
Valorization of Peels of Eight Peach Varieties: GC–MS Profile, Free and Bound Phenolics and Corresponding Biological Activities

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Samples
2.2. Gas Chromatographic–Mass Spectrometry Analysis (GC-MS)
2.3. Free and Bound Phenolic Compounds Extraction
2.3.1. Extraction of Free Phenolic Compounds
2.3.2. Extraction of Bound Phenolic Compounds
Alkaline Hydrolysis Method
Acid Hydrolysis Method
2.4. Determination of Total Phenolic Contents (TPC)
2.5. Determination of Total Flavonoid Content (TFC)
2.6. Determination of Total Monomeric Anthocyanins (TMA)
2.7. Evaluation of Antioxidant Activities of Phenolic (Free and Bound) Fractions
2.7.1. DPPH• Radical Scavenging Assay
2.7.2. ABTS•+ Radical Scavenging Assay
2.7.3. Ferric-Reducing Antioxidant Power (FRAP) Assay
2.7.4. Cupric Ion-Reducing Antioxidant Capacity (CUPRAC) Assay
2.8. Enzyme-Inhibitory Activities
2.9. Antimicrobial Activity
2.9.1. Test Microorganisms
2.9.2. Culture Media
2.9.3. Antimicrobial Activity Assay
2.10. Statistical Analyses
3. Results and Discussion
3.1. GC-MS Volatile Profile Characterization of Analyzed Peach Peels
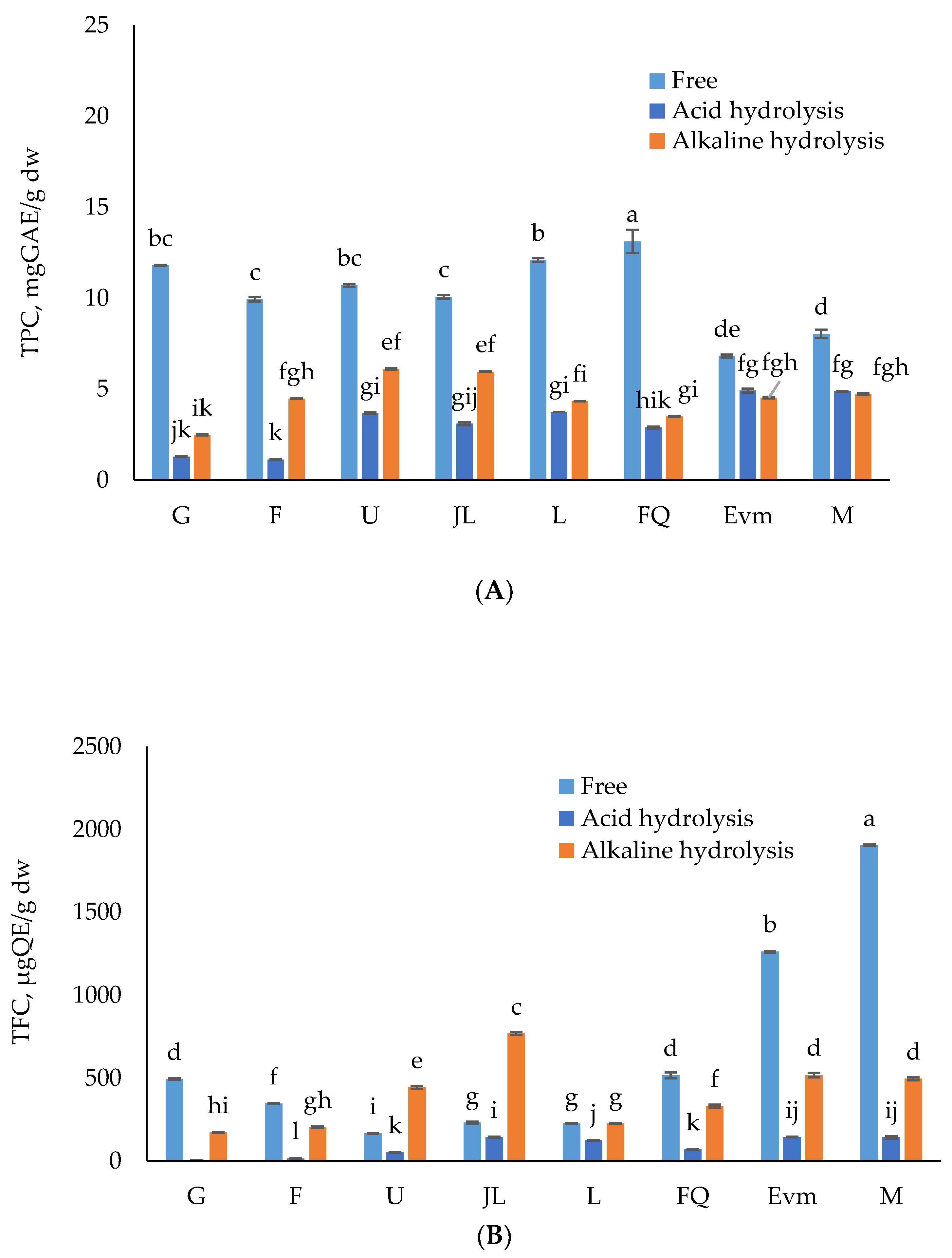
3.2. Total Phenolic, Flavonoid and Total Monomeric Anthocyanins Contents of Free and Bound Insoluble Fractions
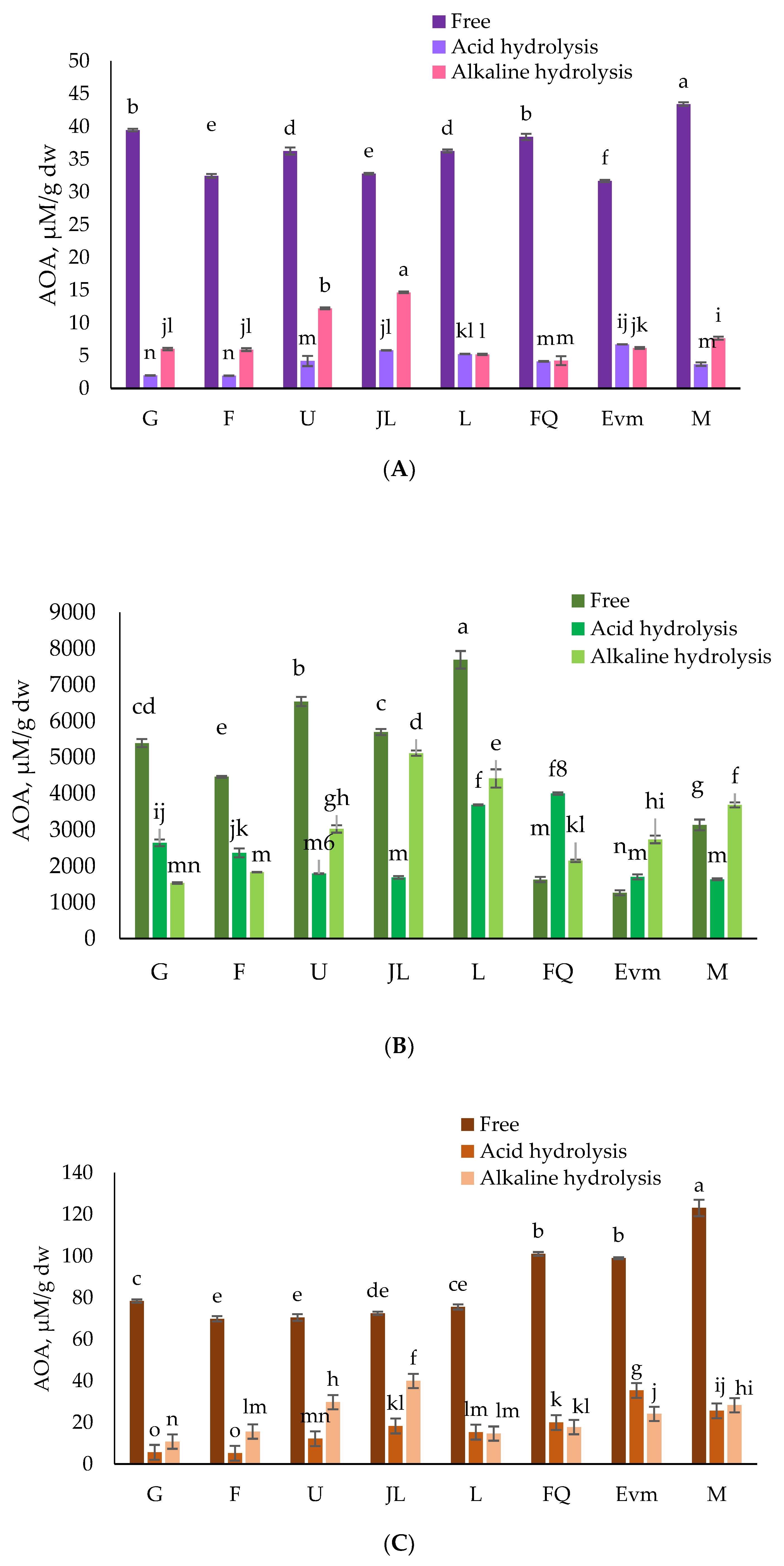
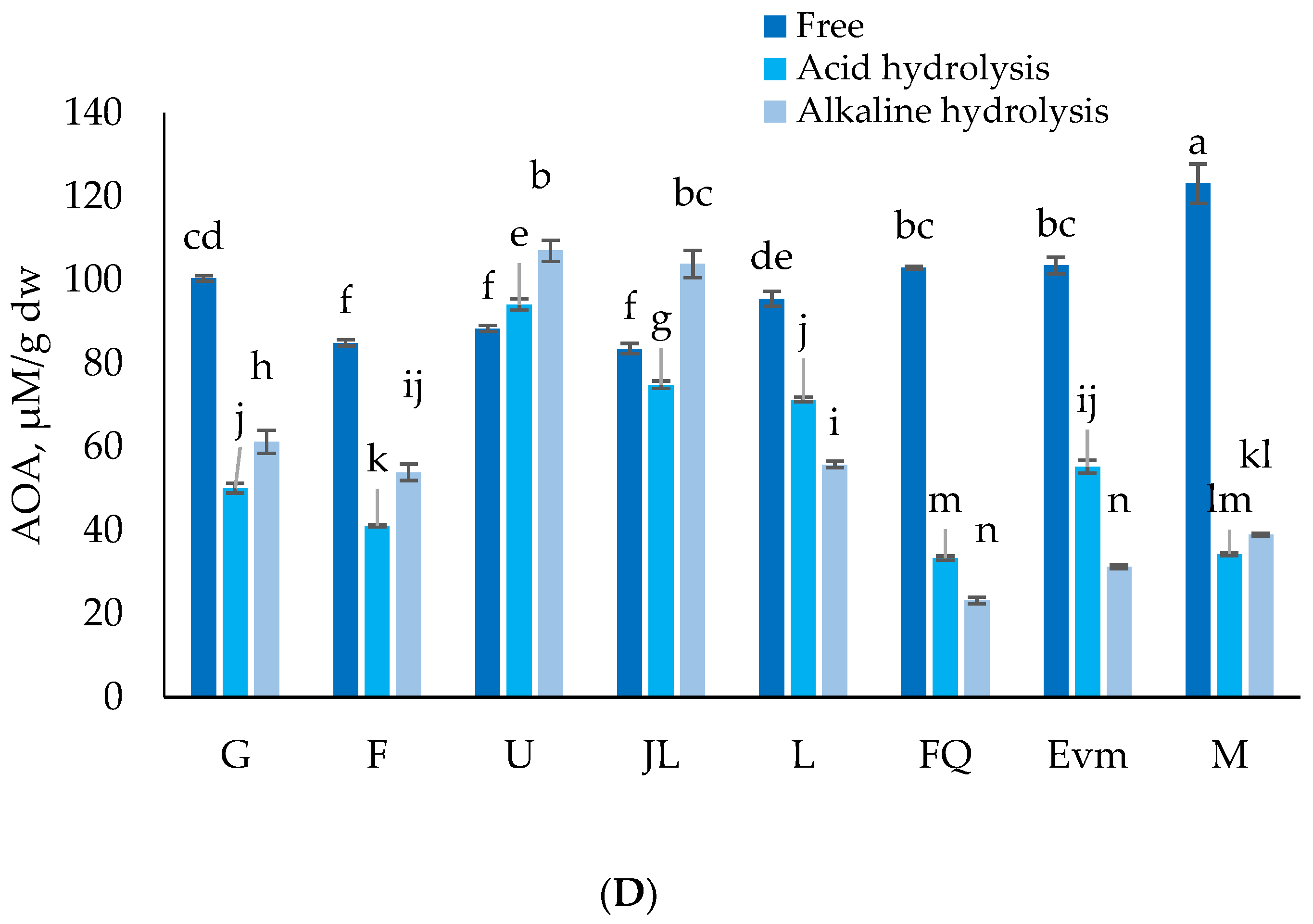
3.3. Antioxidant Activity (AOA)
3.4. Inhibitory Potential towards α-Glucosidase, α-Amylase, Lipase, and Acetylcholinesterase of Analyzed Prunus Persica Peels
3.5. Antimicrobial Activity of Peach Peel Extracts
3.6. Correlation between Phenolic Compounds Content and Antioxidant Activity
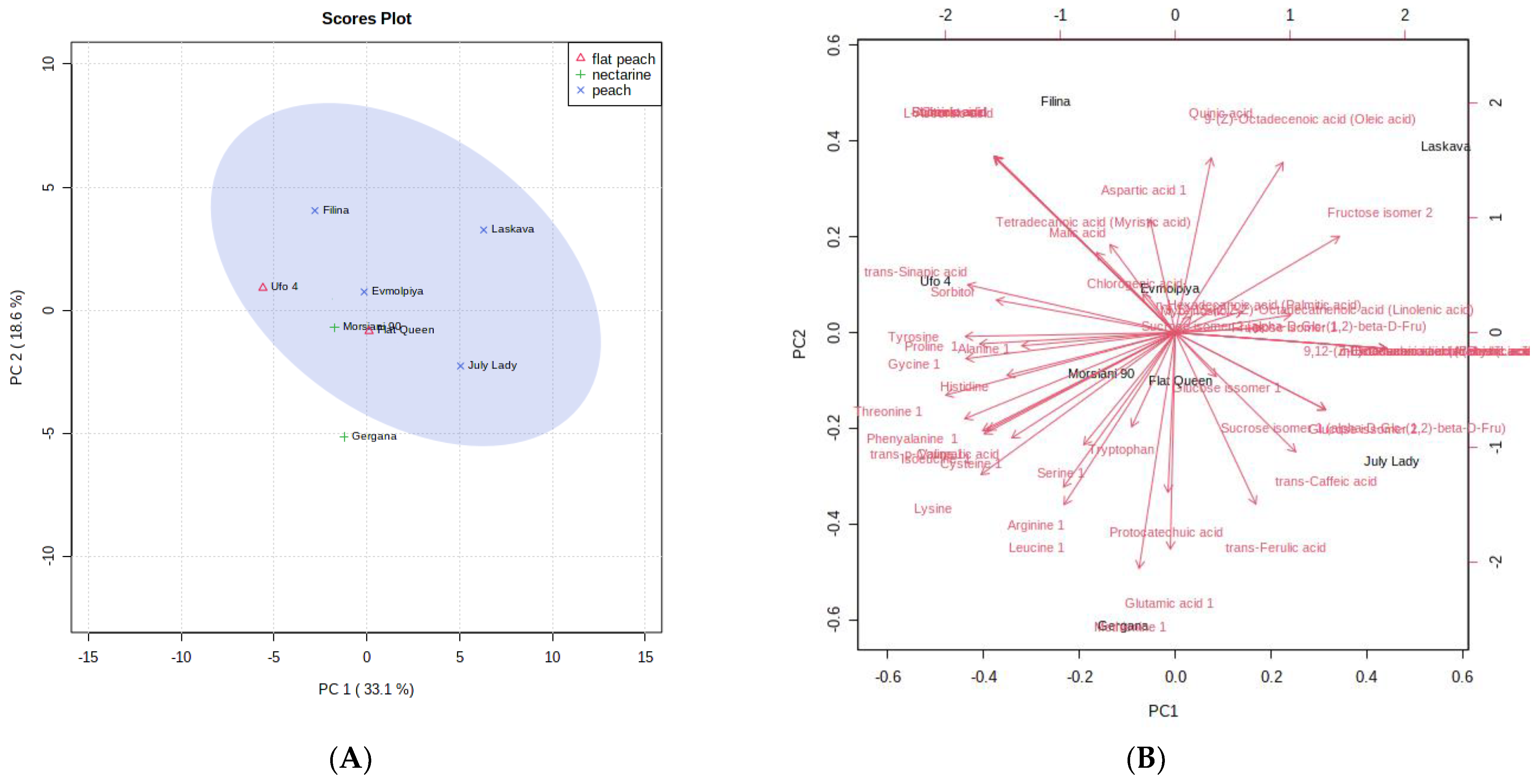
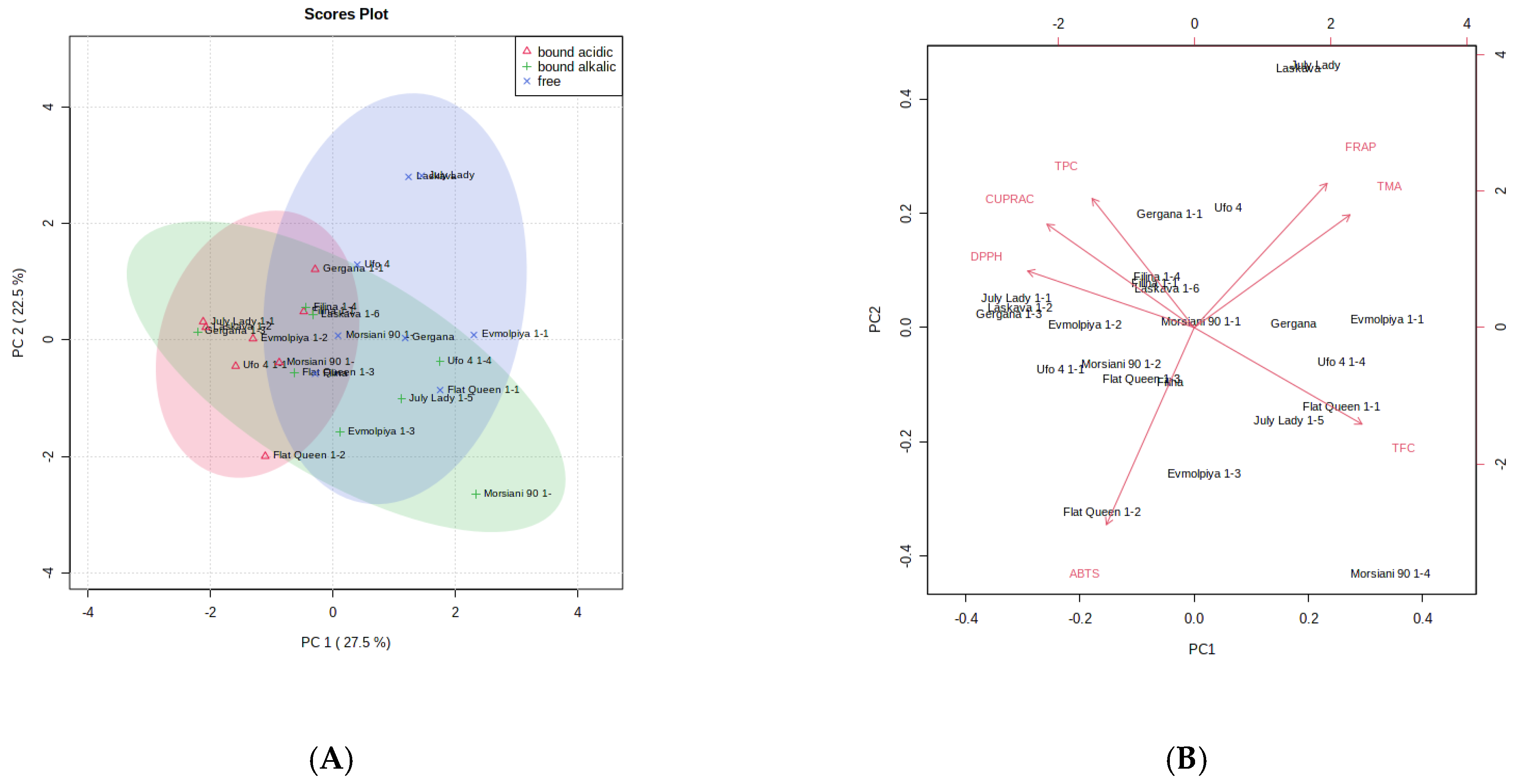
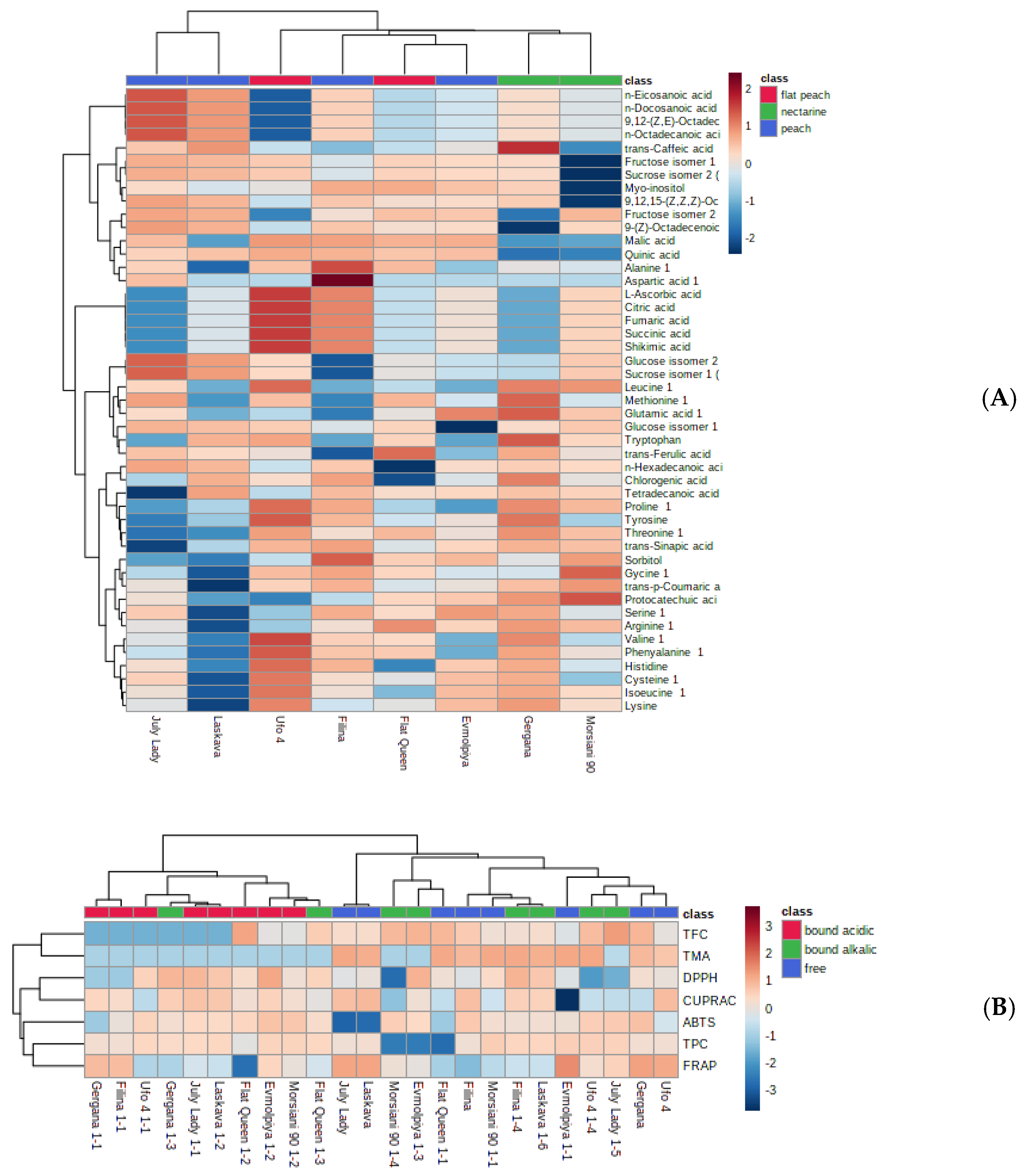
3.7. Principal Component Analysis (PCA) and Hierarchical Cluster Analysis (HCA) of GC-MS and Phenolic Compound and AOA Assays Data
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussein, R.A. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. In Herbal Medicine; Builders, P.F., Ed.; IntechOpen: Rijeka, Croatia, 2018; Chapter 2; ISBN 978-1-78984-783-3. [Google Scholar]
- Dziadek, K.; Kopeć, A.; Piątkowska, E.; Leszczyńska, T. High-Fructose Diet-Induced Metabolic Disorders Were Counteracted by the Intake of Fruit and Leaves of Sweet Cherry in Wistar Rats. Nutrients 2019, 11, 2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzma, J.N.; Schmidt, K.A.; Kratz, M. Prevention of metabolic diseases: Fruits (including fruit sugars) vs. vegetables. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From Central to Specialized Metabolism: An Overview of Some Secondary Compounds Derived From the Primary Metabolism for Their Role in Conferring Nutritional and Organoleptic Characteristics to Fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şahin, S.; Bilgin, M. Valorization of peach (Prunus persica L.) waste into speciality products via green methods. Biomass Convers. Biorefinery 2022, 12, 123–132. [Google Scholar] [CrossRef]
- Yüksekkaya, Ş.; Başyiğit, B.; Sağlam, H.; Pekmez, H.; Cansu, Ü.; Karaaslan, A.; Karaaslan, M. Valorization of fruit processing by-products: Free, esterified, and insoluble bound phytochemical extraction from cherry (Prunus avium) tissues and their biological activities. J. Food Meas. Charact. 2021, 15, 1092–1107. [Google Scholar] [CrossRef]
- Food and Agriculture Organization Reducing Food Loss and Waste in Fruits and Vegetables, for Improving Access to Healthy Diets (Webinar). 2022. Available online: https://www.fao.org/food-loss-reduction/events-and-opportunities/detail/en/c/1458146/ (accessed on 8 January 2023).
- Kumar, H.; Bhardwaj, K.; Sharma, R.; Nepovimova, E.; Kuča, K.; Dhanjal, D.S.; Verma, R.; Bhardwaj, P.; Sharma, S.; Kumar, D. Fruit and Vegetable Peels: Utilization of High Value Horticultural Waste in Novel Industrial Applications. Molecules 2020, 25, 2812. [Google Scholar] [CrossRef]
- Bhardwaj, K.; Najda, A.; Sharma, R.; Nurzyńska-Wierdak, R.; Dhanjal, D.S.; Sharma, R.; Manickam, S.; Kabra, A.; Kuča, K.; Bhardwaj, P. Fruit and Vegetable Peel-Enriched Functional Foods: Potential Avenues and Health Perspectives. Evid.-Based Complement. Altern. Med. 2022, 2022, 8543881. [Google Scholar] [CrossRef]
- Imran, M.; Khan, M.K.; Ahmad, M.H.; Ahmad, R.S.; Javed, M.R.; Nisa, M.U.; Nadeem, M.; Liaqat, F.; Ahmad, U.; Rahim, M.A. Valorization of Peach (Prunus persica) Fruit Waste. In Mediterranean Fruits Bio-Wastes: Chemistry, Functionality and Technological Applications; Ramadan, M.F., Farag, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 589–604. ISBN 978-3-030-84436-3. [Google Scholar]
- Lucarini, M.; Durazzo, A.; Bernini, R.; Campo, M.; Vita, C.; Souto, E.B.; Lombardi-Boccia, G.; Ramadan, M.F.; Santini, A.; Romani, A. Fruit Wastes as a Valuable Source of Value-Added Compounds: A Collaborative Perspective. Molecules 2021, 26, 6338. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Mihaylova, D.; Popova, A.; Alexieva, I.; Krastanov, A.; Lante, A. Polyphenols as Suitable Control for Obesity and Diabetes. Open Biotechnol. J. 2018, 12, 219–228. [Google Scholar] [CrossRef]
- Minatel, I.O. Phenolic compounds: Functional properties, impact of processing and bioavailability. Phenolic Compounds—Biological Activity; Borges, C.V., Ed.; IntechOpen: Rijeka, Croatia, 2017; Chapter 1; ISBN 978-953-51-2960-8. [Google Scholar]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Nat. Prod. Commun. 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Aluko, R.E. Food-derived Acetylcholinesterase Inhibitors as Potential Agents against Alzheimer’s Disease. eFood 2021, 2, 49–58. [Google Scholar] [CrossRef]
- Jo, Y.H.; Kim, S.B.; Liu, Q.; Do, S.-G.; Hwang, B.Y.; Lee, M.K. Comparison of pancreatic lipase inhibitory isoflavonoids from unripe and ripe fruits of Cudrania tricuspidata. PLoS ONE 2017, 12, e0172069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Saura-Calixto, F. Fruit peels as sources of non-extractable polyphenols or macromolecular antioxidants: Analysis and nutritional implications. Food Res. Int. 2018, 111, 148–152. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic Compounds of Pomegranate Byproducts (Outer Skin, Mesocarp, Divider Membrane) and Their Antioxidant Activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef]
- Nayak, B.; Liu, R.H.; Tang, J. Effect of Processing on Phenolic Antioxidants of Fruits, Vegetables, and Grains—A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 887–918. [Google Scholar] [CrossRef]
- Mohd Basri, M.S.; Abdul Karim Shah, N.N.; Sulaiman, A.; Mohamed Amin Tawakkal, I.S.; Mohd Nor, M.Z.; Ariffin, S.H.; Abdul Ghani, N.H.; Mohd Salleh, F.S. Progress in the Valorization of Fruit and Vegetable Wastes: Active Packaging, Biocomposites, By-Products, and Innovative Technologies Used for Bioactive Compound Extraction. Polymers 2021, 13, 3503. [Google Scholar] [CrossRef]
- AL-Huqail, A.A.; Kumar, V.; Kumar, R.; Eid, E.M.; Taher, M.A.; Adelodun, B.; Abou Fayssal, S.; Mioč, B.; Držaić, V.; Goala, M.; et al. Sustainable Valorization of Four Types of Fruit Peel Waste for Biogas Recovery and Use of Digestate for Radish (Raphanus sativus L. cv. Pusa Himani) Cultivation. Sustainability 2022, 14, 10224. [Google Scholar] [CrossRef]
- Mihaylova, D.; Desseva, I.; Popova, A.; Dincheva, I.; Vrancheva, R.; Lante, A.; Krastanov, A. GC-MS Metabolic Profile and α-Glucosidase-, α-Amylase-, Lipase-, and Acetylcholinesterase-Inhibitory Activities of Eight Peach Varieties. Molecules 2021, 26, 4183. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Morozova, K.; Scampicchio, M.; Ferrentino, G. Non-Extractable Polyphenols from Food By-Products: Current Knowledge on Recovery, Characterisation, and Potential Applications. Processes 2020, 8, 925. [Google Scholar] [CrossRef]
- Vargas-Madriz, Á.F.; Kuri-García, A.; Vargas-Madriz, H.; Chávez-Servín, J.L.; Ferriz-Martínez, R.A.; Hernández-Sandoval, L.G.; Guzmán-Maldonado, S.H. Phenolic profile and antioxidant capacity of Pithecellobium dulce (Roxb) Benth: A review. J. Food Sci. Technol. 2020, 57, 4316–4336. [Google Scholar] [CrossRef] [PubMed]
- Kujala, T.S.; Loponen, J.M.; Klika, K.D.; Pihlaja, K. Phenolics and betacyanins in red beetroot (Beta vulgaris) root: Distribution and effect of cold storage on the content of total phenolics and three individual compounds. J. Agric. Food Chem. 2000, 48, 5338–5342. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, D.; Lante, A.; Krastanov, A. Total phenolic content, antioxidant and antimicrobial activity of Haberlea rhodopensis extracts obtained by pressurized liquid extraction. Acta Aliment. 2015, 44, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Kivrak, İ.; Duru, M.E.; Öztürk, M.; Mercan, N.; Harmandar, M.; Topçu, G. Antioxidant, anticholinesterase and antimicrobial constituents from the essential oil and ethanol extract of Salvia potentillifolia. Food Chem. 2009, 116, 470–479. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Collaborators: Determination of Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines by the pH Differential Method: Collaborative Study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 15–27. ISBN 0076-6879. [Google Scholar]
- Apak, R.; Özyürek, M.; Karademir Çelik, S.; Güçlü, K. CUPRAC method 2004. JAFC 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Sigma-Aldrich Enzymatic Assay of α-Amylase (EC 3.2.1.1). Determination of Alfpha-Amylase Activity. Available online: https://www.sigmaaldrich.com/BG/en/technical-documents/protocol/protein-biology/enzyme-activity-assays/enzymatic-assay-of-a-amylase (accessed on 7 September 2020).
- Saifuddin, N.; Raziah, A.Z. Enhancement of Lipase Enzyme Activity in Non-Aqueous Media through a Rapid Three Phase Partitioning and Microwave Irradiation. E-J. Chem. 2008, 5, 920408. [Google Scholar] [CrossRef]
- Dobrev, G.; Zhekova, B.; Dobreva, V.; Strinska, H.; Doykina, P.; Krastanov, A. Lipase biosynthesis by Aspergillus carbonarius in a nutrient medium containing products and byproducts from the oleochemical industry. Biocatal. Agric. Biotechnol. 2015, 4, 77–82. [Google Scholar] [CrossRef]
- Lobbens, E.S.B.; Vissing, K.J.; Jorgensen, L.; van de Weert, M.; Jäger, A.K. Screening of plants used in the European traditional medicine to treat memory disorders for acetylcholinesterase inhibitory activity and anti amyloidogenic activity. J. Ethnopharmacol. 2017, 200, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Tumbarski, Y.; Deseva, I.; Mihaylova, D.; Stoyanova, M.; Krastev, L.; Nikolova, R.; Yanakieva, V.; Ivanov, I. Isolation, Characterization and Amino Acid Composition of a Bacteriocin Produced by Bacillus methylotrophicus Strain BM47. Food Technol. Biotechnol. 2018, 56, 546–552. [Google Scholar] [CrossRef]
- Assaad, H.I.; Zhou, L.; Carroll, R.J.; Wu, G. Rapid publication-ready MS-Word tables for one-way ANOVA. Springerplus 2014, 3, 474. [Google Scholar] [CrossRef] [Green Version]
- Chong, J., Wishart. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Chahardoli, A.; Jalilian, F.; Memariani, Z.; Farzaei, M.H.; Shokoohinia, Y. Chapter 26—Analysis of organic acids. In Recent Advances in Natural Products Analysis; Sanches Silva, A., Nabavi, S.F., Saeedi, M., Nabavi, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 767–823. ISBN 978-0-12-816455-6. [Google Scholar]
- Singh, P.; Gupta, E.; Mishra, N.; Mishra, P. Chapter 16—Shikimic acid as intermediary model for the production of drugs effective against influenza virus. In Phytochemicals as Lead Compounds for New Drug Discovery; Egbuna, C., Kumar, S., Ifemeje, J.C., Ezzat, S.M., Kaliyaperumal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 245–256. ISBN 978-0-12-817890-4. [Google Scholar]
- Rabelo, T.K.; Guimarães, A.G.; Oliveira, M.A.; Gasparotto, J.; Serafini, M.R.; de Souza Araújo, A.A.; Quintans-Júnior, L.J.; Moreira, J.C.F.; Gelain, D.P. Shikimic acid inhibits LPS-induced cellular pro-inflammatory cytokines and attenuates mechanical hyperalgesia in mice. Int. Immunopharmacol. 2016, 39, 97–105. [Google Scholar] [CrossRef]
- Su, C.-H.; Hsu, C.-H.; Ng, L.-T. Inhibitory potential of fatty acids on key enzymes related to type 2 diabetes. BioFactors 2013, 39, 415–421. [Google Scholar] [CrossRef]
- Hoang Anh, L.; Xuan, T.D.; Dieu Thuy, N.T.; Quan, N.V.; Trang, L.T. Antioxidant and α-amylase Inhibitory Activities and Phytocompounds of Clausena indica Fruits. Medicines 2020, 7, 10. [Google Scholar] [CrossRef]
- Loesche, A.; Wiemann, J.; Al Halabi, Z.; Karasch, J.; Sippl, W.; Csuk, R. Unexpected AChE inhibitory activity of (2E)α,β-unsaturated fatty acids. Bioorg. Med. Chem. Lett. 2018, 28, 3315–3319. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, C.; Ravaglia, D.; Ragaini, A.; Costa, G. Phenolic compounds in peach (Prunus persica) cultivars at harvest and during fruit maturation. Ann. Appl. Biol. 2008, 153, 11–23. [Google Scholar] [CrossRef]
- Soto-Hernandez, M.; Palma-Tenango, M.; Garcia-Mateos, M. (Eds.) Phenolic Compounds; IntechOpen: Rijeka, Croatia, 2017; ISBN 978-953-51-2960-8. [Google Scholar]
- Liu, H.; Jiang, W.; Cao, J.; Li, Y. Changes in Extractable and Non-extractable Polyphenols and Their Antioxidant Properties during Fruit On-tree Ripening in Five Peach Cultivars. Hortic. Plant J. 2019, 5, 137–144. [Google Scholar] [CrossRef]
- Hong, Y.; Wang, Z.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. High-Throughput Screening and Characterization of Phenolic Compounds in Stone Fruits Waste by LC-ESI-QTOF-MS/MS and Their Potential Antioxidant Activities. Antioxidants 2021, 10, 234. [Google Scholar] [CrossRef]
- Mihaylova, D.; Popova, A.; Desseva, I.; Petkova, N.; Stoyanova, M.; Vrancheva, R.; Slavov, A.; Slavchev, A.; Lante, A. Comparative Study of Early- and Mid-Ripening Peach (Prunus persica L.) Varieties: Biological Activity, Macro-, and Micro- Nutrient Profile. Foods 2021, 10, 164. [Google Scholar] [CrossRef]
- Esmaeilzadeh Kenari, R.; Razavi, R. Phenolic profile and antioxidant activity of free/bound phenolic compounds of sesame and properties of encapsulated nanoparticles in different wall materials. Food Sci. Nutr. 2022, 10, 525–535. [Google Scholar] [CrossRef]
- Ti, H.; Li, Q.; Zhang, R.; Zhang, M.; Deng, Y.; Wei, Z.; Chi, J.; Zhang, Y. Free and bound phenolic profiles and antioxidant activity of milled fractions of different indica rice varieties cultivated in southern China. Food Chem. 2014, 159, 166–174. [Google Scholar] [CrossRef]
- Tang, W.; Li, W.; Yang, Y.; Lin, X.; Wang, L.; Li, C.; Yang, R. Phenolic Compounds Profile and Antioxidant Capacity of Pitahaya Fruit Peel from Two Red-Skinned Species (Hylocereus polyrhizus and Hylocereus undatus). Foods 2021, 10, 1183. [Google Scholar] [CrossRef]
- Li, W.; Yang, R.; Ying, D.; Yu, J.; Sanguansri, L.; Augustin, M.A. Analysis of polyphenols in apple pomace: A comparative study of different extraction and hydrolysis procedures. Ind. Crops Prod. 2020, 147, 112250. [Google Scholar] [CrossRef]
- Hung, P.V.; Duy, T.L. Effects of drying methods on bioactive compounds of vegetables and correlation between bioactive compounds and their antioxidants. Int. Food Res. J. 2012, 19, 327–332. [Google Scholar]
- Loypimai, P.; Moongngarm, A.; Chottanom, P. Thermal and pH degradation kinetics of anthocyanins in natural food colorant prepared from black rice bran. J. Food Sci. Technol. 2016, 53, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Li, W.; Deng, Z.; Li, H.; Zhang, B. The Composition and Antioxidant Activity of Bound Phenolics in Three Legumes, and Their Metabolism and Bioaccessibility of Gastrointestinal Tract. Foods 2020, 9, 1816. [Google Scholar] [CrossRef]
- Kim, K.-H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Chen, G.-L.; Zhang, X.; Chen, S.-G.; Han, M.-D.; Gao, Y.-Q. Antioxidant activities and contents of free, esterified and insoluble-bound phenolics in 14 subtropical fruit leaves collected from the south of China. J. Funct. Foods 2017, 30, 290–302. [Google Scholar] [CrossRef]
- Kim, J.-S. Antioxidant Activities of Selected Berries and Their Free, Esterified, and Insoluble-Bound Phenolic Acid Contents. Prev. Nutr. Food Sci. 2018, 23, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Zhang, R.; Hou, F.; Zhang, M.; Guo, J.; Huang, F.; Deng, Y.; Wei, Z. Comparison of the free and bound phenolic profiles and cellular antioxidant activities of litchi pulp extracts from different solvents. BMC Complement. Altern. Med. 2014, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Arranz, S.; Saura-calixto, F.; Shaha, S.; Kroon, P.A. High contents of nonextractable polyphenols in fruits suggest that polyphenol contents of plant foods have been underestimated. J. Agric. Food Chem. 2009, 57, 7298–7303. [Google Scholar] [CrossRef]
- Verma, B.K.; Hucl, P.J.; Chibbar, R.N. Phenolic acid composition and antioxidant capacity of acid and alkali hydrolysed wheat bran fractions. Food Chem. 2009, 116, 947–954. [Google Scholar] [CrossRef]
- Manzoor, M.; Anwar, F.; Mahmood, Z.; Rashid, U.; Ashraf, M. Variation in minerals, phenolics and antioxidant activity of peel and pulp of different varieties of peach (Prunus persica L.) fruit from Pakistan. Molecules 2012, 17, 6491–6506. [Google Scholar] [CrossRef] [PubMed]
- Rossato, S.B.; Haas, C.; Raseira, M.D.C.B.; Moreira, J.C.F.; Zuanazzi, J.Â.S. Antioxidant Potential of Peels and Fleshes of Peaches from Different Cultivars. J. Med. Food 2009, 12, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Karagiannis, E.; Nasiopoulou, E.; Skodra, C.; Molassiotis, A.; Tanou, G. Peach, Apple, and Pear Fruit Quality: To Peel or Not to Peel? Horticulturae 2021, 7, 85. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure-Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.; Mamadalieva, N.Z.; Hussain, A.; Hassan, U.; Rabnawaz, A.; Ahmed, I.; Green, I.R. Fruit Peels: Food Waste as a Valuable Source of Bioactive Natural Products for Drug Discovery. Curr. Issues Mol. Biol. 2022, 44, 1960–1994. [Google Scholar] [CrossRef]
- Belhadj, F.; Somrani, I.; Aissaoui, N.; Messaoud, C.; Boussaid, M.; Marzouki, M.N. Bioactive compounds contents, antioxidant and antimicrobial activities during ripening of Prunus persica L. varieties from the North West of Tunisia. Food Chem. 2016, 204, 29–36. [Google Scholar] [CrossRef]
- Koyu, H.; Kazan, A.; Nalbantsoy, A.; Yalcin, H.T.; Yesil-Celiktas, O. Cytotoxic, antimicrobial and nitric oxide inhibitory activities of supercritical carbon dioxide extracted Prunus persica leaves. Mol. Biol. Rep. 2020, 47, 569–581. [Google Scholar] [CrossRef]
- Mocanu, A.; Isopencu, G.; Busuioc, C.; Popa, O.-M.; Dietrich, P.; Socaciu-Siebert, L. Bacterial cellulose films with ZnO nanoparticles and propolis extracts: Synergistic antimicrobial effect. Sci. Rep. 2019, 9, 17687. [Google Scholar] [CrossRef] [Green Version]
- van der Maas, L.; Driessen, J.L.S.P.; Mussatto, S.I. Effects of Inhibitory Compounds Present in Lignocellulosic Biomass Hydrolysates on the Growth of Bacillus subtilis. Energies 2021, 14, 8419. [Google Scholar] [CrossRef]
- Ibraheem, O.; Ndimba, B.K. Molecular adaptation mechanisms employed by ethanologenic bacteria in response to lignocellulose-derived inhibitory compounds. Int. J. Biol. Sci. 2013, 9, 598–612. [Google Scholar] [CrossRef] [Green Version]
- Suriyaprom, S.; Mosoni, P.; Leroy, S.; Kaewkod, T.; Desvaux, M.; Tragoolpua, Y. Antioxidants of Fruit Extracts as Antimicrobial Agents against Pathogenic Bacteria. Antioxidants 2022, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.; Mousa, O.; El-Fitiany, R.; El Gedaily, R. Cytotoxic, antimicrobial activities, and phytochemical investigation of three peach cultivars and acerola leaves. J. Rep. Pharm. Sci. 2020, 9, 221–236. [Google Scholar] [CrossRef]
- Raudone, L.; Vilkickyte, G.; Pitkauskaite, L.; Raudonis, R.; Vainoriene, R.; Motiekaityte, V. Antioxidant Activities of Vaccinium vitis-idaea L. Leaves within Cultivars and Their Phenolic Compounds. Molecules 2019, 24, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaylova, D.; Popova, A.; Vrancheva, R.; Dincheva, I. HS-SPME-GC-MS Volatile Profile Characterization of Peach (Prunus persica L. Batsch) Varieties Grown in the Eastern Balkan Peninsula. Plants 2022, 11, 166. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RI | Polar Metabolites | F | G | U | L | JL | FQ | Evm | M | |
|---|---|---|---|---|---|---|---|---|---|---|
| Amino acids | ||||||||||
| 1 | 1097 | Alanine | 0.705 | 0.104 | 0.209 | 0.171 | 0.011 | 0.228 | 0.041 | 0.086 |
| 2 | 1208 | Valine | 0.142 | 0.307 | 0.549 | 0.079 | 0.014 | 0.123 | 0.027 | 0.052 |
| 3 | 1266 | Leucine | ND | 0.052 | 0.082 | 0.007 | ND | 0.010 | ND | 0.035 |
| 4 | 1285 | Isoleucine | 0.119 | 0.265 | 0.435 | 0.117 | 0.009 | 0.034 | 0.207 | 0.143 |
| 5 | 1293 | Proline | 0.161 | 0.297 | 0.525 | ND | 0.008 | 0.080 | ND | 0.115 |
| 6 | 1299 | Gycine | 0.224 | 0.079 | 0.177 | 0.059 | ND | 0.137 | 0.078 | 0.391 |
| 7 | 1351 | Serine | 0.591 | 0.618 | 1.240 | 0.438 | 0.028 | 0.359 | 0.711 | 0.239 |
| 8 | 1376 | Threonine | 0.201 | 0.932 | 0.777 | 0.600 | 0.011 | 0.477 | 0.178 | 0.417 |
| 9 | 1508 | Aspartic acid | 1.141 | ND | ND | 0.068 | ND | ND | ND | ND |
| 10 | 1515 | Methionine | 0.050 | 0.401 | 0.124 | 0.191 | 0.007 | 0.142 | 0.035 | 0.036 |
| 11 | 1550 | Cysteine | 0.007 | 0.014 | 0.024 | 0.009 | ND | 0.005 | 0.011 | 0.020 |
| 12 | 1609 | Glutamic acid | 0.280 | 0.522 | 0.860 | 0.175 | 0.051 | 0.137 | 0.389 | 0.217 |
| 13 | 1635 | Phenyalanine | 0.215 | 0.283 | 0.467 | 0.106 | 0.033 | 0.205 | 0.058 | 0.151 |
| 14 | 1833 | Arginine | 0.171 | 0.376 | 0.710 | 0.142 | 0.016 | 0.416 | 0.211 | 0.273 |
| 15 | 1910 | Lysine | 0.184 | 0.857 | 1.072 | 0.243 | 0.015 | 0.247 | 0.563 | 0.352 |
| 16 | 1930 | Tyrosine | 0.156 | 0.438 | 0.669 | ND | 0.012 | 0.028 | 0.055 | 0.015 |
| 17 | 2144 | Histidine | 0.384 | 0.425 | 0.827 | 0.203 | ND | ND | 0.287 | 0.122 |
| 18 | 2211 | Tryptophan | ND | 0.148 | 0.076 | ND | 0.068 | 0.046 | ND | 0.044 |
| Total | 4.729 | 6.117 | 8.825 | 2.607 | 0.283 | 2.673 | 2.851 | 2.709 | ||
| Organic acids | ||||||||||
| 1 | 1305 | Succinic acid | 0.728 | 0.547 | 0.783 | 0.526 | 0.619 | 0.602 | 0.642 | 0.662 |
| 2 | 1344 | Fumaric acid | 0.548 | 0.412 | 0.589 | 0.396 | 0.466 | 0.453 | 0.483 | 0.498 |
| 3 | 1477 | Malic acid | 1.012 | 0.760 | 1.088 | 0.731 | 0.860 | 0.836 | 0.892 | 0.920 |
| 4 | 1818 | Shikimic acid | 1.662 | 1.249 | 1.787 | 1.201 | 1.413 | 1.374 | 1.465 | 1.511 |
| 5 | 1841 | Citric acid | 0.849 | 0.638 | 0.913 | 0.613 | 0.722 | 0.702 | 0.748 | 0.772 |
| 6 | 1855 | Quinic acid | 0.825 | 0.620 | 0.887 | 0.596 | 0.701 | 0.682 | 0.727 | 0.750 |
| 7 | 1946 | L-Ascorbic acid | 0.287 | 0.216 | 0.309 | 0.207 | 0.244 | 0.237 | 0.253 | 0.261 |
| Total | 5.911 | 4.441 | 6.357 | 4.271 | 5.025 | 4.885 | 5.211 | 5.374 | ||
| Sugar alcohols | ||||||||||
| 1 | 1932 | Sorbitol | 0.326 | 0.245 | 0.232 | 0.197 | 0.182 | 0.269 | 0.281 | 0.296 |
| 2 | 2034 | Myo-inositol | 0.187 | 0.141 | 0.104 | 0.127 | 0.091 | 0.187 | 0.157 | 0.170 |
| Saccharides (mono-, di-) | ||||||||||
| 1 | 1856 | Fructose isomer | 0.972 | 1.355 | 1.588 | 1.984 | 1.804 | 1.491 | 1.389 | 1.640 |
| 1865 | Fructose isomer | 0.337 | 0.470 | 0.550 | 0.688 | 0.625 | 0.517 | 0.481 | 0.568 | |
| 2 | 1881 | Glucose issomer | 1.855 | 2.586 | 3.029 | 3.786 | 3.442 | 2.844 | 2.650 | 3.129 |
| 1901 | Glucose issomer | 1.397 | 1.947 | 2.281 | 2.851 | 2.592 | 2.142 | 1.996 | 2.356 | |
| 3 | 2620 | Sucrose isomer (alpha-D-Glc-(1.2)-beta-D-Fru) | 2.992 | 4.171 | 4.885 | 6.106 | 5.551 | 4.588 | 4.274 | 5.047 |
| 2833 | Sucrose isomer (alpha-D-Glc-(1.2)-beta-D-Fru) | 1.737 | 2.421 | 2.836 | 3.545 | 3.223 | 2.663 | 2.481 | 2.930 | |
| Total | 9.290 | 12.950 | 15.168 | 18.960 | 17.236 | 14.245 | 13.272 | 15.669 | ||
| Saturated and unsaturated fatty acids | ||||||||||
| 1 | 1719 | Tetradecanoic acid (Myristic acid) | 0.336 | 0.319 | 0.166 | 0.460 | 0.394 | 0.259 | 0.276 | 0.287 |
| 2 | 1926 | n-Hexadecanoic acid (Palmitic acid) | 3.953 | 3.754 | 1.948 | 5.411 | 4.625 | 3.040 | 3.246 | 3.378 |
| 3 | 2095 | 9,12-(Z,E)-Octadecadienoic acid (Linoleic acid) | 2.451 | 2.327 | 1.208 | 3.355 | 2.867 | 1.885 | 2.013 | 2.095 |
| 4 | 2099 | 9-(Z)-Octadecenoic acid (Oleic acid) | 1.285 | 1.220 | 0.633 | 1.758 | 1.503 | 0.988 | 1.055 | 1.098 |
| 5 | 2103 | 9,12,15-(Z,Z,Z)-Octadecatrienoic acid (Linolenic acid) | 0.445 | 0.423 | 0.219 | 0.609 | 0.521 | 0.342 | 0.366 | 0.380 |
| 6 | 2247 | n-Octadecanoic acid (Stearic acid) | 1.648 | 1.565 | 0.812 | 2.256 | 1.928 | 1.268 | 1.354 | 1.409 |
| 7 | 2311 | n-Eicosanoic acid (Arahydic acid) | 0.756 | 0.718 | 0.372 | 1.034 | 0.884 | 0.581 | 0.621 | 0.646 |
| 8 | 2408 | n-Docosanoic acid (Behenic acid) | 0.939 | 0.891 | 0.463 | 1.285 | 1.098 | 0.722 | 0.771 | 0.802 |
| Total | 11.811 | 11.217 | 5.821 | 16.168 | 13.819 | 9.086 | 9.701 | 10.095 | ||
| Phenolic acids | ||||||||||
| 1 | 1836 | Protocatechuic acid | 0.073 | 0.187 | 0.033 | 0.107 | 0.042 | 0.123 | 0.136 | 0.273 |
| 2 | 1945 | trans-p-Coumaric acid | 0.157 | 0.142 | 0.122 | 0.096 | ND | 0.083 | 0.096 | 0.185 |
| 3 | 2103 | trans-Ferulic acid | 0.110 | 0.092 | 0.055 | 0.077 | 0.064 | 0.137 | 0.026 | 0.055 |
| 4 | 2140 | trans-Caffeic acid | 0.034 | 0.128 | 0.045 | 0.068 | 0.085 | 0.044 | 0.053 | 0.026 |
| 5 | 2254 | trans-Sinapic acid | 0.072 | 0.062 | 0.061 | 0.050 | 0.022 | 0.031 | 0.046 | 0.055 |
| 6 | 3191 | Chlorogenic acid | 2.816 | 3.559 | 1.638 | 0.842 | 2.502 | 2.150 | 1.214 | 1.363 |
| Total | 3.263 | 4.170 | 1.955 | 1.240 | 2.714 | 2.569 | 1.572 | 1.957 | ||
| Samples | α-Glucosidase | Lipase | α-Amylase | AChE | |
|---|---|---|---|---|---|
| G | Free | - | - | - | - |
| Acid hydrolysis | - | - | - | - | |
| Alkaline hydrolysis | 3.4 ± 0.11 k | - | 26.67 * | 26.67 * | |
| F | Free | - | - | - | - |
| Acid hydrolysis | - | - | 31.51 * | - | |
| Alkaline hydrolysis | 9.0 ± 0.13 h | - | 20.55 ± 0.51 a | 31.1 ± 0.22 a | |
| U | Free | 39.7 ± 0.05 b | - | - | - |
| Acid hydrolysis | 22.9 ± 0.14 d | - | - | - | |
| Alkaline hydrolysis | 5.9 ± 0.04 i | - | 15.17 * | 15.17 * | |
| JL | Free | - | - | - | - |
| Acid hydrolysis | 26.1 ± 0.54 c | - | - | 21.3 * | |
| Alkaline hydrolysis | 5.1 ± 0.13 j | - | 17.38 ± 0.11 b | 17.8 ± 0.52 b | |
| L | Free | 27.2 ± 0.11 c | - | - | - |
| Acid hydrolysis | 14.1 ± 0.09 f | - | 31.51 * | - | |
| Alkaline hydrolysis | 9.0 ± 0.10 h | - | - | 30.23 * | |
| FQ | Free | 49.3 ± 0.22 a | - | - | - |
| Acid hydrolysis | - | - | - | - | |
| Alkaline hydrolysis | 3.1 ± 0.10 k | - | - | - | |
| Evm | Free | 19.4 ± 0.12 e | - | - | 56.36 * |
| Acid hydrolysis | - | - | - | - | |
| Alkaline hydrolysis | 2.7 ± 0.08 l | - | - | - | |
| M | Free | 11.6 ± 0.14 g | - | - | 55.67 * |
| Acid hydrolysis | - | - | - | - | |
| Alkaline hydrolysis | 2.6 ± 0.08 l | - | - | - | |
| Test Microorganism/ Samples | Sample (Inhibition Zone, mm) * | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G | F | U | JK | L | FQ | Evm | M | G | F | U | JK | L | FQ | Evm | M | G | F | U | JK | L | FQ | Evm | M | |
| Free | Acid Hydrolysis | Alkaline Hydrolysis | ||||||||||||||||||||||
| Gram (+) bacteria | ||||||||||||||||||||||||
| Bacillus subtilis ATCC 6633 | - | - | - | 8 | - | - | 8 | 8 | 9 | 10 | 10 | 10 | 10 | 10 | 10 | 9 | 10 | 11 | 9 | 9 | 10 | 10 | 9 | 10 |
| Staphylococcus aureus ATCC 25923 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | 9 | 8 | 8 | - | - | - | - |
| Listeria monocytogenes NBIMCC 8632 | - | - | - | - | - | - | - | - | 8 | 8 | 8 | 8 | 8 | 9 | 9 | 8 | 8 | 9 | 8 | 9 | 9 | - | - | - |
| Enterococcus faecalis ATCC 19433 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 11 | - | 8 | - | - | - | - |
| Gram (−) bacteria | ||||||||||||||||||||||||
| Salmonella enteritidis ATCC 13076 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 9 | - | 8 | - | - | - | - |
| Escherichia coli ATCC 8739 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Proteus vulgaris ATCC 6380 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | - | 8 | - | - | - | - |
| Pseudomonas aeruginosa ATCC 9027 | 11 | 11 | 10 | 12 | 12 | 9 | 9 | - | 12 | 12 | 12 | 15 | 15 | 9 | 9 | - | 13 | 14 | 13 | 13 | 13 | 8 | - | 9 |
| Yeasts | ||||||||||||||||||||||||
| Candida albicans NBIMCC 74 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | 8 | - | - | - | 9 | 9 | 8 |
| Saccharomyces cerevisiae ATCC 9763 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | 8 | 8 | 8 | - | - | - | 8 | 8 | 8 |
| Fungi | ||||||||||||||||||||||||
| Aspergillus niger ATCC 1015 | - | - | - | - | - | - | - | - | 8 | 8 | 8 | 8 | 8 | 8 | 9 | 9 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| Aspergillus flavus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | 8 | 8 | 8 | 8 | 8 | - | - |
| Penicillium sp. | - | - | - | - | - | - | - | - | 9 | 9 | 8 | 9 | 9 | 8 | 8 | 8 | 9 | 9 | 9 | 9 | 9 | - | 8 | - |
| Rhizopus sp. | 10 | 10 | - | 10 | 10 | - | - | - | 8 | 8 | - | 10 | 10 | - | 10 | 10 | 9 | 9 | - | - | - | - | 8 | - |
| Fusarium moniliforme ATCC 38932 | - | - | - | - | - | - | - | - | 8 | 8 | 8 | 9 | 9 | 8 | 8 | 8 | 10 | 10 | 10 | 10 | 10 | - | - | - |
| Mucor sp. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Variables | TPC | TFC | TMA | DPPH | ABTS | FRAP | CUPRAC |
|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.3249 *** | 0.5957 * | 0.8856 * | 0.5059 * | 0.8231 * | 0.6251 * |
| TFC | 1 | 0.0608 *** | 0.5701 * | 0.1461 *** | 0.7241 * | 0.5234 * | |
| TMA | 1 | 0.6286 * | 0.5618 * | 0.4695 * | 0.3833 * | ||
| DPPH | 1 | 0.4303 * | 0.9562 * | 0.7453 * | |||
| ABTS | 1 | 0.2979 ** | 0.2376 ** | ||||
| FRAP | 1 | 0.7155 * | |||||
| CUPRAC | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihaylova, D.; Popova, A.; Desseva, I.; Dincheva, I.; Tumbarski, Y. Valorization of Peels of Eight Peach Varieties: GC–MS Profile, Free and Bound Phenolics and Corresponding Biological Activities. Antioxidants 2023, 12, 205. https://doi.org/10.3390/antiox12010205
Mihaylova D, Popova A, Desseva I, Dincheva I, Tumbarski Y. Valorization of Peels of Eight Peach Varieties: GC–MS Profile, Free and Bound Phenolics and Corresponding Biological Activities. Antioxidants. 2023; 12(1):205. https://doi.org/10.3390/antiox12010205
Chicago/Turabian StyleMihaylova, Dasha, Aneta Popova, Ivelina Desseva, Ivayla Dincheva, and Yulian Tumbarski. 2023. "Valorization of Peels of Eight Peach Varieties: GC–MS Profile, Free and Bound Phenolics and Corresponding Biological Activities" Antioxidants 12, no. 1: 205. https://doi.org/10.3390/antiox12010205