Cell-Type-Specific Modulation of Hydrogen Peroxide Cytotoxicity and 4-Hydroxynonenal Binding to Human Cellular Proteins In Vitro by Antioxidant Aloe vera Extract

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extract Preparation
2.2. Total Soluble Phenolic Content and Antioxidant Activity
2.2.1. Phenolic Content
2.2.2. Ascorbic Acid Content
2.2.3. Measurement of Total Carotenoids
2.2.4. Antioxidant Activity
2.3. Cell Cultures and Treatments
2.3.1. Cell Cultures
2.3.2. Experimental Treatments with AV, H2O2, and HNE
2.3.3. Determination of Intracellular HNE–Protein Adducts Using Cell-Based HNE–His ELISA
2.3.4. Measurement of Intracellular ROS Production
3. Results
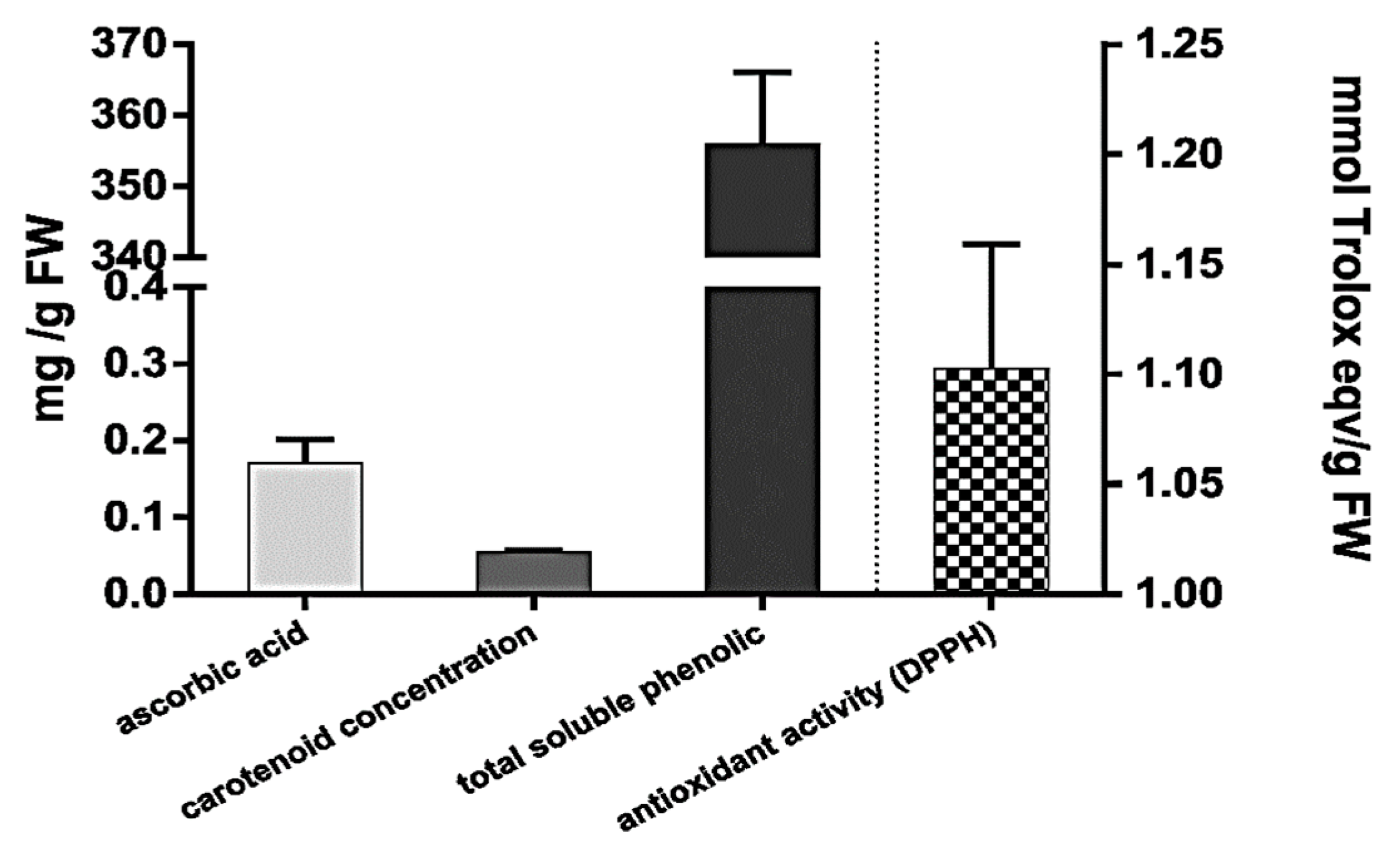
3.1. The Levels of Antioxidants in A. vera Leaves
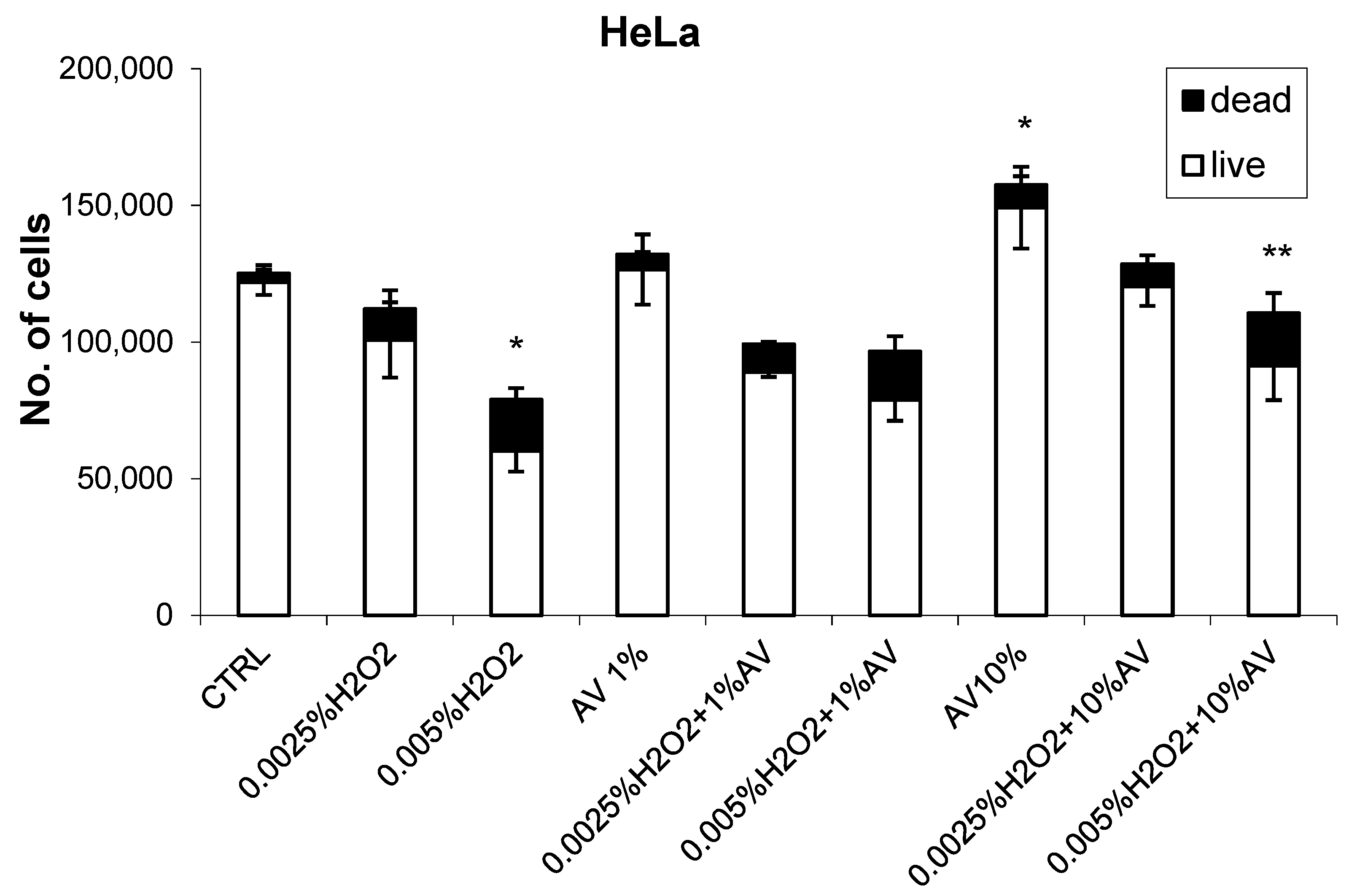
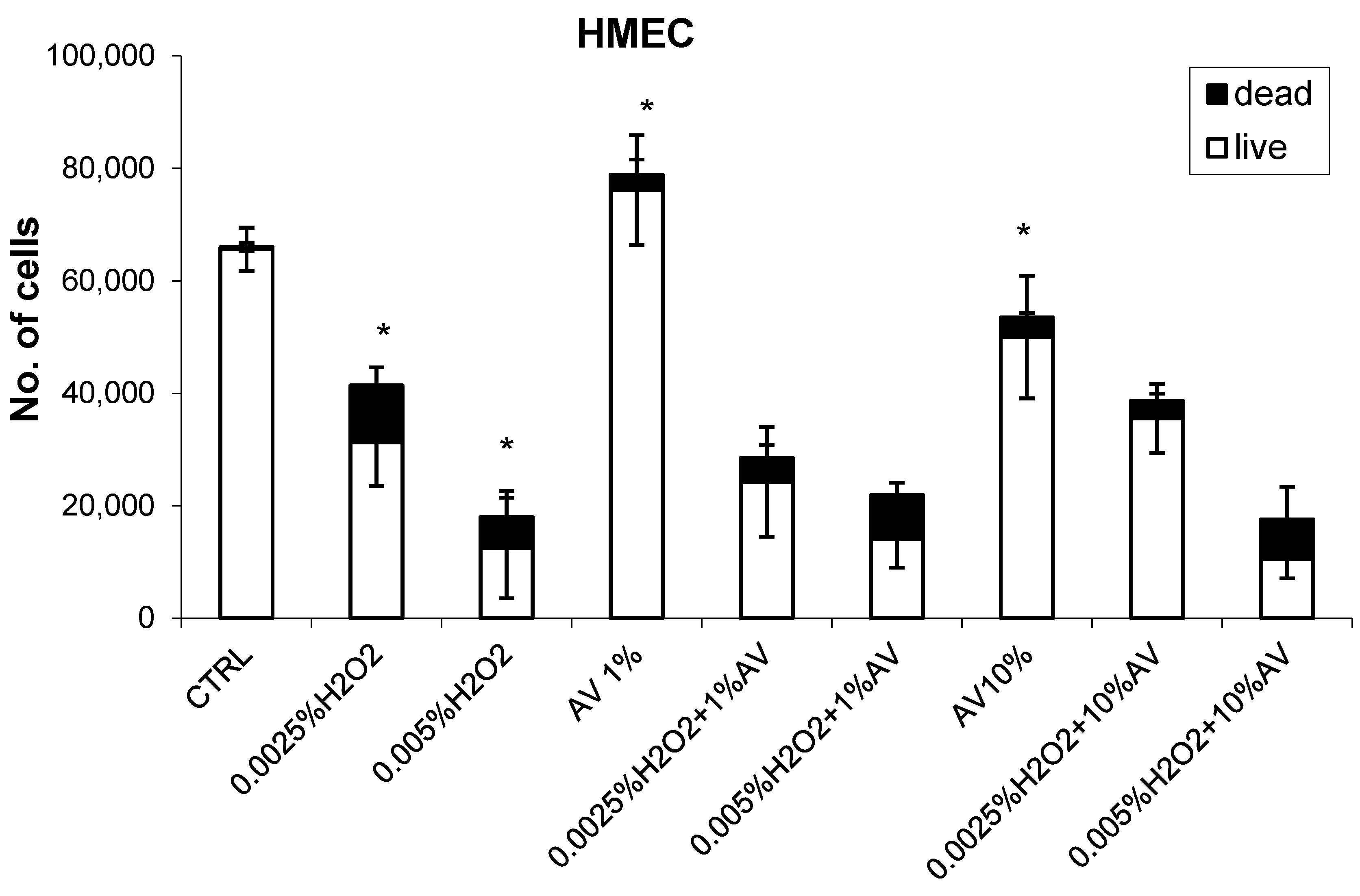
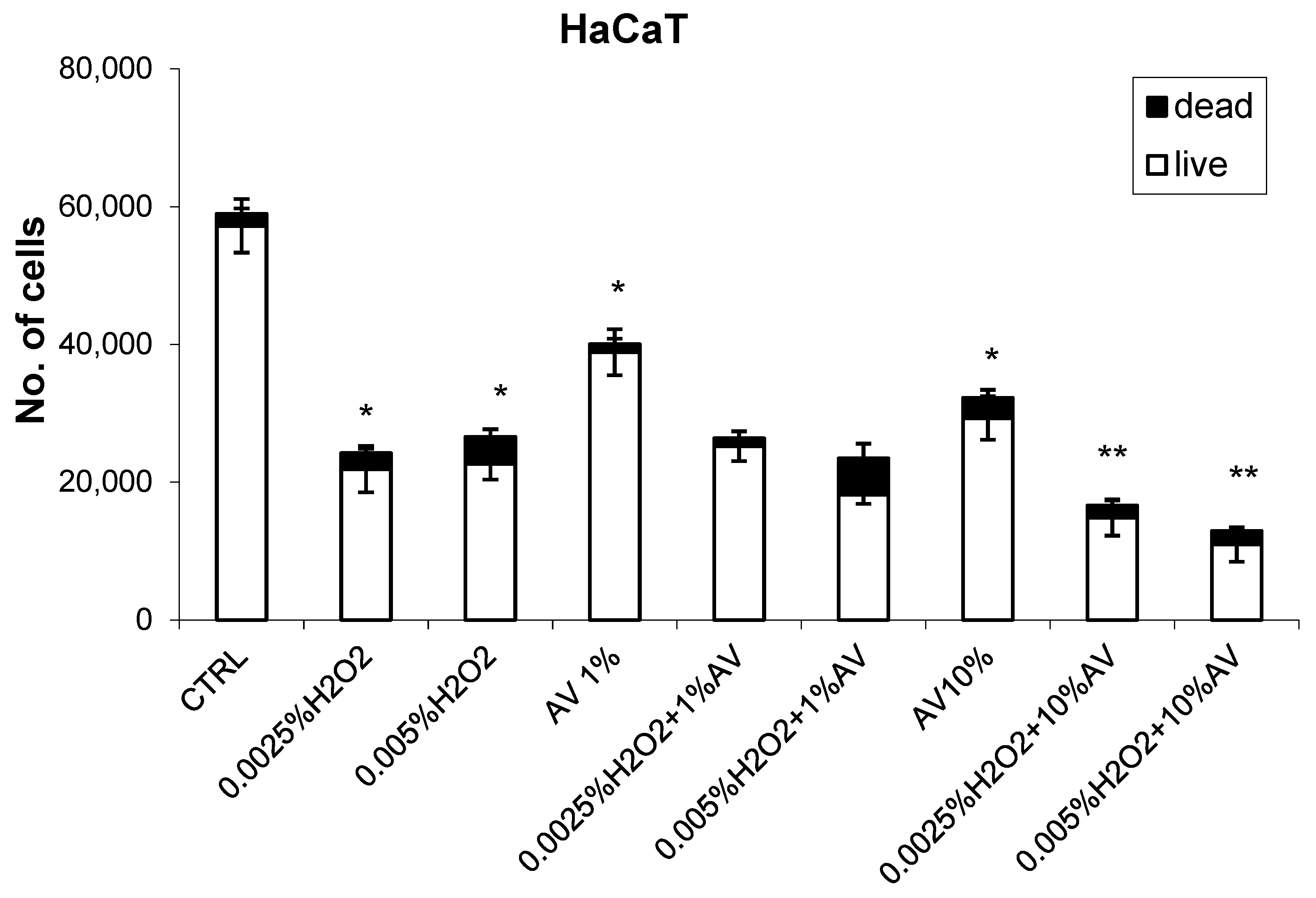
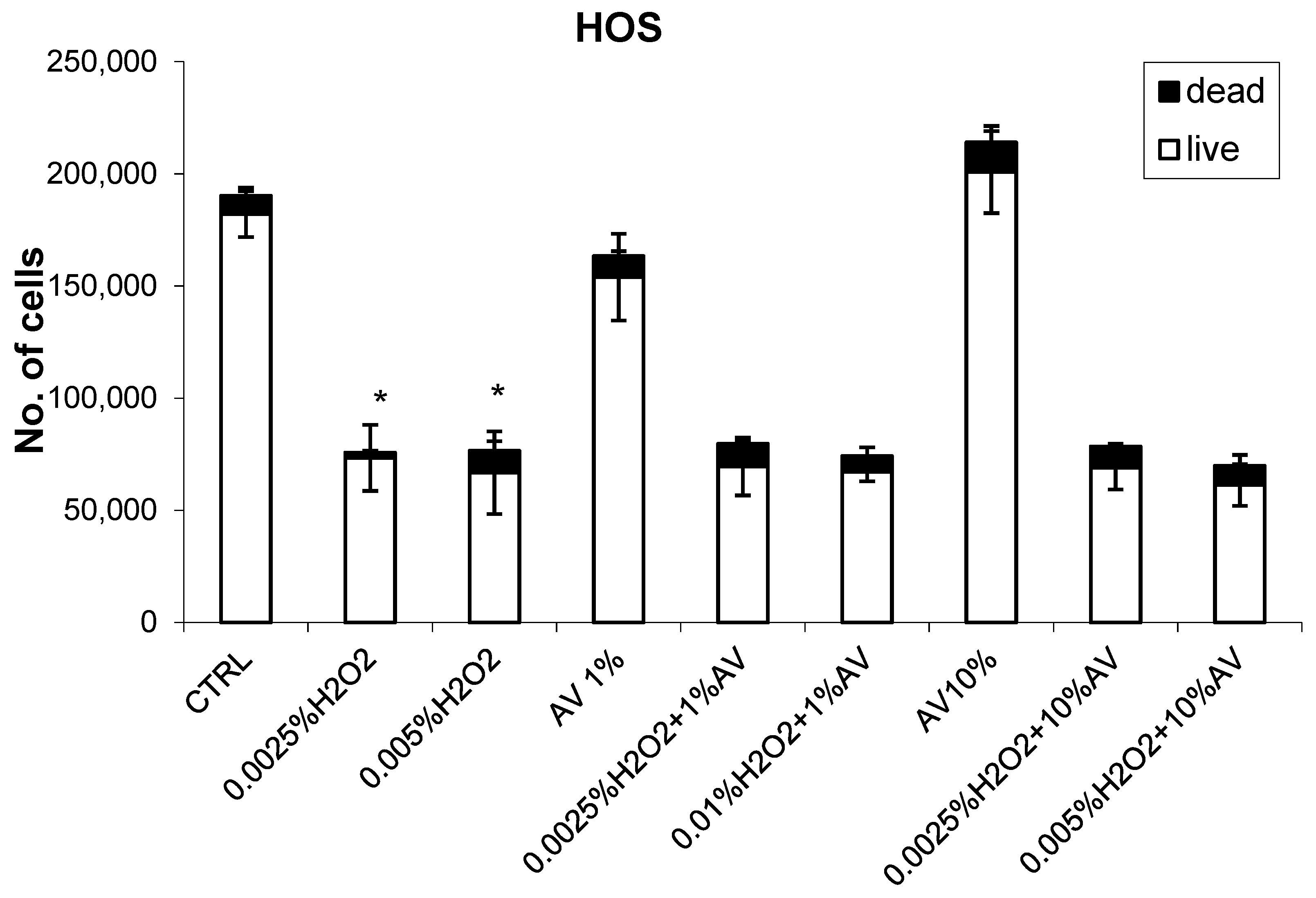
3.2. The Effects of H2O2 and AV Extract on Cell Viability
3.3. The Effects of AV Extract on HNE-Pretreated Cells
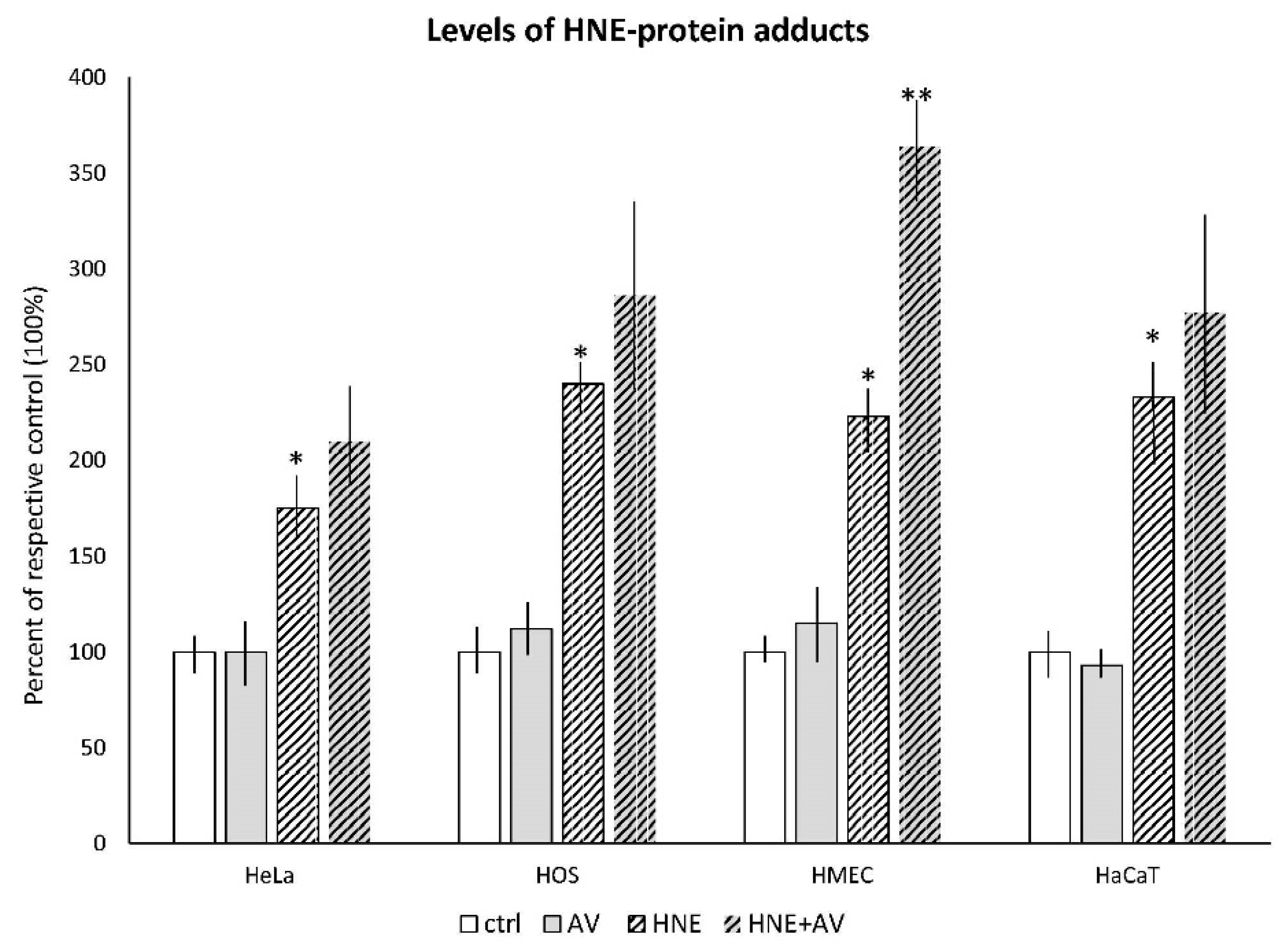
3.3.1. The Effects AV on HNE Binding to Cellular Proteins
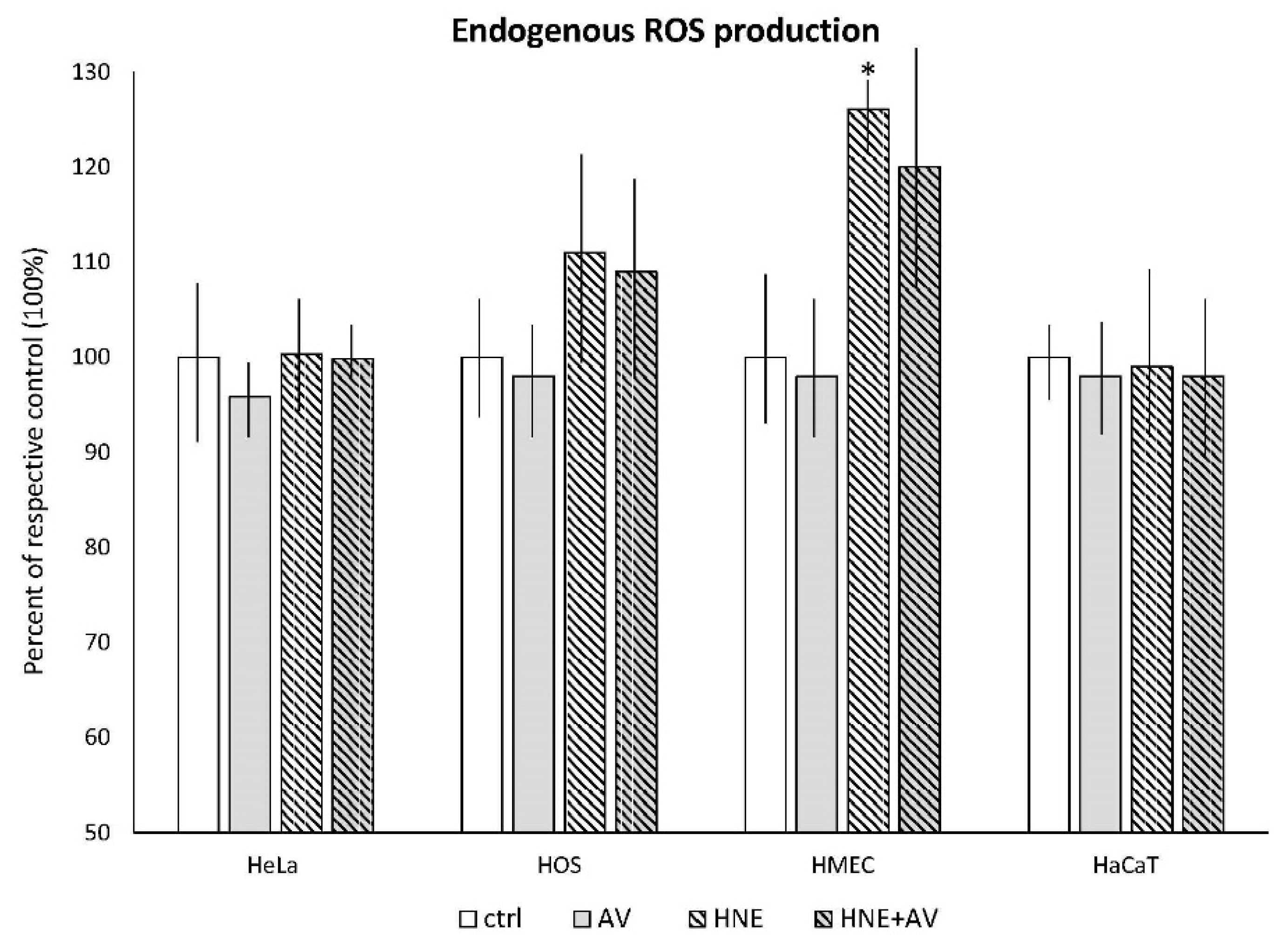
3.3.2. The Effects of AV on Cellular ROS Production Induced by HNE
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgment
Conflicts of Interest
References
- Radha, M.H.; Laxmipriya, N.P. Evaluation of biological properties and clinical effectiveness of Aloe vera: A systematic review. J. Tradit. Complement. Med. 2015, 5, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Eshun, K.; He, Q. Aloe vera: A valuable ingredient for the food, pharmaceutical and cosmetic industries—A review. Crit. Rev. Food Sci. Nutr. 2004, 44, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Hamman, J.H. Composition and applications of Aloe vera leaf gel. Molecules 2008, 13, 1599–1616. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Pellizzoni, M.; Molinari, G.P.; Franchi, F. Aloe anthraquinones against cancer. Med. Aromat. Plant Sci. Biotechnol. 2012, 6, 20–24. [Google Scholar]
- Choi, S.; Chung, M.H. A review on the relationship between Aloe vera components and their biologic effects. Semin. Integr. Med. 2003, 1, 53–62. [Google Scholar] [CrossRef]
- Bourdeau, M.D.; Beland, F.A. An evaluation of the biological and toxicological properties of Aloe barbadensis (Miller), Aloe vera. J. Environ. Sci. Health C 2006, 24, 103–154. [Google Scholar]
- Zarkovic, N. 4-Hydroxynonenal as a bioactive marker of pathophysiological processes. Mol. Asp. Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Vistoli, G.; Maddis, D.D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Zarkovic, N. Editorial Introduction to the Special Issue on 4-Hydroxynonenal and Related Lipid Oxidation Products. Free Radic. Biol. Med. 2017, 111, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Sovic, A.; Borović, S.; Lončarić, I.; Kreuzer, T.; Zarkovic, K.; Vukovic, T.; Wäg, G.; Hrascan, R.; Wintersteiger, R.; Klinger, R.; et al. The carcinostatic and proapoptotic potential of 4-Hydroxynonenal in HeLa cells is associated with its conjugation to cellular proteins. Anticancer Res. 2001, 21, 1997–2004. [Google Scholar] [PubMed]
- Borovic Sunjic, S.; Cipak, A.; Rabuzin, F.; Wildburger, R.; Zarkovic, N. The influence of 4-hydroxy-2-nonenal on proliferation, differentiation and apoptosis of human osteosarcoma cells. Biofactors 2005, 24, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N.; Cipak, A.; Jaganjac, M.; Borovic, S.; Zarkovic, K. Pathophysiological relevance of aldehydic protein modifications. J. Proteom. 2013, 92, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Hoppe, A.; Detsch, R.; Boccaccini, A.R.; Zarkovic, N. Effects of Cu-doped 45S5 bioactive glass on the lipid peroxidation-associated growth of human osteoblast-like cells in vitro. J. Biomed. Mater Res. Part A 2014, 102, 3556–3561. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Cipak Gasparovic, A.; Zarkovic, N. Overview on major lipid peroxidation bioactive factor 4-hydroxynonenal as pluripotent growth regulating factor. Free Radic. Res. 2015, 49, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, K.; Jakovcevic, A.; Zarkovic, N. Contribution of the HNE-immunohistochemistry to modern pathological concepts of major human diseases. Free Radic. Biol. Med. 2017, 111, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Randhir, R.; Shetty, K. Developmental stimulation of total phenolics and related antioxidant activity in light-and dark-germinated corn by natural elicitors. Process Biochem. 2005, 40, 1721–1732. [Google Scholar] [CrossRef]
- Bessey, O.A.; Lowky, O.H.; Brock, M.J. A method for the rapid determination of alkaline phosphatase with five cubic millimeters of serum. J. Biol. Chem. 1946, 164, 321–329. [Google Scholar] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. In Current Protocols in Food Analytical Chemistry (CPFA); Wrolstad, R.E., Acree, T.E., An, H., Decker, E.A., Penner, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Sporns, P., Eds.; John Wiley and Sons: New York, NY, USA, 2001; pp. F4.3.1–F4.3.8. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Tech. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Cheregi, M.C.; Danet, A.F. Total antioxidant capacity of some commercial fruit juices: Electrochemical and spectrophotometrical approaches. Molecules 2009, 14, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Tiveron, A.P.; Melo, P.S.; Bergamaschi, K.B.; Vieira, T.M.; Regitano-d’Arce, M.A.; Alencar, S.M. Antioxidant activity of Brazilian vegetables and its relation with phenolic composition. Int. J. Mol. Sci. 2012, 13, 8943–8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borović, S.; Rabuzin, F.; Waeg, G.; Žarković, N. Enzyme-linked immunosorbent assay for 4-hydroxynonenal-histidine conjugates. Free Radic. Res. 2006, 40, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Živković, M.; Žarković, K.; Škrinjar, L.; Georg, W.; Poljak-Blaži, M.; Šunjić, B.S.; Schaur, R.J.; Žarković, N. A new method for detection of HNE-histidine conjugates in rat inflammatory cells. Croat Chem. Acta 2005, 78, 91–98. [Google Scholar]
- Spickett, C.M.; Wiswedel, I.; Siems, W.; Zarkovic, K.; Zarkovic, N. Advances in methods for the determination of biologically relevant lipid peroxidation products. Free Radic. Res. 2010, 44, 1172–1202. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Teklić, T.; Engler, M.; Cesar, V.; Lepeduš, H.; Parađiković, N.; Lončarić, Z.; Štolfa, I.; Marotti, T.; Mikac, N.; Žarković, N. Copper excess influence on lettuce (Lactuca sativa L.) grown in the soil and nutrient solution. J. Food Agric. Environ. 2008, 6, 439–444. [Google Scholar]
- Wonisch, W.; Kohlwein, S.D.; Schaur, J.; Tatzber, F.; Guttenberger, H.; Zarkovic, N.; Winkler, R.; Esterbauer, H. Treatment of the budding yeast (Saccharomyces cerevisiae) with the lipid peroxidation product 4-HNE provokes a temporary cell cycle arrest in G1 phase. Free Radic. Biol. Med. 1998, 25, 682–687. [Google Scholar] [CrossRef]
- Čipak, A.; Jaganjac, M.; Tehlivets, O.; Kohlwein, S.D.; Žarković, N. Adaptation to oxidative stress induced by polyunsaturated fatty acids in yeast. BBA-Mol. Cell Biol. Lipids 2008, 178, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Žarković, N.; Schaur, R.J.; Puhl, H.; Jurin, M.; Esterbauer, H. Mutual dependence of growth modifying effects of 4-hydroxy-nonenal and fetal calf serum in vitro. Free Radic. Biol. Med. 1994, 16, 877–884. [Google Scholar]
- Žarković, N.; Žarković, K.; Schaur, R.J.; Štolc, S.; Schlag, G.; Redl, H.; Waeg, G.; Borović, S.; Lončarić, L.; Jurić, G.; et al. 4-Hydroxynonenal as a second messenger of free radicals and growth modifying factor. Life Sci. 1999, 65, 1901–1904. [Google Scholar] [CrossRef]
- Semlitsch, T.; Tillian, M.H.; Žarković, N.; Borović, S.; Purtscher, M.; Hohenwarter, O.; Schaur, J.R. Differential Influence of the Lipid Peroxidation Product 4-Hydroxynonenal on the Growth of Human Lymphatic Leukaemia Cells and Human Peripheral Blood Lymphocytes. Anticancer Res. 2002, 22, 1689–1697. [Google Scholar] [PubMed]
- Borović, S.; Čipak, A.; Meinitzer, A.; Kejla, Z.; Perovic, D.; Waeg, G.; Zarkovic, N. Differential effect of 4-hydroxynonenal on normal and malignant mesenchimal cells. Redox Rep. 2007, 207, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Mouthuy, P.A.; Snelling, S.J.B.; Dakin, S.G.; Milković, L.; Gašparović, A.C.; Carr, A.J.; Žarković, N. Biocompatibility of implantable materials: An oxidative stress viewpoint. Biomaterials 2016, 109, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Cipak-Gasparovic, A.; Milkovic, L.; Borovic-Sunjic, S.; Zarkovic, N. Cancer Growth Regulation by 4-Hydroxynonenal Article Type. Free Radic. Biol. Med. 2017, 111, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G.; Zarkovic, N. Revealing mechanisms of selective, concentration-dependent potentials of 4-hydroxy-2-nonenal to induce apoptosis in cancer cells through inactivation of membrane-associated catalase. Free Radic. Biol. Med. 2015, 81, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Xiao, M.; Zarkovic, K.; Zhu, M.; Sa, R.; Lu, J.; Tao, Y.; Chen, Q.; Xia, L.; Cheng, S.; et al. Mitochondrial Control of Apoptosis through Modulation of Cardiolipin Oxidation in Hepatocellular Carcinoma: A Novel Link between Oxidative Stress and Cancer. Free Radic. Biol. Med. 2017, 176, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Piskač Živković, N.; Petrovečki, M.; Lončarić, T.Č.; Nikolić, I.; Waeg, G.; Jaganjac, M.; Žarković, K.; Žarković, N. Positron Emission Tomography-Computed Tomography and 4-Hydroxynonenal-histidine Immunohistochemistry Reveal Differential Onset of Lipid Peroxidation in Primary Lung Cancer and in Pulmonary Metastasis of Remote Malignancies. Redox Biol. 2017, 11, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Negre-Salvayre, A.; Auge, N.; Ayala, V.; Basaga, H.; Boada, J.; Brenke, R.; Chapple, S.; Cohen, G.; Feher, J.; Grune, T.; et al. Pathological aspects of lipid peroxidation. Free Radic. Res. 2010, 44, 1125–1171. [Google Scholar] [CrossRef] [PubMed]
- Kujundžić, R.N.; Žarković, N.; Trošelj, K.G. Pyridine nucleotides in regulation of cell death and survival by redox and non-redox reactions. Crit. Rev. Eukar. Gene Express. 2014, 24, 287–309. [Google Scholar] [CrossRef]
- Milkovic, L.; Siems, W.; Siems, R.; Zarkovic, N. Oxidative stress and antioxidants in carcinogenesis and integrative therapy of cancer. Curr. Pharm. Des. 2014, 20, 6529–6542. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Zarkovic, N.; Saso, L. Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol. 2017, 12, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Pecere, T.; Gazzola, M.V.; Mucignat, C.; Parolin, C.; Vecchia, F.D.; Cavaggioni, A.; Basso, G.; Diaspro, A.; Salvato, B.; Carli, M.; et al. Aloe-emodin is a new type of anticancer agent with selective activity against neuroectodermal tumors. Cancer Res. 2000, 60, 2800–2804. [Google Scholar] [PubMed]
- Grimaudo, S.; Tolomeo, M.; Gancitano, R.; Dalessandro, N.; Aiello, E. Effects of highly purified anthraquinoid compounds from Aloe vera on sensitive and multidrug resistant leukemia cells. Oncol. Rep. 1997, 4, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Z.; Hsu, S.L.; Liu, M.C.; Wu, C.H. Effects and mechanisms of aloe-emodin on cell death in human lung squamous cell carcinoma. Eur. J. Pharmacol. 2001, 431, 287–295. [Google Scholar] [CrossRef]
- Jaganjac, M.; Poljak-Blazi, M.; Schaur, R.J.; Zarkovic, K.; Borović, S.; Čipak, A.; Cindrić, M.; Uchida, K.; Waeg, G.; Žarković, N. Elevated neutrophil elastase and acrolein-protein adducts are associated with W256 regression. Clin. Exp. Immunol. 2012, 170, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasi, F.; Tessitore, L.; Zanetti, D.; Citrin, J.C.; Zingaro, B.; Chiarpotto, E.; Zarkovic, N.; Serviddio, G.; Poli, G. Associated changes of lipid peroxidation and TGF 1 levels in human cancer during tumor progression. Gut 2002, 50, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Žarković, K.; Juric, G.; Waeg, G.; Kolenc, D.; Žarković, N. Immunohistochemical appearance of HNE-protein conjugates in human astrocytomas. Biofactors 2005, 24, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Harlev, E.; Nevo, E.; Lansky, E.P.; Ofir, R.; Bishayee, A. Anticancer potential of Aloes: Antioxidant, antiproliferative, and immunostimulatory attributes. Planta. Med. 2012, 78, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gęgotek, A.; Nikliński, J.; Žarković, N.; Žarković, K.; Waeg, G.; Łuczaj, W.; Charkiewicz, R.; Skrzydlewska, E. Lipid mediators involved in the oxidative stress and antioxidant defence of human lung cancer cells. Redox Biol. 2016, 9, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, M.; Zarkovic, N. Preface to the special issue on 4-hydroxynonenal and related lipid oxidation products. Free Radic. Biol. Med. 2017, 111, 1. [Google Scholar] [CrossRef] [PubMed]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European Contribution to the study of ROS: A Summary of the Findings and Prospects for the Future from the COST Action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cesar, V.; Jozić, I.; Begović, L.; Vuković, T.; Mlinarić, S.; Lepeduš, H.; Borović Šunjić, S.; Žarković, N. Cell-Type-Specific Modulation of Hydrogen Peroxide Cytotoxicity and 4-Hydroxynonenal Binding to Human Cellular Proteins In Vitro by Antioxidant Aloe vera Extract. Antioxidants 2018, 7, 125. https://doi.org/10.3390/antiox7100125
Cesar V, Jozić I, Begović L, Vuković T, Mlinarić S, Lepeduš H, Borović Šunjić S, Žarković N. Cell-Type-Specific Modulation of Hydrogen Peroxide Cytotoxicity and 4-Hydroxynonenal Binding to Human Cellular Proteins In Vitro by Antioxidant Aloe vera Extract. Antioxidants. 2018; 7(10):125. https://doi.org/10.3390/antiox7100125
Chicago/Turabian StyleCesar, Vera, Iva Jozić, Lidija Begović, Tea Vuković, Selma Mlinarić, Hrvoje Lepeduš, Suzana Borović Šunjić, and Neven Žarković. 2018. "Cell-Type-Specific Modulation of Hydrogen Peroxide Cytotoxicity and 4-Hydroxynonenal Binding to Human Cellular Proteins In Vitro by Antioxidant Aloe vera Extract" Antioxidants 7, no. 10: 125. https://doi.org/10.3390/antiox7100125