Phaseolus vulgaris L. var. Venanzio Grown in Tuscany: Chemical Composition and In Vitro Investigation of Potential Effects on Colorectal Cancer

 , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Preparation of Extracts of FV
2.2. Polyphenols Content
2.3. Soluble Carbohydrates Content
2.4. Protein Content
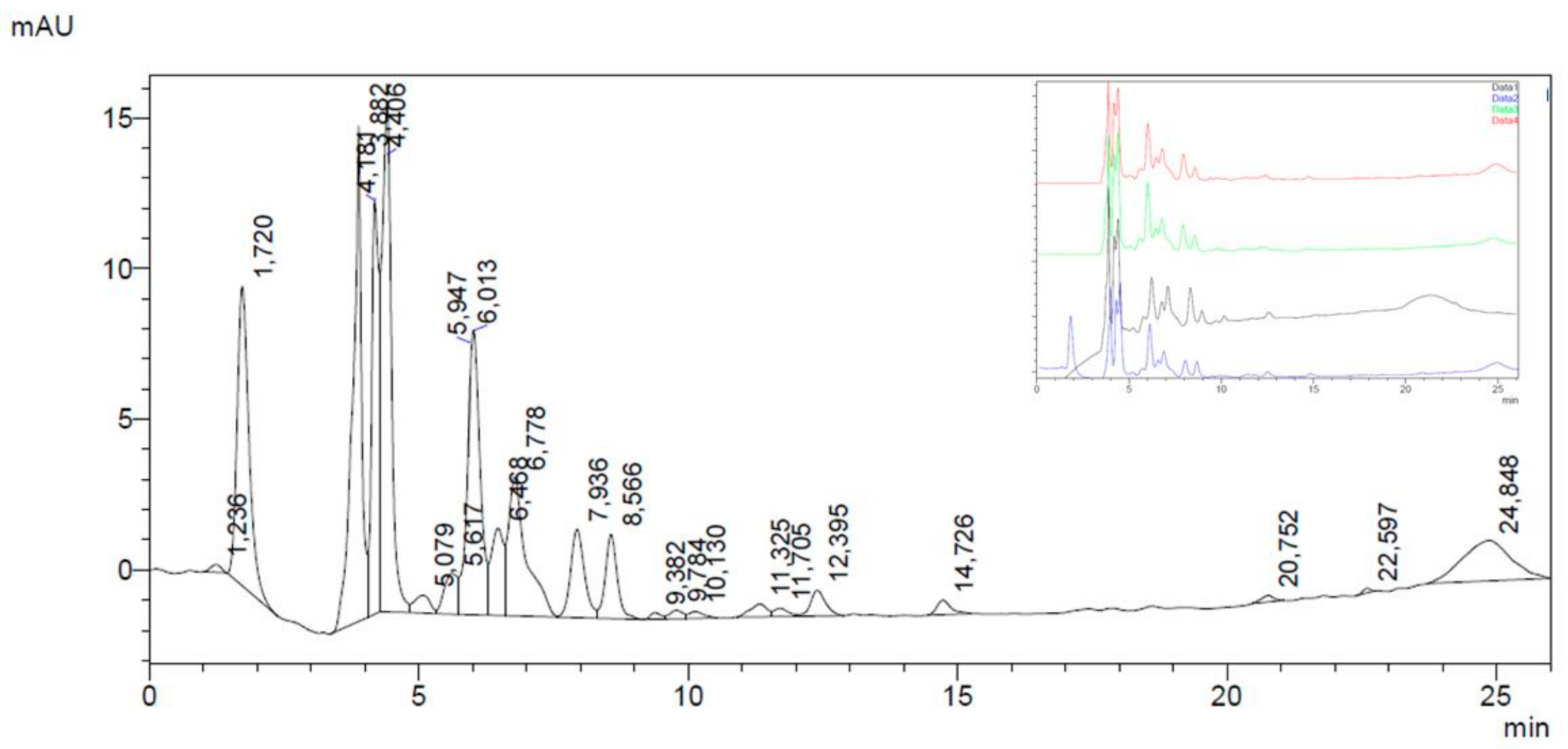
2.5. HPLC-DAD Analysis on Main Polyphenolic Constituents
2.6. HPLC-DAD-DPPH (2,2-Diphenyl-1-picrylhydrazyl)
2.7. Cell Culture
2.8. MTT Assay
2.9. Western Blotting Analysis
2.10. ROS Measurement
2.11. Statistical Analysis
3. Results
3.1. Chemical Composition of FV Extracts
3.2. DPPH Test and HPLC-DAD-DPPH
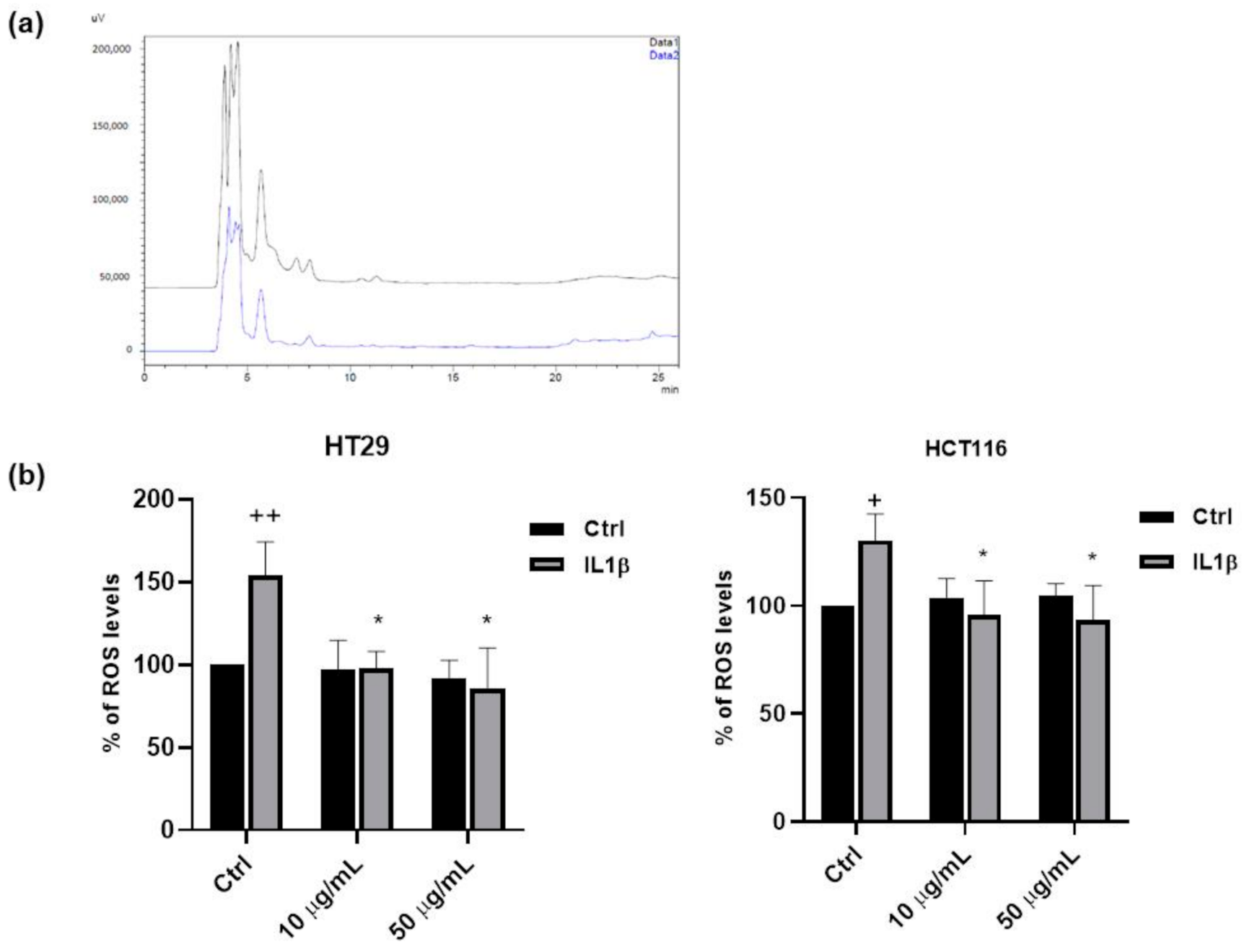
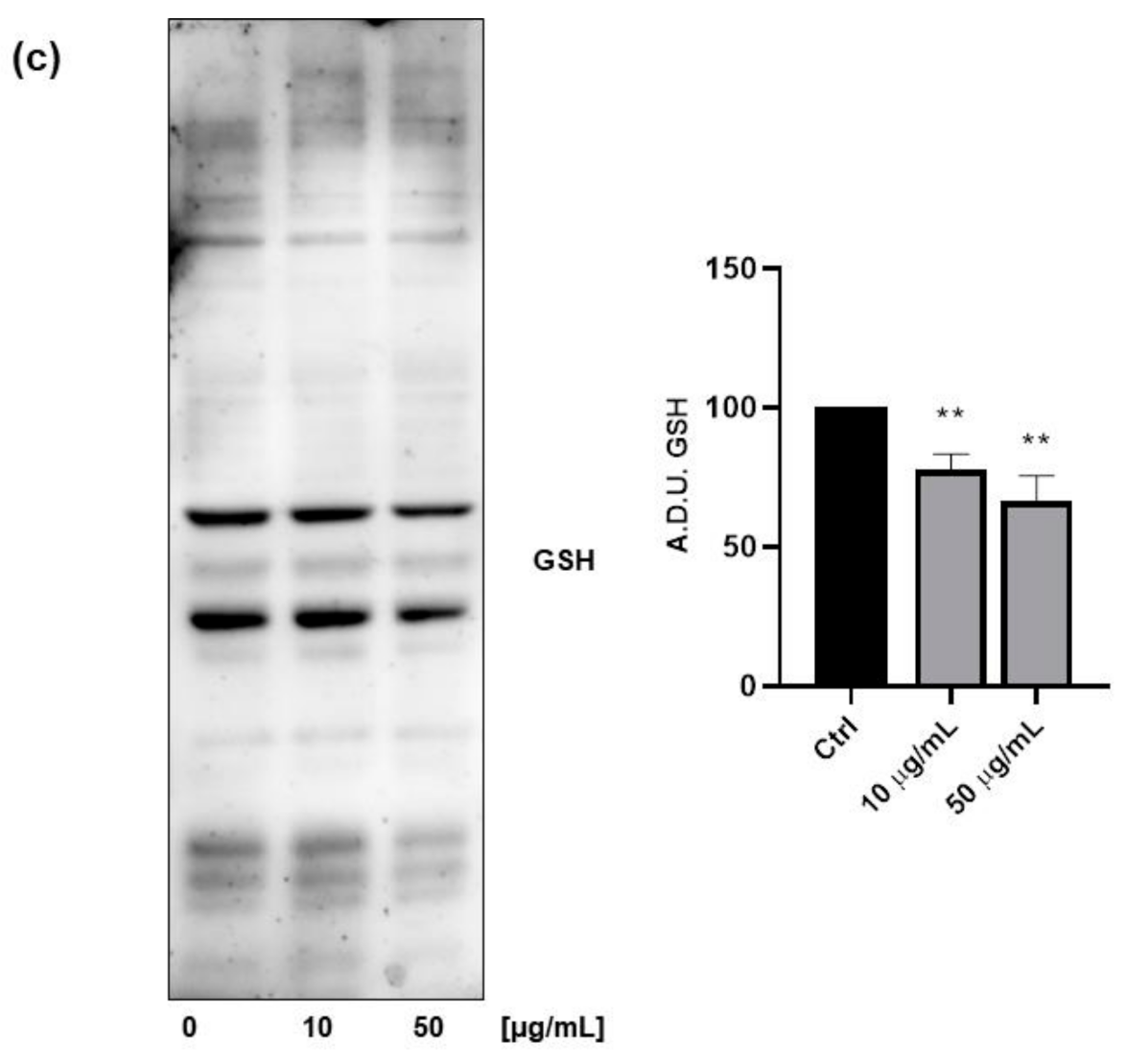
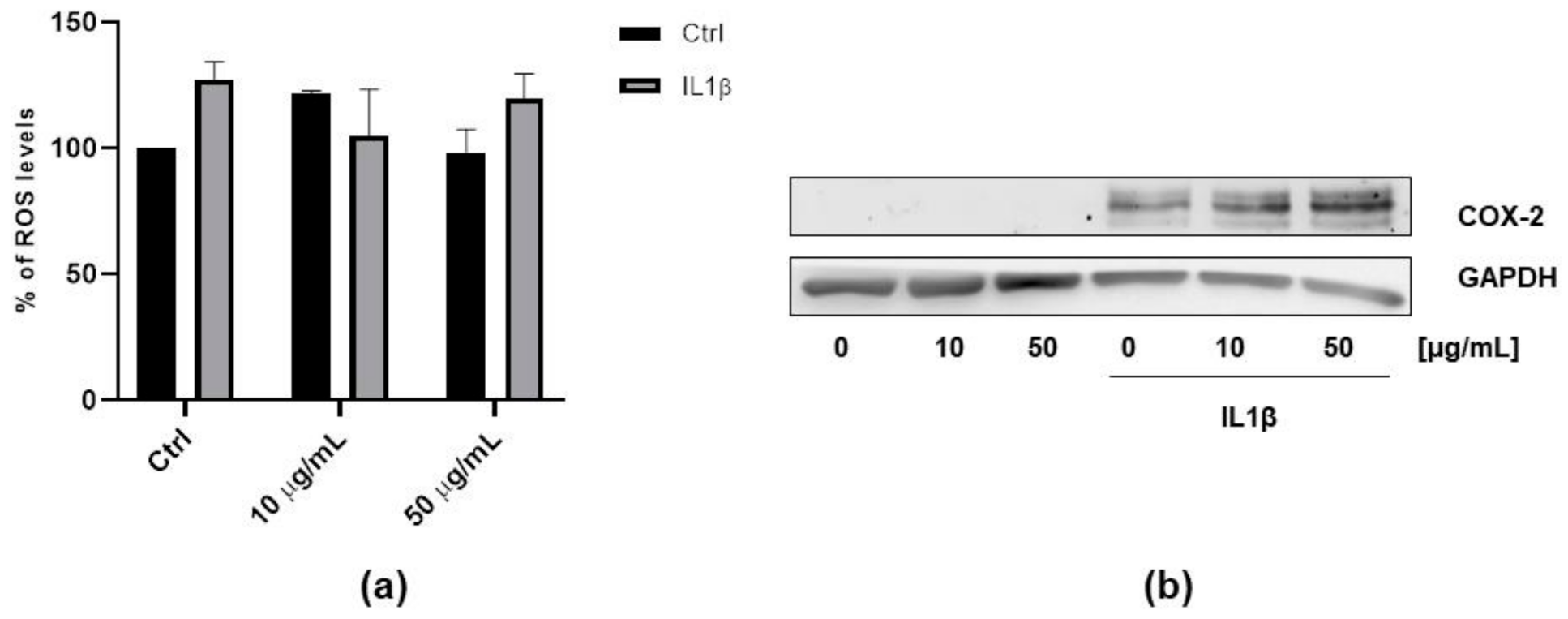
3.3. Antioxidant Properties of FV Extract
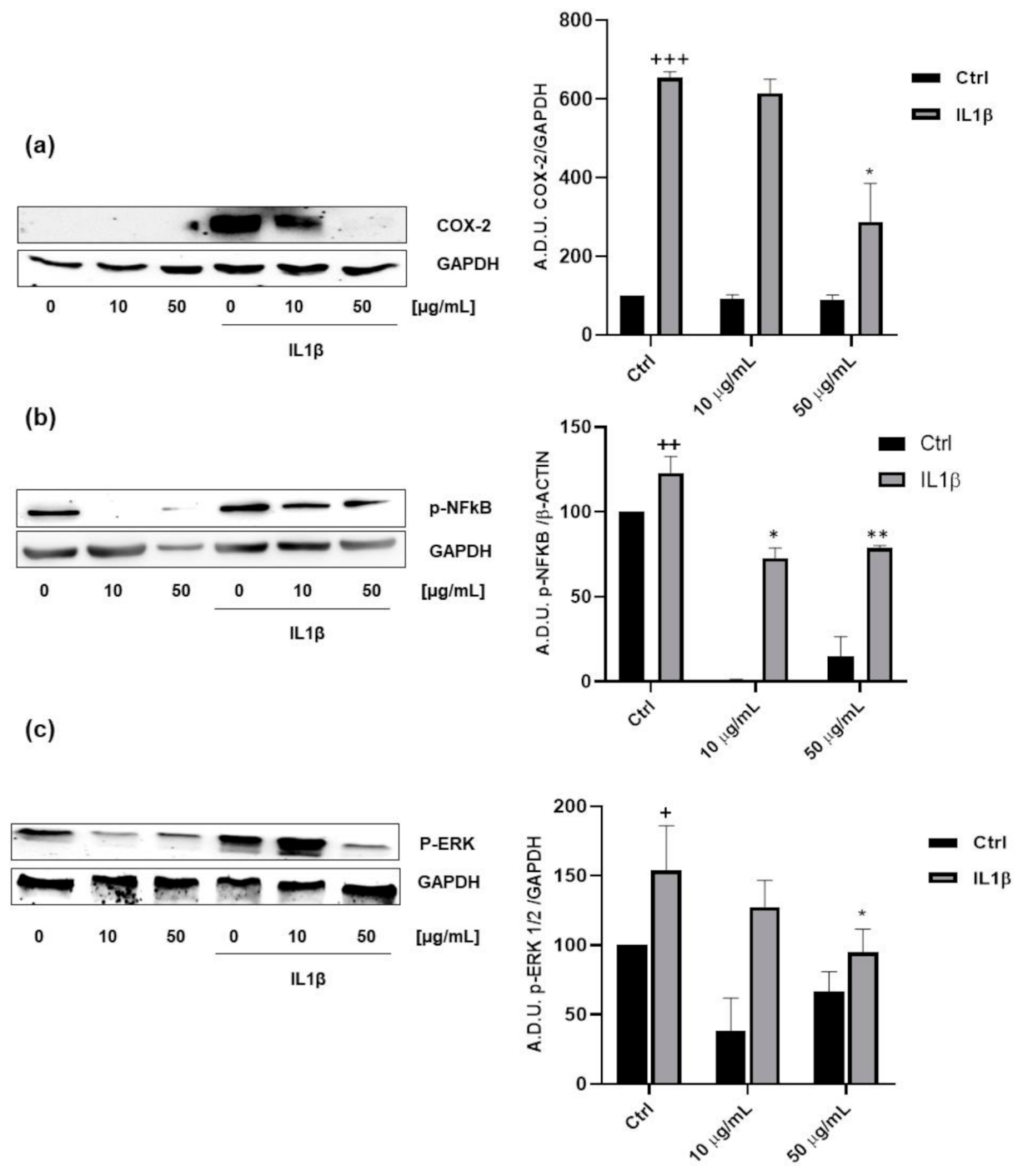
3.4. Anti-Inflammatory Properties of FV Extract
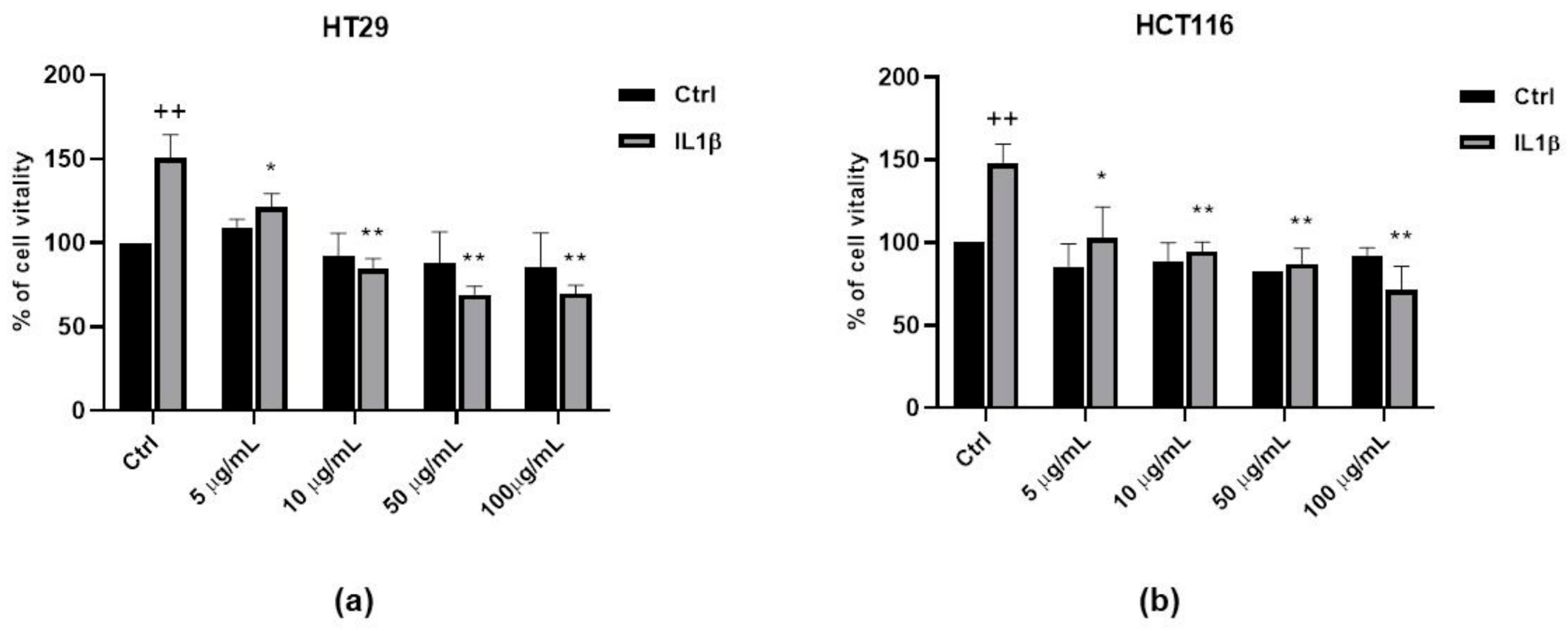
3.5. Anti-Proliferative Activity of FV Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Teodoro, A.J. Bioactive compounds of food: Their role in the prevention and treatment of diseases. Oxid. Med. Cell. Longev. 2019, 2019, 3765986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez-Martínez, S.E.; Ferriz-Martínez, R.A.; Campos-Vega, R.; Elton-Puente, J.E.; Carbot, K.d.; García-Gasca, T. Bean seeds: Leading nutraceutical source for human health. CYTA J. Food 2016, 14, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Ombra, M.N.; D’Acierno, A.; Nazzaro, F.; Riccardi, R.; Spigno, P.; Zaccardelli, M.; Pane, C.; Maione, M.; Fratianni, F. Phenolic Composition and Antioxidant and Antiproliferative Activities of the Extracts of Twelve Common Bean (Phaseolus vulgaris L.) Endemic Ecotypes of Southern Italy before and after Cooking. Oxid. Med. Cell. Longev. 2016, 2016, 1398298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, K.; Xu, B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.Q.; Gan, R.Y.; Ge, Y.Y.; Zhang, D.; Corke, H. Polyphenols in Common Beans (Phaseolus vulgaris L.): Chemistry, Analysis, and Factors Affecting Composition. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1518–1539. [Google Scholar] [CrossRef] [Green Version]
- Hayat, I.; Ahmad, A.; Masud, T.; Ahmed, A.; Bashir, S. Nutritional and Health Perspectives of Beans (Phaseolus vulgaris L.): An Overview. Crit. Rev. Food Sci. Nutr. 2014, 54, 580–592. [Google Scholar] [CrossRef]
- Thompson, S.V.; Winham, D.M.; Hutchins, A.M. Bean and rice meals reduce postprandial glycemic response in adults with type 2 diabetes: A cross-over study. Nutr. J. 2012, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Correa, P. Epidemiological Correlations Between Diet and Cancer Frequency. Cancer Res. 1981, 41, 3685–3689. [Google Scholar]
- Kolonel, L.N.; Hankin, J.H.; Whittemore, A.S.; Wu, A.H.; Gallagher, R.P.; Wilkens, L.R.; John, E.M.; Howe, G.R.; Dreon, D.M.; West, D.W.; et al. Vegetables, fruits, legumes and prostate cancer: A multiethnic case-control study. Cancer Epidemiol. Biomark. Prev. 2000, 9, 795–804. [Google Scholar]
- Hughes, J.S.; Ganthavorn, C.; Wilson-Sanders, S. Dry beans inhibit azoxymethane-induced colon carcinogenesis in F344 rats. J. Nutr. 1997, 127, 2328–2333. [Google Scholar] [CrossRef]
- Hangen, L.; Bennink, M.R. Consumption of black beans and navy beans (Phaseolus vulgaris) reduced azoxymethane-induced colon cancer in rats. Nutr. Cancer 2002, 44, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Feregrino-Pérez, A.A.; Berumen, L.C.; García-Alcocer, G.; Guevara-Gonzalez, R.G.; Ramos-Gomez, M.; Reynoso-Camacho, R.; Acosta-Gallegos, J.A.; Loarca-Piña, G. Composition and chemopreventive effect of polysaccharides from common beans (Phaseolus vulgaris L.) on azoxymethane-induced colon cancer. J. Agric. Food Chem. 2008, 56, 8737–8744. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Xia, L.; Ng, T.B. White kidney bean lectin exerts anti-proliferative and apoptotic effects on cancer cells. Int. J. Biol. Macromol. 2016, 85, 335–345. [Google Scholar] [CrossRef]
- Moreno-Jiménez, M.R.; López-Barraza, R.; Cervantes-Cardoza, V.; Pérez-Ramírez, I.F.; Reyna-Rojas, J.A.; Gallegos-Infante, J.A.; Estrella, I.; Rojas-Contreras, J.A.; González-Laredo, R.F.; Rocha-Guzmán, N.E. Mechanisms associated to apoptosis of cancer cells by phenolic extracts from two canned common beans varieties (Phaseolus vulgaris L.). J. Food Biochem. 2019, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Feregrino-Perez, A.A.N.; Piñol-Felis, C.; Gomez-Arbones, X.; Guevara-González, R.G.; Campos-Vega, R.; Acosta-Gallegos, J.; Loarca-Piña, G. A non-digestible fraction of the common bean (Phaseolus vulgaris L.) induces cell cycle arrest and apoptosis during early carcinogenesis. Plant Foods Hum. Nutr. 2014, 69, 248–254. [Google Scholar] [CrossRef]
- Vergara-Castañeda, H.A.; Guevara-González, R.G.; Ramos-Gómez, M.; Reynoso-Camacho, R.; Guzmán-Maldonado, H.; Feregrino-Pérez, A.A.; Oomah, B.D.; Loarca-Piña, G. Non-digestible fraction of cooked bean (Phaseolus vulgaris L.) cultivar Bayo Madero suppresses colonic aberrant crypt foci in azoxymethane-induced rats. Food Funct. 2010, 1, 294–300. [Google Scholar] [CrossRef]
- Mensack, M.M.; McGinley, J.N.; Thompson, H.J. Metabolomic analysis of the effects of edible dry beans (Phaseolus vulgaris L.) on tissue lipid metabolism and carcinogenesis in rats. Br. J. Nutr. 2012, 108, S155–S165. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.D.; Mensack, M.M.; Jiang, W.; Zhu, Z.; Lewis, M.R.; McGinley, J.N.; Brick, M.A.; Thompson, H.J. Cell signaling pathways associated with a reduction in mammary cancer burden by dietary common bean (Phaseolus vulgaris L.). Carcinogenesis 2012, 33, 226–232. [Google Scholar] [CrossRef]
- Campos-Vega, R.; García-Gasca, T.; Guevara-Gonzalez, R.; Ramos-Gomez, M.; Oomah, B.D.; Loarca-Piña, G. Human gut flora-fermented nondigestible fraction from cooked bean (Phaseolus vulgaris L.) modifies protein expression associated with apoptosis, cell cycle arrest, and proliferation in human adenocarcinoma colon cancer cells. J. Agric. Food Chem. 2012, 60, 12443–12450. [Google Scholar] [CrossRef]
- Biagi, M.; Noto, D.; Corsini, M.; Baini, G.; Cerretani, D.; Cappellucci, G.; Moretti, E. Antioxidant Effect of the Castanea sativa Mill. Leaf Extract on Oxidative Stress Induced upon Human Spermatozoa. Oxid. Med. Cell. Longev. 2019, 2019, 8926075. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Terzuoli, E.; Donnini, S.; Uva, M.; Ziche, M.; Morbidelli, L. Monitoring endothelial and tissue responses to cobalt ferrite Nanoparticles and Hybrid Hydrogels. PLoS ONE 2016, 11, e0168727. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Moglia, A.; Schiavo, I.; Donnini, S.; Berta, G.N.; di Scipio, F.; Perrelli, A.; Fornelli, C.; Trabalzini, L.; Retta, S.F. Yeast-derived recombinant avenanthramides inhibit proliferation, migration and epithelial mesenchymal transition of colon cancer cells. Nutrients 2018, 10, 1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finetti, F.; Schiavo, I.; Ercoli, J.; Zotta, A.; Boda, E.; Retta, S.F.; Trabalzini, L. KRIT1 loss-mediated upregulation of NOX1 in stromal cells promotes paracrine pro-angiogenic responses. Cell. Signal. 2020, 68, 109527. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Colombo, G.; Giustarini, D.; Milzani, A. Protein S-glutathionylation: A regulatory device from bacteria to humans. Trends Biochem. Sci. 2009, 34, 85–96. [Google Scholar] [CrossRef]
- Hsieh, H.J.; Liu, C.A.; Huang, B.; Tseng, A.H.; Wang, D.L. Shear-induced endothelial mechanotransduction: The interplay between reactive oxygen species (ROS) and nitric oxide (NO) and the pathophysiological implications. J. Biomed. Sci. 2014, 21, 3. [Google Scholar] [CrossRef] [Green Version]
- Goradel, N.H.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell. Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef]
- Donnini, S.; Finetti, F.; Terzuoli, E.; Giachetti, A.; Ĩiguez, M.A.; Hanaka, H.; Fresno, M.; Rådmark, O.; Ziche, M. EGFR signaling upregulates expression of microsomal prostaglandin e synthase-1 in cancer cells leading to enhanced tumorigenicity. Oncogene 2012, 31, 3457–3466. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, W.; Sanidad, K.Z.; Shih, P.A.; Zhao, X.; Zhang, G. Eicosanoid signaling in carcinogenesis of colorectal cancer. Cancer Metastasis Rev. 2018, 37, 257–267. [Google Scholar] [CrossRef]
- Piotrowski, I.; Kulcenty, K.; Suchorska, W. Interplay between inflammation and cancer. Rep. Pract. Oncol. Radiother. 2020, 25, 422–427. [Google Scholar] [CrossRef]
- Nasef, N.A.; Mehta, S. Role of inflammation in pathophysiology of colonic disease: An update. Int. J. Mol. Sci. 2020, 21, 4748. [Google Scholar] [CrossRef]
- Patrignani, P.; Patrono, C. Aspirin and Cancer. J. Am. Coll. Cardiol. 2016, 68, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Reinmuth, N.; Stoeltzing, O.; Parikh, A.A.; Tellez, C.; Williams, S.; Jung, Y.D.; Fan, F.; Takeda, A.; Akagi, M.; et al. Cyclooxygenase-2 is up-regulated by interleukin-1β in human colorectal cancer cells via multiple signaling pathways. Cancer Res. 2003, 63, 3632–3636. [Google Scholar] [PubMed]
- Gupta, R.A.; Dubois, R.N. Colorectal Cancer Prevention of Cyclooxygenase-2. Nat. Rev. Cancer 2001, 1, 11–21. [Google Scholar] [CrossRef]
- Owczarek, K.; Lewandowska, U. The Impact of Dietary Polyphenols on COX-2 Expression in Colorectal Cancer. Nutr. Cancer 2017, 69, 1105–1118. [Google Scholar] [CrossRef]
- Monti, M.; Terzuoli, E.; Ziche, M.; Morbidelli, L. H2S dependent and independent anti-inflammatory activity of zofenoprilat in cells of the vascular wall. Pharmacol. Res. 2016, 113, 426–437. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
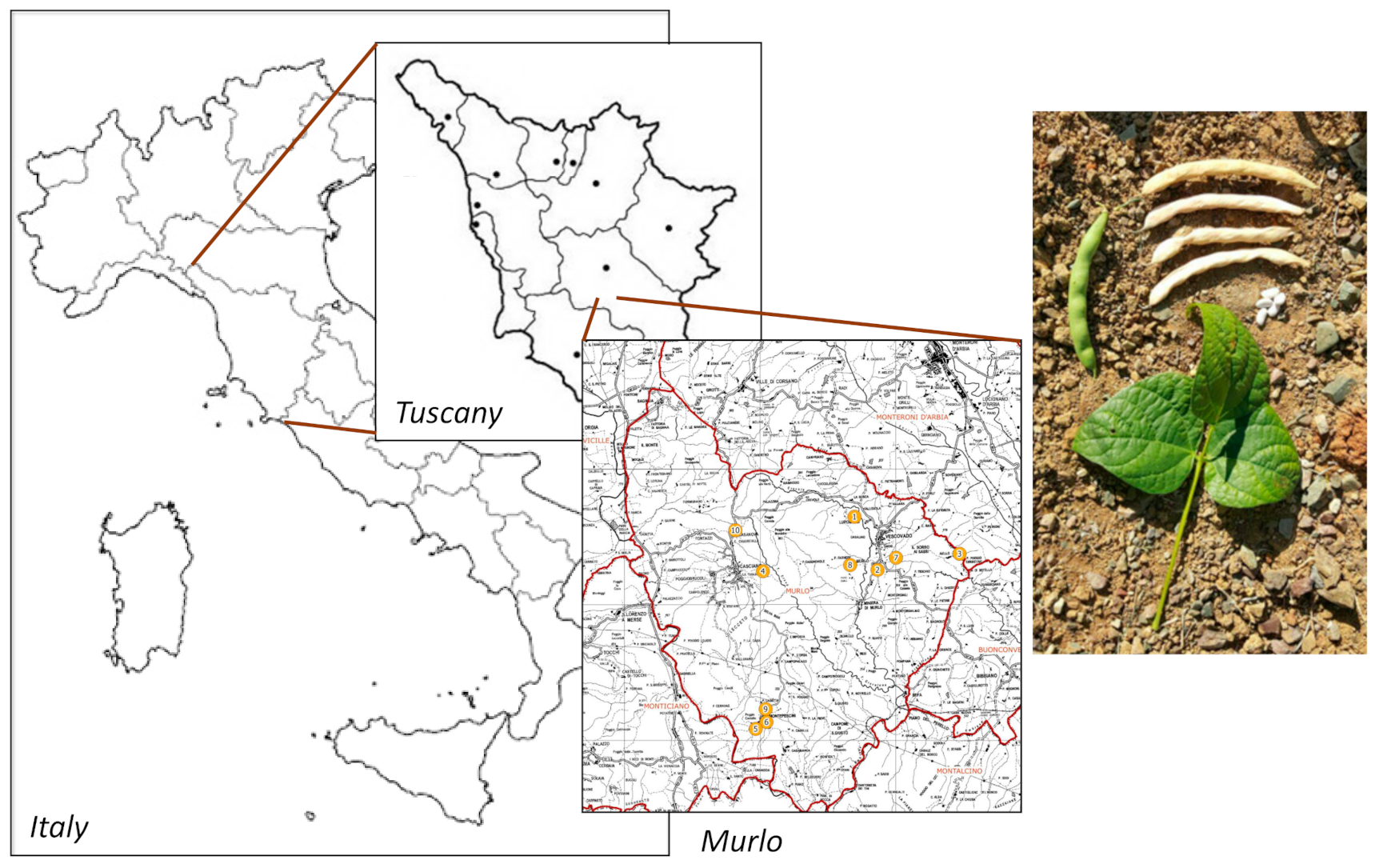
| Grower | # Sample |
|---|---|
| Società Agricola Aiellino | 1 |
| Nicola Ulivieri | 2 |
| Burresi family | 3 |
| Azienda Agricola Podere Vignali | 4 |
| Components | Quantification (mg/g) | |||
|---|---|---|---|---|
| #1 | #2 | #3 | #4 | |
| Total polyphenols | 0.142 ± 0.018 | 0.123 ± 0.011 | 0.129 ± 0.016 | 0.120 ± 0.015 |
| Total hydroxycinnamic derivatives | 0.054 ± 0.004 | 0.051 ± 0.003 | 0.046 ± 0.005 | 0.052 ± 0.005 |
| Isoflavones | <0.005 | <0.005 | <0.005 | <0.005 |
| Composition | Quantification |
|---|---|
| Total polyphenols | 0.131 ± 0.016 mg/g |
| Total hydroxycinnamic derivatives | 0.046 ± 0.004 mg/g |
| Gallic acid | 0.052 ± 0.005 mg/g |
| Chlorogenic acid | 0.011 ± 0.002 mg/g |
| Isoflavones | <0.005 mg/g |
| Total soluble carbohydrates | 10.032 ± 0.820 mg/g |
| Total proteins | 15.190 ± 2.020 mg/g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finetti, F.; Biagi, M.; Ercoli, J.; Macrì, G.; Miraldi, E.; Trabalzini, L. Phaseolus vulgaris L. var. Venanzio Grown in Tuscany: Chemical Composition and In Vitro Investigation of Potential Effects on Colorectal Cancer. Antioxidants 2020, 9, 1181. https://doi.org/10.3390/antiox9121181
Finetti F, Biagi M, Ercoli J, Macrì G, Miraldi E, Trabalzini L. Phaseolus vulgaris L. var. Venanzio Grown in Tuscany: Chemical Composition and In Vitro Investigation of Potential Effects on Colorectal Cancer. Antioxidants. 2020; 9(12):1181. https://doi.org/10.3390/antiox9121181
Chicago/Turabian StyleFinetti, Federica, Marco Biagi, Jasmine Ercoli, Giulia Macrì, Elisabetta Miraldi, and Lorenza Trabalzini. 2020. "Phaseolus vulgaris L. var. Venanzio Grown in Tuscany: Chemical Composition and In Vitro Investigation of Potential Effects on Colorectal Cancer" Antioxidants 9, no. 12: 1181. https://doi.org/10.3390/antiox9121181