
Protracted Oxidative Alterations in the Mechanism of Hematopoietic Acute Radiation Syndrome

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

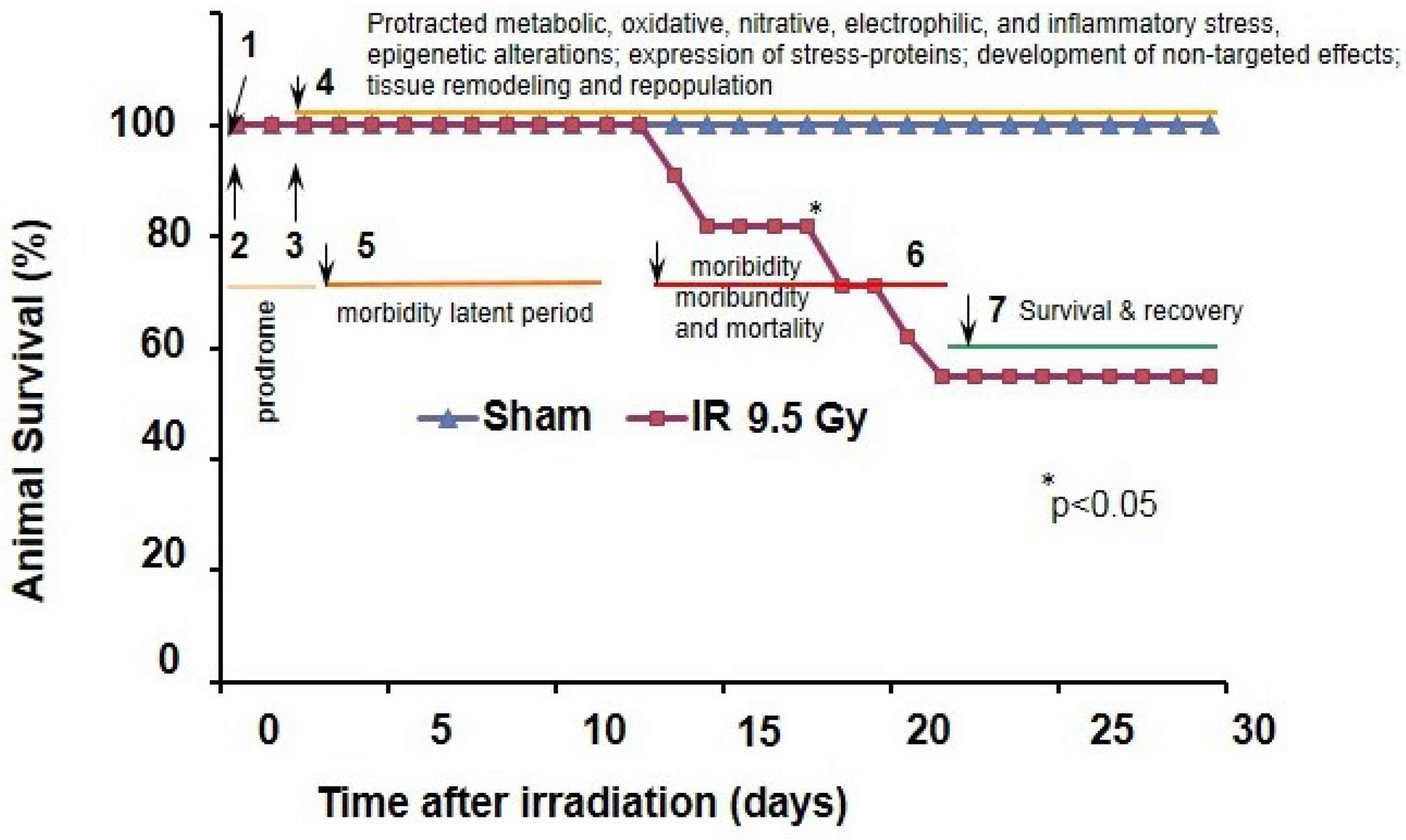
2. Radiation-Related Multistage Activation of Oxidative Reactions and Their Role in hARS

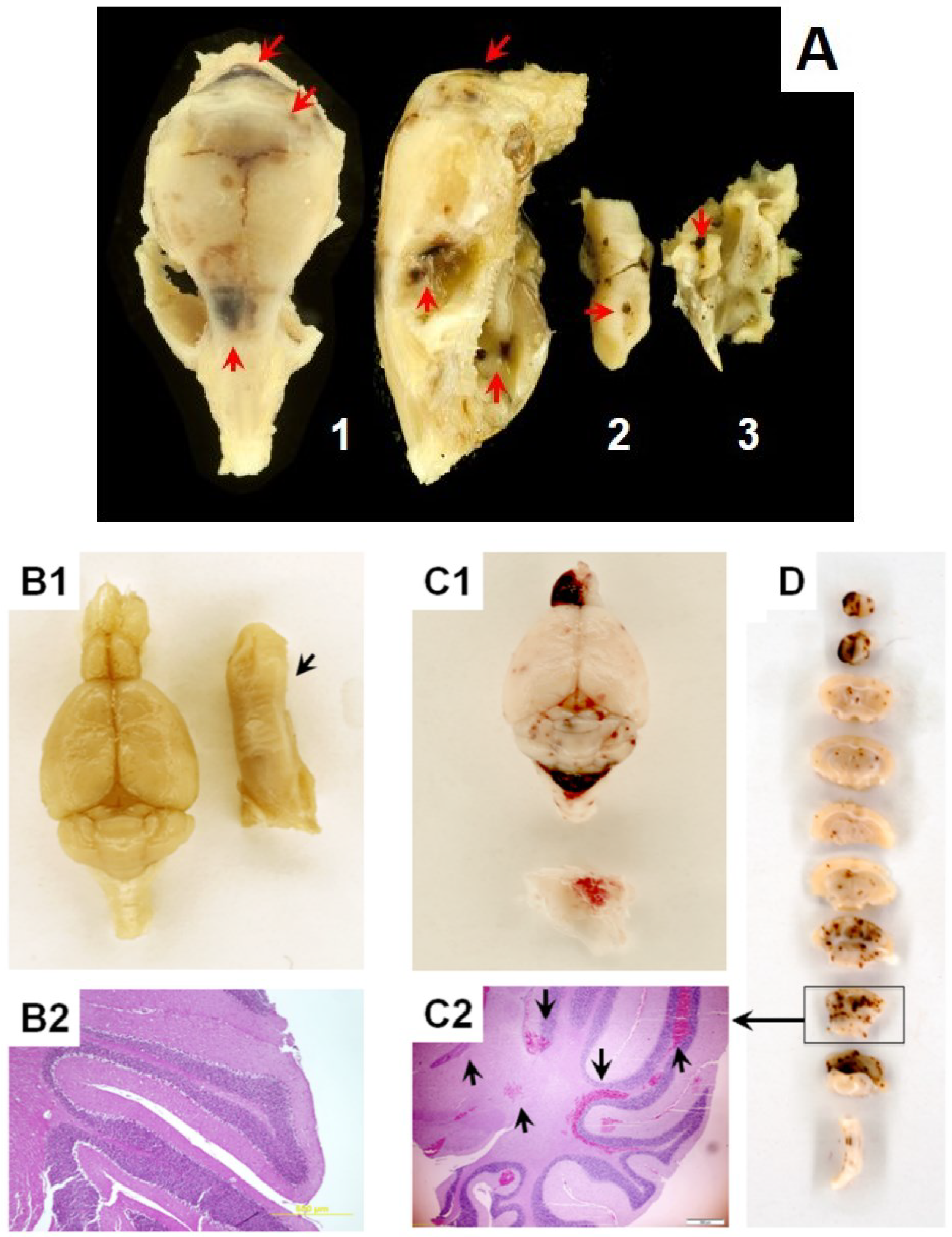
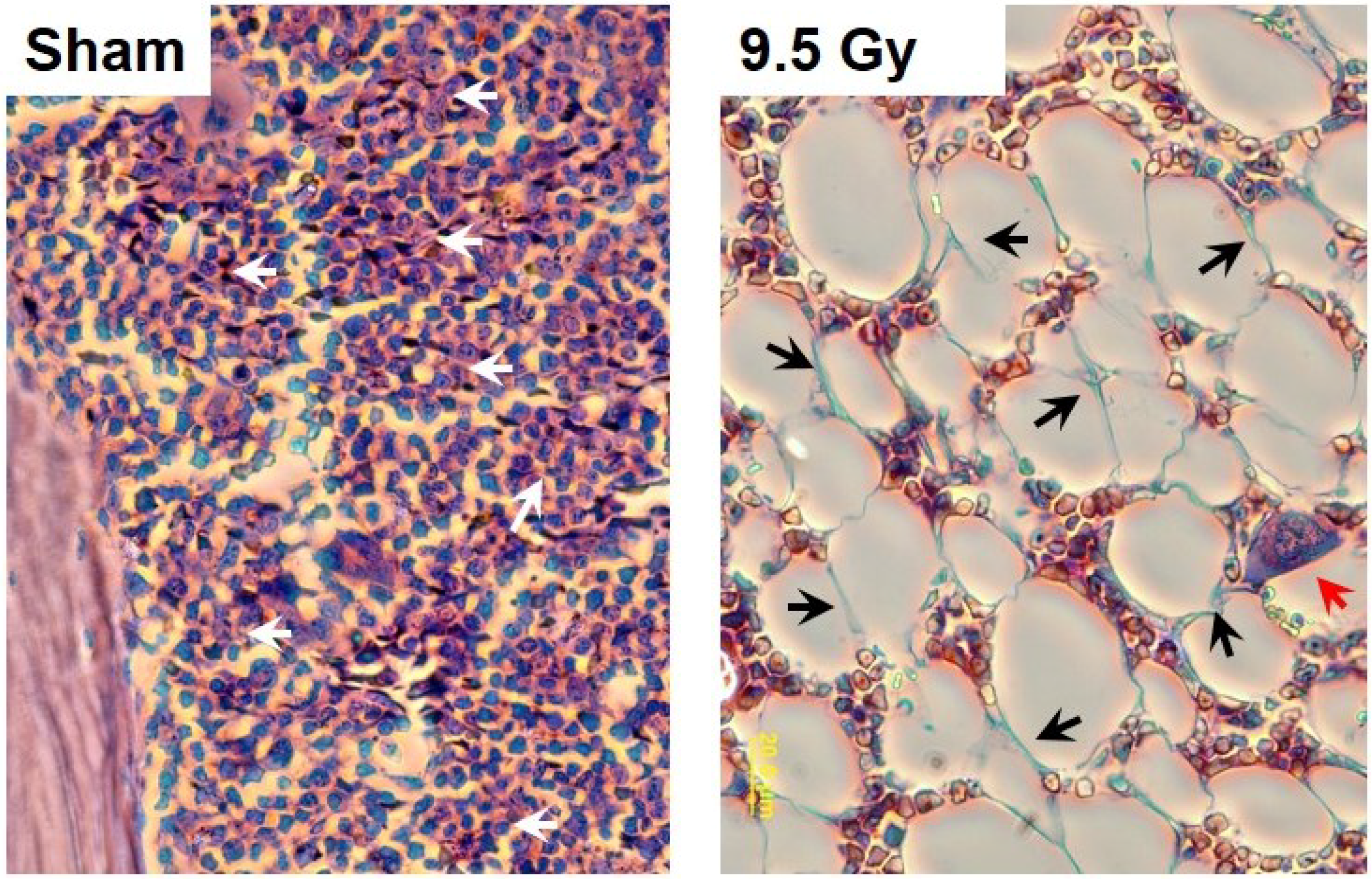
3. Radiation-Induced Oxidative Reactions and Related Alterations in Tissue Barriers

4. Conclusions
Conflicts of Interest
References
- Lawrence, J.H.; Tennant, R. The comparative effects of neutrons and X-rays on the whole body. J. Exp. Med. 1937, 66, 667–688. [Google Scholar] [CrossRef] [PubMed]
- Grahn, D. Acute radiation response of mice from a cross between radiosensitive and radioresistant strains. Genetics 1958, 43, 835–843. [Google Scholar] [PubMed]
- Mettler, F.A., Jr.; Upton, A.C. Radiation effects on cell and organ systems. In Medical Effects of Ionizing Radiation, 3rd ed.; Mettler, F.A., Upton, W.B., Eds.; Elsevier: Philadelphia, PA, USA, 2008; pp. 17–21. [Google Scholar]
- Siegal, T.; Pfeffer, M.R.; Meltzer, A.; Shezen, E.; Nimrod, A.; Ezov, N.; Ovadia, H. Cellular and secretory mechanisms related to delayed radiation-induced microvessel dysfunction in the spinal cord of rats. Int. J. Radiat. Oncol. Biol. Phys. 1996, 36, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Chen, P.; Jain, V.; Reilly, R.M.; Wong, C.S. Early radiation-induced endothelial cell loss and blood-spinal cord barrier breakdown in the rat spinal cord. Radiat. Res. 2004, 161, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Barjaktarovic, Z.; Schmaltz, D.; Shyla, A.; Azimzadeh, O.; Schulz, S.; Haagen, J.; Dörr, W.; Sarioglu, H.; Schäfer, A.; Atkinson, M.J.; et al. Radiation-induced signaling results in mitochondrial impairment in mouse heart at 4 weeks after exposure to X-rays. PLoS One 2011, 6, e27811. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.K.; Pathak, R.; Zandkarimi, F.; Kaur, P.; Alkhalil, L.; Singh, R.; Zhong, X.; Ghosh, S.; Aykin-Burns, N.; Hauer-Jensen, M. Liver metabolomics reveals increased oxidative stress and fibrogenic potential in gfrp transgenic mice in response to ionizing radiation. J. Proteome Res. 2014, 13, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Rodemann, H.P. Role of radiation-induced signaling proteins in the response of vascular and connective tissues. In Modification of Radiation Response; Nieder, C., Milas, L., Ang, K.K., Eds.; Springer-Verlag: Berlin, Germary, 2003; pp. 15–28. [Google Scholar]
- Storer, J.B. Acute responses to ionizing radiation. Biology of the Laboratory Mouse; Green, E.L., Ed.; The Jackson Laboratory: Bar Harbor, ME, USA, 2007. Chapter 22. 2nd ed. Available online: http://www.informatics.jax.org/greenbook/frames/frametitle.shtml (accessed on 12 February 2015).
- Brook, I.; Elliott, T.B.; Ledney, G.D. Infection after ionizing radiation. In Handbook of Animal Models of Infection; Zak, O., Merle, A., Sande, M.A., Eds.; Academic Press: San Diego, CA, USA; London, UK, 1999; Chapter 17; pp. 151–161. [Google Scholar]
- Potten, C.S.; Grant, H.K. The relationship between ionizing radiation-induced apoptosis and stem cells in the small and large intestine. Br. J. Cancer 1998, 78, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Lombardini, E.; Salber, R.; Kazemzedeh, M.; Nagy, V.; Olsen, C.; Whitnall, M.H. Hematological changes as prognostic indicators of survival: Similarities between Gottingen minipigs, humans, and other large animal models. PLoS One 2011, 6, e25210. [Google Scholar] [CrossRef] [PubMed]
- Roth, N.M.; Sontag, M.R.; Kiani, M.F. Early effects of ionizing radiation on the microvascular networks in normal tissue. Radiat. Res. 1999, 151, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Lorne, E.; Dupont, H.; Abraham, E. Toll-like receptors 2 and 4: Initiators of non-septic inflammation in critical care medicine? Intensive Care Med. 2010, 36, 1826–1835. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Torrice, C.D.; Bell, J.F.; Monahan, K.B.; Jiang, Q.; Wang, Y.; Ramsey, M.R.; Jin, J.; Wong, K.K.; Su, L.; et al. Mitigation of hematologic radiation toxicity in mice through pharmacological quiescence induced by CDK4/6 inhibition. J. Clin. Investig. 2010, 120, 2528–2536. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Chai, Y.; Ponnaiya, B.; Ivanov, V.N. Radiation induced non-targeted response: Mechanism and potential clinical implications. Curr. Mol. Pharmacol. 2011, 4, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Brunswig-Spickenheier, B.; Cappallo-Obermann, H.; Eggert, K.; Gehling, U.M.; Rudolph, C.; Schlegelberger, B.; Cornils, K.; Zustin, J.; Spiess, A.N.; et al. Radiation rescue: Mesenchymal stromal cells protect from lethal irradiation. PLoS One 2011, 6, e14486. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Fukumoto, R.; Gorbunov, N.V. Lipid peroxidation after ionizing irradiation leads to apoptosis and autophagy. In Lipid Peroxidation; Angel, C., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2012; pp. 261–278. [Google Scholar]
- Brook, I.; Elliott, T.B.; Ledney, G.D.; Shoemaker, M.O.; Knudson, G.B. Management of postirradiation infection: Lessons learned from animal models. Mil. Med. 2004, 169, 194–197. [Google Scholar] [PubMed]
- Berbée, M.; Fu, Q.; Boerma, M.; Wang, J.; Kumar, K.S.; Hauer-Jensen, M. γ-tocotrienol ameliorates intestinal radiation injury and reduces vascular oxidative stress after total-body irradiation by an HMG-CoA reductase-dependent mechanism. Radiat. Res. 2009, 171, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Zhai, M.; Liao, P.J.; Bolduc, D.L.; Elliott, T.B.; Gorbunov, N.V. Pegylated G-CSF inhibits blood cell depletion, increases platelets, blocks splenomegaly, and improves survival after whole-body ionizing irradiation but not after irradiation combined with burn. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Wattel, F.; Mathieu, D.; Nevière, R.; Bocquillon, N. Role of microcirculation in multiorgan failure of infectious origin. Bull. Acad. Natl. Med. 2000, 184, 1609–1619. [Google Scholar] [PubMed]
- Kiang, J.G.; Zhai, M.; Liao, P.-J.; Elliott, T.B.; Gorbunov, N.V. Ghrelin therapy improves survival after whole-body ionizing irradiation or combined with burn or wound: Amelioration of leukocytopenia, thrombocytopenia, splenomegaly, and bone marrow injury. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Fuks, Z.; Persaud, R.S.; Alfieri, A.; McLoughlin, M.; Ehleiter, D.; Schwartz, J.L.; Seddon, A.P.; Cordon-Cardo, C.; Haimovitz-Friedman, A. Basic fibroblast growth factor protects endothelial cells against radiation-induced programmed cell death in vitro and in vivo. Cancer Res. 1994, 54, 2582–9025. [Google Scholar] [PubMed]
- Wen, S.; Goldberg, L.R.; Dooner, M.S.; Reagan, J.L.; Quesenberry, P.J. Mesenchymal stem cell-derived vesicles reverse hematopoietic radiation damage. Blood 2013, 122, 2459. [Google Scholar]
- Georgakilas, A.G.; O’Neill, P.; Stewart, R.D. Induction and repair of clustered DNA lesions: What do we know so far? Radiat. Res. 2013, 180, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules-mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Hatzi, V.I.; Laskaratou, D.A.; Mavragani, I.V.; Nikitaki, Z.; Mangelis, A.; Panayiotidis, M.I.; Pantelias, G.E.; Terzoudi, G.I.; Georgakilas, A.G. Non-targeted radiation effects in vivo: A critical glance of the future in radiobiology. Cancer Lett. 2015, 356, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Non-Target and Delayed Effects of Exposure to Ionizing Radiation. UNCEAR 2006 Report Vol II Annex C. Available online: http://www.unscear.org/unscear/en/publications/2006_2.html (accessed on the 26 January 2015).
- Roots, R.; Okada, S. Protection of DNA molecules of cultured mammalian cells from radiation induced single strand scissions by various alcohols and SH compounds. Int. J. Radiat. Biol. 1972, 21, 329–342. [Google Scholar] [CrossRef]
- LaVerne, J.A. OH Radicals and Oxidizing Products in the Gamma Radiolysis of Water. Radiat. Res. 2000, 153, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Forni, L.G.; Willson, R.L. Vitamin E analogue Trolox CEsr and pulse-radiolysis studies of free-radical reactions. Biochem. J. 1988, 255, 513–522. [Google Scholar] [PubMed]
- Stadtman, E.R. Oxidation of free amino acids and amino acid residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Duran, L.; Tappel, A.L. Production of carbonyl compounds and sulfur compounds on irradiation of amino acids. Radiat. Res. 1958, 9, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, F.J.; Cipollina, C.; Freeman, B.A. Formation and signaling actions of electrophilic lipids. Chem. Rev. 2011, 111, 5997–6021. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.J.; Wondrak, G.T.; Laurean, D.C.; Jacobson, M.K.; Jacobson, E.L. DNA damage by carbonyl stress in human skin cells. Mutat. Res. 2003, 522, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K. Redox-derived damage-associated molecular patterns: Ligand function of lipid peroxidation adducts. Redox Biol. 2013, 1, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Ilnytskyy, Y.; Kovalchuk, O. Non-targeted radiation effects-an epigenetic connection. Mutat. Res. 2011, 714, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Stewart, J.; Ware, J.H.; Zhou, Z.; Donahue, J.J.; Kennedy, A.R. Effects of dietary supplements on the space radiation-induced reduction in total antioxidant status in CBA mice. Radiat. Res. 2006, 165, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.F.; Landauer, M.R. Radioprotection by antioxidants. Ann. N. Y. Acad. Sci. 2000, 899, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.; Pasquali, E.; Leonardi, S.; Rebessi, S.; Tanori, M.; Giardullo, P.; Borra, F.; Pazzaglia, S.; Naus, C.C.; di Majo, V.; et al. Role of connexin43 and ATP in long-range bystander radiation damage and oncogenesis in vivo. Oncogene 2011, 30, 4601–4608. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.E.; Kim, H.; Rajagopalan, M.S.; Stone, B.; Salimi, U.; Rwigema, J.C.; Epperly, M.W.; Shen, H.; Goff, J.P.; Franicola, D.; et al. Repopulation of the irradiation damaged lung with bone marrow-derived cells. In Vivo 2012, 26, 9–18. [Google Scholar] [PubMed]
- Wang, Y.; Liu, L.; Pazhanisamy, S.K.; Li, H.; Meng, A.; Zhou, D. Total body irradiation causes residual bone marrow injury by induction of persistent oxidative stress in murine hematopoietic stem cells. Free Radic. Biol. Med. 2010, 48, 348–356. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Y.; Pang, X.; Su, Y.; Ai, G.; Wang, T. ER stress induced by ionising radiation in IEC-6 cells. Int. J. Radiat. Biol. 2010, 86, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Naik, E.; Dixit, V.M. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. J. Exp. Med. 2011, 208, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Leach, J.K.; van Tuyle, G.; Lin, P.S.; Schmidt-Ullrich, R.; Mikkelsen, R.B. Ionizing radiation-induced, mitochondria-dependent generation of reactive oxygen/nitrogen. Cancer Res. 2001, 61, 3894–3901. [Google Scholar] [PubMed]
- Leach, J.K.; Black, S.M.; Schmidt-Ullrich, R.K.; Mikkelsen, R.B. Activation of constitutive nitric-oxide synthase activity is an early signaling event induced by ionizing radiation. J. Biol. Chem. 2002, 277, 15400–15406. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yu, H.; Liang, P.H.; Cheng, H.; XuFeng, R.; Yuan, Y.; Zhang, P.; Smith, C.A.; Cheng, T. An acute negative bystander effect of γ-irradiated recipients on transplanted hematopoietic stem cells. Blood 2012, 119, 3629–3637. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Azzam, E.I.; Li, J.J.; Gius, D. Metabolic oxidation/reduction reactions and cellular responses to ionizing radiation: A unifying concept in stress response biology. Cancer Metastasis Rev. 2004, 23, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Robbins, M.E. Inflammation and chronic oxidative stress in radiation-induced late normal tissue injury: Therapeutic implications. Curr. Med. Chem. 2009, 16, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Kobashigawa, S.; Suzuki, K.; Yamashita, S. Ionizing radiation accelerates Drp1-dependent mitochondrial fission, which involves delayed mitochondrial reactive oxygen species production in normal human fibroblast-like cells. Biochem. Biophys. Res. Commun. 2011, 414, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Kam, W.W.; Banati, R.B. Effects of ionizing radiation on mitochondria. Free Radic. Biol. Med. 2013, 65, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, Y.M.; Park, J.W. Regulation of ionizing radiation-induced apoptosis by a manganese porphyrin complex. Biochem. Biophys. Res. Commun. 2005, 334, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Stoyanovsky, D.A.; Huang, Z.; Jiang, J.; Belikova, N.A.; Tyurin, V.; Epperly, M.W.; Greenberger, J.S.; Bayir, H.; Kagan, V.E. A manganese-porphyrin complex decomposes H2O2, inhibits apoptosis, and acts as a radiation mitigator in vivo. ACS Med. Chem. Lett. 2011, 2, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Zabbarova, I.; Kanai, A. Targeted delivery of radioprotective agents to mitochondria. Mol. Interv. 2008, 8, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Pogue-Geile, K.L.; Epperly, M.W.; Bigbee, W.L.; Draviam, R.; Day, B.W.; Wald, N.; Watkins, S.C.; Greenberger, J.S. Activation of the nitric oxide synthase 2 pathway in the response of bone marrow stromal cells to high doses of ionizing radiation. Radiat. Res. 2000, 154, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.E.; Kim, B.Y.; Kwak, S.Y.; Bae, I.H.; Han, Y.H. Ionizing radiation-inducible microRNA miR-193a-3p induces apoptosis by directly targeting Mcl-1. Apoptosis 2013, 18, 896–909. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Xufeng, R.; Park, M.R.; Gu, H.; Hu, L.; Kang, J.W.; Ma, S.; Liang, P.H.; Li, Y.; Cheng, H.; et al. Hematopoietic stem cell regeneration enhanced by ectopic expression of ROS-detoxifying enzymes in transplant mice. Mol. Ther. 2013, 21, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Burdelya, L.G.; Brackett, C.M.; Kojouharov, B.; Gitlin, I.I.; Leonova, K.I.; Gleiberman, A.S.; Aygun-Sunar, S.; Veith, J.; Johnson, C.; Haderski, G.J.; et al. Central role of liver in anticancer and radioprotective activities of Toll-like receptor 5 agonist. Proc. Natl. Acad. Sci. USA 2013, 110, E1857–E1866. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Elliott, T.B.; McDaniel, D.P.; Zhai, M.; Liao, P.-J.; Kiang, J.G. Up-regulation of autophagy defense mechanisms in mouse mesenchymal stromal cells in response to ionizing irradiation followed by bacterial challenge. In Autophagy—A Double-Edged Sword—Cell Survival or Death? Yannick, B., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2013; pp. 331–350. [Google Scholar]
- Gorbunov, N.V.; Garrison, B.R.; McDaniel, D.P.; Zhai, M.; Liao, P.J.; Nurmemet, D.; Kiang, J.G. Adaptive redox response of mesenchymal stromal cells to stimulation with lipopolysaccharide inflammagen: Mechanisms of remodeling of tissue barriers in sepsis. Oxid. Med. Cell. Longev. 2013, 186795, 1–16. [Google Scholar] [CrossRef]
- Friedenstein, A. Stromal-hematopoietic interrelationships: Maximov’s ideas and modern models. Haematol. Blood Transfus. 1989, 32, 159–167. [Google Scholar] [PubMed]
- Powell, D.W.; Pinchuk, I.V.; Saada, J.I.; Chen, X.; Mifflin, R.C. Mesenchymal cells of the intestinal lamina propria. Ann. Rev. Physiol. 2011, 3, 213–237. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Breen, A.P.; Murphy, J.A. Reactions of oxyl radicals with DNA. Free Radic. Biol. Med. 1995, 18, 1033–1077. [Google Scholar] [CrossRef] [PubMed]
- Østdal, H.; Davies, M.J.; Andersen, H.J. Reaction between protein radicals and other biomolecules. Free Radic. Biol. Med. 2002, 33, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Bild, W.; Ciobica, A.; Padurariu, M.; Bild, V. The interdependence of the reactive species of oxygen, nitrogen, and carbon. J. Physiol. Biochem. 2013, 69, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L. Carbonyl modified proteins in cellular regulation, aging, and disease. Free Radic. Biol. Med. 2002, 32, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Marcocci, L.; Liu, L.; Suzuki, Y.J. Cell signaling by protein carbonylation and decarbonylation. Antioxid. Redox Signal. 2010, 12, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Radical-free biology of oxidative stress. Am. J. Physiol. Cell Physiol. 2008, 295, C849–C868. [Google Scholar] [CrossRef] [PubMed]
- Gebicki, J.M.; Nauser, T.; Domazou, A.; Steinmann, D.; Bounds, P.L.; Koppenol, W.H. Reduction of protein radicals by GSH and ascorbate: Potential biological significance. Amino Acids 2010, 39, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Gasparutto, D.; Ravanat, J.L. Oxidative damage to DNA: Formation, measurement and biochemical features. Mutat. Res. 2003, 531, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Higdon, A.; Diers, A.R.; Oh, J.Y.; Landar, A.; Darley-Usmar, V.M. Cell signalling by reactive lipid species: New concepts and molecular mechanisms. Biochem. J. 2012, 442, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Tyurina, Y.Y.; Tyurin, V.A.; Epperly, M.W.; Greenberger, J.S.; Kagan, V.E. Oxidative lipidomics of gamma-irradiation-induced intestinal injury. Free Radic. Biol. Med. 2008, 44, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Anuranjani; Bala, M. Concerted action of Nrf2-ARE pathway, MRN complex, HMGB1 and inflammatory cytokines-implication in modification of radiation damage. Redox Biol. 2014, 2, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Kiang, J.G. Up-regulation of autophagy in small intestine paneth cells in response to total-body γ-irradiation. J. Pathol. 2009, 219, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Barshishat-Kupper, M.; McCart, E.A.; Mueller, G.P.; Regina, M.; Day, R.M. Bone marrow protein oxidation in response to ionizing radiation in C57BL/6J mice. Proteome 2014, 2, 291–302. [Google Scholar] [CrossRef]
- Grimsrud, P.A.; Xie, H.; Griffin, T.J.; Bernlohr, D.A. Oxidative stress and covalent modification of protein with bioactive aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef] [PubMed]
- Haberzettl, P.; Hill, B.G. Oxidized lipids activate autophagy in a JNK-dependent manner by stimulating the endoplasmic reticulum stress response. Redox Biol. 2013, 1, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Picklo, M.J.; Azenkeng, A.; Hoffmann, M.R. Trans-4-oxo-2-nonenal potently alters mitochondrial function. Free Radic. Biol. Med. 2011, 50, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Tulard, A.; Hoffschir, F.; de Boisferon, F.H.; Luccioni, C.; Bravard, A. Persistent oxidative stress after ionizing radiation is involved in inherited radiosensitivity. Free Radic. Biol. Med. 2003, 35, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, O.; Miyahara, M.; Shiraishi, N.; Nakazawa, T. Radiation-induced damage to mitochondrial d-β-hydroxybutyrate dehydrogenase and lipidperoxidation. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1985, 48, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Brady, N.R.; Elmore, S.P.; van Beek, J.J.; Krab, K.; Courtoy, P.J.; Hue, L.; Westerhoff, H.V. Coordinated behavior of mitochondria in both space and time: A reactive oxygen species-activated wave of mitochondrial depolarization. Biophys. J. 2004, 87, 2022–2034. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ROS-induced ROS release: An update and review. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 509–517. [Google Scholar] [CrossRef]
- Warren, S.L.; Whipple, G.H. Roentgen ray intoxication: IV. Intestinal lesions and acute intoxication produced by radiation in a variety of animals. J. Exp. Med. 1923, 38, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.B.; Ledney, G.D. Therapy for bacterial infections following ionizing radiation injury. In Medical Consequences of Radiological and Nuclear Weapons; Mickelson, A.B., Ed.; Office of The Surgeon General United States Army: Falls Church, VA, USA; Borden Institute: Fort Detrick, MD, USA, 2012; pp. 81–100. [Google Scholar]
- Sakaguchi, S.; Furusawa, S. Oxidative stress and septic shock: Metabolica spects of oxygen-derived free radicals generated in the liver during endotoxemia. FEMS Immunol. Med. Microbiol. 2006, 47, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Towner, R.A.; Garteiser, P.; Bozza, F.; Smith, N.; Saunders, D.; d’ Avila, J.C.; Magno, F.; Oliveira, M.F.; Ehrenshaft, M.; Lupu, F.; et al. In vivo detection of free radicals in mouse septic encephalopathy using molecular MRI and immuno-spin trapping. Free Radic. Biol. Med. 2013, 65, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, B.J.; Wei, L.; Zhang, L.; Ping, X.; Epperly, M.; Greenberger, J.; Cheng, T.; Yu, J. Ionizing irradiation induces acute haematopoietic syndrome and gastrointestinal syndrome independently in mice. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Garrison, B.; Kiang, J.G. Response of crypt paneth cells in the small intestine following total-body γ-irradiation. Int. J. Immunopathol. Pharmacol. 2010, 23, 971–983. [Google Scholar] [PubMed]
- Tapio, S. Ionizing radiation effects on cells, organelles and tissues on proteome level. Adv. Exp. Med. Biol. 2013, 990, 37–48. [Google Scholar] [PubMed]
- Criswell, T.; Leskov, K.; Miyamoto, S.; Luo, G.; Boothman, D.A. Transcription factors activated in mammalian cells after clinically relevant doses of ionizing radiation. Oncogene 2003, 22, 5813–5827. [Google Scholar] [CrossRef] [PubMed]
- Kempf, S.J.; Azimzadeh, O.; Atkinson, M.J.; Tapio, S. Long-term effects of ionising radiation on the brain: Cause for concern? Radiat. Environ. Biophys. 2013, 52, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Greenberger, J.S.; Epperly, M. Bone marrow-derived stem cells and radiation response. Semin. Radiat. Oncol. 2009, 9, 133–139. [Google Scholar] [CrossRef]
- Ahmad, T.; Mukherjee, S.; Pattnaik, B.; Kumar, M.; Singh, S.; Kumar, M.; Rehman, R.; Tiwari, B.K.; Jha, K.A.; Barhanpurkar, A.P.; et al. Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy. EMBO J. 2014, 33, 994–1010. [Google Scholar] [PubMed]
- Azzam, E.I.; de Toledo, S.M.; Little, J.B. Stress signaling from irradiated to non-irradiated cells. Curr. Cancer Drug Targets 2004, 4, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.R.; Dias, I.H.; Willetts, R.S.; Devitt, A. Redox regulation of protein damage in plasma. Redox Biol. 2014, 2, 430–435. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorbunov, N.V.; Sharma, P. Protracted Oxidative Alterations in the Mechanism of Hematopoietic Acute Radiation Syndrome. Antioxidants 2015, 4, 134-152. https://doi.org/10.3390/antiox4010134
Gorbunov NV, Sharma P. Protracted Oxidative Alterations in the Mechanism of Hematopoietic Acute Radiation Syndrome. Antioxidants. 2015; 4(1):134-152. https://doi.org/10.3390/antiox4010134
Chicago/Turabian StyleGorbunov, Nikolai V., and Pushpa Sharma. 2015. "Protracted Oxidative Alterations in the Mechanism of Hematopoietic Acute Radiation Syndrome" Antioxidants 4, no. 1: 134-152. https://doi.org/10.3390/antiox4010134