
Attention-Deficit/Hyperactivity Disorder Animal Model Presents Retinal Alterations and Methylphenidate Has a Differential Effect in ADHD versus Control Conditions
 , , , , and
, , , , and 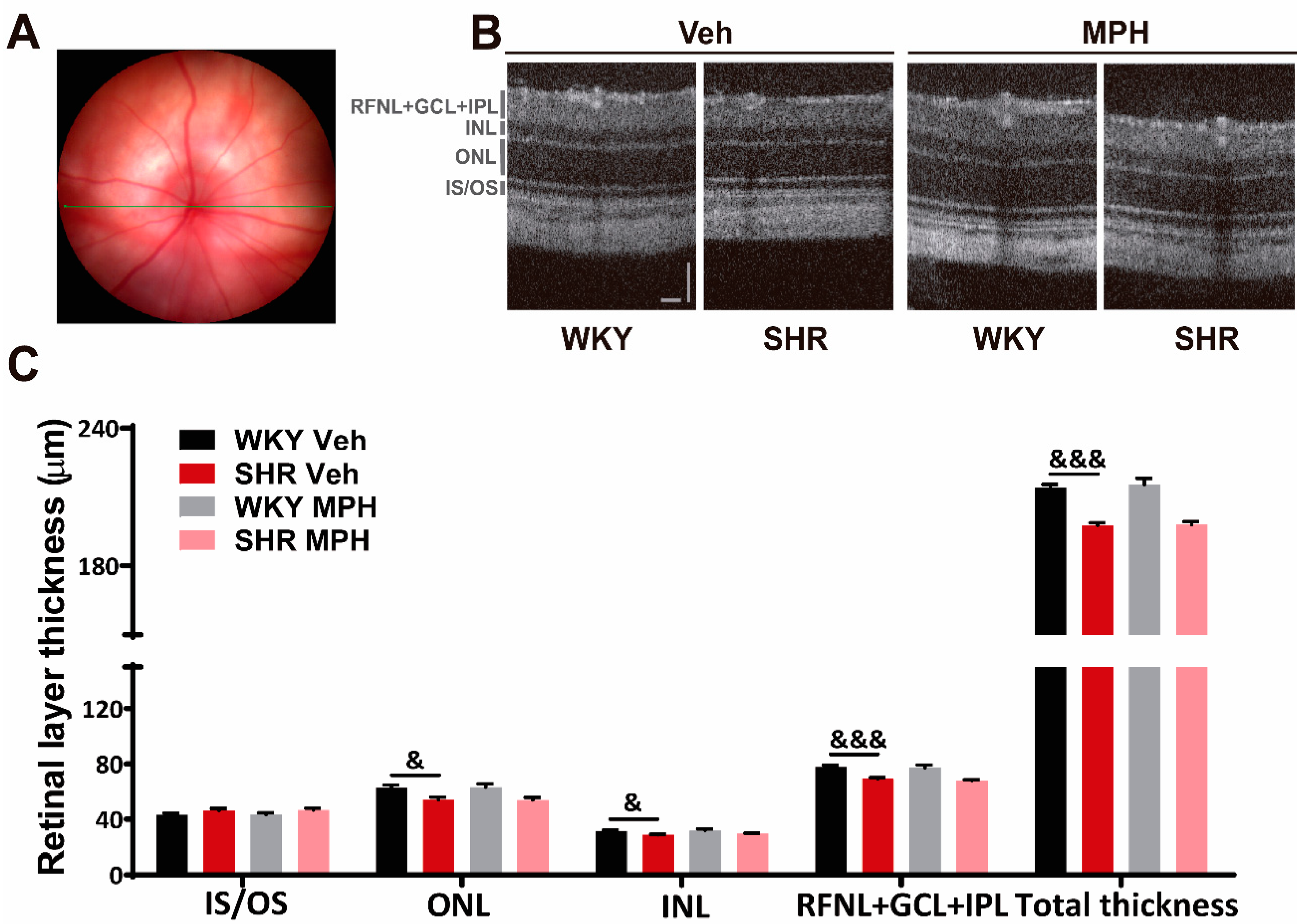{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Electroretinogram (ERG) Recordings
2.3. Optical Coherence Tomography (OCT)
2.4. Immunolabelling
2.4.1. Retinal Cryosections
2.4.2. Retina Wholemounts
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
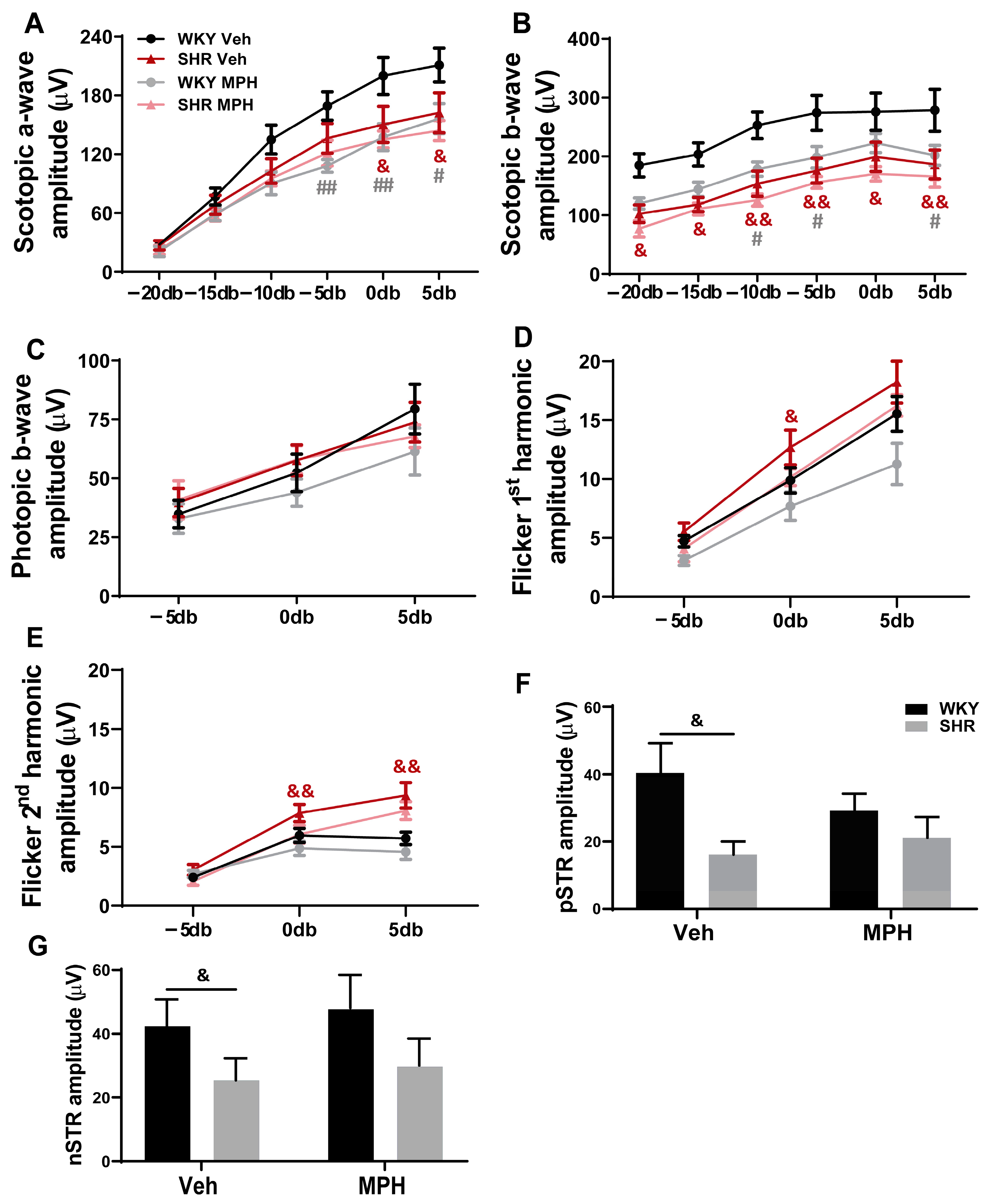
3.1. Impact of MPH Treatment on Retinal Structure and Function
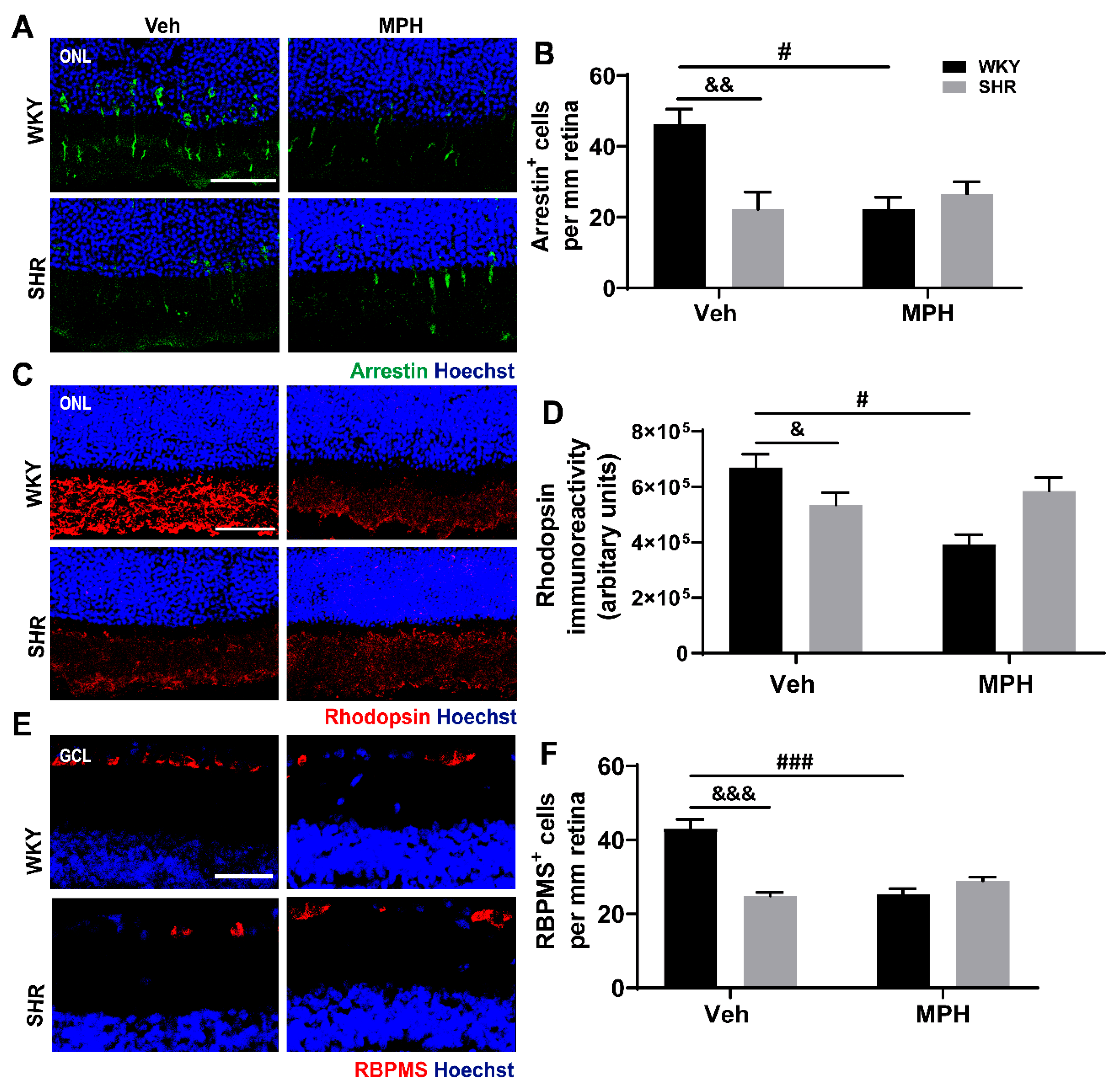
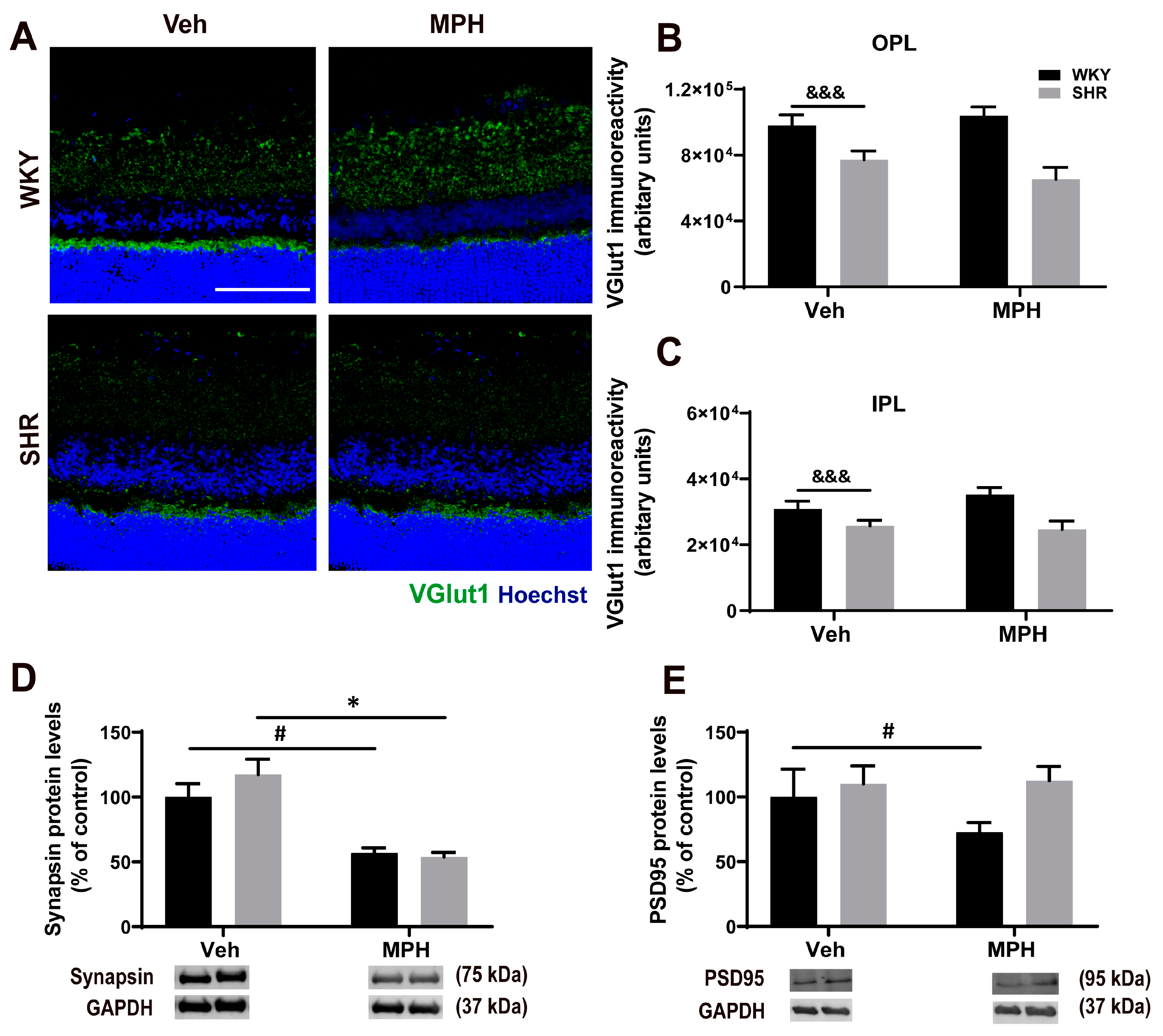
3.2. Neuronal Alterations in the Retina of ADHD Animal Model and the Impact MPH on Control versus ADHD
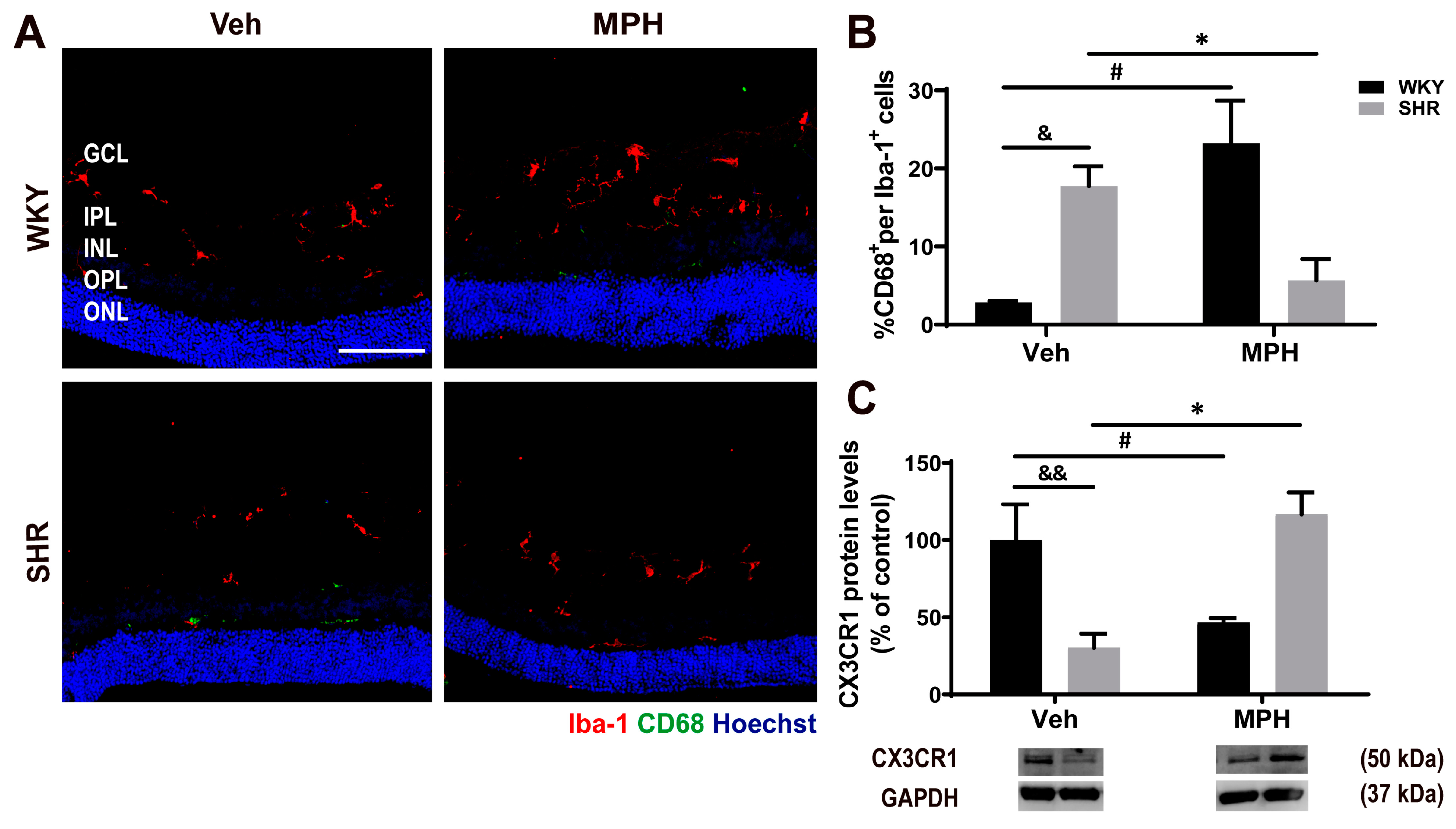
3.3. Microglia Response and Neuroimmune Profile in ADHD and the Effect of MPH Treatment
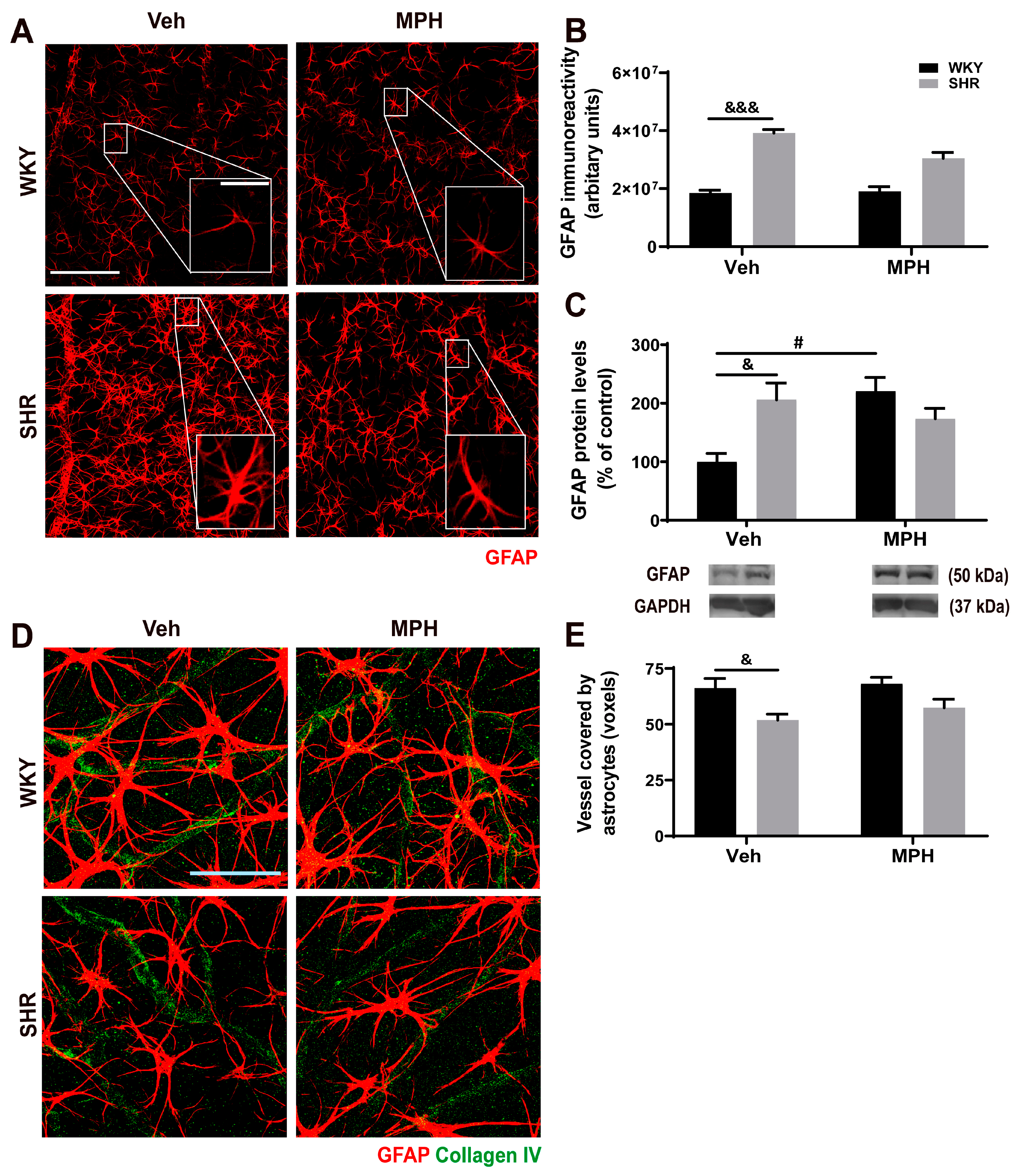
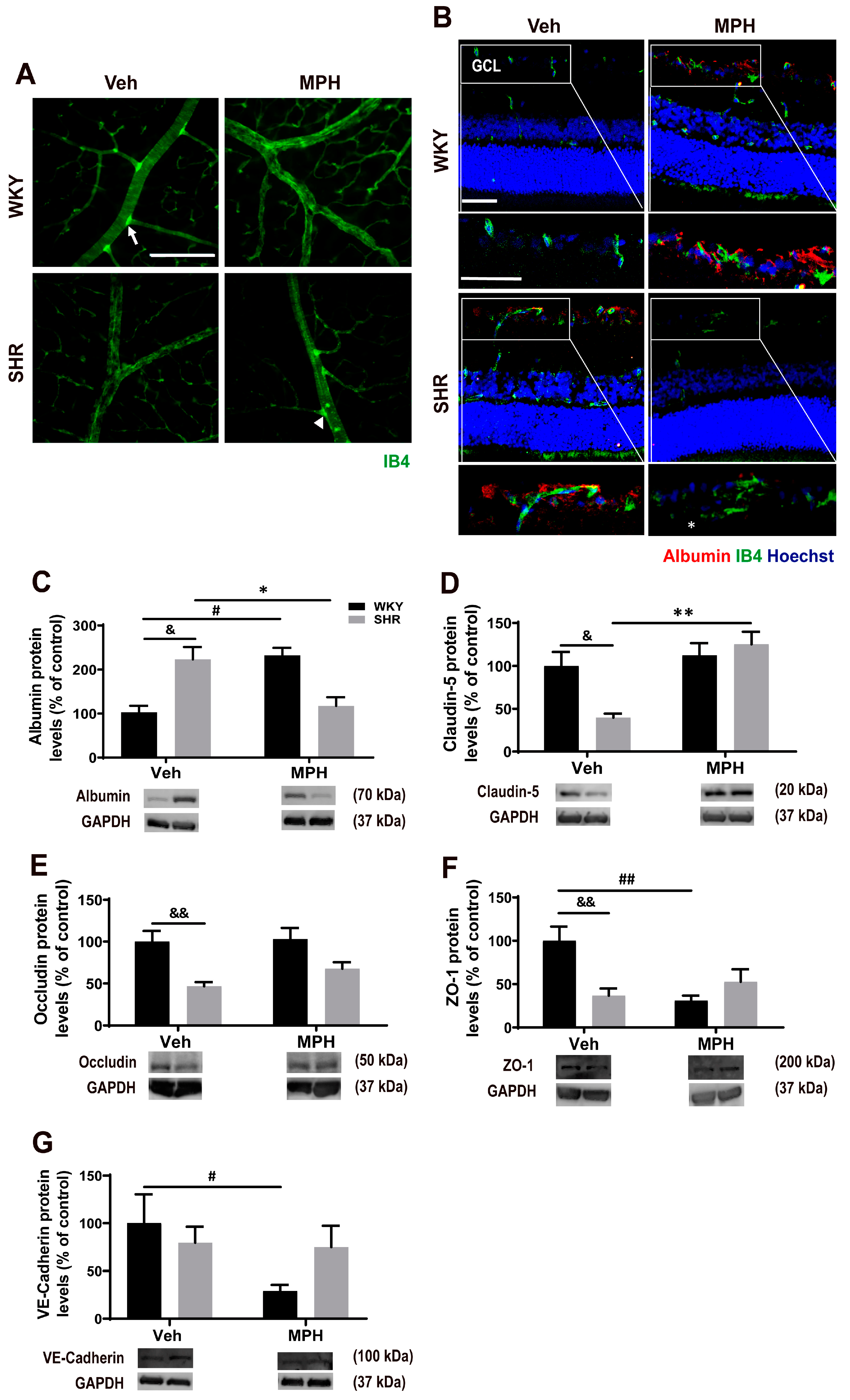
3.4. Astrocytic and Retinal Vascular Alterations in ADHD Animal Model and Impact of MPH Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Posner, J.; Polanczyk, G.V.; Sonuga-Barke, E. Attention-deficit hyperactivity disorder. Lancet 2020, 395, 450–462. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Hoogman, M.; Bralten, J.; Hibar, D.P.; Mennes, M.; Zwiers, M.P.; Schweren, L.S.J.; van Hulzen, K.J.E.; Medland, S.E.; Shumskaya, E.; Jahanshad, N.; et al. Subcortical brain volume differences in participants with attention deficit hyperactivity disorder in children and adults: A cross-sectional mega-analysis. Lancet Psychiatry 2017, 4, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Coelho-Santos, V.; Cardoso, F.L.; Leitão, R.A.; Fontes-Ribeiro, C.A.; Silva, A.P. Impact of developmental exposure to methylphenidate on rat brain’s immune privilege and behavior: Control versus ADHD model. Brain Behav. Immun. 2018, 68, 169–182. [Google Scholar] [CrossRef]
- Markowitz, J.S.; Melchert, P.W. The Pharmacokinetics and pharmacogenomics of psychostimulants. Child Adolesc. Psychiatr. Clin. N. Am. 2022, 31, 393–416. [Google Scholar] [CrossRef]
- Coelho-Santos, V.; Socodato, R.; Portugal, C.; Leitão, R.A.; Rito, M.; Barbosa, M.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Minshall, R.D.; et al. Methylphenidate-triggered ROS generation promotes caveolae-mediated transcytosis via Rac1 signaling and c-Src-dependent caveolin-1 phosphorylation in human brain endothelial cells. Cell Mol. Life Sci. 2016, 73, 4701–4716. [Google Scholar] [CrossRef]
- Coelho-Santos, V.; Cardoso, F.L.; Magalhães, A.; Ferreira-Teixeira, M.; Leitão, R.A.; Gomes, C.; Rito, M.; Barbosa, M.; Fontes-Ribeiro, C.A.; Silva, A.P. Effect of chronic methylphenidate treatment on hippocampal neurovascular unit and memory performance in late adolescent rats. Eur. Neuropsychopharmacol. 2019, 29, 195–210. [Google Scholar] [CrossRef]
- London, A.; Benhar, I.; Schwartz, M. The retina as a window to the brain-from eye research to CNS disorders. Nat. Rev. Neurol. 2013, 9, 44–53. [Google Scholar] [CrossRef]
- Robbins, C.B.; Thompson, A.C.; Bhullar, P.K.; Koo, H.Y.; Agrawal, R.; Soundararajan, S.; Yoon, S.P.; Polascik, B.W.; Scott, B.L.; Grewal, D.S.; et al. Characterization of retinal microvascular and choroidal structural changes in Parkinson disease. JAMA Ophthalmol. 2021, 139, 182–188. [Google Scholar] [CrossRef]
- Pan, J.; Zhou, Y.; Xiang, Y.; Yu, J. Retinal nerve fiber layer thickness changes in Schizophrenia: A meta-analysis of case-control studies. Psychiatry Res. 2018, 270, 786–791. [Google Scholar] [CrossRef]
- Schönfeldt-Lecuona, C.; Schmidt, A.; Kregel, T.; Kassubek, J.; Dreyhaupt, J.; Freudenmann, R.W.; Connemann, B.J.; Pinkhardt, E.H.; Gahr, M. Retinal changes in patients with major depressive disorder—A controlled optical coherence tomography study. J. Affect. Disord. 2018, 227, 665–671. [Google Scholar] [CrossRef]
- Kalenderoglu, A.; Sevgi-Karadag, A.; Celik, M.; Egilmez, O.B.; Han-Almis, B.; Ozen, M.E. Can the retinal ganglion cell layer (GCL) volume be a new marker to detect neurodegeneration in bipolar disorder? Compr. Psychiatry 2016, 67, 66–72. [Google Scholar] [CrossRef] [PubMed]
- DeCarlo, D.K.; Swanson, M.; McGwin, G.; Visscher, K.; Owsley, C. ADHD and vision problems in the national survey of children’s health. Optom. Vis. Sci. 2016, 93, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari, M.A.; Berquin, P.; Missonnier, P.; Aarabi, A.; Debatisse, D.; De Broca, A.; Wallois, F. Visual sensory processing deficit in the occipital region in children with attention-deficit / hyperactivity disorder as revealed by event-related potentials during cued continuous performance test. Neurophysiol. Clin. 2010, 40, 137–149. [Google Scholar] [CrossRef]
- Ulucan Atas, P.B.; Ceylan, O.M.; Dönmez, Y.E.; Ozel Ozcan, O. Ocular findings in patients with attention deficit and hyperactivity. Int. Ophthalmol. 2020, 40, 3105–3113. [Google Scholar] [CrossRef]
- Werner, A.L.; Tebartz van Elst, L.; Ebert, D.; Friedel, E.; Bubl, A.; Clement, H.W.; Lukačin, R.; Bach, M.; Bubl, E. Normalization of increased retinal background noise after ADHD treatment: A neuronal correlate. Schizophr. Res. 2020, 219, 77–83. [Google Scholar] [CrossRef]
- Clements, K.M.; Devonshire, I.M.; Reynolds, J.N.; Overton, P.G. Enhanced visual responses in the superior colliculus in an animal model of attention-deficit hyperactivity disorder and their suppression by D-amphetamine. Neuroscience 2014, 274, 289–298. [Google Scholar] [CrossRef]
- Brace, L.R.; Kraev, I.; Rostron, C.L.; Stewart, M.G.; Overton, P.G.; Dommett, E.J. Altered visual processing in a rodent model of attention-deficit hyperactivity disorder. Neuroscience 2015, 303, 364–377. [Google Scholar] [CrossRef] [Green Version]
- Navarra, R.L.; Clark, B.D.; Zitnik, G.A.; Waterhouse, B.D. Methylphenidate and atomoxetine enhance sensory-evoked neuronal activity in the visual thalamus of male rats. Exp. Clin. Psychopharmacol. 2013, 21, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Sagvolden, T. Behavioral validation of the spontaneously hypertensive rat (SHR) as an animal model of attention-deficit/hyperactivity disorder (AD/HD). Neurosci. Biobehav. Rev. 2000, 24, 31–39. [Google Scholar] [CrossRef]
- Adriani, W.; Caprioli, A.; Granstrem, O.; Carli, M.; Laviola, G. The spontaneously hypertensive-rat as an animal model of ADHD: Evidence for impulsive and non-impulsive subpopulations. Neurosci. Biobehav. Rev. 2003, 27, 639–651. [Google Scholar] [CrossRef]
- Leffa, D.T.; Panzenhagen, A.C.; Salvi, A.A.; Bau, C.H.D.; Pires, G.N.; Torres, I.L.S.; Rohde, L.A.; Rovaris, D.L.; Grevet, E.H. Systematic review and meta-analysis of the behavioral effects of methylphenidate in the spontaneously hypertensive rat model of attention-deficit/hyperactivity disorder. Neurosci. Biobehav. Rev. 2019, 100, 166–179. [Google Scholar] [CrossRef]
- Sagvolden, T.; Russell, V.A.; Aase, H.; Johansen, E.B.; Farshbaf, M. Rodent models of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1239–1247. [Google Scholar] [CrossRef]
- Martins, J.; Castelo-Branco, M.; Batista, A.; Oliveiros, B.; Santiago, A.R.; Galvão, J.; Fernandes, E.; Carvalho, F.; Cavadas, C.; Ambrósio, A.F. Effects of 3,4-methylenedioxymethamphetamine administration on retinal physiology in the rat. PLoS ONE 2011, 6, e29583. [Google Scholar] [CrossRef]
- Boia, R.; Dias, P.A.N.; Galindo-Romero, C.; Ferreira, H.; Aires, I.D.; Vidal-Sanz, M.; Agudo-Barriuso, M.; Bernardes, R.; Santos, P.F.; de Sousa, H.C.; et al. Intraocular implants loaded with A3R agonist rescue retinal ganglion cells from ischemic damage. J. Control. Release 2022, 343, 469–481. [Google Scholar] [CrossRef]
- Boia, R.; Elvas, F.; Madeira, M.H.; Aires, I.D.; Rodrigues-Neves, A.C.; Tralhão, P.; Szabó, E.C.; Baqi, Y.; Müller, C.E.; Tomé, Â.R.; et al. Treatment with A2A receptor antagonist KW6002 and caffeine intake regulate microglia reactivity and protect retina against transient ischemic damage. Cell Death Dis. 2017, 8, e3065. [Google Scholar] [CrossRef]
- Leitão, R.A.; Sereno, J.; Castelhano, J.M.; Gonçalves, S.I.; Coelho-Santos, V.; Fontes-Ribeiro, C.; Castelo-Branco, M.; Silva, A.P. Aquaporin-4 as a New target against methamphetamine-induced brain alterations: Focus on the neurogliovascular unit and motivational behavior. Mol. Neurobiol. 2017, 55, 2056–2069. [Google Scholar] [CrossRef]
- Salinas-Navarro, M.; Mayor-Torroglosa, S.; Jiménez-López, M.; Avilés-Trigueros, M.; Holmes, T.M.; Lund, R.D.; Villegas-Pérez, M.P.; Vidal-Sanz, M. A computerized analysis of the entire retinal ganglion cell population and its spatial distribution in adult rats. Vision Res. 2009, 49, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Leitão, R.A.; Fontes-Ribeiro, C.A.; Silva, A.P. The effect of parthenolide on methamphetamine-induced blood-brain barrier and astrocyte alterations. Eur. J. Clin. Investig. 2021, 52, e13694. [Google Scholar] [CrossRef]
- Khare, P.; Datusalia, A.K.; Sharma, S.S. Parthenolide, an NF-κB inhibitor ameliorates diabetes-induced behavioural deficit, neurotransmitter imbalance and neuroinflammation in type 2 diabetes rat model. Neuromol. Med. 2016, 19, 101–112. [Google Scholar] [CrossRef]
- Baptista, F.I.; Pinto, M.J.; Elvas, F.; Martins, T.; Almeida, R.D.; Ambrósio, A.F. Diabetes induces changes in KIF1A, KIF5B and dynein distribution in the rat retina: Implications for axonal transport. Exp. Eye Res. 2014, 127, 91–103. [Google Scholar] [CrossRef]
- Rashid, K.; Akhtar-Schaefer, I.; Langmann, T. Microglia in retinal degeneration. Front. Immunol. 2019, 10, 1975. [Google Scholar] [CrossRef] [Green Version]
- Cunha-Vaz, J.; Bernardes, R.; Lobo, C. Blood-retinal barrier. Eur. J. Ophthalmol. 2010, 21 (Suppl. S6), S3–S9. [Google Scholar] [CrossRef]
- Martínez-Carmona, M.; Lucas-Ruiz, F.; Gallego-Ortega, A.; Galindo-Romero, C.; Norte-Muñoz, M.; González-Riquelme, M.J.; Valiente-Soriano, F.J.; Vidal-Sanz, M.; Agudo-Barriuso, M. Ly6c as a new marker of mouse blood vessels: Qualitative and quantitative analyses on intact and ischemic retinas. Int. J. Mol. Sci. 2021, 23, 19. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Q.; Muir, E.R.; Kiel, J.W.; Duong, T.Q. Retinal Vascular and Anatomical Features in the Spontaneously Hypertensive Rat. Curr. Eye Res. 2020, 45, 1422–1429. [Google Scholar] [CrossRef]
- Hergüner, A.; Alpfidan, İ.; Yar, A.; Erdoğan, E.; Metin, Ö.; Sakarya, Y.; Hergüner, S. Retinal nerve fiber layer thickness in children with ADHD. J. Atten. Disord. 2016, 22, 619–626. [Google Scholar] [CrossRef]
- Li, S.L.; Kam, K.W.; Chee, A.S.H.; Zhang, X.J.; Chen, L.J.; Yip, W.W.K.; Tham, C.C.; Young, A.L.; Wong, I.C.K.; Ip, P.; et al. The association between attention-deficit/hyperactivity disorder and retinal nerve fiber/ganglion cell layer thickness measured by optical coherence tomography: A systematic review and meta-analysis. Int. Ophthalmol. 2021, 41, 3211–3221. [Google Scholar] [CrossRef]
- Sripada, C.S.; Kessler, D.; Angstadt, M. Lag in maturation of the brain’s intrinsic functional architecture in attention-deficit/hyperactivity disorder. Proc. Natl. Acad. Sci. USA 2014, 111, 14259–14264. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.L.; Thompson, P.M. Typical and atypical brain development: A review of neuroimaging studies. Dialogues Clin. Neurosci. 2013, 15, 359–384. [Google Scholar] [CrossRef]
- Işik, Ü.; Kaygisiz, M. Assessment of intraocular pressure, macular thickness, retinal nerve fiber layer, and ganglion cell layer thicknesses: Ocular parameters and optical coherence tomography findings in attention-deficit/hyperactivity disorder. Braz. J. Psychiatry 2020, 42, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Chen, A.L.; Braverman, E.R.; Comings, D.E.; Chen, T.J.; Arcuri, V.; Blum, S.H.; Downs, B.W.; Waite, R.L.; Notaro, A.; et al. Attention-deficit-hyperactivity disorder and reward deficiency syndrome. Neuropsychiatr. Dis. Treat. 2008, 4, 893–918. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Wang, G.J.; Kollins, S.H.; Wigal, T.L.; Newcorn, J.H.; Telang, F.; Fowler, J.S.; Zhu, W.; Logan, J.; Ma, Y.; et al. Evaluating dopamine reward pathway in ADHD: Clinical implications. JAMA 2009, 302, 1084–1091. [Google Scholar] [CrossRef]
- Roy, S.; Field, G.D. Dopaminergic modulation of retinal processing from starlight to sunlight. J. Pharmacol. Sci. 2019, 140, 86–93. [Google Scholar] [CrossRef]
- Roessner, V.; Banaschewski, T.; Fillmer-Otte, A.; Becker, A.; Albrecht, B.; Uebel, H.; Sergeant, J.; Tannock, R.; Rothenberger, A. Color perception deficits in co-existing attention-deficit/hyperactivity disorder and chronic tic disorders. J. Neural. Transm. 2008, 115, 235–239. [Google Scholar] [CrossRef]
- Marrocco, E.; Indrieri, A.; Esposito, F.; Tarallo, V.; Carboncino, A.; Alvino, F.G.; De Falco, S.; Franco, B.; De Risi, M.; De Leonibus, E. α-synuclein overexpression in the retina leads to vision impairment and degeneration of dopaminergic amacrine cells. Sci. Rep. 2020, 10, 9619. [Google Scholar] [CrossRef]
- Tran, K.K.N.; Wong, V.H.Y.; Lim, J.K.H.; Shahandeh, A.; Hoang, A.; Finkelstein, D.I.; Bui, B.V.; Nguyen, C.T.O. Characterization of retinal function and structure in the MPTP murine model of Parkinson’s disease. Sci. Rep. 2022, 12, 7610. [Google Scholar] [CrossRef]
- Weil, R.S.; Schrag, A.E.; Warren, J.D.; Crutch, S.J.; Lees, A.J.; Morris, H.R. Visual dysfunction in Parkinson’s disease. Brain 2016, 139, 2827–2843. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.H.; Kang, E.Y.; Liu, P.K.; Levi, S.R.; Wang, H.H.; Tseng, Y.J.; Seo, G.H.; Lee, H.; Yeh, L.K.; Chen, K.J.; et al. Photoreceptor manifestations of primary mitochondrial optic nerve disorders. Investig. Ophthalmol. Vis. Sci. 2022, 63, 5. [Google Scholar] [CrossRef]
- Fabian, I.D.; Kinori, M.; Ancri, O.; Spierer, A.; Tsinman, A.; Ben Simon, G.J. The possible association of attention deficit hyperactivity disorder with undiagnosed refractive errors. J. AAPOS 2013, 17, 507–511. [Google Scholar] [CrossRef]
- Martin, L.; Aring, E.; Landgren, M.; Hellström, A.; Andersson Grönlund, M. Visual fields in children with attention-deficit/hyperactivity disorder before and after treatment with stimulants. Acta. Ophthalmol. 2008, 86, 259–264. [Google Scholar] [CrossRef]
- Skalny, A.V.; Mazaletskaya, A.L.; Zaitseva, I.P.; Skalny, A.A.; Spandidos, D.A.; Tsatsakis, A.; Lobanova, Y.N.; Skalnaya, M.G.; Aschner, M.; Tinkov, A.A. Alterations in serum amino acid profiles in children with attention deficit/hyperactivity disorder. Biomed. Rep. 2021, 14, 47. [Google Scholar] [CrossRef]
- Huang, X.; Wang, M.; Zhang, Q.; Chen, X.; Wu, J. The role of glutamate receptors in attention-deficit/hyperactivity disorder: From physiology to disease. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 272–286. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, A.; Shi, M.Y.; Yan, Z. Disrupted glutamatergic transmission in prefrontal cortex contributes to behavioral abnormality in an animal model of ADHD. Neuropsychopharmacology 2017, 42, 2096–2104. [Google Scholar] [CrossRef] [Green Version]
- Antes, R.; Ezra-Elia, R.; Weinberger, D.; Solomon, A.; Ofri, R.; Michaelson, D.M. ApoE4 induces synaptic and ERG impairments in the retina of young targeted replacement apoE4 mice. PLoS ONE 2013, 8, e64949. [Google Scholar] [CrossRef]
- Navarra, R.L.; Clark, B.D.; Gargiulo, A.T.; Waterhouse, B.D. Methylphenidate enhances early-stage sensory processing and rodent performance of a visual signal detection task. Neuropsychopharmacology 2016, 42, 1326–1337. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Tao, L.; Li, L. Long-term systemic treatment with methamphetamine causes retinal damage in CD1 mice. Int. J. Toxicol. 2018, 37, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.H.; Boia, R.; Santos, P.F.; Ambrósio, A.F.; Santiago, A.R. Contribution of microglia-mediated neuroinflammation to retinal degenerative diseases. Mediat. Inflamm. 2015, 2015, 673090. [Google Scholar] [CrossRef] [Green Version]
- Zabel, M.K.; Zhao, L.; Zhang, Y.; Gonzalez, S.R.; Ma, W.; Wang, X.; Fariss, R.N.; Wong, W.T. Microglial phagocytosis and activation underlying photoreceptor degeneration is regulated by CX3CL1-CX3CR1 signaling in a mouse model of retinitis pigmentosa. Glia 2016, 64, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- García-Ayuso, D.; Di Pierdomenico, J.; Vidal-Sanz, M.; Villegas-Pérez, M.P. Retinal ganglion cell death as a late remodeling effect of photoreceptor degeneration. Int. J. Mol. Sci. 2019, 20, 4649. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Peng, B.; Lin, B. Fractalkine receptor regulates microglial neurotoxicity in an experimental mouse glaucoma model. Glia 2014, 62, 1943–1954. [Google Scholar] [CrossRef]
- Yokokura, M.; Takebasashi, K.; Takao, A.; Nakaizumi, K.; Yoshikawa, E.; Futatsubashi, M.; Suzuki, K.; Nakamura, K.; Yamasue, H.; Ouchi, Y. In vivo imaging of dopamine D1 receptor and activated microglia in attention-deficit/hyperactivity disorder: A positron emission tomography study. Mol. Psychiatry 2020, 26, 4958–4967. [Google Scholar] [CrossRef]
- Vecino, E.; Rodriguez, F.D.; Ruzafa, N.; Pereiro, X.; Sharma, S.C. Glia-neuron interactions in the mammalian retina. Prog. Retin. Eye Res. 2016, 51, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudnicky, F.; Zhang, J.D.; Kim, B.K.; Pandya, N.J.; Lan, Y.; Sach-Peltason, L.; Ragelle, H.; Strassburger, P.; Gruener, S.; Lazendic, M.; et al. Inducers of the endothelial cell barrier identified through chemogenomic screening in genome-edited hPSC-endothelial cells. Proc. Natl. Acad. Sci. USA 2020, 117, 19854–19865. [Google Scholar] [CrossRef]
- Wang, J.; Li, K.L.; Shukla, A.; Beroun, A.; Ishikawa, M.; Huang, X.; Wang, Y.; Wang, Y.Q.; Yang, Y.; Bastola, N.D.; et al. Cocaine triggers astrocyte-mediated synaptogenesis. Biol. Psychiatry 2021, 89, 386–397. [Google Scholar] [CrossRef]
- Lee, M.; Leskova, W.; Eshaq, R.S.; Harris, N.R. Acute changes in the retina and central retinal artery with methamphetamine. Exp. Eye Res. 2020, 193, 107964. [Google Scholar] [CrossRef]
- Coelho-Santos, V.; Leitão, R.A.; Cardoso, F.L.; Palmela, I.; Rito, M.; Barbosa, M.; Brito, M.A.; Fontes-Ribeiro, C.A.; Silva, A.P. The TNF-α/NF-κB signaling pathway has a key role in methamphetamine-induced blood-brain barrier dysfunction. J. Cereb. Blood Flow Metab. 2015, 35, 1260–1271. [Google Scholar] [CrossRef] [Green Version]
- Martins, T.; Burgoyne, T.; Kenny, B.A.; Hudson, N.; Futter, C.E.; Ambrósio, A.F.; Silva, A.P.; Greenwood, J.; Turowski, P. Methamphetamine-induced nitric oxide promotes vesicular transport in blood-brain barrier endothelial cells. Neuropharmacology 2013, 65, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Albarral, J.A.; Salazar, J.J.; de Hoz, R.; Marco, E.M.; Martín-Sánchez, B.; Flores-Salguero, E.; Salobrar-García, E.; López-Cuenca, I.; Barrios-Sabador, V.; Avilés-Trigueros, M.; et al. Retinal molecular changes are associated with neuroinflammation and loss of RGCs in an experimental model of glaucoma. Int. J. Mol. Sci. 2021, 22, 2066. [Google Scholar] [CrossRef]
- Oksanen, M.; Lehtonen, S.; Jaronen, M.; Goldsteins, G.; Hämäläinen, R.H.; Koistinaho, J. Astrocyte alterations in neurodegenerative pathologies and their modeling in human induced pluripotent stem cell platforms. Cell. Mol. Life. Sci. 2019, 76, 2739–2760. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Trotta, M.C.; Gesualdo, C.; Petrillo, F.; Lepre, C.C.; Della Corte, A.; Cavasso, G.; Maggiore, G.; Hermenean, A.; Simonelli, F.; D’Amico, M.; et al. Resolution of inflammation in retinal disorders: Briefly the state. Int. J. Mol. Sci. 2022, 23, 4501. [Google Scholar] [CrossRef]
- Canedo, T.; Portugal, C.C.; Socodato, R.; Almeida, T.O.; Terceiro, A.F.; Bravo, J.; Silva, A.I.; Magalhães, J.D.; Guerra-Gomes, S.; Oliveira, J.F.; et al. Astrocyte-derived TNF and glutamate critically modulate microglia activation by methamphetamine. Neuropsychopharmacology 2021, 46, 2358–2370. [Google Scholar] [CrossRef]
- Rohowetz, L.J.; Kraus, J.G.; Koulen, P. Reactive Oxygen Species-Mediated Damage of Retinal Neurons: Drug Development Targets for Therapies of Chronic Neurodegeneration of the Retina. Int. J. Mol. Sci. 2018, 19, 3362. [Google Scholar] [CrossRef] [Green Version]
- Dinh, Q.N.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Roles of inflammation, oxidative stress, and vascular dysfunction in hypertension. Biomed Res. Int. 2014, 2014, 406960. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanches, E.S.; Boia, R.; Leitão, R.A.; Madeira, M.H.; Fontes-Ribeiro, C.A.; Ambrósio, A.F.; Fernandes, R.; Silva, A.P. Attention-Deficit/Hyperactivity Disorder Animal Model Presents Retinal Alterations and Methylphenidate Has a Differential Effect in ADHD versus Control Conditions. Antioxidants 2023, 12, 937. https://doi.org/10.3390/antiox12040937
Sanches ES, Boia R, Leitão RA, Madeira MH, Fontes-Ribeiro CA, Ambrósio AF, Fernandes R, Silva AP. Attention-Deficit/Hyperactivity Disorder Animal Model Presents Retinal Alterations and Methylphenidate Has a Differential Effect in ADHD versus Control Conditions. Antioxidants. 2023; 12(4):937. https://doi.org/10.3390/antiox12040937
Chicago/Turabian StyleSanches, Eliane S., Raquel Boia, Ricardo A. Leitão, Maria H. Madeira, Carlos A. Fontes-Ribeiro, António Francisco Ambrósio, Rosa Fernandes, and Ana Paula Silva. 2023. "Attention-Deficit/Hyperactivity Disorder Animal Model Presents Retinal Alterations and Methylphenidate Has a Differential Effect in ADHD versus Control Conditions" Antioxidants 12, no. 4: 937. https://doi.org/10.3390/antiox12040937