
Phenylethanoid Glycoside-Enriched Extract Prepared from Clerodendrum chinense Leaf Inhibits A549 Lung Cancer Cell Migration and Apoptosis Induction through Enhancing ROS Production
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Method
2.1. Materials
2.2. Plant Collection and Identification
2.3. Plant Extraction
2.4. Phytochemical Analysis of C. chinense Leaf Extract
2.5. Determination of Total Phenolics in C. chinense Leaf Extract
2.6. Determination of Total Flavonoids in C. chinense Leaf Extract
2.7. Method Validation of HPLC Analysis for the Identification and Quantification Bioactive Compounds in C. chinense Leaf Extract
2.7.1. Linearity
2.7.2. Precision
2.7.3. Accuracy
2.7.4. Limit of Detection (LOD) and Limit of Quantitation (LOQ)
2.7.5. Identification and Quantification Bioactive Compounds in C. chinense Leaf Extract
2.8. Antioxidant Assay
2.8.1. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Activity Assay
2.8.2. 2,2′-Azino-bis (3-Ethylbenzthiazoline-6-sulphonic Acid (ABTS) Radical Scavenging Activity Assay
2.8.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.9. Determination of Cytotoxicity of C. chinense Leaf Extract and Bioactive Compounds Using Sulforhodamine B (SRB) Assay
2.10. Apoptosis Assay
2.11. ROS Formation Assay
2.12. Colony Formation Assay
2.13. Cell Migration Assay
2.14. Statistical Analysis
3. Results
3.1. Yield of C. chinense Leaf Ethanolic Extract
3.2. Phytochemical Screening of C. chinense Leaf Ethanol Extract
3.3. Total Phenolic and Flavonoid Contents of C. chinense Leaf Extract
3.4. HPLC Method Validation
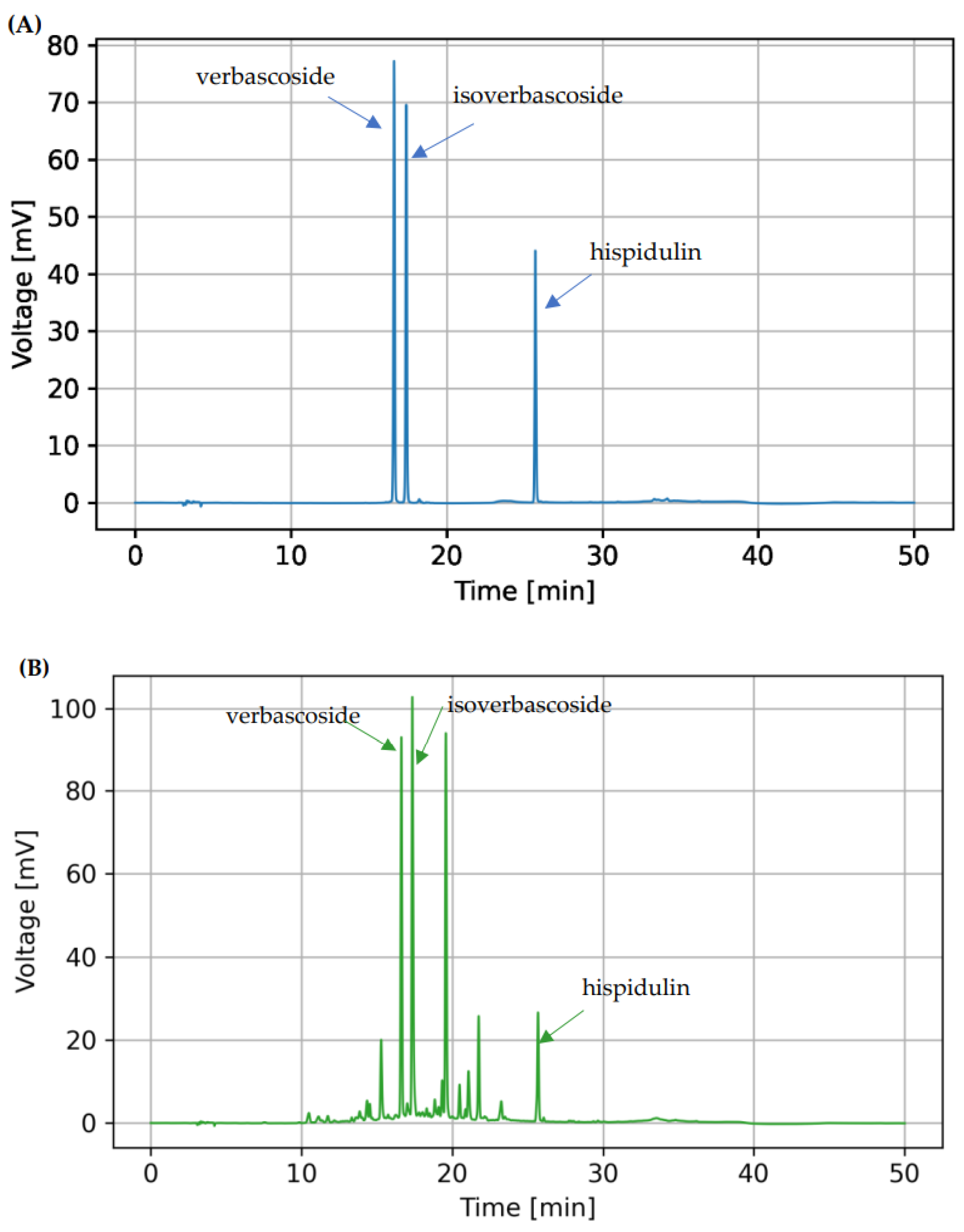
3.5. Bioactive Compounds in C. chinense Leaf Extract Determined by HPLC Analysis
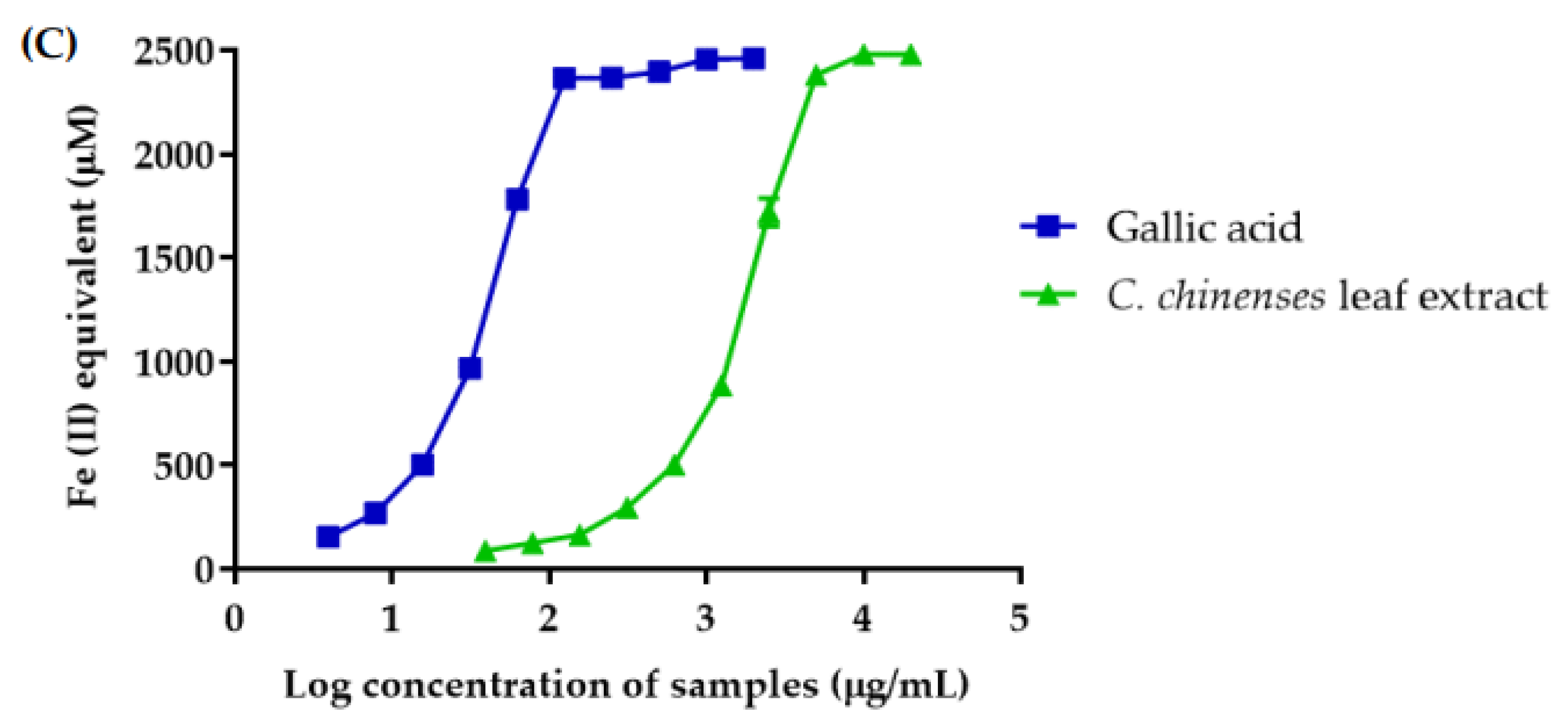
3.6. Antioxidant Activities of C. chinense Leaf Extract
3.7. Pearson Correlation of Total Phenolic and Flavonoid Contents with Antioxidant Activity of C. chinense Leaf Extract
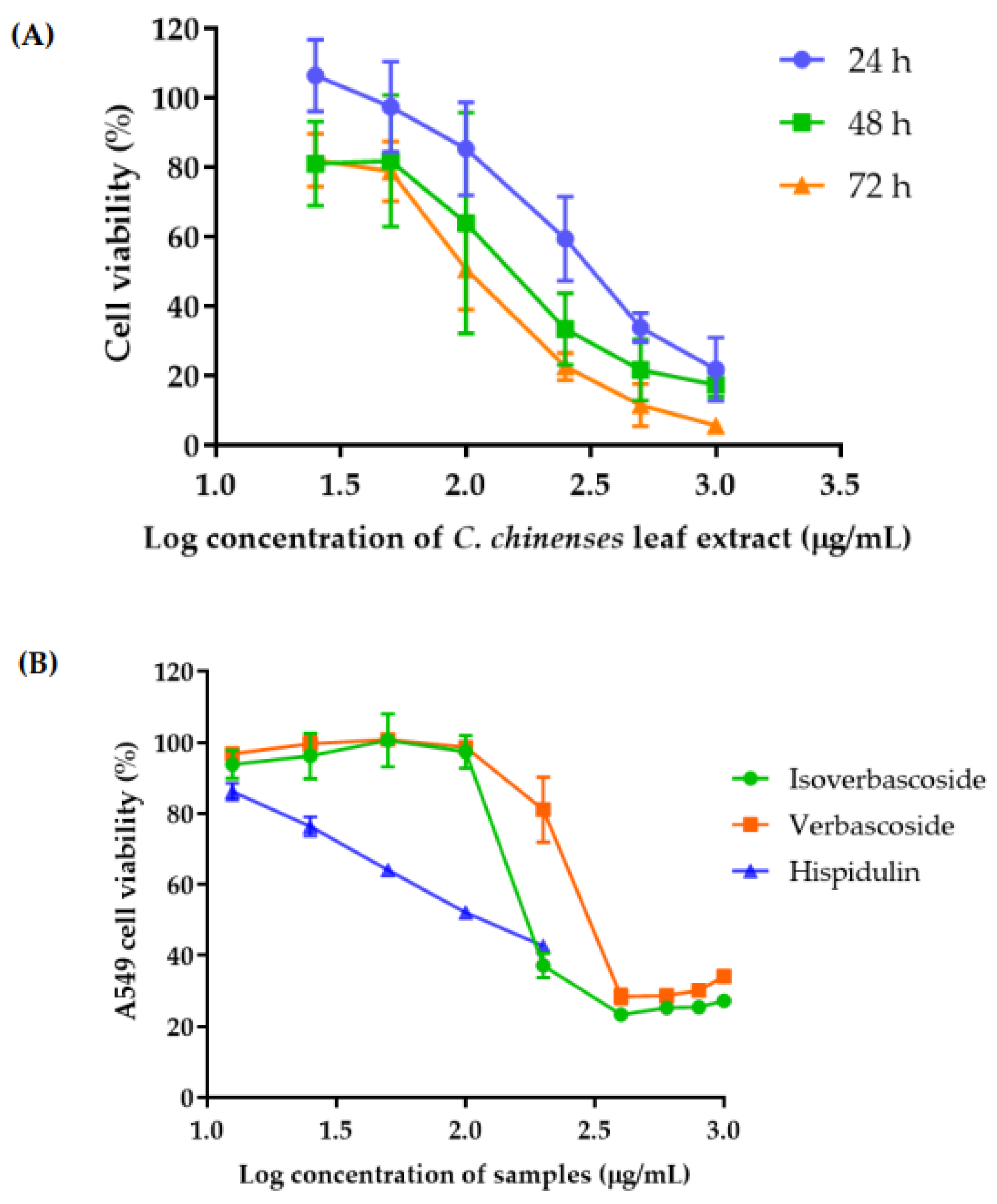
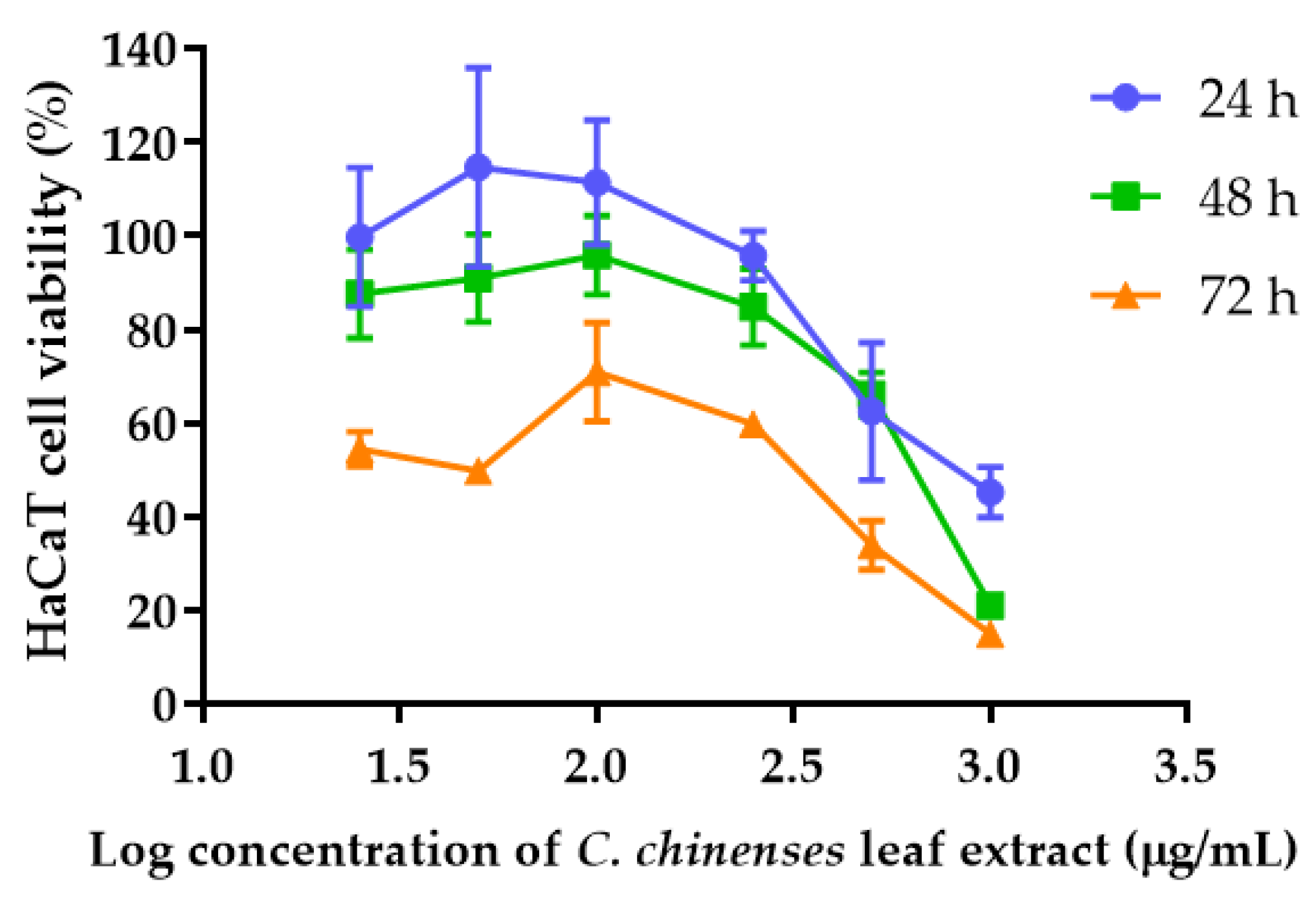
3.8. Determination of Cytotoxicity of C. chinense Leaf Extract and Bioactive Compounds Using Sulforhodamine B (SRB) Assay
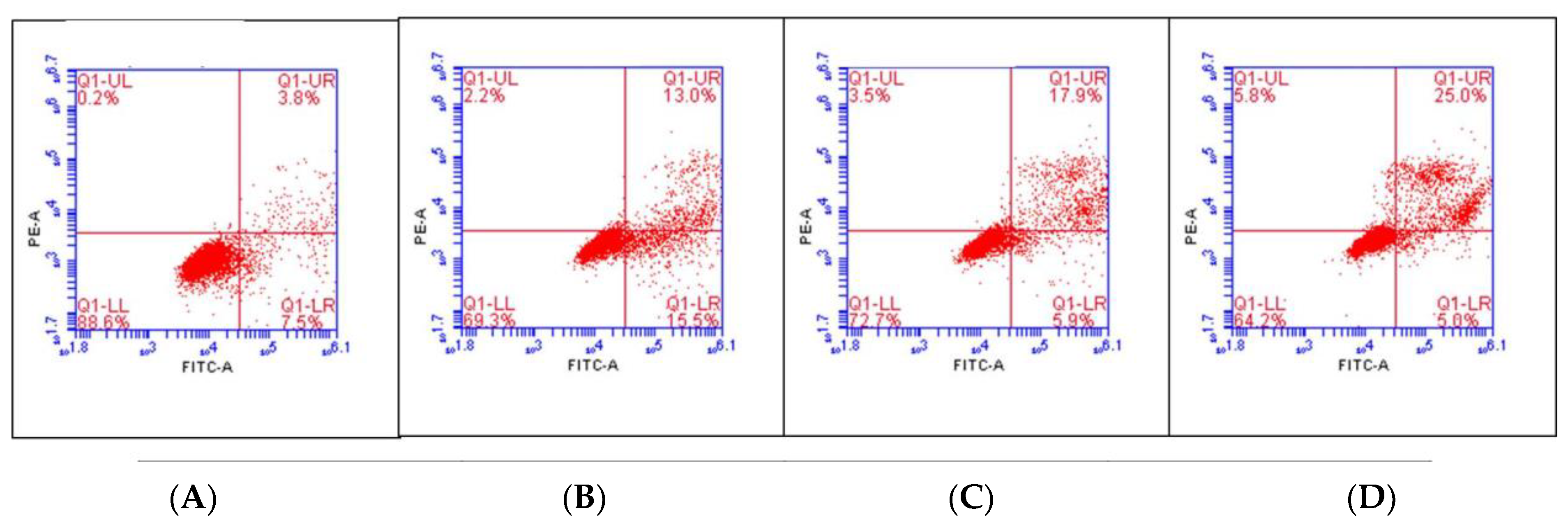
3.9. Annexin V-FITC/PI for Apoptosis Detection
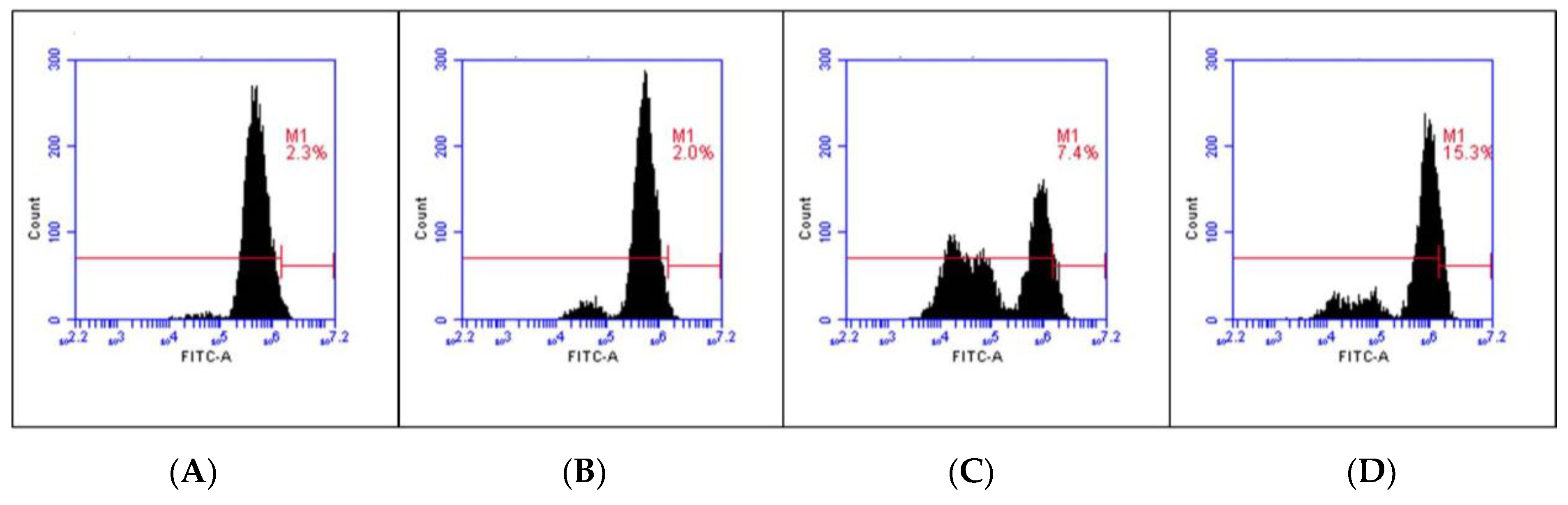
3.10. C. chinense Leaf Extract Induced ROS Generation in A549 Cells
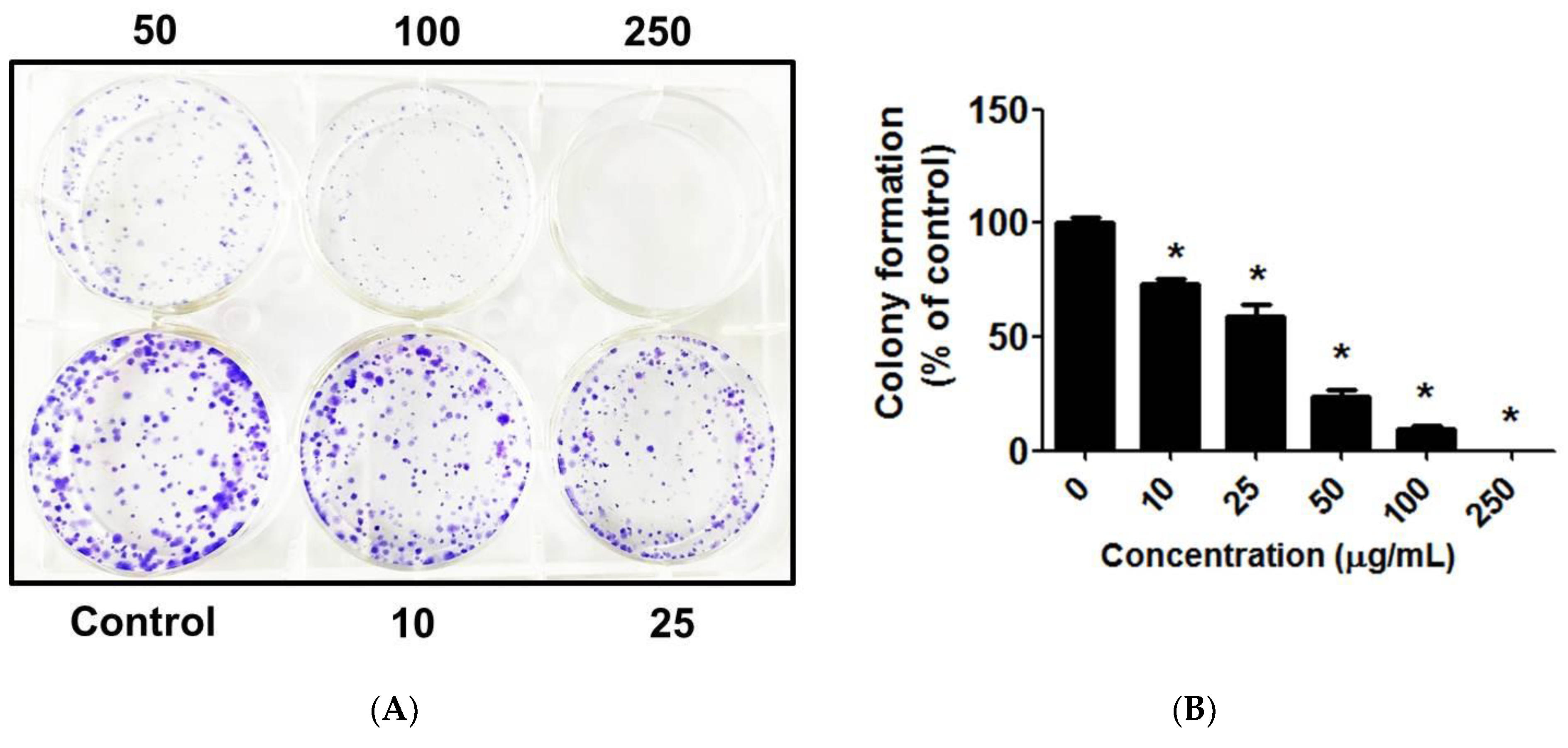
3.11. C. chinense Leaf Extract Decreased Lung Cancer Cell Colony Formation
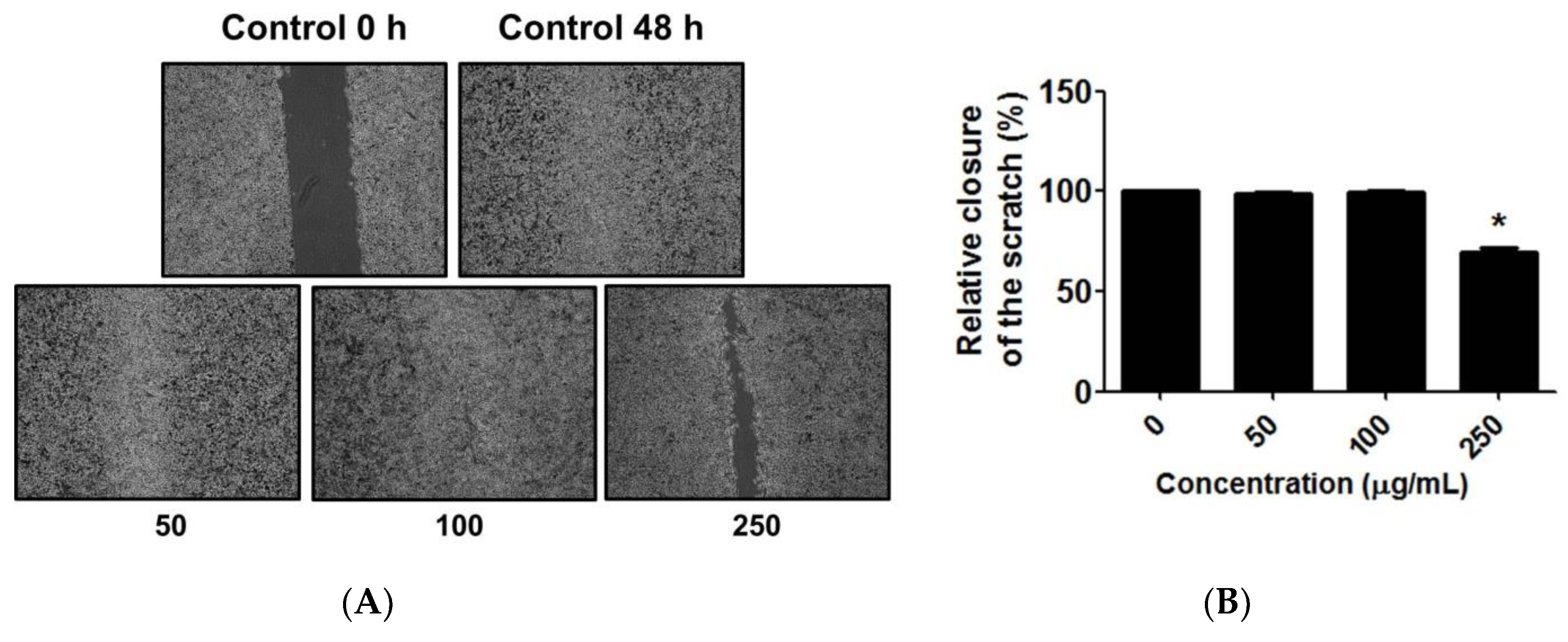
3.12. Effect of C. chinense Leaf Extract on Migration of A549 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer treatment and survivorship statistics, 2022. CA A Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Kung, P.-T.; Kuo, W.-Y.; Tsai, W.-C. Effect of time interval from diagnosis to treatment for non-small cell lung cancer on survival: A national cohort study in Taiwan. BMJ Open 2020, 10, e034351. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef]
- Huang, R.; Chen, H.; Liang, J.; Li, Y.; Yang, J.; Luo, C.; Tang, Y.; Ding, Y.; Liu, X.; Yuan, Q.; et al. Dual Role of Reactive Oxygen Species and their Application in Cancer Therapy. J. Cancer 2021, 12, 5543–5561. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Pei, X.; Xiao, J.; Wei, G.; Zhang, Y.; Lin, F.; Xiong, Z.; Lu, L.; Wang, X.; Pang, G.; Jiang, Y.; et al. Oenothein B inhibits human non-small cell lung cancer A549 cell proliferation by ROS-mediated PI3K/Akt/NF-κB signaling pathway. Chem. Biol. Interact. 2019, 298, 112–120. [Google Scholar] [CrossRef]
- Wahba, H.M.; AbouZid, S.F.; Sleem, A.A.; Apers, S.; Pieters, L.; Shahat, A.A. Chemical and biological investigation of some Clerodendrum species cultivated in Egypt. Pharm. Biol. 2011, 49, 66–72. [Google Scholar] [CrossRef]
- Gao, L.-M.; Wei, X.-M.; He, Y.-Q. Studies on chemical constituents in leafs of Clerodendron fragrans. China J. Chin. Mater. Med. 2003, 28, 948–951. [Google Scholar]
- Shrivastava, N.; Patel, T. Clerodendrum and Healthcare: An Overview-Part II Phytochemistry and Biotechnology. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 209–223. [Google Scholar]
- Wang, J.H.; Luan, F.; He, X.D.; Wang, Y.; Li, M.X. Traditional uses and pharmacological properties of Clerodendrum phytochemicals. J. Tradit. Complement. Med. 2018, 8, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Abouzid, S.F.; Wahba, H.M.; Elshamy, A.; Cos, P.; Maes, L.; Apers, S.; Pieters, L.; Shahat, A.A. Antimicrobial activity of some Clerodendrum species from Egypt. Nat. Prod. Res. 2013, 27, 1032–1036. [Google Scholar] [CrossRef]
- Chiangnoon, R.; Samee, W.; Uttayarat, P.; Jittachai, W.; Ruksiriwanich, W.; Sommano, S.R.; Athikomkulchai, S.; Chittasupho, C. Phytochemical Analysis, Antioxidant, and Wound Healing Activity of Pluchea indica L. (Less) Branch Extract Nanoparticles. Molecules 2022, 27, 635. [Google Scholar] [CrossRef]
- Tunit, P.; Thammarat, P.; Okonogi, S.; Chittasupho, C. Hydrogel Containing Borassus flabellifer L. Male Flower Extract for Antioxidant, Antimicrobial, and Anti-Inflammatory Activity. Gels 2022, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Byrne, F.P.; Jin, S.; Paggiola, G.; Petchey, T.H.M.; Clark, J.H.; Farmer, T.J.; Hunt, A.J.; Robert McElroy, C.; Sherwood, J. Tools and techniques for solvent selection: Green solvent selection guides. Sustain. Chem. Process. 2016, 4, 7. [Google Scholar] [CrossRef]
- Sapiun, Z.; Pangalo, P.; Imran, A.K.; Wicita, P.S.; Astuti Daud, R.P. Determination of Total Flavonoid Levels of Ethanol Extract Sesewanua Leaf (Clerodendrum Fragrans Wild) With Maceration Method Using UV-Vis Spectrofotometry. Polym. J. 2020, 12, 356–360. [Google Scholar] [CrossRef]
- Simorangkir, M.; Nainggolan, B.; Silaban, S. Secondary Metabolites Phytochemical Analysis of n-Hexane, Ethyl Acetate and Ethanol Extracts of Sarang Banua (Clerodendrum fragrans Vent Willd) Leaves. In AISTSSE 2018, Proceedings of the 5th Annual International Seminar on Trends in Science and Science Education, AISTSSE 2018, 18–19 October 2018; European Alliance for Innovation: Ghent, Belgium, 2019. [Google Scholar] [CrossRef]
- Lee, K.J.; Oh, Y.C.; Cho, W.K.; Ma, J.Y. Antioxidant and Anti-Inflammatory Activity Determination of One Hundred Kinds of Pure Chemical Compounds Using Offline and Online Screening HPLC Assay. Evid.-Based Complement. Altern. Med. 2015, 2015, 165457. [Google Scholar] [CrossRef] [PubMed]
- Swargiary, A.; Brahma, K.; Boro, T.; Daimari, M.; Roy, M. Study of phytochemical content, antioxidant and larvicidal property of different solvent extracts of Clerodendrum infortunatum and Citrus grandis. Indian J. Tradit. Knowl. 2021, 20, 329–334. [Google Scholar]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Barung, E.N.; Kalonio, D.E.; Banne, Y.; Kambuno, N.T. Anticancer Activities of Sesewanua Leaf Extracts (Clerodendrum fragrans (Vent.) Willd) against A549 Lung Cancer Cell. Open Access Maced. J. Med. Sci. 2021, 9, 1226–1230. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Seyfi, D.; Behzad, S.B.; Nabiuni, M.; Parivar, K.; Tahmaseb, M.; Amini, E. Verbascoside Attenuates Rac-1 and HIF-1α Signaling Cascade in Colorectal Cancer Cells. Anti-Cancer Agents Med. Chem. 2018, 18, 2149–2155. [Google Scholar] [CrossRef]
- Jia, W.Q.; Wang, Z.T.; Zou, M.M.; Lin, J.H.; Li, Y.H.; Zhang, L.; Xu, R.X. Verbascoside Inhibits Glioblastoma Cell Proliferation, Migration and Invasion While Promoting Apoptosis through Upregulation of Protein Tyrosine Phosphatase SHP-1 and Inhibition of STAT3 Phosphorylation. Cell. Physiol. Biochem. 2018, 47, 1871–1882. [Google Scholar] [CrossRef]
- Puca, R.; Nardinocchi, L.; Sacchi, A.; Rechavi, G.; Givol, D.; D’Orazi, G. HIPK2 modulates p53 activity towards pro-apoptotic transcription. Mol. Cancer 2009, 8, 85. [Google Scholar] [CrossRef] [PubMed]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Sun, L.; Davis, N.M.; Abrams, S.L.; Franklin, R.A.; Cocco, L.; Evangelisti, C.; Chiarini, F.; et al. Advances in targeting signal transduction pathways. Oncotarget 2012, 3, 1505–1521. [Google Scholar] [CrossRef] [PubMed]
- Vasincu, A.; Neophytou, C.M.; Luca, S.V.; Skalicka-Woźniak, K.; Miron, A.; Constantinou, A.I. 6-O-(3″, 4″-di-O-trans-cinnamoyl)-α-l-rhamnopyranosylcatalpol and verbascoside: Cytotoxicity, cell cycle kinetics, apoptosis, and ROS production evaluation in tumor cells. J. Biochem. Mol. Toxicol. 2020, 34, e22443. [Google Scholar] [CrossRef]
- Lin, H.P.; Jiang, S.S.; Chuu, C.P. Caffeic acid phenethyl ester causes p21 induction, Akt signaling reduction, and growth inhibition in PC-3 human prostate cancer cells. PLoS ONE 2012, 7, e31286. [Google Scholar] [CrossRef]
- Wu, C.H.; Chen, C.H.; Hsieh, P.F.; Lee, Y.H.; Kuo, W.W.; Wu, R.C.; Hung, C.H.; Yang, Y.L.; Lin, V.C. Verbascoside inhibits the epithelial-mesenchymal transition of prostate cancer cells through high-mobility group box 1/receptor for advanced glycation end-products/TGF-β pathway. Environ. Toxicol. 2021, 36, 1080–1089. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, Y.; Huang, B.; Yang, X.; Wu, Y.; Liu, B.; Yuan, Y.; Zhang, G. Evaluation of the Antitumor Activity by Ni Nanoparticles with Verbascoside. J. Nanomater. 2013, 2013, 623497. [Google Scholar] [CrossRef]
- Zhao, X.H.; Yue, H.L.; Li, P.; Zeng, X.; Zhang, G. Evaluation of the antitumor activity by CdTe QDs with verbascoside. Nano 2013, 08, 1350031. [Google Scholar] [CrossRef]
- Chen, R.C.; Su, J.H.; Yang, S.M.; Li, J.; Wang, T.J.; Zhou, H. Effect of isoverbascoside, a phenylpropanoid glycoside antioxidant, on proliferation and differentiation of human gastric cancer cell. Acta Pharmacol. Sin. 2002, 23, 997–1001. [Google Scholar]
- Yang, X.; Guo, F.; Peng, Q.; Liu, Y.; Yang, B. Suppression of in vitro and in vivo human ovarian cancer growth by isoacteoside is mediated via sub-G1 cell cycle arrest, ROS generation, and modulation of AKT/PI3K/m-TOR signalling pathway. J. BUON 2019, 24, 285–290. [Google Scholar] [PubMed]
- Lv, L.; Zhang, W.; Li, T.; Jiang, L.; Lu, X.; Lin, J. Hispidulin exhibits potent anticancer activity in vitro and in vivo through activating ER stress in non-small-cell lung cancer cells. Oncol. Rep. 2020, 43, 1995–2003. [Google Scholar] [CrossRef]
- Ashaq, A.; Maqbool, M.F.; Maryam, A.; Khan, M.; Shakir, H.A.; Irfan, M.; Qazi, J.I.; Li, Y.; Ma, T. Hispidulin: A novel natural compound with therapeutic potential against human cancers. Phytother. Res. 2021, 35, 771–789. [Google Scholar] [CrossRef]
- Han, M.; Gao, H.; Ju, P.; Gao, M.Q.; Yuan, Y.P.; Chen, X.H.; Liu, K.L.; Han, Y.T.; Han, Z.W. Hispidulin inhibits hepatocellular carcinoma growth and metastasis through AMPK and ERK signaling mediated activation of PPARγ. Biomed. Pharmacother. 2018, 103, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhao, F.; Yan, J.; Xia, Z.; Jiang, D.; Ma, P. Hispidulin: A promising flavonoid with diverse anti-cancer properties. Life Sci. 2020, 259, 118395. [Google Scholar] [CrossRef]
- Das, S.K.; Reddy, K.A.; Mukkanti, K. Total synthesis of phenylpropanoid glycosides, grayanoside A and syringalide B, through a common intermediate. Carbohydr. Res. 2007, 342, 2309–2315. [Google Scholar] [CrossRef]
- Kawada, T.; Asano, R.; Makino, K.; Sakuno, T. Synthesis of isoacteoside, a dihydroxyphenylethyl glycoside. J. Wood Sci. 2002, 48, 512–515. [Google Scholar] [CrossRef]
- Kavvadias, D.; Sand, P.; Youdim, K.A.; Qaiser, M.Z.; Rice-Evans, C.; Baur, R.; Sigel, E.; Rausch, W.D.; Riederer, P.; Schreier, P. The flavone hispidulin, a benzodiazepine receptor ligand with positive allosteric properties, traverses the blood-brain barrier and exhibits anticonvulsive effects. Br. J. Pharmacol. 2004, 142, 811–820. [Google Scholar] [CrossRef]
- Chao, S.-W.; Su, M.-Y.; Chiou, L.-C.; Chen, L.-C.; Chang, C.-I.; Huang, W.-J. Total Synthesis of Hispidulin and the Structural Basis for Its Inhibition of Proto-oncogene Kinase Pim-1. J. Nat. Prod. 2015, 78, 1969–1976. [Google Scholar] [CrossRef]
- Lin, H.; Zhang, W.; Dong, Z.X.; Gu, T.; Li, N.G.; Shi, Z.H.; Kai, J.; Qu, C.; Shang, G.X.; Tang, Y.P.; et al. A new and practical synthetic method for the synthesis of 6-O-methyl-scutellarein: One metabolite of scutellarin in vivo. Int. J. Mol. Sci. 2015, 16, 7587–7594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Jiang, Q.; Han, Y.; Peng, J.; Wang, C. Hispidulin potentiates the antitumor effect of sunitinib against human renal cell carcinoma in laboratory models. Cell Biochem. Biophys. 2015, 71, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, W.; He, X.; Fei, Z. Hispidulin enhances the anti-tumor effects of temozolomide in glioblastoma by activating AMPK. Cell Biochem. Biophys. 2015, 71, 701–706. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenolic Content (mg GAE/g) | Total Flavonoid Content (mg GAE/g) | |
|---|---|---|
| C. chinense leaf ethanol extract | 1029.99 ± 21.29 mg | 326.37 ± 24.54 |
| Parameters | Verbascoside | Isoverbascoside | Hispidulin |
|---|---|---|---|
| Regression equation | y = 89.026x − 2.301 | y = 89.577x − 32.786 | y = 318.83x − 21.409 |
| Correlation coefficient (r) | 0.9999 | 0.9999 | 0.9993 |
| Linear range (µg/mL) | 1–9 | 1–9 | 0.3–1.5 |
| LOD (µg/mL) | 0.0039 | 0.0790 | 0.0213 |
| LOQ (µg/mL) | 0.0130 | 0.2633 | 0.0711 |
| Concentration | Verbascoside | Isoverbascoside | Hispidulin | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1.5 | 4.8 | 8.0 | 2.0 | 5.0 | 8.0 | 0.5 | 0.9 | 1.3 | |
| Mean recovery, % | 95.69 ± 0.31 | 98.04 ± 1.30 | 102.51 ± 2.17 | 104.45 ± 1.42 | 100.90 ± 0.34 | 103.64 ± 1.36 | 97.70 ± 0.09 | 102.56 ± 0.95 | 103.35 ± 0.40 |
| Day 1 | Day 2 | Day 3 | Day 1 | Day 2 | Day 3 | Day 1 | Day 2 | Day 3 | |
| Intraday, %RSD | 0.25–0.78 | 0.11–0.46 | 0.12–0.50 | 0.20–0.64 | 0.32–0.38 | 0.13–0.82 | 0.26–0.77 | 0.29–0.68 | 0.32–0.65 |
| Interday, % RSD | 0.33–2.12 | 0.33–1.36 | 0.09–0.93 | ||||||
| Total Phenolic Content (Gallic Acid Equivalent) | Total Flavonoid Content (Quercetin Equivalent) | |
|---|---|---|
| DPPH assay | 0.9557 * | 0.8829 * |
| ABTS assay | 0.9990 **** | 0.9997 **** |
| FRAP assay | 0.9994 **** | 0.9987 **** |
| Group of Treatment | Viable Cells (%) | Late Apoptosis (%) | Necrosis (%) |
|---|---|---|---|
| Control | 83.12 ± 3.02 | 3.95 ± 0.14 | 0.12 ± 0.07 |
| CCL 50 µg/mL | 75.60 ± 1.18 * | 15.28 ± 1.37 * | 1.97 ± 0.11 |
| CCL 100 µg/mL | 68.10 ± 1.29 * | 14.50 ± 1.25 * | 3.97 ± 0.48 |
| CCL 250 µg/mL | 63.57 ± 3.35 * | 21.67 ± 1.55 * | 5.75 ± 0.61 * |
| Control | Extract 50 µg/mL | Extract 100 µg/mL | Extract 250 µg/mL | |
|---|---|---|---|---|
| ROS formation (%) | 2.22 ± 0.59 | 1.70 ± 0.33 | 5.42 ± 1.96 | 15.95 ± 3.95 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chittasupho, C.; Athikomkulchai, S.; Samee, W.; Na Takuathung, M.; Yooin, W.; Sawangrat, K.; Saenjum, C. Phenylethanoid Glycoside-Enriched Extract Prepared from Clerodendrum chinense Leaf Inhibits A549 Lung Cancer Cell Migration and Apoptosis Induction through Enhancing ROS Production. Antioxidants 2023, 12, 461. https://doi.org/10.3390/antiox12020461
Chittasupho C, Athikomkulchai S, Samee W, Na Takuathung M, Yooin W, Sawangrat K, Saenjum C. Phenylethanoid Glycoside-Enriched Extract Prepared from Clerodendrum chinense Leaf Inhibits A549 Lung Cancer Cell Migration and Apoptosis Induction through Enhancing ROS Production. Antioxidants. 2023; 12(2):461. https://doi.org/10.3390/antiox12020461
Chicago/Turabian StyleChittasupho, Chuda, Sirivan Athikomkulchai, Weerasak Samee, Mingkwan Na Takuathung, Wipawadee Yooin, Kasirawat Sawangrat, and Chalermpong Saenjum. 2023. "Phenylethanoid Glycoside-Enriched Extract Prepared from Clerodendrum chinense Leaf Inhibits A549 Lung Cancer Cell Migration and Apoptosis Induction through Enhancing ROS Production" Antioxidants 12, no. 2: 461. https://doi.org/10.3390/antiox12020461