Antioxidant Properties and Protective Effects of Some Species of the Annonaceae, Lamiaceae, and Geraniaceae Families against Neuronal Damage Induced by Excitotoxicity and Cerebral Ischemia


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material
2.3. Preparation of the Plant Extracts
2.4. OH• Scavenging Assay
2.5. ROO• Scavenging Assay
2.6. O₂•− Scavenging Assay
2.7. Experimental Design
2.8. Treatment with Plant Extracts
2.9. Oxidative Stress in Primary Neuronal Cultures Induced by Excitotoxicity
2.10. Determination of Cytotoxicity
2.11. Immunofluorescence to Identify Neurons
2.12. Measurement of Intracellular O₂•− Production Induced by Excitotoxicity
2.13. Induction of Cerebral Ischemia with the Middle Cerebral Artery Occlusion (MCAO) Model
2.14. Quantification of Tissue Damage, Neurological Outcome, and Survival after MCAO
2.15. Statistical Analysis
3. Results
3.1. OH• Scavenging Activity of Annonaceae, Lamiaceae, and Geraniaceae Extracts
3.2. ROO• Scavenging Activity of Annonaceae, Lamiaceae, and Geraniaceae Extracts
3.3. O₂•− Scavenging Activity of Annonaceae, Lamiaceae, and Geraniaceae Extracts
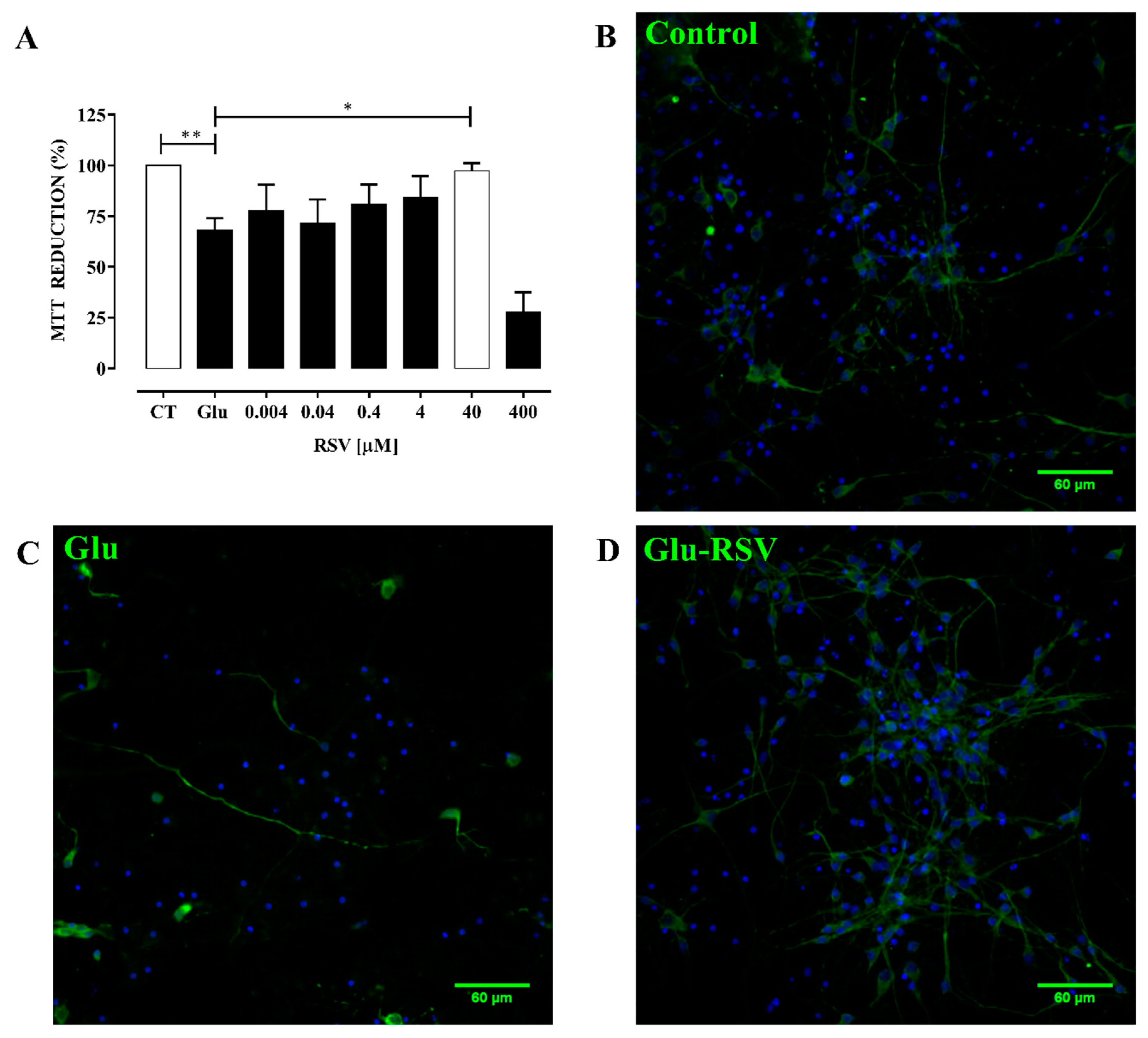
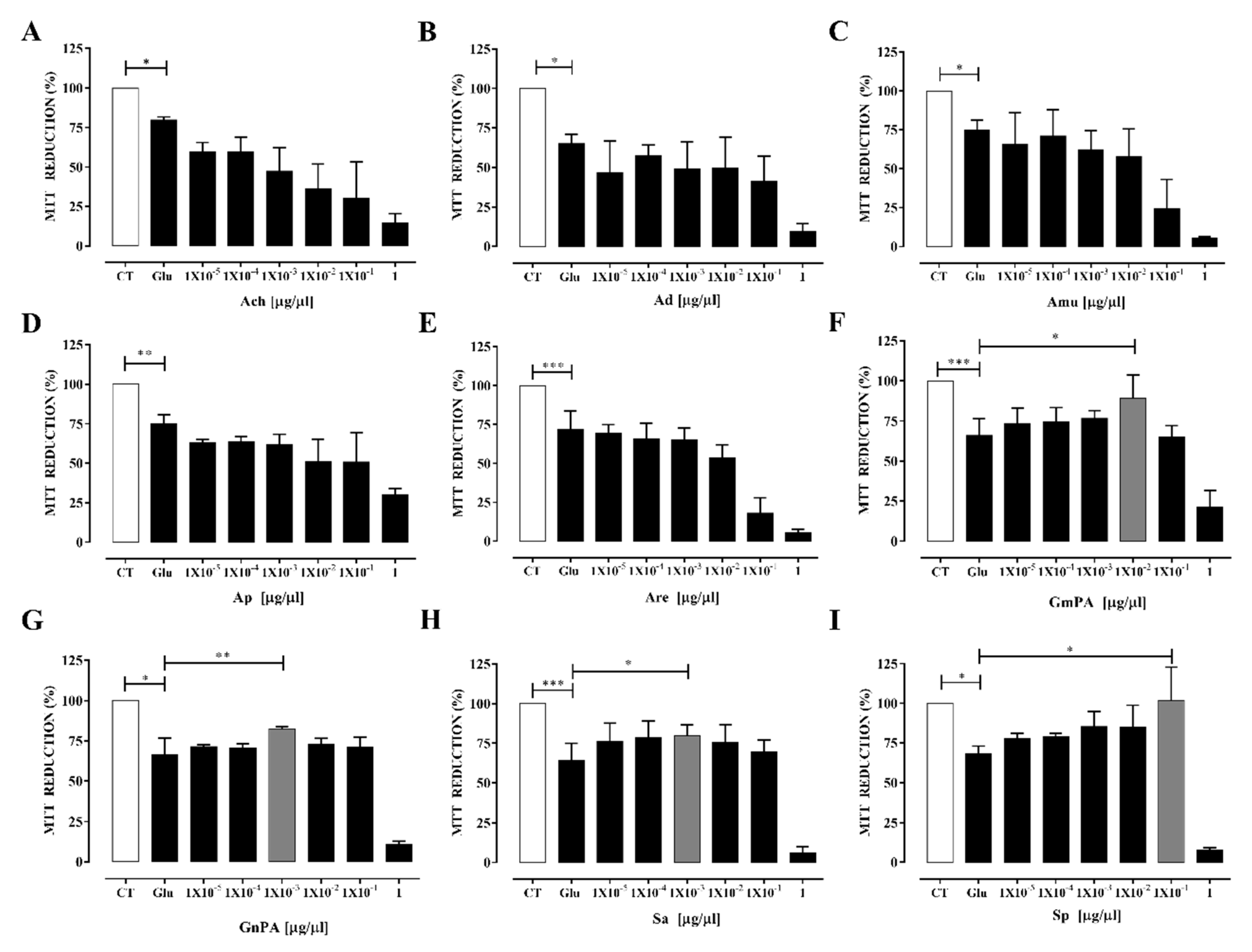
3.4. Plant Extracts Obtained from Geraniaceae and Lamiaceae Families Protect Neurons from Excitotoxicity-Induced Oxidative Stress
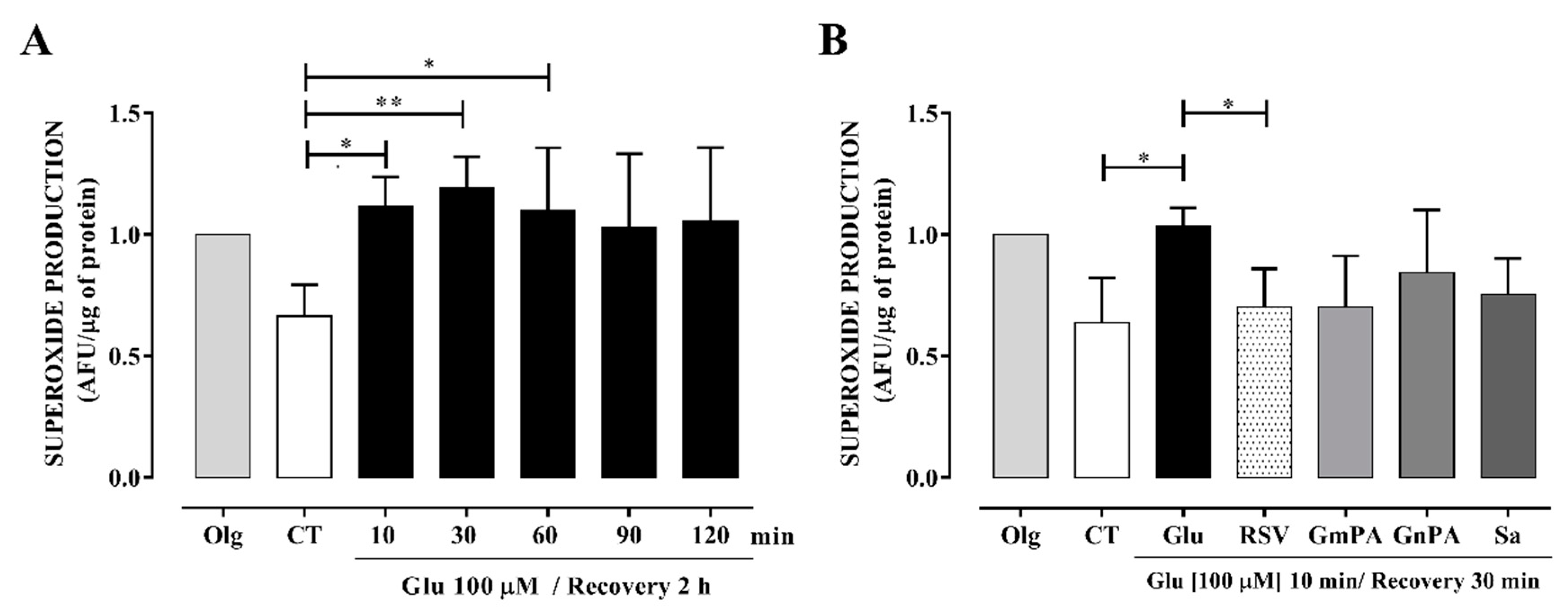
3.5. Extracts Obtained from the Geraniaceae and Lamiaceae Families Reduced Intracellular Production of O₂•−
3.6. Sp Extract Prevented the Cerebral Ischemia-Induced Damage
4. Discussion
4.1. Extracts Prevent Oxidative Stress Induced by Excitotoxicity in Cultured Neurons
4.2. Sp Extract Protects against Cerebral Ischemia-Induced Damage
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The science of stroke: Mechanisms in search of treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Ramírez, N.; Gutiérrez Aguilar, G.F.; Espinoza-Rojo, M.; Aguilera, P. Current evidence for AMPK activation involvement on resveratrol-induced neuroprotection in cerebral ischemia. Nutr. Neurosci. 2018, 21, 229–247. [Google Scholar] [CrossRef]
- Huang, W.Y.; Davidge, S.T.; Wu, J. Bioactive Natural Constituents from Food Sources-Potential Use in Hypertension Prevention and Treatment. Crit. Rev. Food Sci. Nutr. 2013, 53, 615–630. [Google Scholar] [CrossRef]
- Karimi, A.; Majlesi, M.; Rafieian-Kopaei, M. Herbal versus synthetic drugs; beliefs and facts. J. nephropharmacology 2015, 4, 27–30. [Google Scholar]
- Sentkowska, A.; Pyrzyńska, K. Investigation of antioxidant activity of selenium compounds and their mixtures with tea polyphenols. Mol. Biol. Rep. 2019, 46, 3019–3024. [Google Scholar] [CrossRef] [Green Version]
- Jardim, F.R.; de Rossi, F.T.; Nascimento, M.X.; da Silva Barros, R.G.; Borges, P.A.; Prescilio, I.C.; de Oliveira, M.R. Resveratrol and Brain Mitochondria: a Review. Mol. Neurobiol. 2018, 55, 2085–2101. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, K.; Wan, W.; Cheng, Y.; Pu, X.; Ye, X. Resveratrol provides neuroprotection by regulating the JAK2/STAT3/PI3K/AKT/mTOR pathway after stroke in rats. Genes Dis. 2018, 5, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Ramírez, N.; Alquisiras-Burgos, I.; Ortiz-Plata, A.; Ruiz-Tachiquín, M.E.; Espinoza-Rojo, M.; Aguilera, P. Resveratrol Activates Neuronal Autophagy Through AMPK in the Ischemic Brain. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelle, M.G.; Wahla, D.; Dieguezb, C.; Berniera, M.; de Caboa, R. Resveratrol supplementation, where are we now and where should we go? Ageing Res Rev 2015, 21, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.; Krishan, P.; Singh, N.; Kumar, A.; Shri, R. Amelioration of ischemia-reperfusion induced functional and biochemical deficit in mice by Ocimum kilimandscharicum leaf extract. Biomed. Pharmacother. 2017, 85, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Bora, K.S.; Arora, S.; Shri, R. Role of Ocimum basilicum L. in prevention of ischemia and reperfusion-induced cerebral damage, and motor dysfunctions in mice brain. J. Ethnopharmacol. 2011, 137, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Angeles-López, G.E.; González-Trujano, M.E.; Déciga-Campos, M.; Ventura-Martínez, R. Neuroprotective evaluation of tilia americana and annona diversifolia in the neuronal damage induced by intestinal ischemia. Neurochem. Res. 2013, 38, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- De-la-Cruz-Chacón, I. La familia Annonaceae Juss. en México. Rev. Ciencias UNICACH 2016, 10, 71–82. [Google Scholar]
- Yeh, S.H.; Chang, F.R.; Wu, Y.C.; Yang, Y.L.; Zhuo, S.K.; Hwang, T.L. An anti-inflammatory ent-kaurane from the stems of Annona squamosa that inhibits various human neutrophil functions. Planta Med. 2005, 71, 904–909. [Google Scholar] [CrossRef]
- Peters, O.; Back, T.; Lindauer, U.; Busch, C.; Megow, D.; Dreier, J.; Dirnagl, U. Increased Formation of Reactive Oxygen Species After Permanent and Reversible Middle Cerebral Artery Occlusion in the Rat. J. Cereb. Blood Flow Metab. 1998, 18, 196–205. [Google Scholar] [CrossRef] [Green Version]
- González-Trujano, M.E.; Martínez, A.L.; Reyes-Ramírez, A.; Reyes-Trejo, B.; Navarrete, A. Palmitone isolated from Annona diversifolia induces an anxiolytic-like effect in mice. Planta Med. 2006, 72, 703–707. [Google Scholar] [CrossRef]
- Lis-Balchin, M. Aromatherapy science: A guide for healthcare professionals; Pharmaceutical Press: London, UK, 2005. [Google Scholar]
- Fiz, O.; Vargas, P.; Alarcón, M.; Aedo, C.; García, J.L.; Aldasoro, J.J. Phylogeny and Historical Biogeography of Geraniaceae in Relation to Climate Changes and Pollination Ecology. Syst. Bot. 2008, 33, 326–342. [Google Scholar] [CrossRef] [Green Version]
- Calzada, F.; Cedillo-Rivera, R.; Bye, R.; Mata, R. Geranins C and D, Additional New Antipropozoal A-Type Proanthocyanidins from Geranium niveum. Planta Med 2001, 67, 677–680. [Google Scholar] [CrossRef]
- Calzada, F.; Cervantes-Martínez, J.A.; Yépez-Mulia, L. In vitro antiprotozoal activity from the roots of Geranium mexicanum and its constituents on Entamoeba histolytica and Giardia lamblia. J. Ethnopharmacol. 2005, 98, 191–193. [Google Scholar] [CrossRef]
- Ashafaq, M.; Raza, S.S.; Khan, M.M.; Ahmad, A.; Javed, H.; Ahmad, M.E.; Tabassum, R.; Islam, F.; Siddiqui, M.S.; Safhi, M.M.; et al. Catechin hydrate ameliorates redox imbalance and limits inflammatory response in focal cerebral ischemia. Neurochem. Res. 2012, 37, 1747–1760. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Su, Y.; Wei, D.; Qian, L.; Xing, D.; Pan, J.; Chen, Y.; Huang, M. Catechin relieves hypoxia/reoxygenation-induced myocardial cell apoptosis via down-regulating lncRNA MIAT. J. Cell. Mol. Med. 2020, 24, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Bentham, G. Labiatae. In Genera Plantarum; Bentham, G., Hoocker, J., Eds.; Taylor and Francis, Ltd.: London, UK, 1876; Volume 2. [Google Scholar]
- Walker, J.; Sytsma, K.; Treutlein, J.; Wink, M. Salvia (Lamiaceae) is not monophyletic: implications for the systematics, radiation, and ecological specializations of Salvia and tribe Mentheae. Am. J. Bot. 2004, 91, 1115–1125. [Google Scholar] [CrossRef] [PubMed]
- Calzada, F.; Bautista, E.; Yépez-Mulia, L.; García-Hernandez, N.; Ortega, A. Antiamoebic and antigiardial activity of clerodane diterpenes from Mexican Salvia species used for the treatment of diarrhea. Phyther. Res. 2015, 29, 1600–1604. [Google Scholar] [CrossRef]
- Chen, C.; Xi, C.; Liang, X.; Ma, J.; Su, D.; Abel, T.; Liu, R. The Role of κ Opioid Receptor in Brain Ischemia. Crit. Care Med. 2016, 44, e1219–e1225. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, A.; Camacho, J.; Chino, S.; Jacque, P.; Lopez, M. Herbario medicinal del Instituto Mexicano del Seguro Social; Información Etnobotánica; IMSS: Mexico City, Mexico, 1994. [Google Scholar]
- Yan, E.B.; Unthank, J.K.; Castillo-melendez, M.; Miller, S.L.; Langford, J.; Walker, D.W. Novel method for in vivo hydroxyl radical measurement by microdialysis in fetal sheep brain in utero. J. Appl. Physiol. 2005, 98, 2304–2310. [Google Scholar] [CrossRef] [Green Version]
- Sachs, J.; McArthur, J.; Schmidt-Traub, G.; Bahadur, C.; Faye, M.; Kruk, M. Millennium Development Goals Needs Assessments: Country Case Studies of Bangladesh, Cambodia, Ghana, Tanzania and Uganda; Millenium Project: Washington, DC, USA, 2004; p. 225. [Google Scholar]
- Oberley, L.W.; Spitz, D.R. Assay of Superoxide Dismutase Activity in Tumor Tissue. Methods Enzymol. 1984, 105, 457–464. [Google Scholar]
- Fath, T.; Ke, Y.D.; Gunning, P.; Götz, J.; Ittner, L.M. Primary support cultures of hippocampal and substantia nigra neurons. Nat. Protoc. 2009, 4, 78–85. [Google Scholar] [CrossRef]
- Longa, E.Z.; Weinstein, P.; Carlson, S.; Cummins, R. Reversible Middle Cerebral Artery Oclussion Without Craniectomy in Rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Rasband, W.S. ImageJ; U. S. National Institutes of Health (NIH): Bethesda, MD, USA, 1997.
- Roof, R.L.; Schielke, G.P.; Ren, X.; Hall, E.D. A Comparison of Long-Term Functional Outcome After 2 Middle Cerebral Artery Occlusion Models in Rats. Stroke 2001, 32, 2648–2657. [Google Scholar]
- Kelsey, N.A.; Wilkins, H.M.; Linseman, D.A. Neutraceutical Antioxidants as Novel Neuroprotective Agents. Molecules 2015, 15, 7792–7814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant properties of resveratrol: A structure-activity insight. Innov. Food Sci. Emerg. Technol. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Gatz, S.A.; Wiesmüller, L. Take a break - Resveratrol in action on DNA. Carcinogenesis 2008, 29, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H. Bin Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8. [Google Scholar]
- Iuga, C.; Alvarez-Idaboy, J.R.; Russo, N. Antioxidant activity of trans -resveratrol toward hydroxyl and hydroperoxyl radicals: A quantum chemical and computational kinetics study. J. Org. Chem. 2012, 77, 3868–3877. [Google Scholar] [CrossRef]
- Agu, K.C.; Okolie, P.N. Proximate composition, phytochemical analysis, and in vitro antioxidant potentials of extracts of Annona muricata (Soursop). Food Sci. Nutr. 2017, 5, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Schichiri, M. The role of lipid peroxidation in neurological disorders. J. Clin. Biochem. Nutr. 2014, 54, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; Silva, N.M.D.; Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against α-amylase, α-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Kandimalla, R.; Dash, S.; Kalita, S.; Choudhury, B.; Malampati, S.; Kalita, K.; Kotoky, J. Bioactive guided fractions of Annona reticulata L. bark: Protection against liver toxicity and inflammation through inhibiting oxidative stress and proinflammatory cytokines. Front. Pharmacol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzada, F.; Correa-Basurto, J.; Barbosa, E.; Mendez-Luna, D.; Yepez-Mulia, L. Antiprotozoal Constituents from Annona cherimola Miller, a Plant Used in Mexican Traditional Medicine for the Treatment of Diarrhea and Dysentery. Pharmacogn. Mag. 2017, 13, 148–152. [Google Scholar]
- Durant, A.; Murillo, E.; Britton, G. Antioxidant activity and polyphenol content in cultivated and wild edible fruits grown in Panama. J. Pharm. Bioallied Sci. 2012, 4, 313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chinese Med. (United Kingdom) 2018, 13, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Zhu, H.; Misra, B.R.; Mahaney, J.E.; Li, Y.; Misra, H.P. EPR studies on the superoxide-scavenging capacity of the nutraceutical resveratrol. Mol Cell Biochem. 2008, 313, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Sueishi, Y.; Nii, R.; Kakizaki, N. Resveratrol analogues like piceatannol are potent antioxidants as quantitatively demonstrated through the high scavenging ability against reactive oxygen species and methyl radical. Bioorganic Med. Chem. Lett. 2017, 27, 5203–5206. [Google Scholar] [CrossRef] [PubMed]
- Acésio, N.O.; Carrijo, G.S.; Batista, T.H.; Damasceno, J.L.; Côrrea, M.B.; Tozatti, M.G.; Cunha, W.R.; Tavares, D.C. Assessment of the antioxidant, cytotoxic, and genotoxic potential of the Annona muricata leaves and their influence on genomic stability. J. Toxicol. Environ. Heal. Part A Curr. Issues 2017, 80, 1290–1300. [Google Scholar] [CrossRef]
- Wahab, S.M.A.; Jantan, I.; Haque, M.A.; Arshad, L. Exploring the leaves of Annona muricata L. as a source of potential anti-inflammatory and anticancer agents. Front. Pharmacol. 2018, 9, 1–20. [Google Scholar]
- Şöhretoğlu, D.; Genç, Y.; Harput, Ş. Comparative evaluation of phenolic profile, antioxidative and cytotoxic activities of different Geranium species. Iran. J. Pharm. Res. 2017, 16, 178–187. [Google Scholar]
- Xu, J.; Wei, K.; Zhang, G.; Lei, L.; Yang, D.; Wang, W.; Han, Q.; Xia, Y.; Bi, Y.; Yang, M.; et al. Ethnopharmacology, phytochemistry, and pharmacology of Chinese Salvia species: A review. J. Ethnopharmacol. 2018, 225, 18–30. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, W.; Xu, L.; Chen, L. In Salvia miltiorrhiza, phenolic acids possess protective properties against amyloid β-induced cytotoxicity, and tanshinones act as acetylcholinesterase inhibitors. Environ. Toxicol. Pharmacol. 2011, 31, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Bautista, E.; Ortiz-Pastrana, N.; Pastor-Palacios, G.; Montoya-Contreras, A.; Toscano, R.A.; Morales-Jiménez, J.; Salazar-Olivo, L.A.; Ortega, A. Neo-Clerodane Diterpenoids from Salvia polystachya Stimulate the Expression of Extracellular Matrix Components in Human Dermal Fibroblasts. J. Nat. Prod. 2017, 80, 3003–3009. [Google Scholar] [CrossRef] [PubMed]
- Jeanne, M.; Jorgensen, J.; Gould, D.B.; Francisco, S. Identify Mechanisms for Stroke Prevention. Circulation 2016, 131, 1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.Y.; Fan, J.Y.; Horie, Y.; Miura, S.; Cui, D.H.; Ishii, H.; Hibi, T.; Tsuneki, H.; Kimura, I. Ameliorating effects of compounds derived from Salvia miltiorrhiza root extract on microcirculatory disturbance and target organ injury by ischemia and reperfusion. Pharmacol. Ther. 2008, 117, 280–295. [Google Scholar] [CrossRef]
- Li, R.; Morris-Natschke, S.L.; Lee, K.H. Clerodane diterpenes: Sources, structures, and biological activities. Nat. Prod. Rep. 2016, 33, 1166–1226. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Botanical Species (Family) | Voucher Specimen | Parts Used a | Yield [% (w/w)] | Common Name |
|---|---|---|---|---|---|
| Ach | Annona cherimola Miller (Annonaceae) | 15795 | L | 4.5 | cherimoya |
| Ad | Annona diversifolia Safford (Annonaceae) | 16248 | L | 5.9 | ilama |
| Amu | Annona muricata Linn (Annonaceae) | 15943 | L | 15.4 | Soursop |
| Ap | Annona purpurea Moc & Sesse (Annonaceae) | 16293 | L | 9.4 | toreta |
| Are | Annona reticulata Linn (Annonaceae) | 15944 | L | 5.2 | custard apple |
| GmPA | Geranium mexicanum H. B & K (Geraniaceae) | 14405 | AP | 15.5 | geranium |
| GnPA | Geranium niveum S. Watson (Geraniaceae) | 36899 | AP | 5.5 | geranium |
| Sa | Salvia amaríssima Ortega (Lamiaceae) | 16263 | AP | 3.8 | sage |
| Sp | Salvia polystachya Ortega (Lamiaceae) | 16386 | AP | 7.3 | sage |
| Extracts/Standards | O2•− | OH• | ROO• |
|---|---|---|---|
| Ach | X | 155.04 ± 4.40 | 102.43 ± 42.92 |
| Ad | X | 131.39 ± 26.77 | 27.14 ± 8.92 |
| Amu | X | 98.96 ± 19.58 | 30.42 ± 8.00 |
| Ap | 23.49 ± 9.12 | 151.73 ± 36.54 | 4.48 ± 0.24 |
| Are | X | 126.98 ± 31.60 | 13.13 ± 3.67 |
| GmPA | 26.83 ± 10.52 | 147.14 ± 43.43 | 12.33 ± 4.26 |
| GnPA | 20.27 ± 4.09 | 131.15 ± 40.37 | 10.46 ± 2.47 |
| Sa | 46.64 ± 9.49 | 127.99 ± 36.85 | 24.91 ± 5.88 |
| Sp | 118.97 ± 31.98 | 154.18 ± 52.21 | 18.21 ± 3.05 |
| Resveratrol | 28.28 ± 9.27 | 30.02 ± 6.58 | 0.23 ± 0.06 |
| DMTU | 226.09 ± 45.68 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pineda-Ramírez, N.; Calzada, F.; Alquisiras-Burgos, I.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Ortiz-Plata, A.; Pinzón Estrada, E.; Torres, I.; Aguilera, P. Antioxidant Properties and Protective Effects of Some Species of the Annonaceae, Lamiaceae, and Geraniaceae Families against Neuronal Damage Induced by Excitotoxicity and Cerebral Ischemia. Antioxidants 2020, 9, 253. https://doi.org/10.3390/antiox9030253
Pineda-Ramírez N, Calzada F, Alquisiras-Burgos I, Medina-Campos ON, Pedraza-Chaverri J, Ortiz-Plata A, Pinzón Estrada E, Torres I, Aguilera P. Antioxidant Properties and Protective Effects of Some Species of the Annonaceae, Lamiaceae, and Geraniaceae Families against Neuronal Damage Induced by Excitotoxicity and Cerebral Ischemia. Antioxidants. 2020; 9(3):253. https://doi.org/10.3390/antiox9030253
Chicago/Turabian StylePineda-Ramírez, Narayana, Fernando Calzada, Iván Alquisiras-Burgos, Omar Noel Medina-Campos, José Pedraza-Chaverri, Alma Ortiz-Plata, Enrique Pinzón Estrada, Ismael Torres, and Penélope Aguilera. 2020. "Antioxidant Properties and Protective Effects of Some Species of the Annonaceae, Lamiaceae, and Geraniaceae Families against Neuronal Damage Induced by Excitotoxicity and Cerebral Ischemia" Antioxidants 9, no. 3: 253. https://doi.org/10.3390/antiox9030253