
Syrah Grape Polyphenol Extracts Protect Human Skeletal Muscle Cells from Oxidative and Metabolic Stress Induced by Excess of Palmitic Acid: Effect of Skin/Seed Ripening Stage

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Grape Materials
2.2. Determination of Total Phenolic Content and Total Antioxidant Capacity
2.3. Qualitative Profiling of GPP Extracts
2.3.1. Anthocyanins
2.3.2. Stilbenes
2.3.3. Monomeric Flavan-3-ols and Proanthocyanidins
2.4. Human Primary SM Cell Culture and Treatment
2.5. Western Blot Analysis
2.6. GPP Extracts Toxicity/Cell Viability
2.7. Citrate Synthase Activity
2.8. Statistical Analysis
3. Results
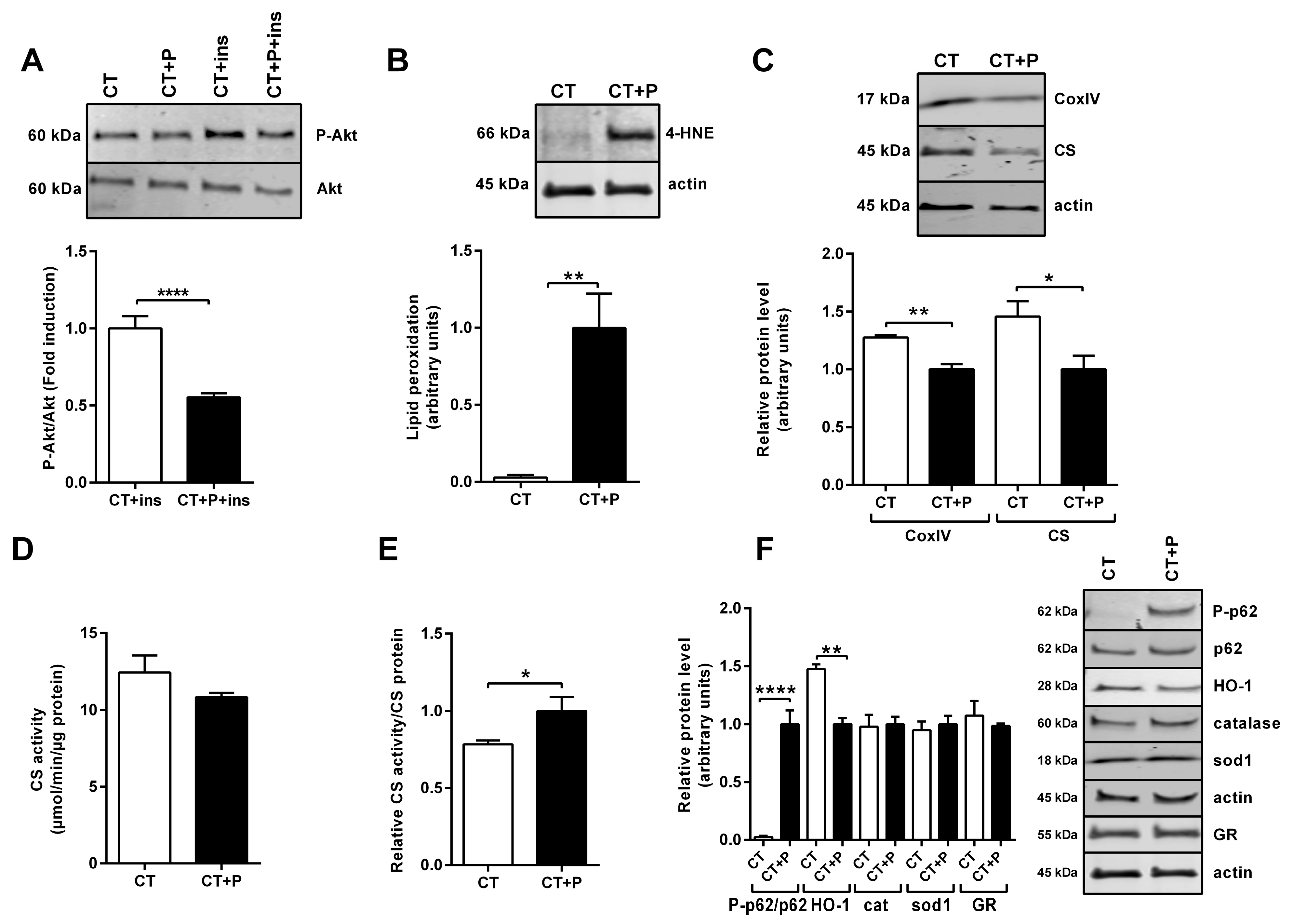
3.1. Characterization of Human SM Cells’ Metabolic Alterations Induced by Lipid Overload
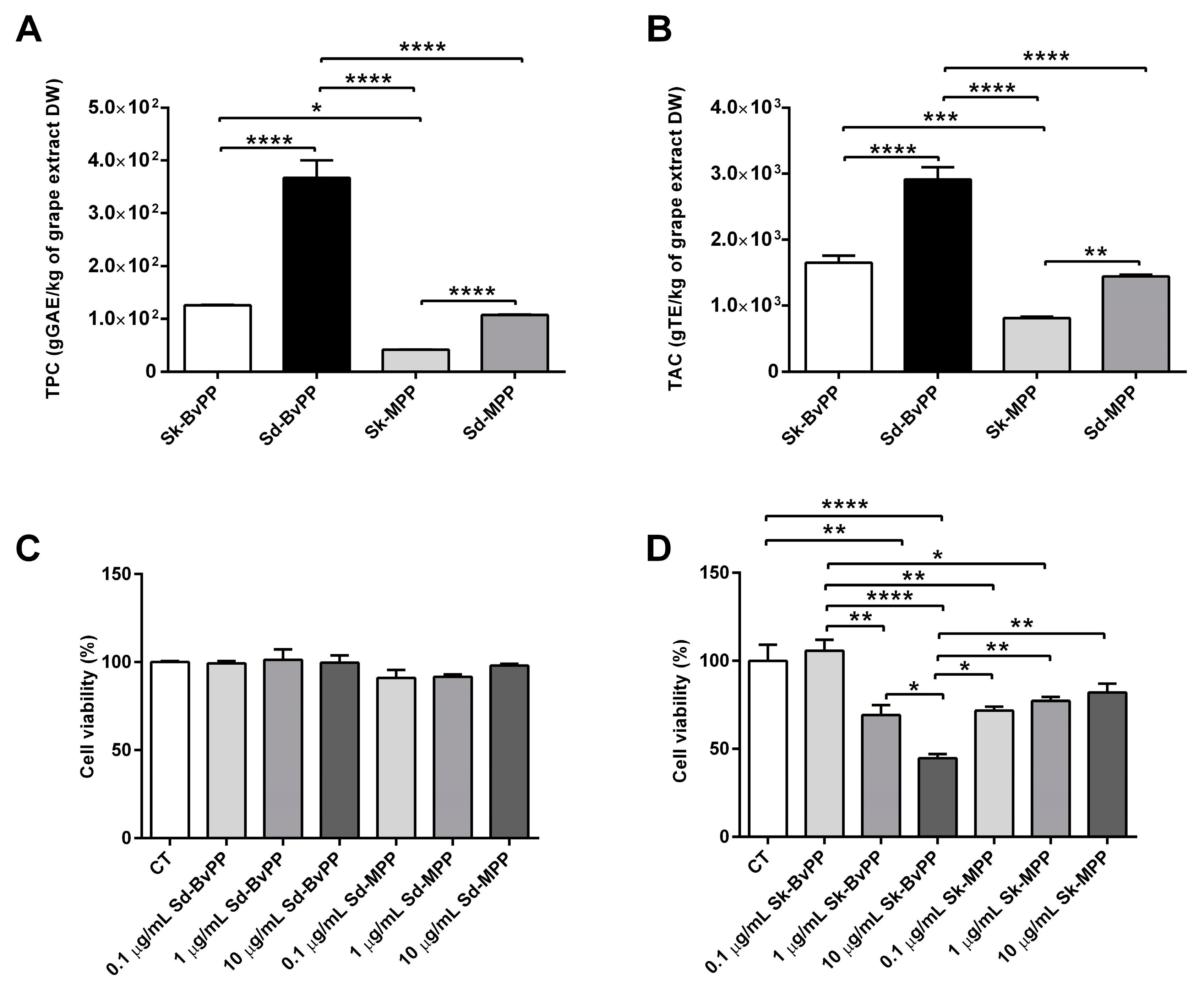
3.2. Grape Extracts Characterization and Cell Viability
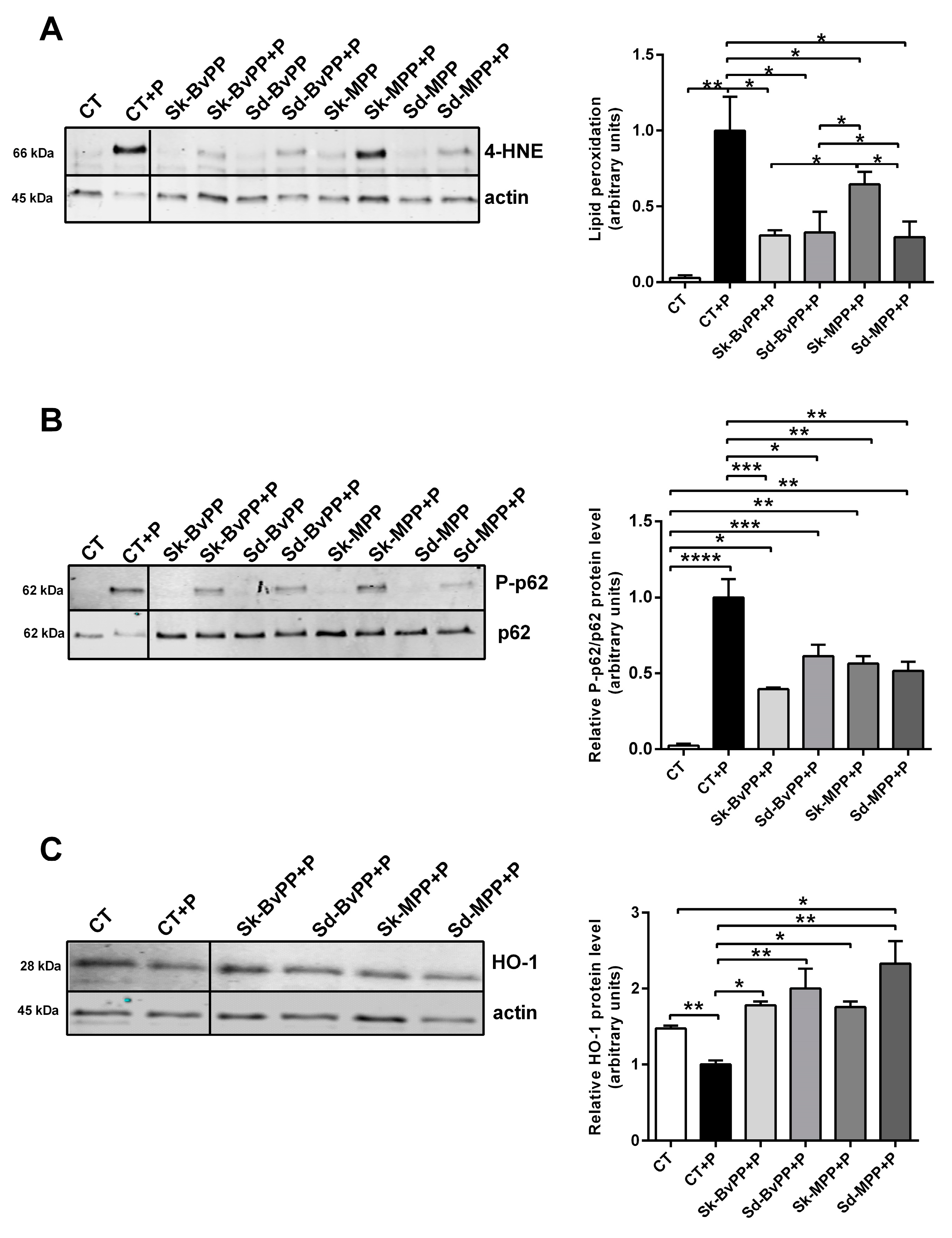
3.3. Effect of GPP Extracts on Antioxidant Response in Lipid-Overload Human SM Cells
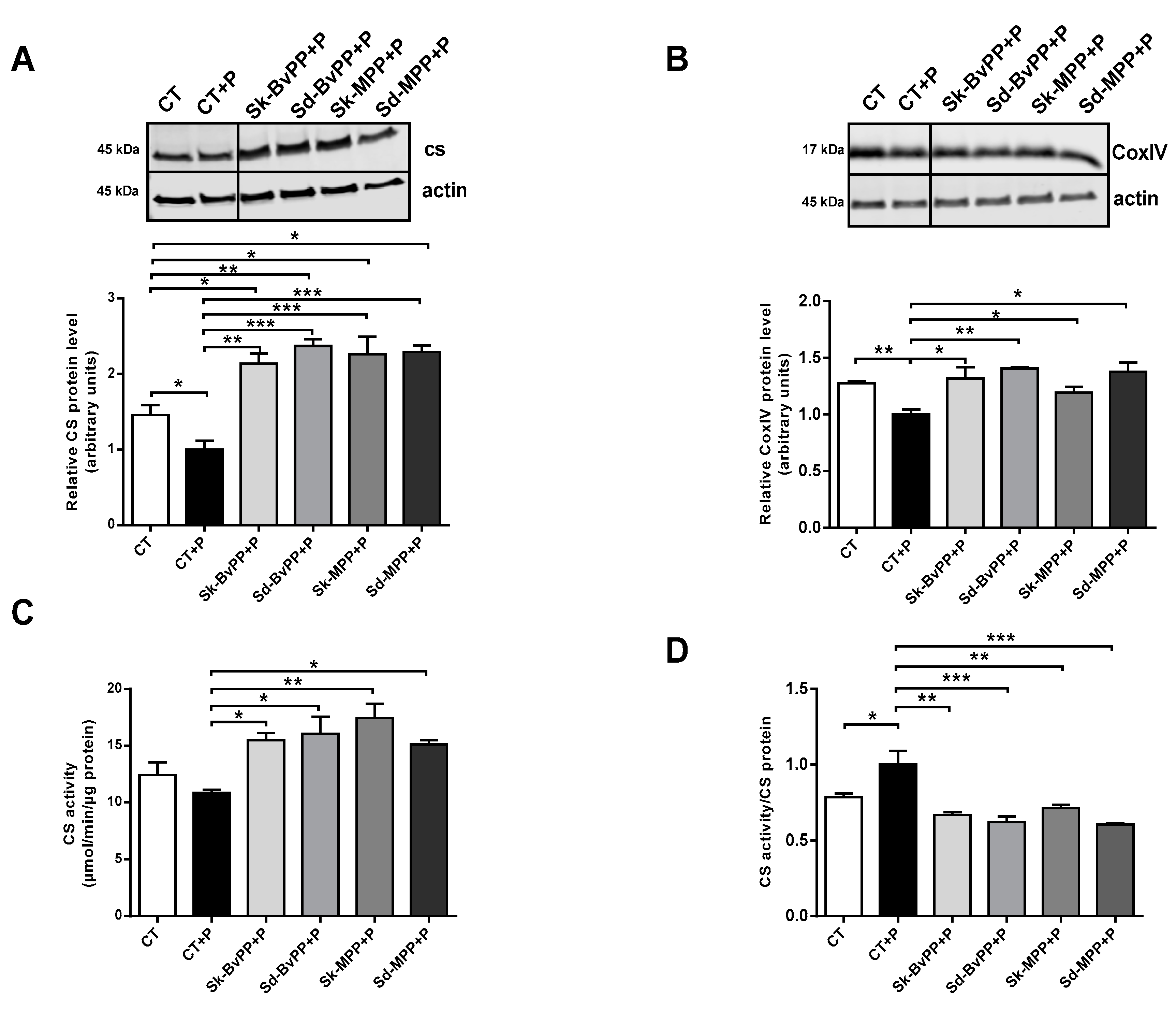
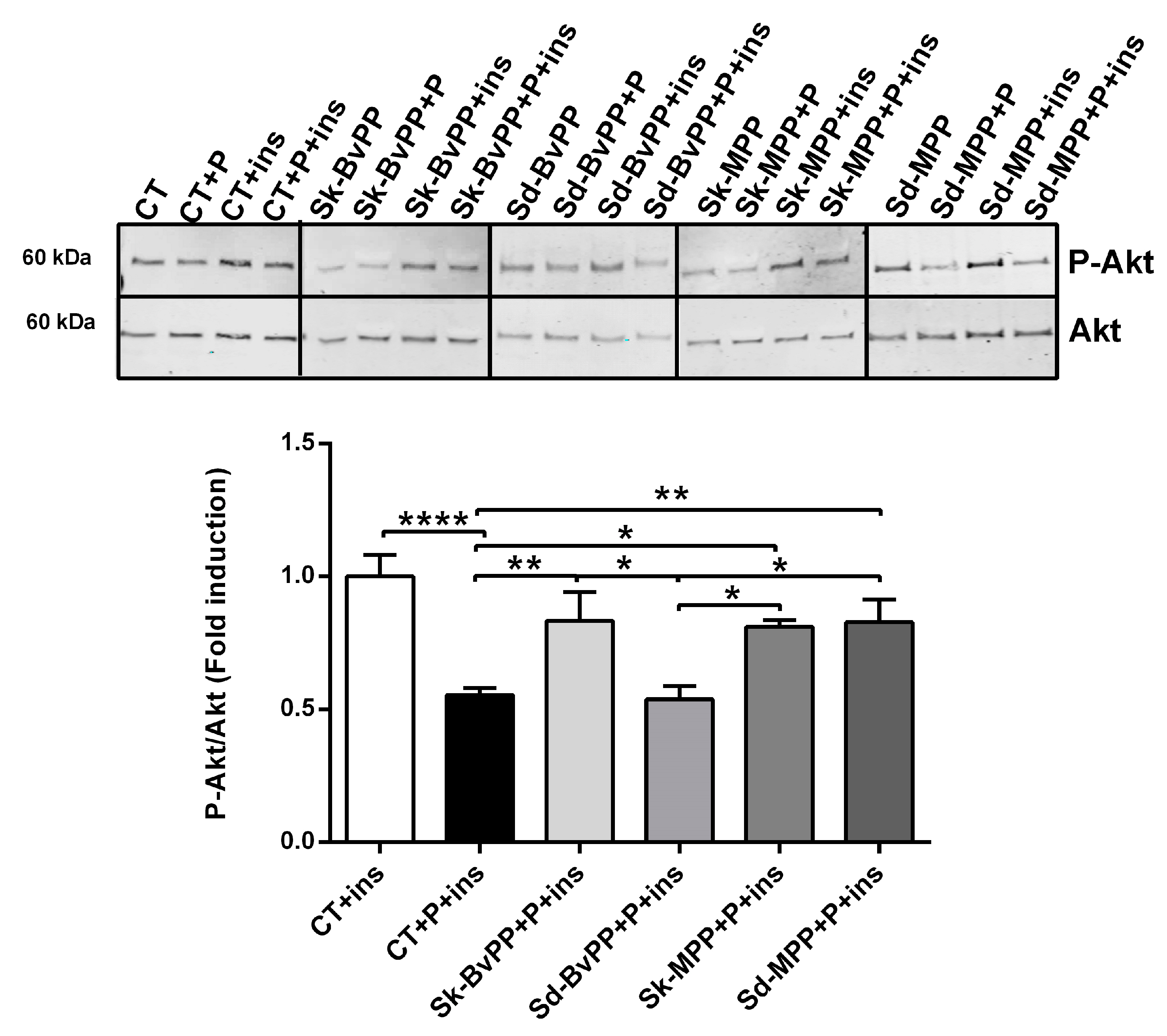
3.4. Effect of GPP Extracts on Human SM Cells Mitochondrial Activity and Insulin Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, H.; Ballantyne, C.M. Skeletal Muscle Inflammation and Insulin Resistance in Obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef]
- Amouzou, C.; Breuker, C.; Fabre, O.; Bourret, A.; Lambert, K.; Birot, O.; Fédou, C.; Dupuy, A.-M.; Cristol, J.-P.; Sutra, T.; et al. Skeletal Muscle Insulin Resistance and Absence of Inflammation Characterize Insulin-Resistant Grade I Obese Women. PLoS ONE 2016, 11, e0154119. [Google Scholar] [CrossRef]
- Gilbert, M. Role of Skeletal Muscle Lipids in the Pathogenesis of Insulin Resistance of Obesity and Type 2 Diabetes. J. Diabetes Investig. 2021, 12, 1934–1941. [Google Scholar] [CrossRef]
- Di Meo, S.; Iossa, S.; Venditti, P. Skeletal Muscle Insulin Resistance: Role of Mitochondria and Other ROS Sources. J. Endocrinol. 2017, 233, R15–R42. [Google Scholar] [CrossRef]
- Hood, D.A.; Memme, J.M.; Oliveira, A.N.; Triolo, M. Maintenance of Skeletal Muscle Mitochondria in Health, Exercise, and Aging. Annu. Rev. Physiol. 2019, 81, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial Dysfunction in Type 2 Diabetes Mellitus: An Organ-Based Analysis. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E268–E285. [Google Scholar] [CrossRef] [PubMed]
- Mthembu, S.X.H.; Dludla, P.V.; Nyambuya, T.M.; Kappo, A.P.; Madoroba, E.; Ziqubu, K.; Nyawo, T.A.; Nkambule, B.B.; Silvestri, S.; Muller, C.J.F.; et al. Experimental Models of Lipid Overload and Their Relevance in Understanding Skeletal Muscle Insulin Resistance and Pathological Changes in Mitochondrial Oxidative Capacity. Biochimie 2022, 196, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Yuzefovych, L.; Wilson, G.; Rachek, L. Different Effects of Oleate vs. Palmitate on Mitochondrial Function, Apoptosis, and Insulin Signaling in L6 Skeletal Muscle Cells: Role of Oxidative Stress. Am. J. Physiol.-Endocrinol. Metab. 2010, 299, E1096–E1105. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Xiao, M.; Zhong, H.; Xia, L.; Tao, Y.; Yin, H. Pathophysiology of Mitochondrial Lipid Oxidation: Role of 4-Hydroxynonenal (4-HNE) and Other Bioactive Lipids in Mitochondria. Free Radic. Biol. Med. 2017, 111, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Pillon, N.J.; Croze, M.L.; Vella, R.E.; Soulère, L.; Lagarde, M.; Soulage, C.O. The Lipid Peroxidation By-Product 4-Hydroxy-2-Nonenal (4-HNE) Induces Insulin Resistance in Skeletal Muscle through Both Carbonyl and Oxidative Stress. Endocrinology 2012, 153, 2099–2111. [Google Scholar] [CrossRef]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.-S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The Selective Autophagy Substrate P62 Activates the Stress Responsive Transcription Factor Nrf2 through Inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Sun, C.-C.; Tang, C.-F. Heme Oxygenase-1: A Potential Therapeutic Target for Improving Skeletal Muscle Atrophy. Exp. Gerontol. 2023, 184, 112335. [Google Scholar] [CrossRef]
- Gasier, H.G.; Suliman, H.B.; Piantadosi, C.A. The HO-1/CO System and Mitochondrial Quality Control in Skeletal Muscle. Exerc. Sport Sci. Rev. 2022, 50, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.F.; Garcia, D.; Scaffa, A.; Peterson, A.L.; Ghio, A.J.; Dennery, P.A. Heme Oxygenase-1 Supports Mitochondrial Energy Production and Electron Transport Chain Activity in Cultured Lung Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 6941. [Google Scholar] [CrossRef]
- Murakami, S.; Kusano, Y.; Okazaki, K.; Akaike, T.; Motohashi, H. NRF2 Signalling in Cytoprotection and Metabolism. Br. J. Pharmacol. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nwozo, O.S.; Effiong, E.M.; Aja, P.M.; Awuchi, C.G. Antioxidant, Phytochemical, and Therapeutic Properties of Medicinal Plants: A Review. Int. J. Food Prop. 2023, 26, 359–388. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Silva, P. The Wine Industry By-Products: Applications for Food Industry and Health Benefits. Antioxidants 2022, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Mazza, K.E.L.; Santiago, M.C.P.A.; Do Nascimento, L.S.M.; Godoy, R.L.O.; Souza, E.F.; Brígida, A.I.S.; Borguini, R.G.; Tonon, R.V. Syrah Grape Skin Valorisation Using Ultrasound-assisted Extraction: Phenolic Compounds Recovery, Antioxidant Capacity and Phenolic Profile. Int. J. Food Sci. Technol. 2019, 54, 641–650. [Google Scholar] [CrossRef]
- Hokayem, M.; Blond, E.; Vidal, H.; Lambert, K.; Meugnier, E.; Feillet-Coudray, C.; Coudray, C.; Pesenti, S.; Luyton, C.; Lambert-Porcheron, S.; et al. Grape Polyphenols Prevent Fructose-Induced Oxidative Stress and Insulin Resistance in First-Degree Relatives of Type 2 Diabetic Patients. Diabetes Care 2013, 36, 1454–1461. [Google Scholar] [CrossRef]
- Lambert, K.; Hokayem, M.; Thomas, C.; Fabre, O.; Cassan, C.; Bourret, A.; Bernex, F.; Feuillet-Coudray, C.; Notarnicola, C.; Mercier, J.; et al. Combination of Nutritional Polyphenols Supplementation with Exercise Training Counteracts Insulin Resistance and Improves Endurance in High-Fat Diet-Induced Obese Rats. Sci. Rep. 2018, 8, 2885. [Google Scholar] [CrossRef]
- Pan, X.; Liu, C.; Wang, X.; Zhao, M.; Zhang, Z.; Zhang, X.; Wang, C.; Song, G. Resveratrol Improves Palmitic Acid-induced Insulin Resistance via the DDIT4/mTOR Pathway in C2C12 Cells. Mol. Med. Rep. 2023, 28, 181. [Google Scholar] [CrossRef]
- Godjevac, D.; Tesevic, V.; Velickovic, M.; Vujisic, L.; Vajs, V.; Milosavljevic, S. Polyphenolic Compounds in Seeds from Some Grape Cultivars Grown in Serbia. J. Serbian Chem. Soc. 2010, 75, 1641–1652. [Google Scholar] [CrossRef]
- De Andrade, R.B.; Machado, B.A.S.; Barreto, G.D.A.; Nascimento, R.Q.; Corrêa, L.C.; Leal, I.L.; Tavares, P.P.L.G.; Ferreira, E.D.S.; Umsza-Guez, M.A. Syrah Grape Skin Residues Has Potential as Source of Antioxidant and Anti-Microbial Bioactive Compounds. Biology 2021, 10, 1262. [Google Scholar] [CrossRef]
- Favre, G.; Gómez-Alonso, S.; Pérez-Navarro, J.; García-Romero, E.; Mena-Morales, A.; Piccardo, D.; González-Neves, G. Seed and Skin-Derived Flavanols in Red Wine: A Study of Syrah, Marselan, and Tannat Cultivars. Eur. Food Res. Technol. 2024, 250, 845–857. [Google Scholar] [CrossRef]
- Qi, M.; Luo, Z.; Wu, B.; Wang, L.; Yang, M.; Zhang, X.; Lin, X.; Xu, Y.; Li, X.; Li, L. Spatial Distribution and Time-Course of Polyphenol Accumulation in Grape Berry (Vitis Labruscana Cv. ‘Kyoho’). J. Food Compos. Anal. 2022, 106, 104353. [Google Scholar] [CrossRef]
- Capozzi, A.; Saucier, C.; Bisbal, C.; Lambert, K. Grape Polyphenols in the Treatment of Human Skeletal Muscle Damage Due to Inflammation and Oxidative Stress during Obesity and Aging: Early Outcomes and Promises. Molecules 2022, 27, 6594. [Google Scholar] [CrossRef] [PubMed]
- Benbouguerra, N.; Valls-Fonayet, J.; Krisa, S.; Garcia, F.; Saucier, C.; Richard, T.; Hornedo-Ortega, R. Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells. Foods 2021, 10, 541. [Google Scholar] [CrossRef] [PubMed]
- Dudoit, A.; Benbouguerra, N.; Richard, T.; Hornedo-Ortega, R.; Valls-Fonayet, J.; Coussot, G.; Saucier, C. α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules 2020, 10, 1088. [Google Scholar] [CrossRef] [PubMed]
- Tey, S.R.; Mueller, M.; Reilly, M.; Switalski, C.; Robertson, S.; Sakanaka-Yokoyama, M.; Suzuki, M. Cell Surface Proteins for Enrichment and In Vitro Characterization of Human Pluripotent Stem Cell-Derived Myogenic Progenitors. Stem Cells Int. 2022, 2022, 2735414. [Google Scholar] [CrossRef] [PubMed]
- Fabre, O.; Breuker, C.; Amouzou, C.; Salehzada, T.; Kitzmann, M.; Mercier, J.; Bisbal, C. Defects in TLR3 Expression and RNase L Activation Lead to Decreased MnSOD Expression and Insulin Resistance in Muscle Cells of Obese People. Cell Death Dis. 2014, 5, e1136. [Google Scholar] [CrossRef] [PubMed]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
- Lambertucci, R.H.; Hirabara, S.M.; Silveira, L.D.R.; Levada-Pires, A.C.; Curi, R.; Pithon-Curi, T.C. Palmitate Increases Superoxide Production through Mitochondrial Electron Transport Chain and NADPH Oxidase Activity in Skeletal Muscle Cells. J. Cell. Physiol. 2008, 216, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.B.; Sousa, C.; Costa, A.; Andrade, P.B.; Valentão, P. Glutathione and the Antioxidant Potential of Binary Mixtures with Flavonoids: Synergisms and Antagonisms. Molecules 2013, 18, 8858–8872. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and Its Impact on Metabolic Syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.P.; Gastaldi, G.; Bobbioni-Harsch, E.; Arboit, P.; Gobelet, C.; Dériaz, O.; Golay, A.; Witztum, J.L.; Giacobino, J.-P. Lipid Peroxidation in Skeletal Muscle of Obese as Compared to Endurance-Trained Humans: A Case of Good vs. Bad Lipids? FEBS Lett. 2003, 551, 104–106. [Google Scholar] [CrossRef]
- Morris, R.T.; Laye, M.J.; Lees, S.J.; Rector, R.S.; Thyfault, J.P.; Booth, F.W. Exercise-Induced Attenuation of Obesity, Hyperinsulinemia, and Skeletal Muscle Lipid Peroxidation in the OLETF Rat. J. Appl. Physiol. 2008, 104, 708–715. [Google Scholar] [CrossRef]
- Pillon, N.J.; Vella, R.E.; Soulère, L.; Becchi, M.; Lagarde, M.; Soulage, C.O. Structural and Functional Changes in Human Insulin Induced by the Lipid Peroxidation Byproducts 4-Hydroxy-2-Nonenal and 4-Hydroxy-2-Hexenal. Chem. Res. Toxicol. 2011, 24, 752–762. [Google Scholar] [CrossRef]
- Shearn, C.T.; Fritz, K.S.; Reigan, P.; Petersen, D.R. Modification of Akt2 by 4-Hydroxynonenal Inhibits Insulin-Dependent Akt Signaling in HepG2 Cells. Biochemistry 2011, 50, 3984–3996. [Google Scholar] [CrossRef] [PubMed]
- Drummond, G.S.; Baum, J.; Greenberg, M.; Lewis, D.; Abraham, N.G. HO-1 Overexpression and Underexpression: Clinical Implications. Arch. Biochem. Biophys. 2019, 673, 108073. [Google Scholar] [CrossRef] [PubMed]
- Caimari, A.; Mariné-Casadó, R.; Boqué, N.; Crescenti, A.; Arola, L.; del Bas, J.M. Maternal Intake of Grape Seed Procyanidins during Lactation Induces Insulin Resistance and an Adiponectin Resistance-like Phenotype in Rat Offspring. Sci. Rep. 2017, 7, 12573. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Jimenez-García, M.; Tejada, S.; Moranta, D.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Sureda, A.; Novellino, E.; Capó, X. Grape Polyphenols Ameliorate Muscle Decline Reducing Oxidative Stress and Oxidative Damage in Aged Rats. Nutrients 2020, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Nikawa, T.; Ulla, A.; Sakakibara, I. Polyphenols and Their Effects on Muscle Atrophy and Muscle Health. Molecules 2021, 26, 4887. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic Acids: Natural Versatile Molecules with Promising Therapeutic Applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, D.; Moon, H.Y.; van Praag, H. Exercise in a Pill: The Latest on Exercise-Mimetics. Brain Plast. 2017, 2, 153–169. [Google Scholar] [CrossRef]
- Gubert, C.; Hannan, A.J. Exercise Mimetics: Harnessing the Therapeutic Effects of Physical Activity. Nat. Rev. Drug Discov. 2021, 20, 862–879. [Google Scholar] [CrossRef]
- Jang, Y.J.; Byun, S. Molecular Targets of Exercise Mimetics and Their Natural Activators. BMB Rep. 2021, 54, 581–591. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | ||||
|---|---|---|---|---|
| Sd | Sk | |||
| Ripening Stage | Ripening Stage | |||
| Polyphenol Subclass | Bv | M | Bv | M |
| Flavan-3-ols | ||||
| (+)-Catechin | 25.2 ± 0.31 | 29.4 ± 0.21 * | 9.43 ± 0.16 † | 1.57 ± 0.12 *,† |
| (−)-Epicatechin | 0.70 ± 0.03 | 28.8 ± 0.11 * | nd | 0.33 ± 0.01 *,† |
| Epicatechin gallate | 37.2 ± 0.08 | 22.3 ± 0.03 * | 0.25 ± 0.01 † | 1.40 ± 0.36 *,† |
| Epigallocatechin | 13.9 ± 0.19 | 0.88 ± 0.77 * | nd | nd |
| mDP | 11.28 ± 0.17 | 5.38 ± 0.12 * | 18.9 ± 0.22 | 11.6 ± 0.83 *,† |
| Sk | ||
|---|---|---|
| Ripening Stage | ||
| Polyphenol Subclass | Bv | M |
| Anthocyanins | ||
| Delphinidin 3-O-Glucoside | nd | 0.69 ± 0.01 |
| Cyanidin 3-O-Glucoside | nd | 0.29 ± 0.01 |
| Petunidin 3-O-Glucoside | nd | 0.95 ± 0.03 |
| Peonidin 3-O-Glucoside | nd | 1.86 ± 0.01 |
| Malvidin 3-O-Glucoside | nd | 4.59 ± 0.04 |
| Delphinidin 3-O-Acetyl Glucoside | nd | 0.13 ± 0.01 |
| Cyanidin 3-O-Acetyl Glucoside | nd | 0.05 ± 0.01 |
| Petunidin 3-O-Acetyl Glucoside | nd | 0.22 ± 0.01 |
| Peonidin 3-O-Acetyl Glucoside | nd | 0.76 ± 0.01 |
| Malvidin 3-O-Acetyl Glucoside | nd | 2.27 ± 0.05 |
| Delphinidin 3-O-Coumaroyl Glucoside | nd | 0.02 ± 0.01 |
| Cyanidin 3-O-Coumaroyl Glucoside | nd | 0.23 ± 0.01 |
| Petunidin 3-O-Coumaroyl Glucoside | nd | 0.33 ± 0.01 |
| Peonidin 3-O-Coumaroyl Glucoside | nd | 1.04 ± 0.02 |
| Malvidin 3-O-Coumaroyl Glucoside | nd | 2.02 ± 0.02 |
| Stilbenes | ||
| Trans-resveratrol | 0.03 ± 0.01 | 0.19 ± 0.01 * |
| Cis-resveratrol | 0.01 ± 0.01 | 0.03 ± 0.01 * |
| Trans-piceid | 0.15 ± 0.01 | 1.01 ± 0.01 * |
| Cis-piceid | 0.02 ± 0.01 | 0.21 ± 0.01 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capozzi, A.; Dudoit, A.; Garcia, L.; Carnac, G.; Hugon, G.; Saucier, C.; Bisbal, C.; Lambert, K. Syrah Grape Polyphenol Extracts Protect Human Skeletal Muscle Cells from Oxidative and Metabolic Stress Induced by Excess of Palmitic Acid: Effect of Skin/Seed Ripening Stage. Antioxidants 2024, 13, 373. https://doi.org/10.3390/antiox13030373
Capozzi A, Dudoit A, Garcia L, Carnac G, Hugon G, Saucier C, Bisbal C, Lambert K. Syrah Grape Polyphenol Extracts Protect Human Skeletal Muscle Cells from Oxidative and Metabolic Stress Induced by Excess of Palmitic Acid: Effect of Skin/Seed Ripening Stage. Antioxidants. 2024; 13(3):373. https://doi.org/10.3390/antiox13030373
Chicago/Turabian StyleCapozzi, Adriana, Auriane Dudoit, Luca Garcia, Gilles Carnac, Gérald Hugon, Cédric Saucier, Catherine Bisbal, and Karen Lambert. 2024. "Syrah Grape Polyphenol Extracts Protect Human Skeletal Muscle Cells from Oxidative and Metabolic Stress Induced by Excess of Palmitic Acid: Effect of Skin/Seed Ripening Stage" Antioxidants 13, no. 3: 373. https://doi.org/10.3390/antiox13030373