
A Scientometric Study to a Critical Review on Promising Anticancer and Neuroprotective Compounds: Citrus Flavonoids

 , ,
, ,
Abstract
:
1. Introduction
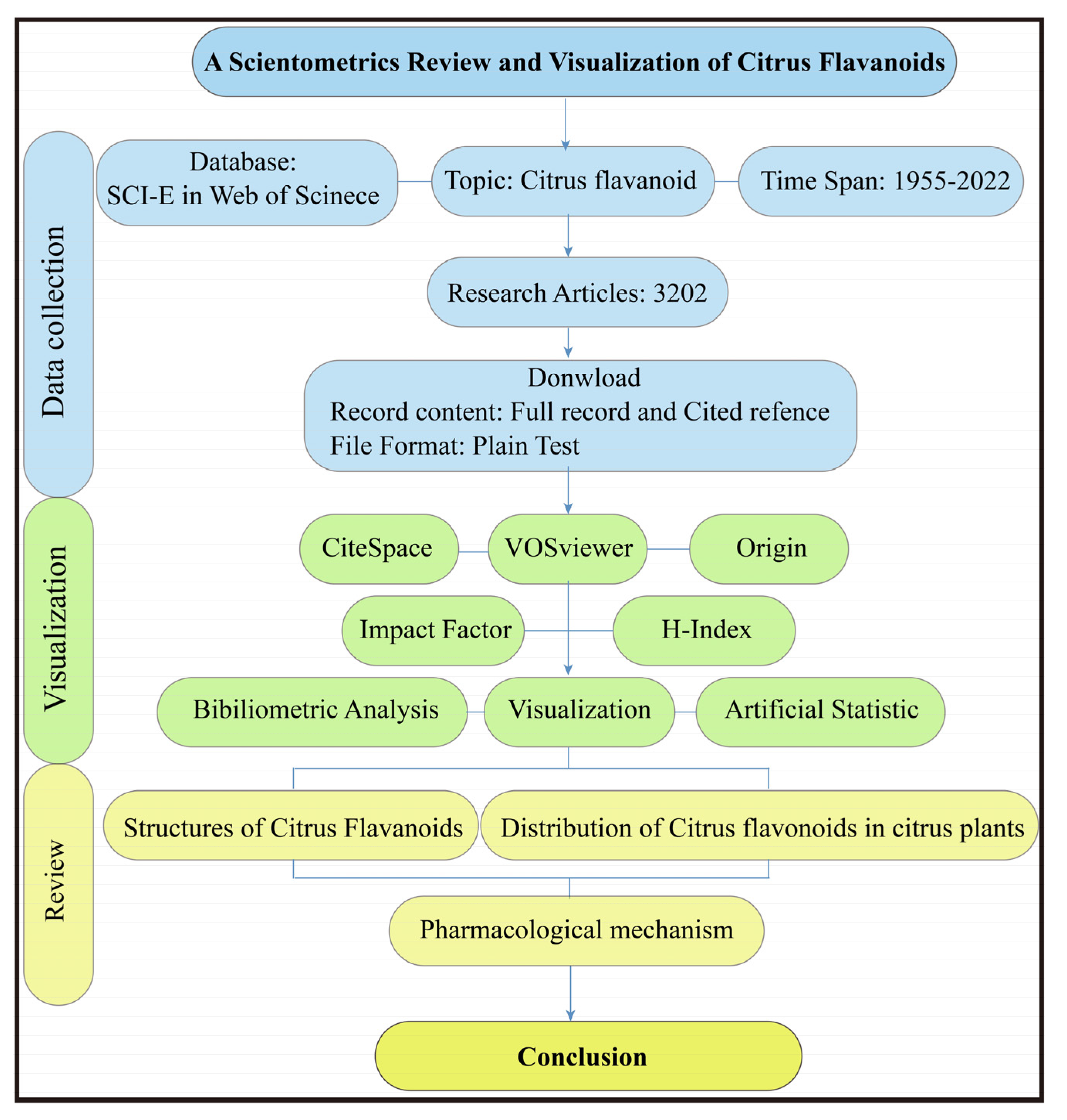
2. Materials and Methods
2.1. Literature Search and Data Download
2.2. Scientometric Analysis and Visualization
3. Results and Discussion
3.1. Scientometric Analysis
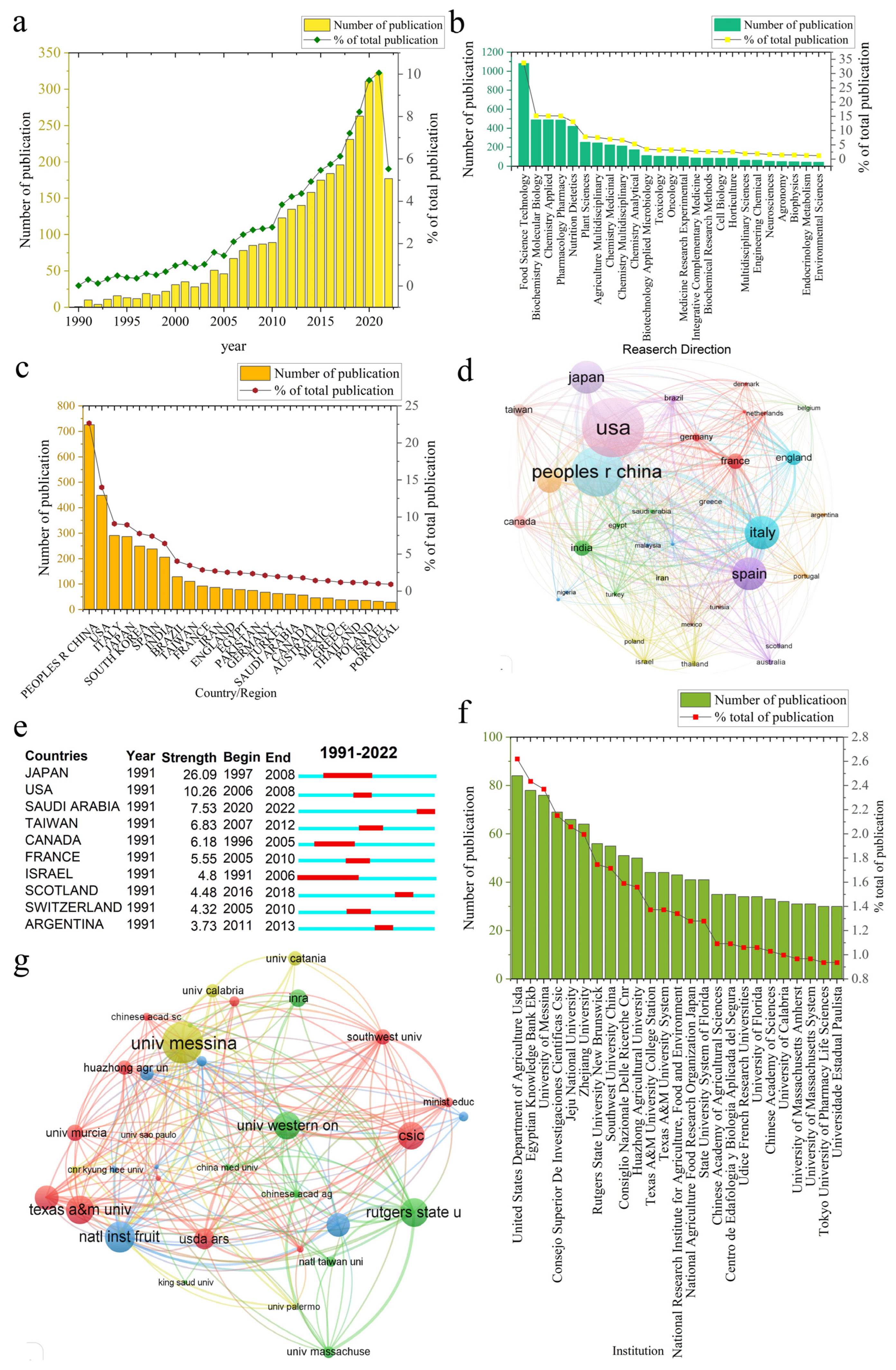
3.1.1. General Analysis
3.1.2. Journal Analysis
3.1.3. Country/Region and Institution Analysis
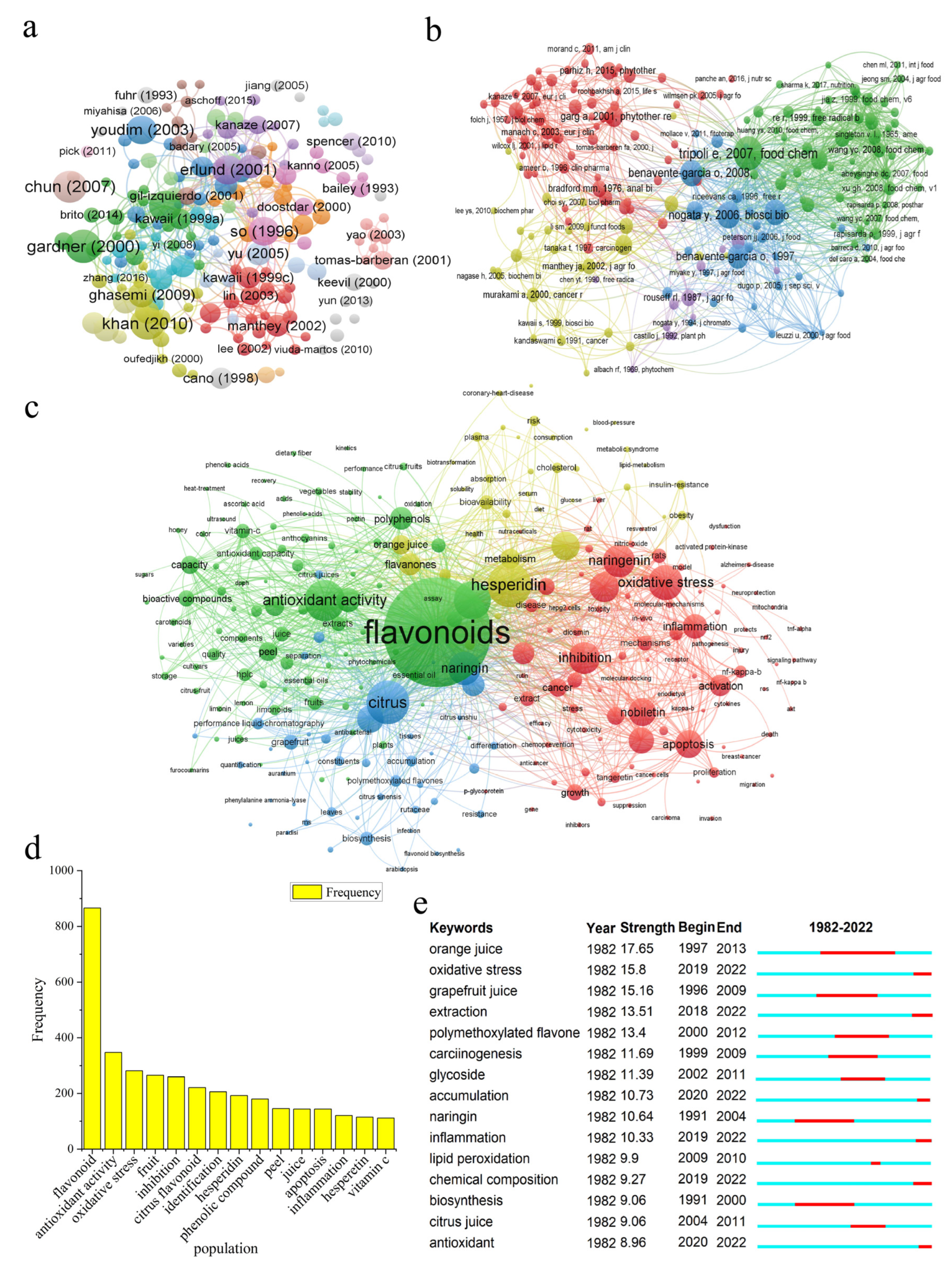
3.1.4. Literature and Cited Reference Analysis
3.1.5. Keywords Analysis
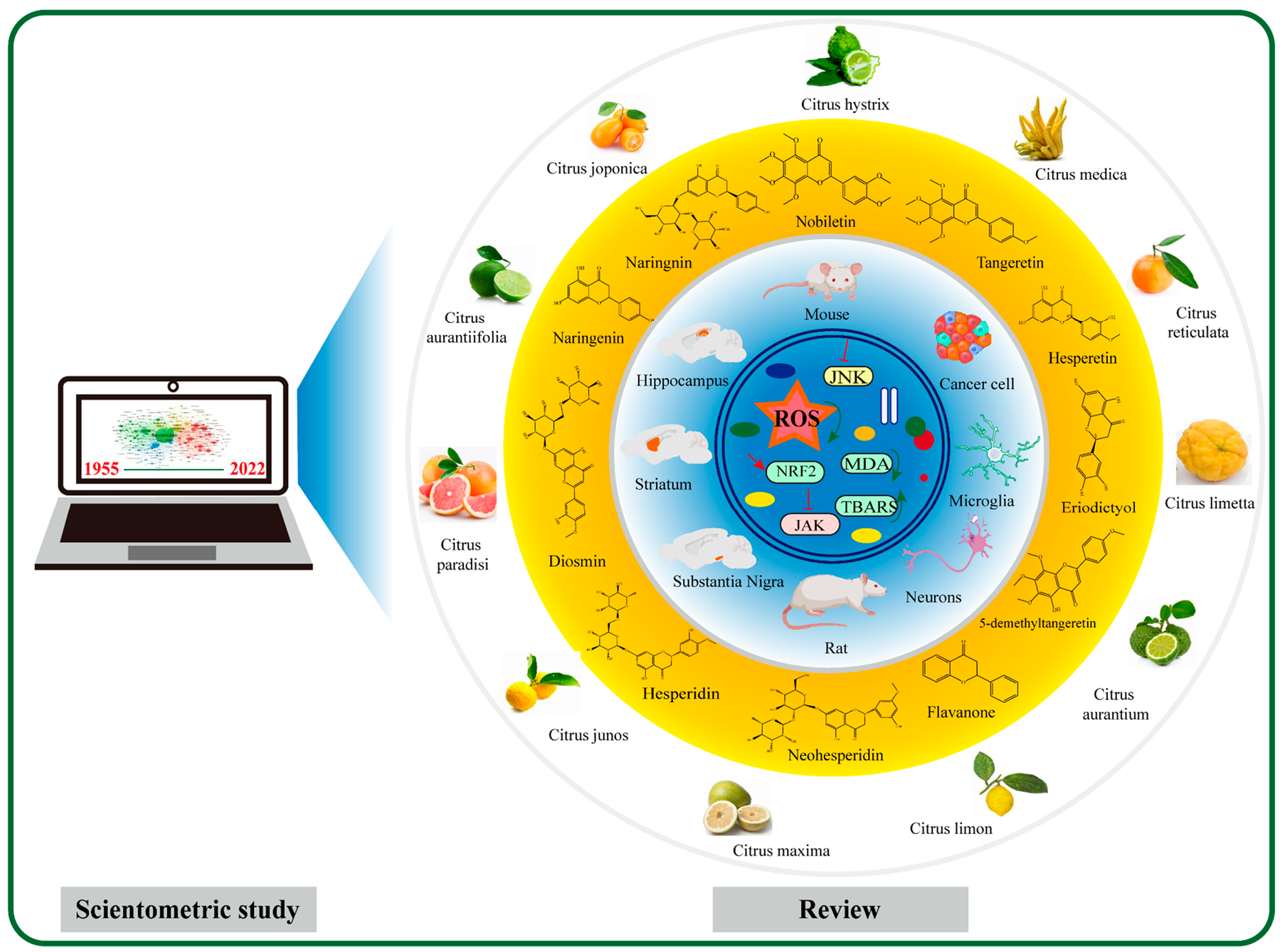
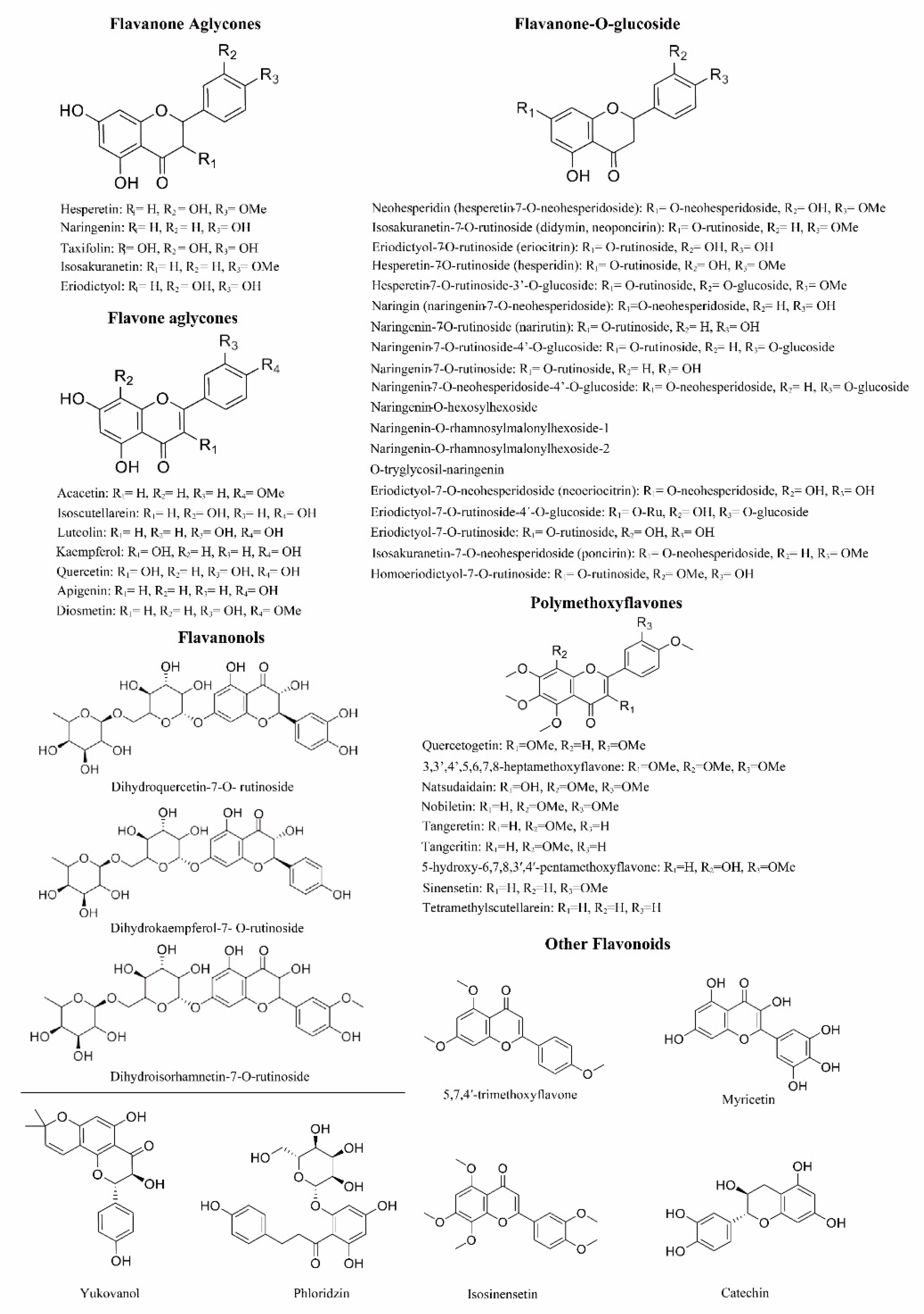
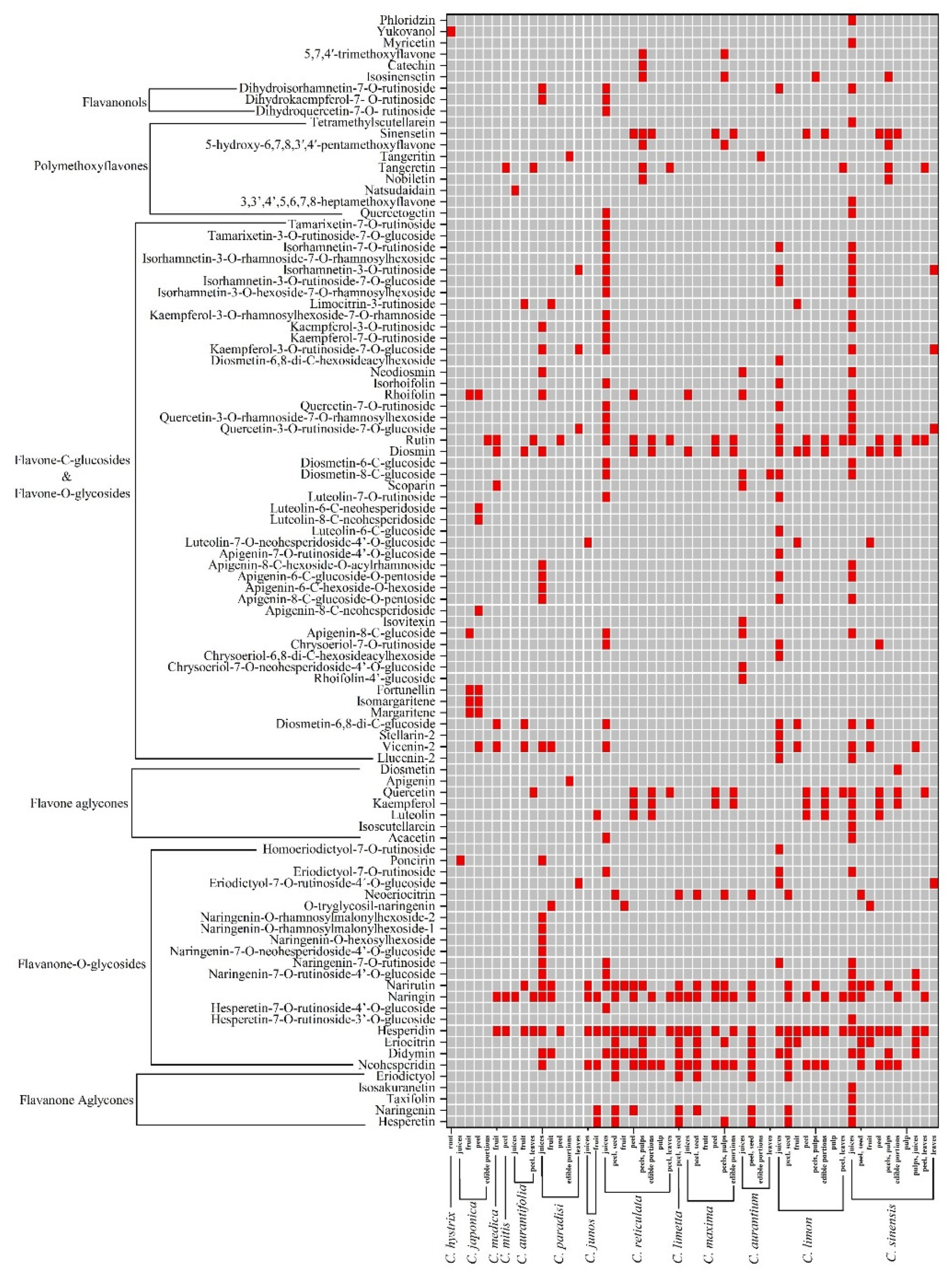
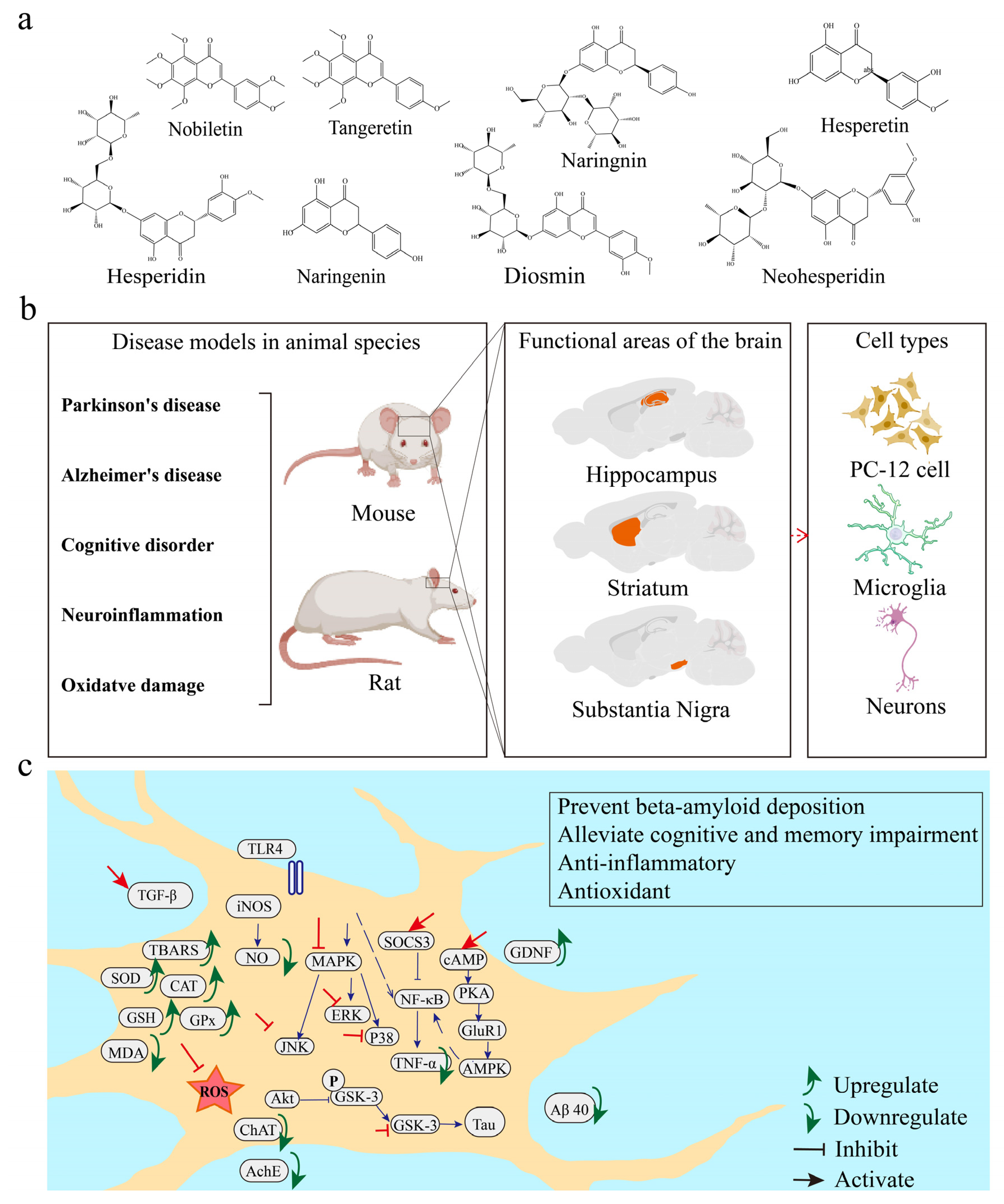
3.2. Chemical Structures and Sources of Citrus Flavonoids
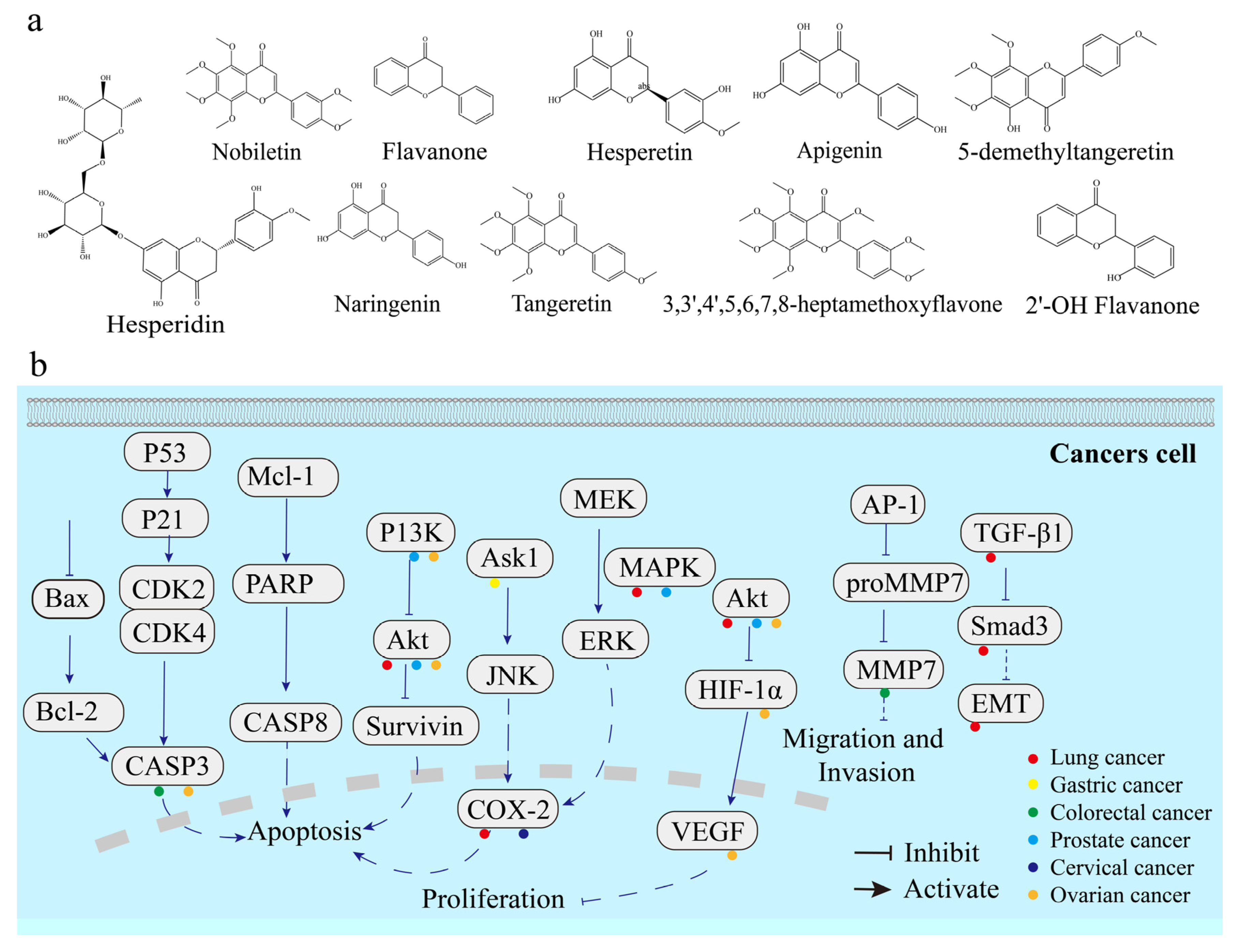
3.3. Citrus Flavonoids and Cancers
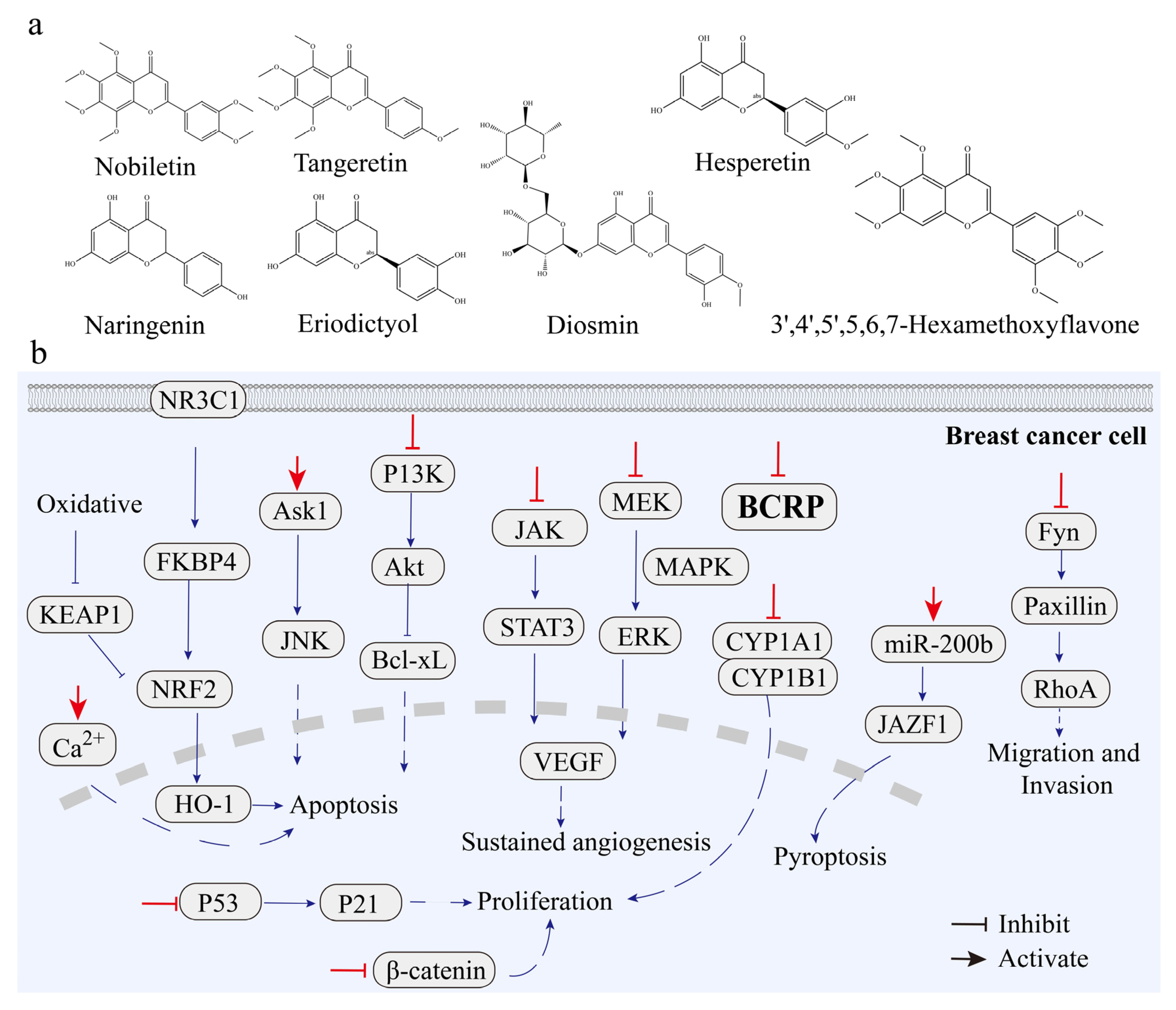
3.3.1. Breast Cancer
3.3.2. Colorectal Cancer
3.3.3. Gastric Cancer
3.3.4. Lung Cancer
3.3.5. Liver Cancer
3.3.6. Prostate Cancer
3.3.7. Cervical Cancer
3.3.8. Ovarian Cancer
3.3.9. Epidermal Carcinoma
3.4. Neuroprotective Effects of Citrus Flavonoids
3.5. Citrus Flavonoid and Metabolic Disease
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, G.A.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Food and Agriculture Statistical Yearbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Han, N.D.; Cheng, J.Y.; Delannoy-Bruno, O.; Webber, D.; Terrapon, N.; Henrissat, B.; Rodionov, D.A.; Arzamasov, A.A.; Osterman, A.L.; Hayashi, D.K.; et al. Microbial liberation of N-methylserotonin from orange fiber in gnotobiotic mice and humans. Cell 2022, 185, 2495–2509. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.Y.T.; Le, T.D.; Dao, P.T.; Bach, G.L.; Huynh, P.X.; Tran, Q.N. Evaluation of different extraction methods on the polyphenols yield, flavonoids yield, and antioxidant activity of the pomelo flavedo extract from Da Xanh (Citrus maxima burm merr.) variety. Food Sci. Technol. 2022, 42, 9. [Google Scholar] [CrossRef]
- Roowi, S.; Crozier, A. Flavonoids in Tropical Citrus Species. J. Agric. Food Chem. 2011, 59, 12217–12225. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; Wang, F.; Lian, Y.H.; Xiao, H.; Zheng, J.K. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Mahato, N.; Lee, Y.R. Extraction, characterization and biological activity of citrus flavonoids. Rev. Chem. Eng. 2019, 35, 265–284. [Google Scholar] [CrossRef]
- Peritz, B.C.; Bar-Ilan, J. The sources used by bibliometrics-scientometrics as reflected in references. Scientometrics 2002, 54, 269–284. [Google Scholar] [CrossRef]
- Hou, J.; Su, H.J.; Kuang, X.C.; Qin, W.C.; Liu, K.B.; Pan, K.X.; Zhang, B.K.; Yang, S.J.; Yang, S.H.; Peng, X.; et al. Knowledge Domains and Emerging Trends of Osteoblasts-Osteoclasts in Bone Disease From 2002 to 2021: A Bibliometrics Analysis and Visualization Study. Front. Endocrinol. 2022, 13, 14. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Zhu, Y.C.; Wang, Q.F.; Chang, T.T.; Liu, C.L.; Zhu, Y.; Wang, X.Q.; Cao, X.Y. Global Trends and Research Hotspots of Exercise for Intervening Diabetes: A Bibliometric Analysis. Front. Public Health 2022, 10, 15. [Google Scholar] [CrossRef]
- Giannos, P.; Kechagias, K.S.; Triantafyllidis, K.K.; Falagas, M.E. Spotlight on Early COVID-19 Research Productivity: A 1-Year Bibliometric Analysis. Front. Public Health 2022, 10, 8. [Google Scholar] [CrossRef]
- Long, J.Z.; Zhang, Y.S.; Liu, X.M.; Pan, M.Y.; Gao, Q. Exosomes in the Field of Neuroscience: A Scientometric Study and Visualization Analysis. Front. Neurol. 2022, 13, 13. [Google Scholar] [CrossRef]
- Gardner, P.T.; White, T.A.C.; McPhail, D.B.; Duthie, G.G. The relative contributions of vitamin C, carotenoids and phenolics to the antioxidant potential of fruit juices. Food Chem. 2000, 68, 471–474. [Google Scholar] [CrossRef]
- Khan, M.K.; Abert-Vian, M.; Fabiano-Tixier, A.S.; Dangles, O.; Chemat, F. Ultrasound-assisted extraction of polyphenols (flavanone glycosides) from orange (Citrus sinensis L.) peel. Food Chem. 2010, 119, 851–858. [Google Scholar] [CrossRef]
- Chun, O.K.; Chung, S.J.; Song, W.O. Estimated dietary flavonoid intake and major food sources of US adults. J. Nutr. 2007, 137, 1244–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid composition of fruit tissues of Citrus species. Biosci. Biotechnol. Biochem. 2006, 70, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Erlund, I.; Meririnne, E.; Alfthan, G.; Aro, A. Plasma kinetics and urinary excretion of the flavanones naringenin and hesperetin in humans after ingestion of orange juice and grapefruit juice. J. Nutr. 2001, 131, 235–241. [Google Scholar] [CrossRef] [Green Version]
- So, F.V.; Guthrie, N.; Chambers, A.F.; Moussa, M.; Carroll, K.K. Inhibition of human breast cancer cell proliferation and delay of mammary tumorigenesis by flavonoids and citrus juices. Nutr. Cancer 1996, 26, 167–181. [Google Scholar] [CrossRef]
- Youdim, K.A.; Dobbie, M.S.; Kuhnle, G.; Proteggente, A.R.; Abbott, N.J.; Rice-Evans, C. Interaction between flavonoids and the blood-brain barrier: In vitro studies. J. Neurochem. 2003, 85, 180–192. [Google Scholar] [CrossRef]
- Ghasemi, K.; Ghasemi, Y.; Ebrahimzadeh, M.A. Antioxidant Activity, Phenol and Flavonoid Contents of 13 Citrus Species Peels nd Tissues. Pak. J. Pharm. Sci. 2009, 22, 277–281. [Google Scholar]
- Manach, C.; Morand, C.; Gil-Izquierdo, A.; Bouteloup-Demange, C.; Remesy, C. Bioavailability in humans of the flavanones hesperidin and narirutin after the ingestion of two doses of orange juice. Eur. J. Clin. Nutr. 2003, 57, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Marin, F.R.; Soler-Rivas, C.; Benavente-Garcia, O.; Castillo, J.; Perez-Alvarez, J.A. By-products from different citrus processes as a source of customized functional fibres. Food Chem. 2007, 100, 736–741. [Google Scholar] [CrossRef]
- Gorinstein, S.; Martin-Belloso, O.; Park, Y.S.; Haruenkit, R.; Lojek, A.; Ciz, M.; Caspi, A.; Libman, I.; Trakhtenberg, S. Comparison of some biochemical characteristics of different citrus fruits. Food Chem. 2001, 74, 309–315. [Google Scholar] [CrossRef]
- Yu, J.; Wang, L.M.; Walzem, R.L.; Miller, E.G.; Pike, L.M.; Patil, B.S. Antioxidant activity of Citrus limonoids, flavonoids, and coumarins. J. Agric. Food Chem. 2005, 53, 2009–2014. [Google Scholar] [CrossRef] [PubMed]
- Anagnostopoulou, M.A.; Kefalas, P.; Papageorgiou, V.P.; Assimopoulou, A.N.; Boskou, D. Radical scavenging activity of various extracts and fractions of sweet orange-peel (Citrus sinensis). Food Chem. 2006, 94, 19–25. [Google Scholar] [CrossRef]
- Rapisarda, P.; Tomaino, A.; Lo Cascio, R.; Bonina, F.; De Pasquale, A.; Saija, A. Antioxidant effectiveness as influenced by phenolic content of fresh orange juices. J. Agric. Food Chem. 1999, 47, 4718–4723. [Google Scholar] [CrossRef]
- Pfaltzgraff, L.A.; De Bruyn, M.; Cooper, E.C.; Budarin, V.; Clark, J.H. Food waste biomass: A resource for high-value chemicals. Green Chem. 2013, 15, 307–314. [Google Scholar] [CrossRef]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of Citrus flavonolds: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of their Molecular Mechanisms and Experimental Models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of Citrus Flavonoids, Naringin and Naringenin, on Metabolic Syndrome and Their Mechanisms of Action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef] [Green Version]
- Gattuso, G.; Barreca, D.; Gargiulli, C.; Leuzzi, U.; Caristi, C. Flavonoid composition of citrus juices. Molecules 2007, 12, 1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Z.; Xi, W.P.; Hu, Y.; Nie, C.; Zhou, Z.Q. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Khan, M.K.; Zill, E.H.; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Sun, Y.J.; Xi, W.P.; Shen, Y.; Qiao, L.P.; Zhong, L.Z.; Ye, X.Q.; Zhou, Z.Q. Phenolic compositions and antioxidant capacities of Chinese wild mandarin (Citrus reticulata Blanco) fruits. Food Chem. 2014, 145, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Assini, J.M.; Mulvihill, E.E.; Huff, M.W. Citrus flavonoids and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Morand, C.; Dubray, C.; Milenkovic, D.; Lioger, D.; Martin, J.F.; Scalbert, A.; Mazur, A. Hesperidin contributes to the vascular protective effects of orange juice: A randomized crossover study in healthy volunteers. Am. J. Clin. Nutr. 2011, 93, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Manthey, J.A.; Guthrie, N.; Grohmann, K. Biological properties of citrus flavonoids pertaining to cancer and inflammation. Curr. Med. Chem. 2001, 8, 135–153. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chuang, Y.C.; Hsu, H.W. The flavonoid, carotenoid and pectin content in peels of citrus cultivated in Taiwan. Food Chem. 2008, 106, 277–284. [Google Scholar] [CrossRef]
- Kanno, S.; Tomizawa, A.; Hiura, T.; Osani, Y.; Shouji, A.; Ujibe, M.; Ohtake, T.; Kimupa, K.; Ishikawa, M. Inhibitory effects of naringenin on tumor growth in human cancer cell lines and sarcoma S-180-implanted mice. Biol. Pharm. Bull. 2005, 28, 527–530. [Google Scholar] [CrossRef] [Green Version]
- Sp, N.; Kang, D.Y.; Kim, D.H.; Park, J.H.; Lee, H.G.; Kim, H.J.; Darvin, P.; Park, Y.M.; Yang, Y.M. Nobiletin Inhibits CD36-Dependent Tumor Angiogenesis, Migration, Invasion, and Sphere Formation Through the Cd36/Stat3/Nf-Kb Signaling Axis. Nutrients 2018, 10, 772. [Google Scholar] [CrossRef] [Green Version]
- Werk, A.N.; Cascorbi, I. Functional Gene Variants of CYP3A4. Clin. Pharm. Ther. 2014, 96, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Gougis, P.; Hilmi, M.; Geraud, A.; Mir, O.; Funck-Brentano, C. Potential cytochrome P450-mediated pharmacokinetic interactions between herbs, food, and dietary supplements and cancer treatments. Crit. Rev. Oncol. Hemat. 2021, 166, 103342. [Google Scholar] [CrossRef] [PubMed]
- Wangensteen, H.; Molden, E.; Christensen, H.; Malterud, K.E. Identification of epoxybergamottin as a CYP3A4 inhibitor in grapefruit peel. Eur. J. Clin. Pharm. 2003, 58, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.; Gattuso, G.; Barreca, D.; Haefeli, W.E. Nobiletin, sinensetin, and tangeretin are the main perpetrators in clementines provoking food-drug interactions in vitro. Food Chem. 2020, 319, 126578. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Cui, M.; Zhao, L.L.; Wang, D.S.; Tang, T.; Wang, W.B.; Wang, S.; Huang, H.Y.; Qiu, X.J. Potential Metabolic Drug-Drug Interaction of Citrus aurantium L. (Rutaceae) Evaluating by Its Effect on 3 CYP450. Front. Pharm. 2018, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- Sp, N.; Kang, D.Y.; Joung, Y.H.; Park, J.H.; Kim, W.S.; Lee, H.K.; Song, K.D.; Park, Y.M.; Yang, Y.M. Nobiletin Inhibits Angiogenesis by Regulating Src/FAK/STAT3-Mediated Signaling through PXN in ER+ Breast Cancer Cells. Int. J. Mol. Sci. 2017, 18, 935. [Google Scholar] [CrossRef] [Green Version]
- Surichan, S.; Androutsopoulos, V.P.; Sifakis, S.; Koutala, E.; Tsatsakis, A.; Arroo, R.R.J.; Boarder, M.R. Bioactivation of the citrus flavonoid nobiletin by CYP1 enzymes in MCF7 breast adenocarcinoma cells. Food Chem. Toxicol. 2012, 50, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Goan, Y.G.; Wu, W.T.; Liu, C.I.; Neoh, C.A.; Wu, Y.J. Involvement of mitochondrial dysfunction, endoplasmic reticulum stress, and the PI3K/AKT/mTOR pathway in nobiletin-induced apoptosis of human bladder cancer cells. Molecules 2019, 24, 2881. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Jin, L.T.; Lu, L.L.; Lu, X.Y.; Zhang, C.L.; Zhang, F.Y.; Liang, W. Naringenin reduces lung metastasis in a breast cancer resection model. Protein Cell 2011, 2, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Schindler, R.; Mentlein, R. Flavonoids and vitamin E reduce the release of the angiogenic peptide vascular endothelial growth factor from human tumor cells. J. Nutr. 2006, 136, 1477–1482. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Z.; Yang, B.; Huang, J.; Xiang, T.X.; Yin, X.D.; Wan, J.Y.; Luo, F.; Zhang, L.; Li, H.Y.; Ren, G.S. Naringin inhibits growth potential of human triple-negative breast cancer cells by targeting beta-catenin signaling pathway. Toxicol. Lett. 2013, 220, 219–228. [Google Scholar] [CrossRef]
- Palit, S.; Kar, S.; Sharma, G.; Das, P.K. Hesperetin Induces Apoptosis in Breast Carcinoma by Triggering Accumulation of ROS and Activation of ASK1/JNK Pathway. J. Cell. Physiol. 2015, 230, 1729–1739. [Google Scholar] [CrossRef]
- Sergeev, I.N.; Li, S.M.; Colby, J.; Ho, C.T.; Dushenkov, S. Polymethoxylated flavones induce Ca2+-mediated apoptosis in breast cancer cells. Life Sci. 2006, 80, 245–253. [Google Scholar] [CrossRef]
- Sergeev, I.N.; Ho, C.T.; Li, S.M.; Colby, J.; Dushenkov, S. Apoptosis-inducing activity of hydroxylated polymethoxyflavones and polymethoxyflavones from orange peel in human breast cancer cells. Mol. Nutr. Food Res. 2007, 51, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Pick, A.; Muller, H.; Mayer, R.; Haenisch, B.; Pajeva, I.K.; Weigt, M.; Bonisch, H.; Muller, C.E.; Wiese, M. Structure-activity relationships of flavonoids as inhibitors of breast cancer resistance protein (BCRP). Bioorg. Med. Chem. 2011, 19, 2090–2102. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.J.; Dong, P.; Guan, H.S.; Li, S.M.; Ho, C.T.; Pan, M.H.; McClements, D.J.; Xiao, H. Inhibitory effects of 5-hydroxy polymethoxyflavones on colon cancer cells. Mol. Nutr. Food Res. 2010, 54, S244–S252. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Murakami, A.; Ohigashi, H. Nobiletin, a citrus flavonoid, down-regulates matrix metalloproteinase-7 (matrilysin) expression in HT-29 human colorectal cancer cells. Biosci. Biotechnol. Biochem. 2005, 69, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Koike, L.; Miyata, Y.; Hirata, M.; Mimaki, Y.; Sashida, Y.; Yano, M.; Ito, A. Inhibition of activator protein-1 binding activity and phosphatidylinositol 3-kinase pathway by nobiletin, a polymethoxy flavonoid, results in augmentation of tissue inhibitor of metalloproteinases-1 production and suppression of production of matrix metalloproteinases-1 and-9 in human fibrosarcoma HT-1080 cells. Cancer Res. 2002, 62, 1025–1029. [Google Scholar]
- Miyamoto, S.; Yasui, Y.; Tanaka, T.; Ohigashi, H.; Murakami, A. Suppressive effects of nobiletin on hyperleptinemia and colitis-related colon carcinogenesis in male ICR mice. Carcinogenesis 2008, 29, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Song, M.Y.; Wang, M.Q.; Zheng, J.K.; Gao, Z.L.; Xu, F.; Zhang, G.D.; Xiao, H. Chemopreventive effects of nobiletin and its colonic metabolites on colon carcinogenesis. Mol. Nutr. Food Res. 2015, 59, 2383–2394. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.H.; Chen, W.J.; Lin-Shiau, S.Y.; Ho, C.T.; Lin, J.K. Tangeretin induces cell-cycle G(1) arrest through inhibiting cyclin-dependent kinases 2 and 4 activities as well as elevating Cdk inhibitors p21 and p27 in human colorectal carcinoma cells. Carcinogenesis 2002, 23, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kim, M.J.; Ha, E.; Chung, J.H. Apoptotic effect of hesperidin through caspase3 activation in human colon cancer cells, SNU-C4. Phytomedicine 2008, 15, 147–151. [Google Scholar] [CrossRef]
- Leonardi, T.; Vanamala, J.; Taddeo, S.S.; Davidson, L.A.; Murphy, M.E.; Patil, B.S.; Wang, N.Y.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R.; et al. Apigenin and naringenin suppress colon carcinogenesis through the aberrant crypt stage in azoxymethane-treated rats. Exp. Biol. Med. 2010, 235, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Aranganathan, S.; Selvam, J.P.; Nalini, N. Effect of hesperetin, a citrus flavonoid, on bacterial enzymes and carcinogen-induced aberrant crypt foci in colon cancer rats: A dose-dependent study. J. Pharm. Pharmacol. 2008, 60, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, A.; Otani, Y.; Kubota, T.; Wada, N.; Furukawa, T.; Kumai, K.; Kameyama, K.; Okada, Y.; Fujii, M.; Yano, M.; et al. The citrus flavonoid, nobiletin, inhibits peritoneal dissemination of human gastric carcinoma in SCID mice. Jpn. J. Cancer Res. 2001, 92, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Yoshimizu, N.; Otani, Y.; Saikawa, Y.; Kubota, T.; Yoshida, M.; Furukawa, T.; Kumai, K.; Kameyama, K.; Fujii, M.; Yano, M.; et al. Anti-tumour effects of nobiletin, a citrus flavonoid, on gastric cancer include: Antiproliferative effects, induction of apoptosis and cell cycle deregulation. Aliment. Pharmacol. Ther. 2004, 20, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Ekambaram, G.; Rajendran, P.; Magesh, V.; Sakthisekaran, D. Naringenin reduces tumor size and weight lost in N-methyl-N ‘-nitro-N-nitrosoguanidine-induced gastric carcinogenesis in rats. Nutr. Res. 2008, 28, 106–112. [Google Scholar] [CrossRef]
- Bao, L.; Liu, F.; Guo, H.B.; Li, Y.; Tan, B.B.; Zhang, W.X.; Peng, Y.H. Naringenin inhibits proliferation, migration, and invasion as well as induces apoptosis of gastric cancer SGC7901 cell line by downregulation of AKT pathway. Tumor Biol. 2016, 37, 11365–11374. [Google Scholar] [CrossRef]
- Park, H.J.; Choi, Y.J.; Lee, J.H.; Nam, M.J. Naringenin causes ASK1-induced apoptosis via reactive oxygen species in human pancreatic cancer cells. Food Chem. Toxicol. 2017, 99, 1–8. [Google Scholar] [CrossRef]
- Kohno, H.; Taima, M.; Sumida, T.; Azuma, Y.; Ogawa, H.; Tanaka, T. Inhibitory effect of mandarin juice rich in beta-cryptoxanthin and hesperidin on 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced pulmonary tumorigenesis in mice. Cancer Lett. 2001, 174, 141–150. [Google Scholar] [CrossRef]
- Chen, K.H.; Weng, M.S.; Lin, J.K. Tangeretin suppresses IL-1 beta-induced cyclooxygenase (COX)-2 expression through inhibition of p38 MAPK, JNK, and AKT activation in human lung carcinoma cells. Biochem. Pharmacol. 2007, 73, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Da, C.L.; Liu, Y.T.; Zhan, Y.Y.; Liu, K.; Wang, R.Z. Nobiletin inhibits epithelial-mesenchymal transition of human non-small cell lung cancer cells by antagonizing the TGF-beta 1/Smad3 signaling pathway. Oncol. Rep. 2016, 35, 2767–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensinphon, N.; Qiu, P.J.; Dong, P.; Zheng, J.K.; Ngauv, P.; Cao, Y.; Li, S.M.; Ho, C.T.; Xiao, H. 5-Demethyltangeretin inhibits human nonsmall cell lung cancer cell growth by inducing G2/M cell cycle arrest and apoptosis. Mol. Nutr. Food Res. 2013, 57, 2103–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, Y.C.; Hsieh, Y.S.; Kuo, W.H.; Chiou, H.L.; Yang, S.F.; Chiang, W.L.; Chu, S.C. The tumor-growth inhibitory activity of flavanone and 2 ‘-OH flavanone in vitro and in vivo through induction of cell cycle arrest and suppression of cyclins and CDKs. J. Biomed. Sci. 2007, 14, 107–119. [Google Scholar] [CrossRef]
- Cincin, Z.B.; Unlu, M.; Kiran, B.; Bireller, E.S.; Baran, Y.; Cakmakoglu, B. Anti-proliferative, apoptotic and signal transduction effects of hesperidin in non-small cell lung cancer cells. Cell. Oncol. 2015, 38, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Arul, D.; Subramanian, P. Naringenin (Citrus Flavonone) Induces Growth Inhibition, Cell Cycle Arrest and Apoptosis in Human Hepatocellular Carcinoma Cells. Pathol. Oncol. Res. 2013, 19, 763–770. [Google Scholar] [CrossRef]
- Banjerdpongchai, R.; Wudtiwai, B.; Khaw-on, P.; Rachakhom, W.; Duangnil, N.; Kongtawelert, P. Hesperidin from Citrus seed induces human hepatocellular carcinoma HepG2 cell apoptosis via both mitochondrial and death receptor pathways. Tumor Biol. 2016, 37, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.; Park, S.; Bazer, F.W.; Song, G. Naringenin-Induced Apoptotic Cell Death in Prostate Cancer Cells Is Mediated via the PI3K/AKT and MAPK Signaling Pathways. J. Cell. Biochem. 2017, 118, 1118–1131. [Google Scholar] [CrossRef]
- Gao, K.; Henning, S.M.; Niu, Y.T.; Youssefian, A.A.; Seeram, N.P.; Xu, A.L.; Heber, D. The citrus flavonoid naringenin stimulates DNA repair in prostate cancer cells. J. Nutr. Biochem. 2006, 17, 89–95. [Google Scholar] [CrossRef]
- Alshatwi, A.A.; Ramesh, E.; Periasamy, V.S.; Subash-Babu, P. The apoptotic effect of hesperetin on human cervical cancer cells is mediated through cell cycle arrest, death receptor, and mitochondrial pathways. Fundam. Clin. Pharmacol. 2013, 27, 581–592. [Google Scholar] [CrossRef]
- Zeng, L.; Zhen, Y.L.; Chen, Y.M.; Zou, L.; Zhang, Y.; Hu, F.; Feng, J.Q.; Shen, J.H.; Wei, B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-kappa B/COX-2-caspase-1 pathway in HeLa cervical cancer cells. Int. J. Oncol. 2014, 45, 1929–1936. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.; Chen, A.Y.; Huang, H.Z.; Ye, X.Q.; Rollyson, W.D.; Perry, H.E.; Brown, K.C.; Rojanasakul, Y.; Rankin, G.O.; Dasgupta, P.; et al. The flavonoid nobiletin inhibits tumor growth and angiogenesis of ovarian cancers via the Akt pathway. Int. J. Oncol. 2015, 46, 2629–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafa, E.S.A.; Zhu, Q.Z.; Barakat, B.M.; Wani, G.; Zhao, Q.; El-Mahdy, M.A.; Wani, A.A. Tangeretin Sensitizes Cisplatin-Resistant Human Ovarian Cancer Cells through Downregulation of Phosphoinositide 3-Kinase/Akt Signaling Pathway. Cancer Res. 2009, 69, 8910–8917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandaswami, C.; Perkins, E.; Soloniuk, D.S.; Drzewiecki, G.; Middleton, E. Antiproliferative effects of citrus flavonoids on a human squamous-cell carcinoma invitro. Cancer Letters 1991, 56, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Ahamad, M.S.; Siddiqui, S.; Jafri, A.; Ahmad, S.; Afzal, M.; Arshad, M. Induction of Apoptosis and Antiproliferative Activity of Naringenin in Human Epidermoid Carcinoma Cell through ROS Generation and Cell Cycle Arrest. PLoS ONE 2014, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Spencer, J.P.E. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104, S40–S47. [Google Scholar] [CrossRef] [Green Version]
- Rezai-Zadeh, K.; Shytle, R.D.; Bai, Y.; Tian, J.; Hou, H.Y.; Mori, T.; Zeng, J.; Obregon, D.; Town, T.; Tan, J. Flavonoid-mediated presenilin-1 phosphorylation reduces Alzheimer’s disease beta-amyloid production. J. Cell. Mol. Med. 2009, 13, 574–588. [Google Scholar] [CrossRef]
- Hwang, S.L.; Yen, G.C. Neuroprotective effects of the citrus flavanones against H2O2-induced cytotoxicity in PC12 cells. J. Agric. Food Chem. 2008, 56, 859–864. [Google Scholar] [CrossRef]
- Johnson, J.; Maher, P.; Hanneken, A. The Flavonoid, Eriodictyol, Induces Long-term Protection in ARPE-19 Cells through Its Effects on Nrf2 Activation and Phase 2 Gene Expression. Invest. Ophthalmol. Vis. Sci. 2009, 50, 2398–2406. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Miyazaki, K.; Sakai, S.; Yawo, H.; Nakata, N.; Moriguchi, S.; Fukunaga, K.; Yokosuka, A.; Sashida, Y.; Mimaki, Y.; et al. Nobiletin, a citrus flavonoid with neurotrophic action, augments protein kinase A-mediated phosphorylation of the AMPA receptor subunit, GluR1, and the postsynaptic receptor response to glutamate in murine hippocampus. Eur. J. Pharmacol. 2008, 578, 194–200. [Google Scholar] [CrossRef]
- Nakajima, A.; Yamakuni, T.; Haraguchi, M.; Omae, N.; Song, S.Y.; Kato, C.; Nakagawasai, O.; Tadano, T.; Yokosuka, A.; Mimaki, Y.; et al. Nobiletin, a citrus flavonoid that improves memory impairment, rescues bulbectomy-induced cholinergic neurodegeneration in mice. J. Pharmacol. Sci. 2007, 105, 122–126. [Google Scholar] [CrossRef] [Green Version]
- Onozuka, H.; Nakajima, A.; Matsuzaki, K.; Shin, R.W.; Ogino, K.; Saigusa, D.; Tetsu, N.; Yokosuka, A.; Sashida, Y.; Mimaki, Y.; et al. Nobiletin, a citrus flavonoid, improves memory impairment and A beta pathology in a transgenic mouse model of Alzheimer’s Disease. J. Pharmacol. Exp. Ther. 2008, 326, 739–744. [Google Scholar] [CrossRef]
- Nakajima, A.; Aoyama, Y.; Shin, E.J.; Nam, Y.; Kim, H.C.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; Yokoi, T.; Ohizumi, Y.; et al. Nobiletin, a citrus flavonoid, improves cognitive impairment and reduces soluble A beta levels in a triple transgenic mouse model of Alzheimer’s disease (3XTg-AD). Behav. Brain Res. 2015, 289, 69–77. [Google Scholar] [CrossRef]
- Nakajima, A.; Aoyama, Y.; Nguyen, T.T.L.; Shin, E.J.; Kim, H.C.; Yamada, S.; Nakai, T.; Nagai, T.; Yokosuka, A.; Mimakic, Y.; et al. Nobiletin, a citrus flavonoid, ameliorates cognitive impairment, oxidative burden, and hyperphosphorylation of tau in senescence-accelerated mouse. Behav. Brain Res. 2013, 250, 351–360. [Google Scholar] [CrossRef]
- Yabuki, Y.; Ohizumi, Y.; Yokosuka, A.; Mimaki, Y.; Fukunaga, K. Nobiletin treatment improves motor and cognitive deficits seen in mptp-induced parkinson model mice. Neuroscience 2014, 259, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Yamakuni, T.; Matsuzaki, K.; Nakata, N.; Onozuka, H.; Yokosuka, A.; Sashida, Y.; Mimaki, Y.; Ohizumi, Y. Nobiletin, a citrus flavonoid, reverses learning impairment associated with N-methyl-D-aspartate receptor antagonism by activation of extracellular signal-regulated kinase signaling. J. Pharmacol. Exp. Ther. 2007, 321, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.D.; Jeong, K.H.; Jung, U.J.; Kim, S.R. Naringin treatment induces neuroprotective effects in a mouse model of Parkinson’s disease in vivo, but not enough to restore the lesioned dopaminergic system. J. Nutr. Biochem. 2016, 28, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Leem, E.; Nam, J.H.; Jeon, M.T.; Shin, W.H.; Won, S.Y.; Park, S.J.; Choi, M.S.; Jin, B.K.; Jung, U.J.; Kim, S.R. Naringin protects the nigrostriatal dopaminergic projection through induction of GDNF in a neurotoxin model of Parkinson’s disease. J. Nutr. Biochem. 2014, 25, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dogra, S.; Prakash, A. Protective Effect of Naringin, a Citrus Flavonoid, Against Colchicine-Induced Cognitive Dysfunction and Oxidative Damage in Rats. J. Med. Food 2010, 13, 976–984. [Google Scholar] [CrossRef]
- Vafeiadou, K.; Vauzour, D.; Lee, H.Y.; Rodriguez-Mateos, A.; Williams, R.J.; Spencer, J.P.E. The citrus flavanone naringenin inhibits inflammatory signalling in glial cells and protects against neuroinflammatory injury. Arch. Biochem. Biophys. 2009, 484, 100–109. [Google Scholar] [CrossRef]
- Wu, L.H.; Lin, C.; Lin, H.Y.; Liu, Y.S.; Wu, C.Y.J.; Tsai, C.F.; Chang, P.C.; Yeh, W.L.; Lu, D.Y. Naringenin Suppresses Neuroinflammatory Responses Through Inducing Suppressor of Cytokine Signaling 3 Expression. Mol. Neurobiol. 2016, 53, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Vaibhav, K.; Ahmed, M.E.; Khan, A.; Tabassum, R.; Islam, F.; Safhi, M.M.; Islam, F. Effect of hesperidin on neurobehavioral, neuroinflammation, oxidative stress and lipid alteration in intracerebroventricular streptozotocin induced cognitive impairment in mice. J. Neurol. Sci. 2015, 348, 51–59. [Google Scholar] [CrossRef]
- Li, C.Y.; Zug, C.; Qu, H.C.; Schluesener, H.; Zhang, Z.Y. Hesperidin ameliorates behavioral impairments and neuropathology of transgenic APP/PS1 mice. Behav. Brain Res. 2015, 281, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a Citrus Flavonoid, Attenuates LPS-Induced Neuroinflammation, Apoptosis and Memory Impairments by Modulating TLR4/NF-kappa B Signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef] [Green Version]
- Kheradmand, E.; Moghaddam, A.H.; Zare, M. Neuroprotective effect of hesperetin and nano-hesperetin on recognition memory impairment and the elevated oxygen stress in rat model of Alzheimer’s disease. Biomed. Pharmacother. 2018, 97, 1096–1101. [Google Scholar] [CrossRef]
- Datla, K.P.; Christidou, M.; Widmer, W.W.; Rooprai, H.K.; Dexter, D.T. Tissue distribution and neuroprotective effects of citrus flavonoid tangeretin in a rat model of Parkinson’s disease. Neuroreport 2001, 12, 3871–3875. [Google Scholar] [CrossRef]
- Shu, Z.P.; Yang, B.Y.; Zhao, H.; Xu, B.Q.; Jiao, W.J.; Wang, Q.H.; Wang, Z.B.; Kuang, H.X. Tangeretin exerts anti-neuroinflammatory effects via NF-kappa B modulation in lipopolysaccharide-stimulated microglial cells. Int. Immunopharmacol. 2014, 19, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Abe, K.; Gotoh, M.; Oka, K. Citrus flavone tangeretin inhibits leukemic hl-60 cell-growth partially through induction of apoptosis with less cytotoxicity on normal lymphocytes. Br. J. Cancer 1995, 72, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Toth, P.P.; Patti, A.M.; Nikolic, D.; Giglio, R.V.; Castellino, G.; Biancucci, T.; Geraci, F.; David, S.; Montalto, G.; Rizvi, A.; et al. Bergamot Reduces Plasma Lipids, Atherogenic Small Dense LDL, and Subclinical Atherosclerosis in Subjects with Moderate Hypercholesterolemia: A 6 Months Prospective Study. Front. Pharmacol. 2016, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Hu, Y.; Zhang, W.N.; Zhao, X.Y.; Chen, Y.H.; Sun, C.D.; Li, X.; Chen, K.S. Hypoglycemic and hypolipidemic effects of neohesperidin derived from Citrus aurantium L. in diabetic KK-A(y) mice. Food Funct. 2015, 6, 878–886. [Google Scholar] [CrossRef]
- Whitman, S.C.; Kurowska, E.M.; Manthey, J.A.; Daugherty, A. Nobiletin, a citrus flavonoid isolated from tangerines, selectively inhibits class A scavenger receptor-mediated metabolism of acetylated LDL by mouse macrophages. Atherosclerosis 2005, 178, 25–32. [Google Scholar] [CrossRef]
- Mollace, V.; Sacco, I.; Janda, E.; Malara, C.; Ventrice, D.; Colica, C.; Visalli, V.; Muscoli, S.; Ragusa, S.; Muscoli, C.; et al. Hypolipemic and hypoglycaemic activity of bergamot polyphenols: From animal models to human studies. Fitoterapia 2011, 82, 309–316. [Google Scholar] [CrossRef] [PubMed]
- He, B.K.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.Y.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvihill, E.E.; Assini, J.M.; Sutherland, B.G.; DiMattia, A.S.; Khami, M.; Koppes, J.B.; Sawyez, C.G.; Whitman, S.C.; Huff, M.W. Naringenin Decreases Progression of Atherosclerosis by Improving Dyslipidemia in High-Fat-Fed Low-Density Lipoprotein Receptor-Null Mice. Thromb. Vasc. Biol. 2010, 30, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Mulvihill, E.E.; Allister, E.M.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Edwards, J.Y.; Markle, J.M.; Hegele, R.A.; Huff, M.W. Naringenin Prevents Dyslipidemia, Apolipoprotein B Overproduction, and Hyperinsulinemia in LDL Receptor-Null Mice with Diet-Induced Insulin Resistance. Diabetes 2009, 58, 2198–2210. [Google Scholar] [CrossRef] [Green Version]
- Allister, E.M.; Borradaile, N.M.; Edwards, J.Y.; Huff, M.W. Inhibition of microsomal triglyceride transfer protein expression and apolipoprotein B100 secretion by the citrus flavonoid naringenin and by insulin involves activation of the mitogen-activated protein kinase pathway in hepatocytes. Diabetes 2005, 54, 1676–1683. [Google Scholar] [CrossRef] [Green Version]
- Assini, J.M.; Mulvihill, E.E.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Felder, S.L.; Chhoker, S.; Edwards, J.Y.; Gros, R.; Huff, M.W. Naringenin prevents cholesterol-induced systemic inflammation, metabolic dysregulation, and atherosclerosis in Ldlr(-/-) mice. J. Lipid Res. 2013, 54, 711–724. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, L.J.; Borradaile, N.M.; de Dreu, L.E.; Huff, M.W. Secretion of hepatocyte apoB is inhibited by the flavonoids, naringenin and hesperetin, via reduced activity and expression of ACAT2 and MTP. J. Lipid Res. 2001, 42, 725–734. [Google Scholar] [CrossRef]
- Priscilla, D.H.; Jayakumar, M.; Thirumurugan, K. Flavanone naringenin: An effective antihyperglycemic and antihyperlipidemic nutraceutical agent on high fat diet fed streptozotocin induced type 2 diabetic rats. J. Funct. Food. 2015, 14, 363–373. [Google Scholar] [CrossRef]
- Borradaile, N.M.; de Dreu, L.E.; Huff, M.W. Inhibition of net HepG2 cell apolipoprotein B secretion by the citrus flavonoid naringenin involves activation of phosphatidylinositol 3-kinase, independent of insulin receptor substrate-1 phosphorylation. Diabetes 2003, 52, 2554–2561. [Google Scholar] [CrossRef] [Green Version]
- Rahigude, A.; Bhutada, P.; Kaulaskar, S.; Aswar, M.; Otari, K. Participation of antioxidant and cholinergic system in protective effect of naringenin against type-2 diabetes-induced memory dysfunction in rats. Neuroscience 2012, 226, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Nyane, N.A.; Tlaila, T.B.; Malefane, T.G.; Ndwandwe, D.E.; Owira, P.M.O. Metformin-like antidiabetic, cardio-protective and non-glycemic effects of naringenin: Molecular and pharmacological insights. Eur. J. Pharmacol. 2017, 803, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Chanet, A.; Milenkovic, D.; Deval, C.; Potier, M.; Constans, J.; Mazur, A.; Bennetau-Pelissero, C.; Morand, C.; Berard, A.M. Naringin, the major grapefruit flavonoid, specifically affects atherosclerosis development in diet-induced hypercholesterolemia in mice. J. Nutr. Biochem. 2012, 23, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Jeong, T.S.; Choi, Y.K.; Hyun, B.H.; Oh, G.T.; Kim, E.H.; Kim, J.R.; Han, J.I.; Bok, S.H. Anti-atherogenic effect of citrus flavonoids, naringin and naringenin, associated with hepatic ACAT and aortic VCAM-1 and MCP-1 in high cholesterol-fed rabbits. Biochem. Biophys. Res. Commun. 2001, 284, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kauter, K.; Brown, L. Naringin Improves Diet-Induced Cardiovascular Dysfunction and Obesity in High Carbohydrate. High Fat Diet Fed Rats 2013, 5, 637–650. [Google Scholar]
- Pang, W.Y.; Wang, X.L.; Mok, S.K.; Lai, W.P.; Chow, H.K.; Leung, P.C.; Yao, X.S.; Wong, M.S. Naringin improves bone properties in ovariectomized mice and exerts oestrogen-like activities in rat osteoblast-like (UMR-106) cells. Br. J. Pharmacol. 2010, 159, 1693–1703. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, S.; Katsumata, S.I.; Suzuki, K.; Ishimi, Y.; Wu, J.; Uehara, M. Dietary Hesperidin Exerts Hypoglycemic and Hypolipidemic Effects in Streptozotocin-Induced Marginal Type 1 Diabetic Rats. J. Clin. Biochem. Nutr. 2010, 46, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurowska, E.A.; Manthey, J.A. Hypolipidemic effects and absorption of citrus polymethoxylated flavones in hamsters with diet-induced hypercholesterolemia. J. Agric. Food Chem. 2004, 52, 2879–2886. [Google Scholar] [CrossRef]
- Rizza, S.; Muniyappa, R.; Iantorno, M.; Kim, J.A.; Chen, H.; Pullikotil, P.; Senese, N.; Tesauro, M.; Lauro, D.; Cardillo, C.; et al. Citrus Polyphenol Hesperidin Stimulates Production of Nitric Oxide in Endothelial Cells while Improving Endothelial Function and Reducing Inflammatory Markers in Patients with Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E782–E792. [Google Scholar] [CrossRef]
- Revathy, J.; Srinivasan, S.; Abdullah, S.H.S.; Muruganathan, U. Antihyperglycemic effect of hesperetin, a citrus flavonoid, extenuates hyperglycemia and exploring the potential role in antioxidant and antihyperlipidemic in streptozotocin-induced diabetic rats. Biomed. Pharm. 2018, 97, 98–106. [Google Scholar] [CrossRef]
- Samie, A.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Hesperetin, a citrus flavonoid, attenuates testicular damage in diabetic rats via inhibition of oxidative stress, inflammation, and apoptosis. Life Sci. 2018, 210, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Pari, L. Ameliorative effect of diosmin, a citrus flavonoid against streptozotocin-nicotinamide generated oxidative stress induced diabetic rats. Chem. Biol. Interact. 2012, 195, 43–51. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Number of Publication | % of Total Publication | Journal | |||
|---|---|---|---|---|---|---|
| Name | Country/Region | Impact Factor (5 Years) | H-Index (2020–2021) | |||
| 1 | 188 | 5.866 | Journal of Agricultural and Food Chemistry | United States | 5.105 | 297.0 |
| 2 | 156 | 4.867 | Food Chemistry | United Kingdom | 7.341 | 262.0 |
| 3 | 75 | 2.34 | Molecules | Switzerland | 4.189 | 149.0 |
| 4 | 47 | 1.466 | Food & Function | United Kingdom | 6.317 | 89.0 |
| 5 | 40 | 1.248 | Journal of Functional Foods | United Kingdom | 4.432 | 84.0 |
| 6 | 40 | 1.248 | Journal of The Science of Food and Agriculture | United Kingdom | 3.547 | 142.0 |
| 7 | 38 | 1.186 | LWT Food Science and Technology | United States | 4.991 | 133.0 |
| 8 | 29 | 0.905 | Nutrients | Switzerland | 5.644 | 115.0 |
| 9 | 28 | 0.874 | Scientia Horticulturae | Netherlands | 3.612 | 112.0 |
| 10 | 27 | 0.842 | Molecular Nutrition Food Research | Germany | 6.575 | 141.0 |
| 11 | 26 | 0.811 | British Journal of Nutrition | United Kingdom | 3.59 | 188.0 |
| 12 | 25 | 0.78 | Food Research International | United Kingdom | 6.355 | 163.0 |
| 13 | 25 | 0.78 | Food Science and Biotechnology | South Korea | 2.468 | 38.0 |
| 14 | 25 | 0.78 | International Journal of Molecular Sciences | Switzerland | 5.708 | 162.0 |
| 15 | 24 | 0.749 | European Food Research and Technology | Germany | 2.898 | 102.0 |
| 16 | 24 | 0.749 | Journal of Food Processing and Preservation | United States | 2.1 | 48.0 |
| 17 | 24 | 0.749 | Journal of Pharmaceutical and Biomedical Analysis | Netherlands | 3.663 | 127.0 |
| 18 | 23 | 0.718 | Food and Chemical Toxicology | United Kingdom | 4.449 | 172.0 |
| 19 | 22 | 0.686 | Bioscience Biotechnology and Biochemistry | United Kingdom | 1.913 | 116.0 |
| 20 | 22 | 0.686 | European Journal of Pharmacology | Netherlands | 4.064 | 180.0 |
| 21 | 22 | 0.686 | Journal of Chromatography A | Netherlands | 4.321 | 229.0 |
| 22 | 22 | 0.686 | Journal of Separation Science | Germany | 3.201 | 102.0 |
| 23 | 22 | 0.686 | Life Sciences | United States | 4.615 | 164.0 |
| 24 | 22 | 0.686 | Phytotherapy Research | United Kingdom | 5.374 | 129.0 |
| 25 | 21 | 0.655 | Antioxidants | Switzerland | 6.084 | 46.0 |
| Rank | Cited Number | Journal | |||
|---|---|---|---|---|---|
| Name | Country/Region | Impact Factor (2020–2021) | H-Index (202–2021) | ||
| 1 | 2157 | Journal of Agricultural and Food Chemistry | United States | 5.105 | 297.0 |
| 2 | 1454 | Food Chemistry | United Kingdom | 7.341 | 262.0 |
| 3 | 703 | Molecules | Switzerland | 4.189 | 149.0 |
| 4 | 680 | Journal of Biological Chemistry | United States | 4.562 | 513.0 |
| 5 | 611 | Life Sciences | United States | 4.615 | 164.0 |
| 6 | 611 | Journal of Nutrition | United States | 4.019 | 265.0 |
| 7 | 607 | PLoS ONE | United States | 3.272 | 332.0 |
| 8 | 605 | Phytochemistry | United Kingdom | 3.814 | 176.0 |
| 9 | 582 | Journal of the Science of Food and Agriculture | United Kingdom | 3.547 | 142.0 |
| 10 | 566 | Food and Chemical Toxicology | United Kingdom | 4.449 | 172.0 |
| 11 | 553 | Biochemical Pharmacology | United States | 5.494 | 198.0 |
| 12 | 550 | Free Radical Biology and Medicine | United States | 265.0 | 265.0 |
| 13 | 548 | Phytotherapy Research | United Kingdom | 5.374 | 129.0 |
| 14 | 509 | Nature | United Kingdom | 49.962 | 1226.0 |
| 15 | 507 | Proceedings of the National Academy of Sciences of the United States of America | United States | 12.779 | 805 |
| 16 | 500 | Biochemical and Biophysical Research Communications | United States | 3.575 | 271 |
| 17 | 494 | Bioscience, Biotechnology, and Biochemistry | United Kingdom | 2.337 | 123 |
| 18 | 481 | American Journal of Clinical Nutrition | United States | 7.045 | 351 |
| 19 | 463 | Food Research International | United Kingdom | 6.475 | 177 |
| 20 | 458 | Journal of Chromatography A | Netherlands | 4.049 | 237 |
| 21 | 444 | Journal of Food Science | United States | 2.470 | 160 |
| 22 | 428 | Analytical Biochemistry | United States | 3.365 | 195 |
| 23 | 425 | Journal of Pharmaceutical and Biomedical Analysis | Netherlands | 3.571 | 133 |
| 24 | 385 | Journal of Ethnopharmacology | Ireland | 3.690 | 205 |
| 25 | 383 | Biological and Pharmaceutical Bulletin | Japan | 2.233 | 122 |
| Rank | Times Cited | Journal | References | |||
|---|---|---|---|---|---|---|
| Year | Name | Country | Impact Factor (2021) | |||
| 1 | 457 | 2000 | Food Chemistry | United Kingdom | 9.231 | [13] |
| 2 | 441 | 2010 | Food Chemistry | United Kingdom | 9.231 | [14] |
| 3 | 431 | 2007 | Journal of Nutrition | United States | 4.687 | [15] |
| 4 | 410 | 2006 | Bioscience, Biotechnology, and Biochemistry | United Kingdom | 2.337 | [16] |
| 5 | 390 | 2001 | Journal of Nutrition | United States | 4.687 | [17] |
| 6 | 382 | 1996 | Nutrition and Cancer | United States | 2.816 | [18] |
| 7 | 378 | 2003 | Journal of Neurochemistry | United Kingdom | 5.546 | [19] |
| 8 | 322 | 2009 | Pakistan Journal of Pharmaceutical Sciences | Pakistan | 0.863 | [20] |
| 9 | 306 | 2003 | European Journal of Clinical Nutrition | United Kingdom | 4.884 | [21] |
| 10 | 301 | 2007 | Food Chemistry | United Kingdom | 9.231 | [22] |
| 11 | 298 | 2001 | Food Chemistry | United Kingdom | 9.231 | [23] |
| 12 | 296 | 2005 | Journal of Agricultural and Food Chemistry | United States | 5.105 | [24] |
| 13 | 292 | 2006 | Food Chemistry | United Kingdom | 9.231 | [25] |
| 14 | 284 | 1999 | Journal of Agricultural and Food Chemistry | United States | 5.105 | [26] |
| 15 | 280 | 2013 | Green Chemistry | United Kingdom | 11.034 | [27] |
| Rank | Times Cited | Journal | References | |||
|---|---|---|---|---|---|---|
| Year | Name | Country | Impact Factor (2021) | |||
| 1 | 113 | 2008 | Journal of Agricultural and Food Chemistry | United States | 5.105 | [28] |
| 2 | 106 | 2007 | Food Chemistry | United Kingdom | 9.231 | [29] |
| 3 | 96 | 2015 | Phytotherapy Research | United Kingdom | 6.388 | [30] |
| 4 | 78 | 2006 | Bioscience, Biotechnology, and Biochemistry | United Kingdom | 2.337 | [16] |
| 5 | 75 | 2014 | Advances in Nutrition | United States | 11.567 | [31] |
| 6 | 68 | 2007 | Molecules | Switzerland | 4.189 | [32] |
| 7 | 59 | 2016 | Food Chemistry | United Kingdom | 9.231 | [33] |
| 8 | 58 | 2014 | Journal of Food Composition and Analysis | United States | 4.520 | [34] |
| 9 | 49 | 2014 | Food Chemistry | United Kingdom | 9.231 | [35] |
| 10 | 46 | 2013 | Current Opinion in Lipidology | United States | 4.616 | [36] |
| 11 | 43 | 2011 | American Journal of Clinical Nutrition | United States | 8.472 | [37] |
| 12 | 42 | 2001 | Current Medicinal Chemistry | United Arab Emirates | 4.740 | [38] |
| 13 | 42 | 2003 | European Journal of Clinical Nutrition | United Kingdom | 4.884 | [21] |
| 14 | 41 | 2001 | Journal of Nutrition | United States | 4.687 | [17] |
| 15 | 38 | 2008 | Food Chemistry | United Kingdom | 9.231 | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, M.; Wei, W.; Zhang, J.; Wang, H.; Bai, Y.; Guo, D.-a. A Scientometric Study to a Critical Review on Promising Anticancer and Neuroprotective Compounds: Citrus Flavonoids. Antioxidants 2023, 12, 669. https://doi.org/10.3390/antiox12030669
Qiu M, Wei W, Zhang J, Wang H, Bai Y, Guo D-a. A Scientometric Study to a Critical Review on Promising Anticancer and Neuroprotective Compounds: Citrus Flavonoids. Antioxidants. 2023; 12(3):669. https://doi.org/10.3390/antiox12030669
Chicago/Turabian StyleQiu, Mingyang, Wenlong Wei, Jianqing Zhang, Hanze Wang, Yuxin Bai, and De-an Guo. 2023. "A Scientometric Study to a Critical Review on Promising Anticancer and Neuroprotective Compounds: Citrus Flavonoids" Antioxidants 12, no. 3: 669. https://doi.org/10.3390/antiox12030669