through regulation of transforming growth factor beta 2. PLoS ONE 2017, 12, e0185619. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cao, W.W.; Xing, H.; Chen, Y.L.; Li, Q.; Shen, T.T.; Jiang, C.; Zhu, D.L. Activation of ERK pathway is required for 15-HETE-induced angiogenesis in human umbilical vascular endothelial cells. J. Recept. Sig. Transd. 2016, 36, 225–232. [Google Scholar] [CrossRef]
- Zhao, X.C.; Nedvetsky, P.; Stanchi, F.; Vion, A.C.; Popp, O.; Zuhlke, K.; Dittmar, G.; Klussmann, E.; Gerhardt, H. Endothelial PKA activity regulates angiogenesis by limiting autophagy through phosphorylation of ATG16L1. Elife 2019, 8, e46380. [Google Scholar] [CrossRef]
- Bakhashab, S.; Ahmed, F.; Schulten, H.J.; Ahmed, F.W.; Glanville, M.; Al-Qahtani, M.H.; Weaver, J.U. Proangiogenic Effect of Metformin in Endothelial Cells Is via Upregulation of VEGFR1/2 and Their Signaling under Hyperglycemia-Hypoxia. Int. J. Mol. Sci. 2018, 19, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.J.; Xiang, X.L.; Xu, H.; Shi, Y.F. Cilostazol Promotes Angiogenesis and Increases Cell Proliferation After Myocardial Ischemia-Reperfusion Injury Through a cAMP-Dependent Mechanism. Cardiovasc. Eng. Techn. 2019, 10, 638–647. [Google Scholar] [CrossRef]
- Voellenkle, C.; Rooij, J.; Guffanti, A.; Brini, E.; Fasanaro, P.; Isaia, E.; Croft, L.; David, M.; Capogrossi, M.C.; Moles, A.; et al. Deep-sequencing of endothelial cells exposed to hypoxia reveals the complexity of known and novel microRNAs. RNA 2012, 18, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Zhang, X.; Zhang, W.; Zhao, S.; Chen, Y. Identification and characterization of cell-bound membrane vesicles. Biochim. Biophys. Acta Biomembr. 2017, 1859, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.Y.; Lee, C.K.; Huang, C.; Ou, Y.H.; Charles, C.J.; Richards, A.M.; Neupane, Y.R.; Pavon, M.V.; Zharkova, O.; Pastorin, G.; et al. Extracellular Vesicles in Cardiovascular Diseases: Alternative Biomarker Sources, Therapeutic Agents, and Drug Delivery Carriers. Int. J. Mol. Sci. 2019, 20, 3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, M.N.; Liu, X.F.; Xu, G.L. Extracellular Vesicles as Messengers in Atherosclerosis. J. Cardiovasc. Transl. 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Diehl, P.; Fricke, A.; Sander, L.; Stamm, J.; Bassler, N.; Htun, N.; Ziemann, M.; Helbing, T.; El-Osta, A.; Jowett, J.B.; et al. Microparticles: Major transport vehicles for distinct microRNAs in circulation. Cardiovasc. Res. 2012, 93, 633–644. [Google Scholar] [CrossRef]
- Burger, D.; Turner, M.; Xiao, F.; Munkonda, M.N.; Akbari, S.; Burns, K.D. High glucose increases the formation and pro-oxidative activity of endothelial microparticles. Diabetologia 2017, 60, 1791–1800. [Google Scholar] [CrossRef] [Green Version]
- Hosseinkhani, B.; Kuypers, S.; van den Akker, N.M.S.; Molin, D.G.M.; Michiels, L. Extracellular Vesicles Work as a Functional Inflammatory Mediator Between Vascular Endothelial Cells and Immune Cells. Front. Immunol. 2018, 9, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, S.M.; Riquelme, J.A.; Zheng, Y.; Vicencio, J.M.; Lavandero, S.; Yellon, D.M. Endothelial cells release cardioprotective exosomes that may contribute to ischaemic preconditioning. Sci Rep. UK 2018, 8, 15885. [Google Scholar] [CrossRef]
- Lin, X.; Li, S.; Wang, Y.J.; Wang, Y.; Zhong, J.Y.; He, J.Y.; Cui, X.J.; Zhan, J.K.; Liu, Y.S. Exosomal Notch3 from high glucose-stimulated endothelial cells regulates vascular smooth muscle cells calcification/aging. Life Sci. 2019, 232, 116582. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, W.; Zhang, R.; Wu, J.; Li, L.; Tang, Y. Proteomic analysis of TNF-alpha-activated endothelial cells and endothelial microparticles. Mol. Med. Rep. 2013, 7, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Yang, S.H.; Kwon, I.; Lee, O.H.; Heo, J.H. Role of tumour necrosis factor receptor-1 and nuclear factor-kappaB in production of TNF-alpha-induced pro-inflammatory microparticles in endothelial cells. Thromb. Haemost. 2014, 112, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Sung, H.C.; Lin, S.R.; Wu, C.W.; Lee, C.W.; Lee, I.T.; Yang, Y.F.; Yu, I.S.; Lin, S.W.; Chiang, M.H.; et al. Resveratrol attenuates ICAM-1 expression and monocyte adhesiveness to TNF-alpha-treated endothelial cells: Evidence for an anti-inflammatory cascade mediated by the miR-221/222/AMPK/p38/NF-kappaB pathway. Sci. Rep. 2017, 7, 44689. [Google Scholar] [CrossRef] [PubMed]
- Holnthoner, W.; Bonstingl, C.; Hromada, C.; Muehleder, S.; Zipperle, J.; Stojkovic, S.; Redl, H.; Wojta, J.; Schochl, H.; Grillari, J.; et al. Endothelial Cell-derived Extracellular Vesicles Size-dependently Exert Procoagulant Activity Detected by Thromboelastometry. Sci. Rep. 2017, 7, 3707. [Google Scholar] [CrossRef]
- Tang, F.; Yang, T.L. MicroRNA-126 alleviates endothelial cells injury in atherosclerosis by restoring autophagic flux via inhibiting of PI3K/Akt/mTOR pathway. Biochem. Biophys. Res. Commun. 2018, 495, 1482–1489. [Google Scholar] [CrossRef]
- Tomberli, B.; Mattesini, A.; Baldereschi, G.I.; Di Mario, C. A Brief History of Coronary Artery Stents. Rev. Esp Cardiol (Engl Ed.) 2018, 71, 312–319. [Google Scholar] [CrossRef]
- Giordano, A.; Romano, S.; Corcione, N.; Frati, G.; Zoccai, G.B.; Ferraro, P.; Messina, S.; Ottolini, S.; Romano, M.F. Tirofiban Positively Regulates beta1 Integrin and Favours Endothelial Cell Growth on Polylactic Acid Biopolymer Vascular Scaffold (BVS). J. Cardiovasc. Transl. Res. 2018, 11, 201–209. [Google Scholar] [CrossRef]
- Wang, J.; Song, C.; Xiao, Y.; Liu, B. In vivo and in vitro analyses of the effects of a novel high-nitrogen low-nickel coronary stent on reducing in-stent restenosis. J. Biomater. Appl. 2018, 33, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Beshchasna, N.; Ho, A.Y.K.; Saqib, M.; Kraskiewicz, H.; Wasyluk, L.; Kuzmin, O.; Duta, O.C.; Ficai, D.; Trusca, R.D.; Ficai, A.; et al. Surface evaluation of titanium oxynitride coatings used for developing layered cardiovascular stents. Mat. Sci Eng. C 2019, 99, 405–416. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, Y.; Xiong, K.; Li, X.; Qi, P.; Tu, Q.; Jing, F.; Weng, Y.; Wang, J.; Huang, N. Nitric oxide producing coating mimicking endothelium function for multifunctional vascular stents. Biomaterials 2015, 63, 80–92. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution | Composition | Conservation time | References |
|---|---|---|---|
| Cord Buffer, pH 7.4 | NaCl 0.14 M KCl 0.004 M Phosphate buffer solution 0.001 M Glucose 0.011 M | <3 h | [28] |
| Phosphate buffer solution (cold) | Calcium, magnesium, streptomycin and penicillin. | 2-3 h | [41] |
| Transportation buffer | NaCl 1.4M Na2HPO4 5.2mM KCl 40 mM. | <24 h | [42] |
| Transportation and conservation buffer | 50 mL of PBS 1X 1 mL of penicillin 1 mL of colistin | <6 h | [11] |
| HEPES (1X) buffer | NaCl 137 mM KCL 4 mM HEPES 10 mM Glucose 11.1 mM. | <3 h | [43] |
| Supplemented PBS | PBS supplemented with 1.5% antibiotic-antimycotic. | <3 h | [44] |
| Culture Medium | Additional reagents | Temperature and humidity | Periodicity of change of medium | References |
|---|---|---|---|---|
| TC199 | FBS, penicillin (200 U/ml), streptomycin (200 µg/mL), L-glutamine (2mM). | 37°C, 5% CO2 | Twice a week | [28] |
| M199 | 15 mg/L of HEPES, penicillin (60 mg/mL), streptomycin (129 mg/mL), 20% of FBS | 37°C, 5% CO2 | Every 48 hours | [29] |
| M199 or RPMI 1640 | HEPES 25mM, 20% of FBS, L- glutamine (0.3 mg/mL), penicillin (100 U/mL), streptomycin (100 µg/ mL) and anti-PPLO agent (0.03 mg/mL). | 37°C, 5% CO2. | Twice a week | [41] |
| Minimum Essential Medium Eagle | 10% of FBS, L-glutamine (1%) | 37°C, 5% CO2. | Every 72 hours | [46] |
| M199 | 20% of FBS, penicillin (100 UI/ mL), streptomycin (100 µg/mL) y L-glutamine (2 mM), ECGF. | 37°C, 5% CO2. | Every 48 hours | [42] |
| MCDB 131 | 20% of FBS, penicillin and streptomycin (50 U/mL-50 U/mL), 20 µg/mL of ECGF, heparin (16 U/mL). | 37°C, 5% CO2. | Every 72 hours | [50] |
| M199 | 20% of FBS, 1 mL of penicillin-streptomycin (10,000 U by 10,000 U). | 37°C, 5% CO2. | Every 48 hours | [11] |
| M199 | 20% of FBS, 50 µg/ mL of ECGF, 50 µg/ mL of heparin, L-glutamine (2mM), penicillin (100 U/mL), streptomycin (100 µg/ mL), amphotericin (2 µg/mL). | 37°C, 5% CO2. | Twice a week | [43] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Leyte, D.J.; Domínguez-Pérez, M.; Mercado, I.; Villarreal-Molina, M.T.; Jacobo-Albavera, L. Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review. Appl. Sci. 2020, 10, 938. https://doi.org/10.3390/app10030938
Medina-Leyte DJ, Domínguez-Pérez M, Mercado I, Villarreal-Molina MT, Jacobo-Albavera L. Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review. Applied Sciences. 2020; 10(3):938. https://doi.org/10.3390/app10030938
Chicago/Turabian StyleMedina-Leyte, Diana J., Mayra Domínguez-Pérez, Ingrid Mercado, María T. Villarreal-Molina, and Leonor Jacobo-Albavera. 2020. "Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review" Applied Sciences 10, no. 3: 938. https://doi.org/10.3390/app10030938