
Anmyungambi Decoction Ameliorates Obesity through Activation of Non-Shivering Thermogenesis in Brown and White Adipose Tissues
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of AMGB Decoctions
2.2. Animal Study
2.3. Serum Analysis
2.4. Hematoxylin and Eosin (H&E) Staining
2.5. Western Blot Assay
2.6. Blood Glucose Measurement
2.7. FFA Level
Catalase Activity
2.8. Statistical Analysis
3. Results
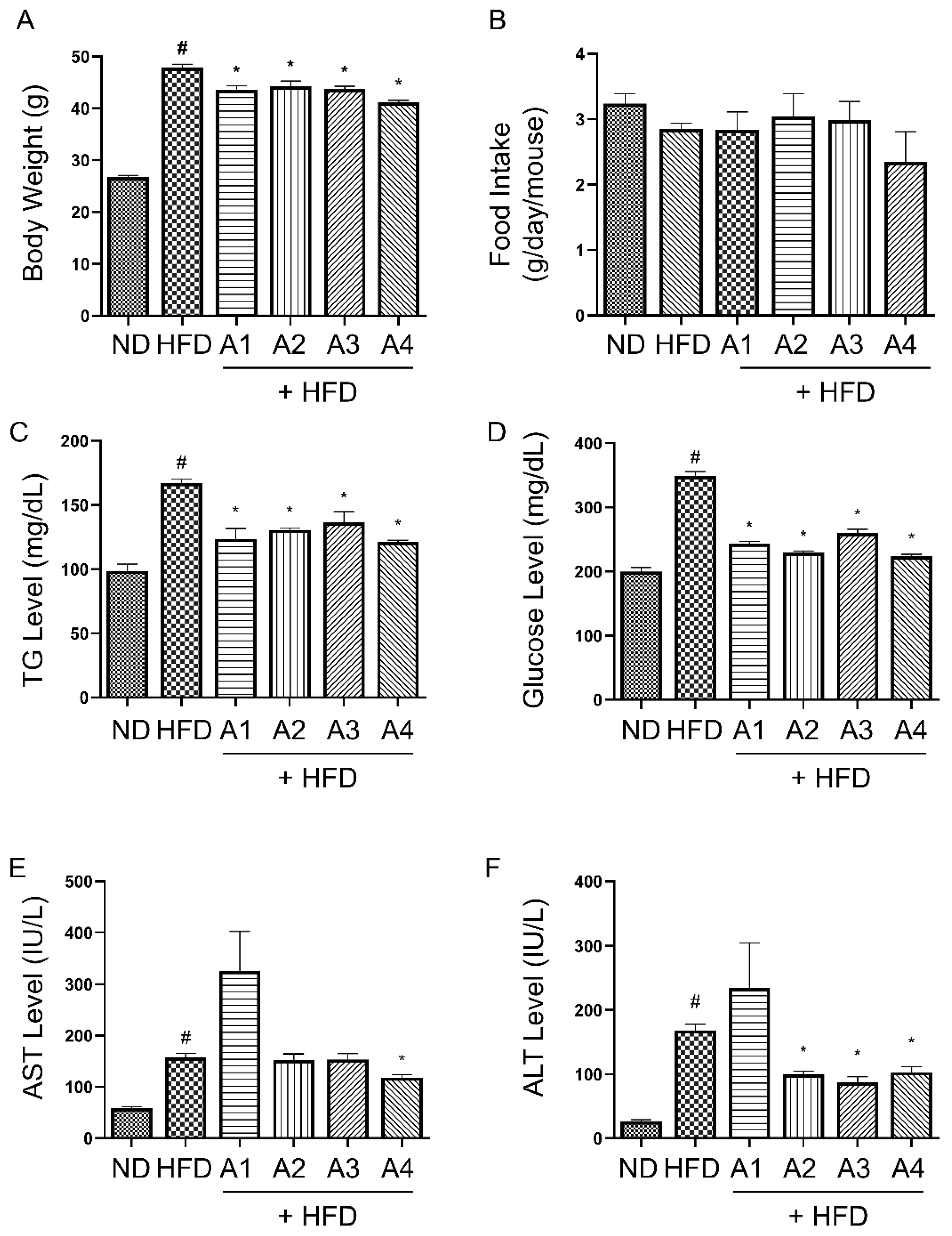
3.1. Various Versions of AMGB Decoction Inhibit Body Weight Gain and Obesity-Related Serum Parameters in HFD-Induced Obese Mice
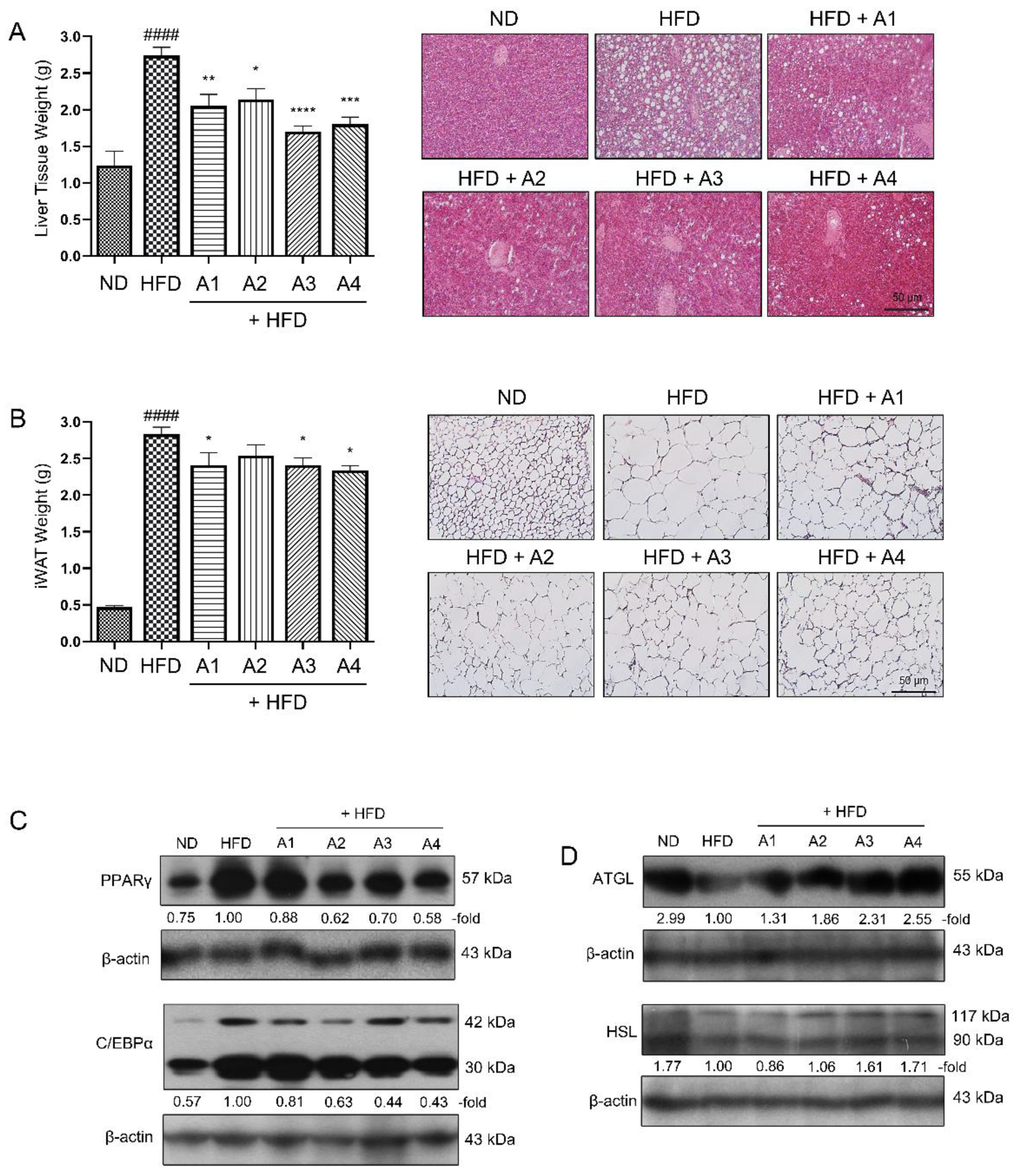
3.2. Various Versions of AMGB Decoction Suppress Adipogenesis and Increase Lipolysis in HFD-Induced Obese Mice
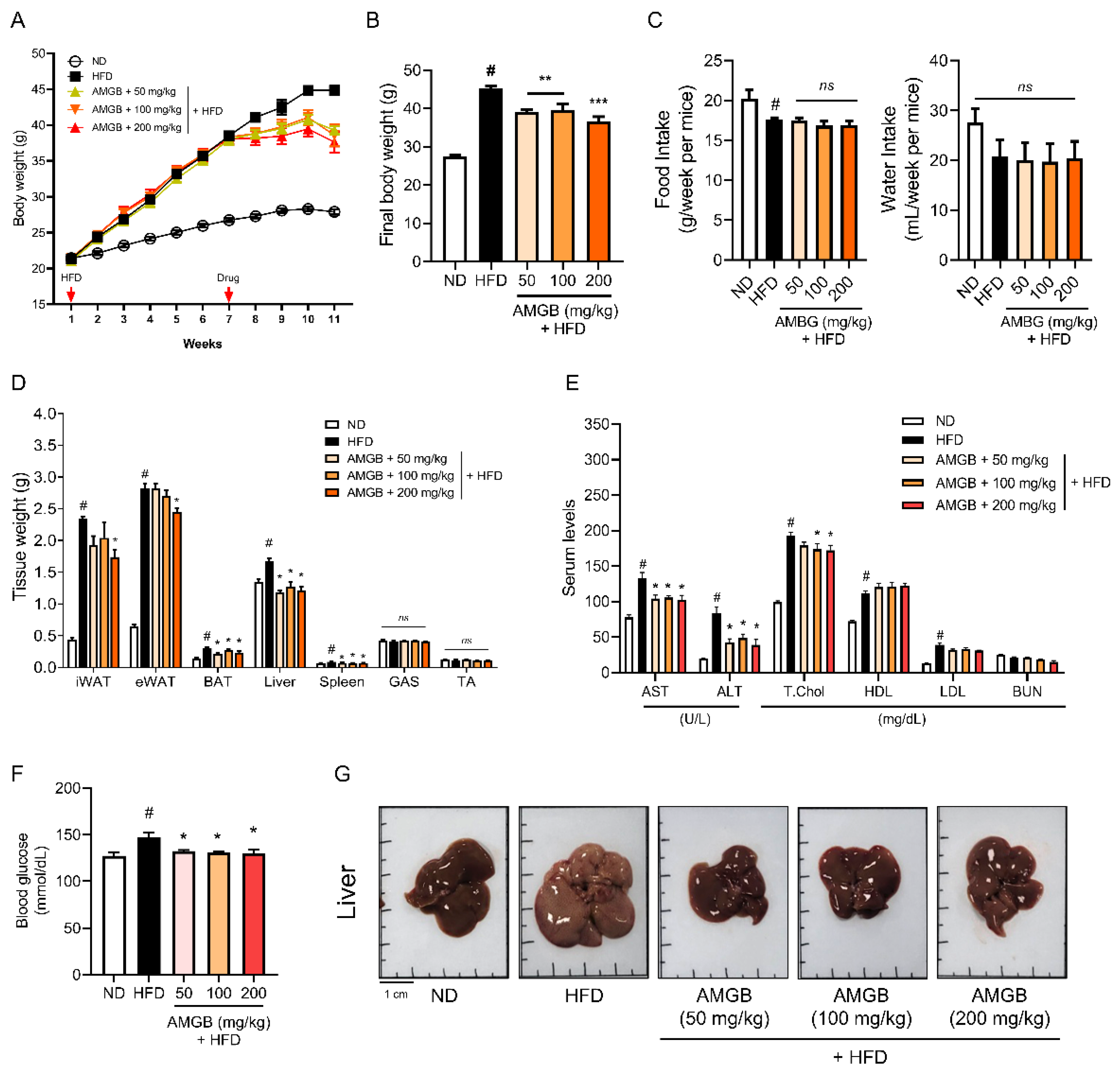
3.3. AMGB-4 Decoction Alleviates HFD-Induced Obesity and Hepatic Steatosis in Mice
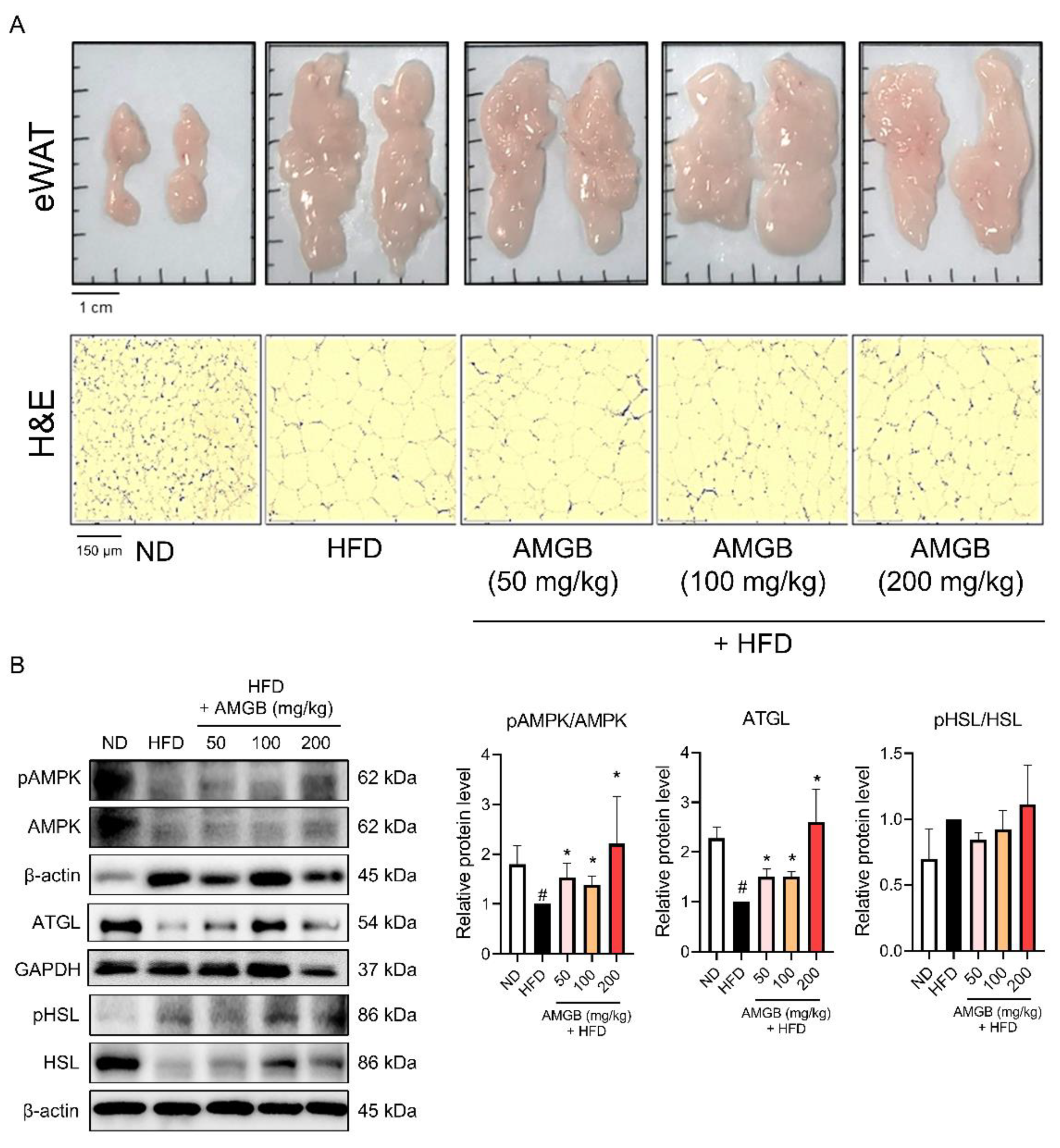
3.4. AMGB-4 Decoction Induces Lipolysis in the eWAT of HFD-Induced Obese Mice
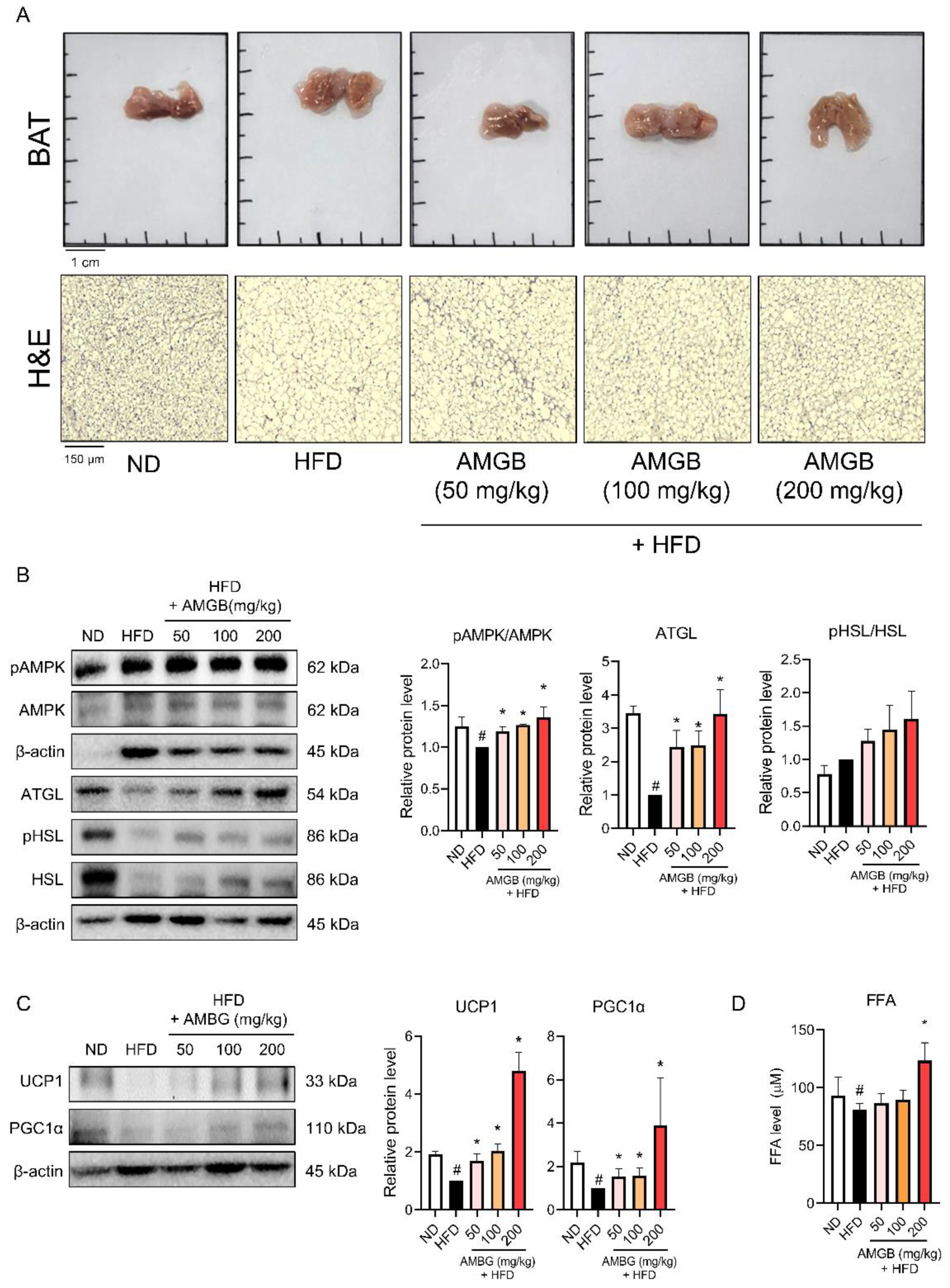
3.5. AMGB-4 Decoction Induces Non-Shivering Thermogenesis in the BAT of HFD-Induced Obese Mice
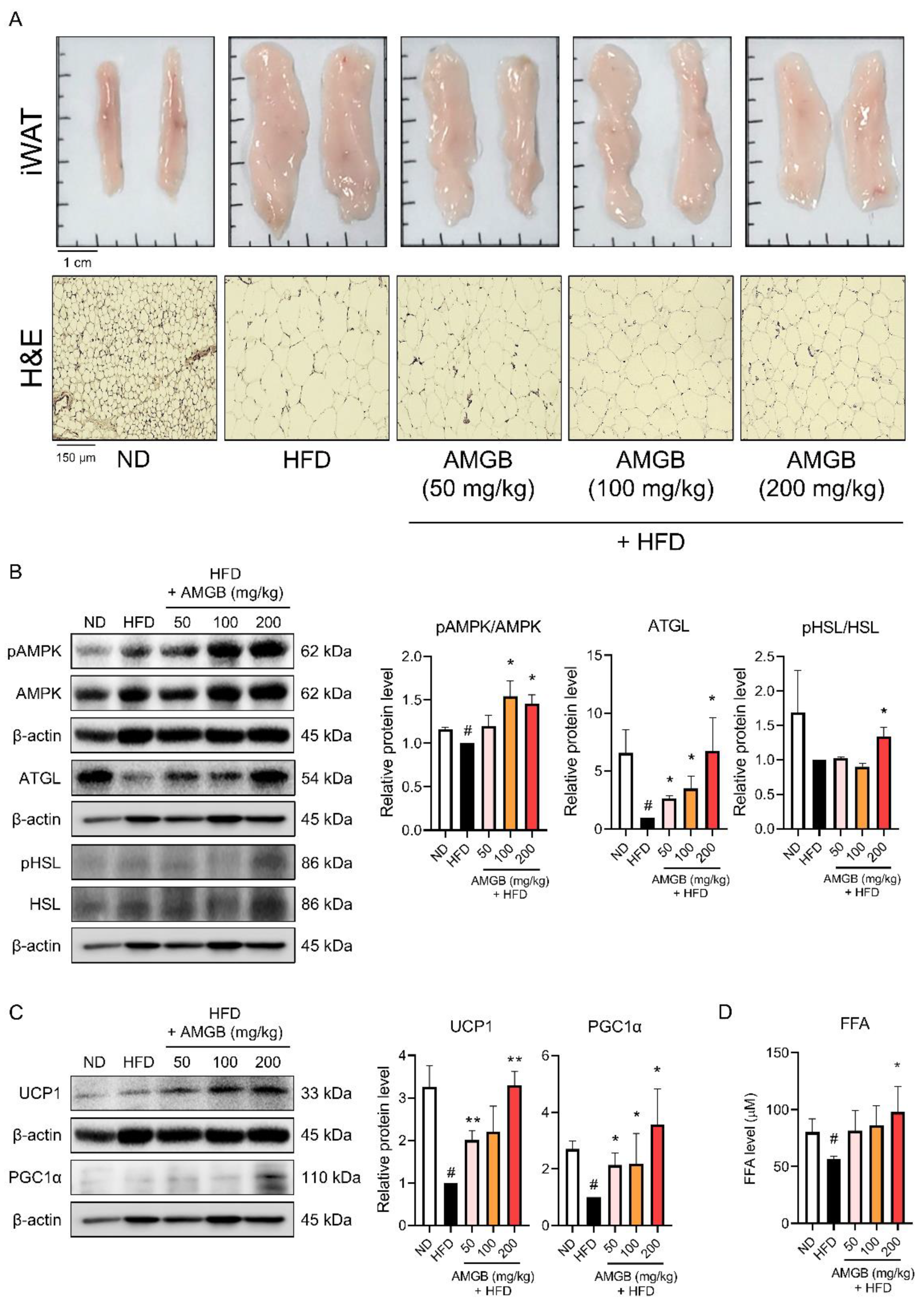
3.6. AMGB-4 Decoction Induces Non-Shivering Thermogenesis in the iWAT of HFD-Induced Obese Mice
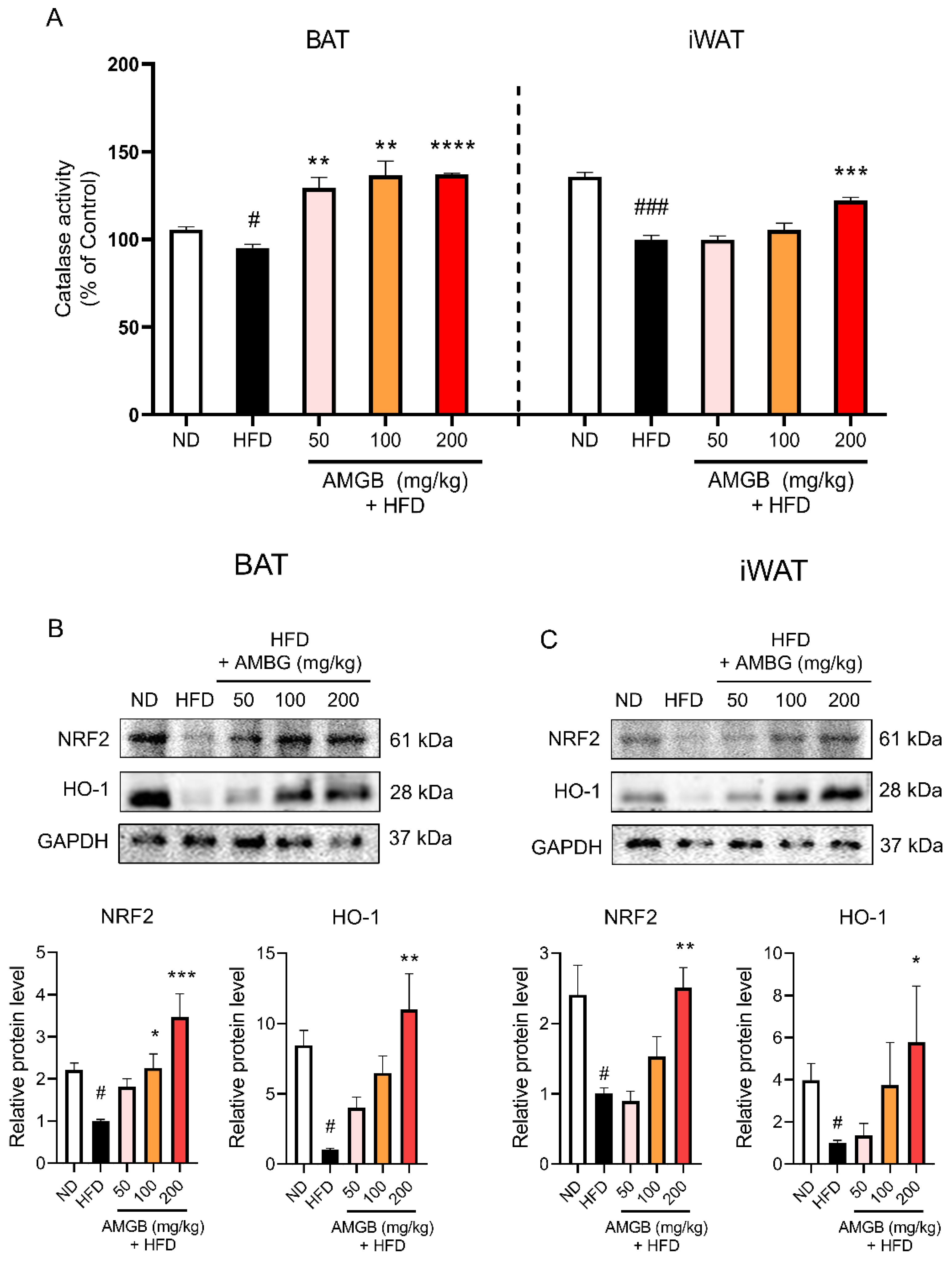
3.7. AMGB-4 Decoction Reduces Pathologic Oxidative Stress by Increasing The Nuclear Factor Erythroid 2–Related Factor 2 (NRF2)-Heme Oxygenase-1 (HO-1) Axis in iWAT and BAT of HFD-Induced Obese Mice
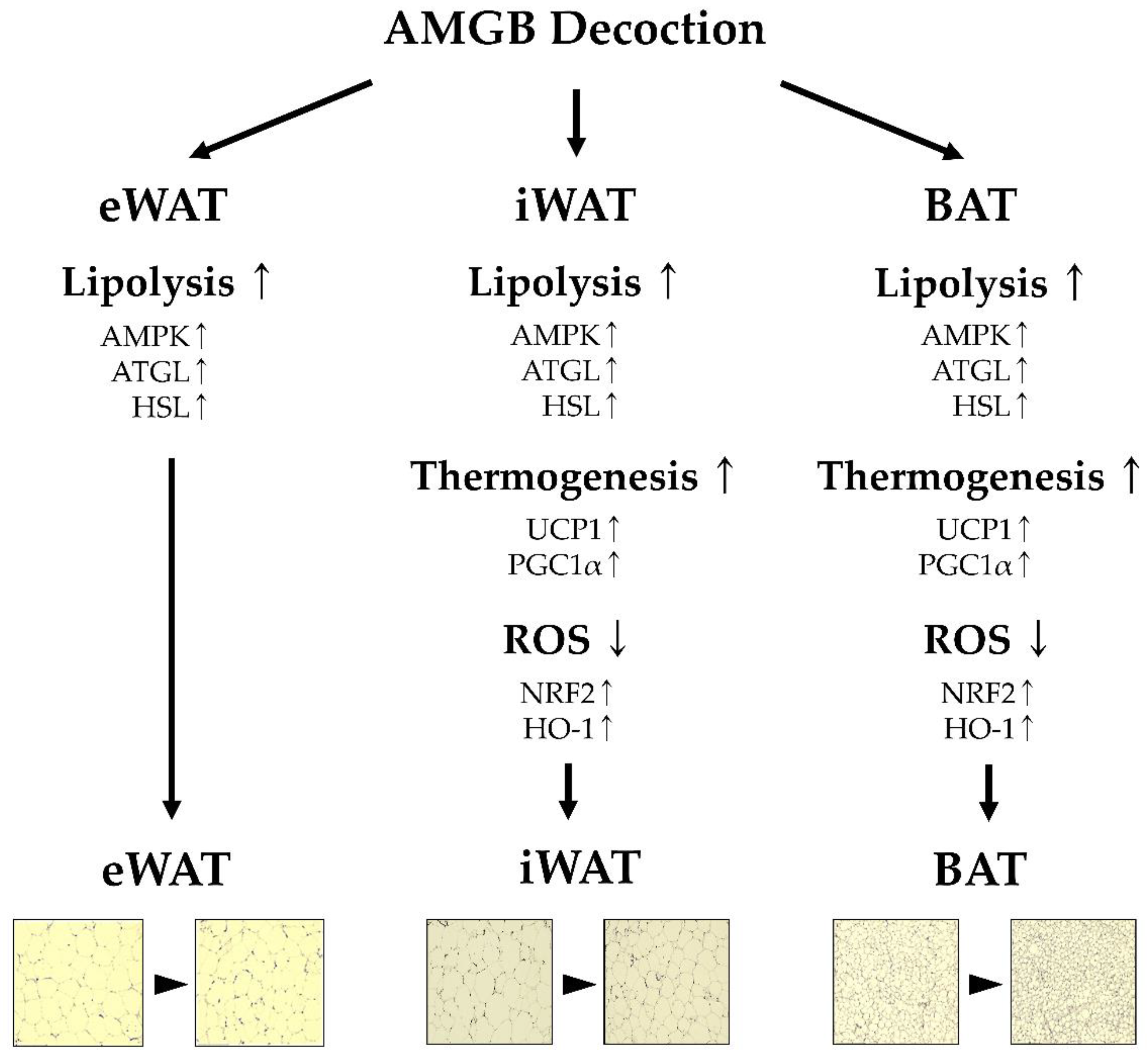
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barnes, A.S. The epidemic of obesity and diabetes: Trends and treatments. Tex. Heart Inst. J. 2011, 38, 142–144. [Google Scholar] [PubMed]
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Després, J.P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef] [PubMed]
- De Pergola, G.; Silvestris, F. Obesity as a major risk factor for cancer. J. Obes. 2013, 2013, 291546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Fact Sheets: Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 19 November 2022).
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected evidence for active brown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Arch, J.R. New Physiological Aspects of Brown Adipose Tissue. Curr. Obes. Rep. 2014, 3, 414–421. [Google Scholar] [CrossRef]
- Li, Y.; Fromme, T. Uncoupling Protein 1 Does Not Produce Heat without Activation. Int. J. Mol. Sci. 2022, 23, 2406. [Google Scholar] [CrossRef]
- Ouellet, V.; Labbé, S.M.; Blondin, D.P.; Phoenix, S.; Guérin, B.; Haman, F.; Turcotte, E.E.; Richard, D.; Carpentier, A.C. Brown adipose tissue oxidative metabolism contributes to energy expenditure during acute cold exposure in humans. J. Clin. Investig. 2012, 122, 545–552. [Google Scholar] [CrossRef]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fromme, T.; Schweizer, S.; Schöttl, T.; Klingenspor, M. Taking control over intracellular fatty acid levels is essential for the analysis of thermogenic function in cultured primary brown and brite/beige adipocytes. EMBO Rep. 2014, 15, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef]
- Himms-Hagen, J.; Cui, J.; Danforth, E., Jr.; Taatjes, D.J.; Lang, S.S.; Waters, B.L.; Claus, T.H. Effect of CL-316,243, a thermogenic beta 3-agonist, on energy balance and brown and white adipose tissues in rats. Am. J. Physiol. 1994, 266, R1371–R1382. [Google Scholar] [CrossRef]
- Baba, S.; Tatsumi, M.; Ishimori, T.; Lilien, D.L.; Engles, J.M.; Wahl, R.L. Effect of nicotine and ephedrine on the accumulation of 18F-FDG in brown adipose tissue. J. Nucl. Med. 2007, 48, 981–986. [Google Scholar] [CrossRef] [Green Version]
- Bonet, M.L.; Oliver, P.; Palou, A. Pharmacological and nutritional agents promoting browning of white adipose tissue. Biochim. Biophys. Acta 2013, 1831, 969–985. [Google Scholar] [CrossRef]
- Song, N.J.; Chang, S.H.; Li, D.Y.; Villanueva, C.J.; Park, K.W. Induction of thermogenic adipocytes: Molecular targets and thermogenic small molecules. Exp. Mol. Med. 2017, 49, e3532017. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.C.; Hsieh, P.S. The Role and Regulatory Mechanism of Brown Adipose Tissue Activation in Diet-Induced Thermogenesis in Health and Diseases. Int. J. Mol. Sci. 2022, 23, 9448. [Google Scholar] [CrossRef]
- Horvath, C.; Wolfrum, C. Feeding brown fat: Dietary phytochemicals targeting non-shivering thermogenesis to control body weight. Proc. Nutr. Soc. 2020, 79, 338–356. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Yoon, S.H.; Kim, H.; Kim, Y.D.; Leem, J.; Park, J. Ephedrae Herba in combination with herbal medicine (Zhizichi decoction and Phellodendri Cortex) for weight reduction: A case series. Integr. Med. Res. 2020, 9, 100408. [Google Scholar] [CrossRef]
- Jang, D.; Jeong, H.; Kim, C.E.; Leem, J. A System-Level Mechanism of Anmyungambi Decoction for Obesity: A Network Pharmacological Approach. Biomolecules 2021, 11, 1881. [Google Scholar] [CrossRef]
- Park, W.Y.; Song, G.; Park, J.Y.; Kim, K.I.; Ahn, K.S.; Kwak, H.J.; Leem, J.; Um, J.Y.; Park, J. Fruit of Gardenia jasminoides Induces Mitochondrial Activation and Non-Shivering Thermogenesis through Regulation of PPARγ. Antioxidants 2021, 10, 1418. [Google Scholar] [CrossRef]
- Kang, J.; Park, J.; Park, W.Y.; Jiao, W.; Lee, S.; Jung, Y.; Youn, D.H.; Song, G.; Cho, S.Y.; Kim, W.Y.; et al. A phytoestrogen secoisolariciresinol diglucoside induces browning of white adipose tissue and activates non-shivering thermogenesis through AMPK pathway. Pharmacol. Res. 2020, 158, 104852. [Google Scholar] [CrossRef]
- Ahmadian, M.; Duncan, R.E.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Triacylglycerol metabolism in adipose tissue. Future Lipidol. 2007, 2, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joe, A.W.; Yi, L.; Even, Y.; Vogl, A.W.; Rossi, F.M. Depot-specific differences in adipogenic progenitor abundance and proliferative response to high-fat diet. Stem. Cells 2009, 27, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, C.; Feng, J.; Chen, J.; Jiang, Q.; Kuang, S.; Wang, Y. Depot-specific differences in fat mass expansion in WT and ob/ob mice. Oncotarget 2017, 8, 46326–46336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The Subtle Balance between Lipolysis and Lipogenesis: A Critical Point in Metabolic Homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase To Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol. Cell Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Frayn, K.N.; Karpe, F.; Fielding, B.A.; Macdonald, I.A.; Coppack, S.W. Integrative physiology of human adipose tissue. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.; Chen, Y. Management of Oxidative Stress: Crosstalk Between Brown/Beige Adipose Tissues and Skeletal Muscles. Front. Physiol. 2021, 12, 712372. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.H.; Jang, J.; Oh, S.; Yoon, J.H.; Jo, D.G.; Yun, U.J.; Park, K.W. Nrf2 induces Ucp1 expression in adipocytes in response to β3-AR stimulation and enhances oxygen consumption in high-fat diet-fed obese mice. BMB Rep. 2021, 54, 419–424. [Google Scholar] [CrossRef]
- Jegal, K.H.; Ko, M.M.; Kim, B.Y.; Son, M.J.; Kim, S. A national survey on current clinical practice pattern of Korean Medicine doctors for treating obesity. PLoS ONE 2022, 17, e0266034. [Google Scholar] [CrossRef]
- Takemoto, H.; Takahashi, J.; Hyuga, S.; Odaguchi, H.; Uchiyama, N.; Maruyama, T.; Yamashita, T.; Hyuga, M.; Oshima, N.; Amakura, Y.; et al. Ephedrine Alkaloids-Free Ephedra Herb Extract, EFE, Has No Adverse Effects Such as Excitation, Insomnia, and Arrhythmias. Biol. Pharm. Bull. 2018, 41, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.L.; Zhang, X.; Liu, W.J.; Li, M.; Zhang, Y.H. Suan zao ren tang in combination with zhi zi chi tang as a treatment protocol for insomniacs with anxiety: A randomized parallel-controlled trial. Evid. Based Complement Alternat Med. 2015, 2015, 913252. [Google Scholar] [CrossRef] [Green Version]
- Steinmann, D.; Baumgartner, R.R.; Heiss, E.H.; Bartenstein, S.; Atanasov, A.G.; Dirsch, V.M.; Ganzera, M.; Stuppner, H. Bioguided isolation of (9Z)-octadec-9-enoic acid from Phellodendron amurense Rupr. and identification of fatty acids as PTP1B inhibitors. Planta Med. 2012, 78, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Kang, B.; Youn, B.Y.; Kim, K.I.; Park, J.; Leem, J. Comprehensive traditional East Asian medicine treatment strategy for obesity considering the therapeutic effects and adverse events: A protocol for systematic review and network meta-analysis. Medicine (Baltimore) 2022, 101, e28673. [Google Scholar] [CrossRef]
- Manzoni, M.S.; Rossi, E.A.; Carlos, I.Z.; Vendramini, R.C.; Duarte, A.C.; Dâmaso, A.R. Fermented soy product supplemented with isoflavones affected fat depots in juvenile rats. Nutrition 2005, 21, 1018–1024. [Google Scholar] [CrossRef]
- Choi, I.; Kim, Y.; Park, Y.; Seog, H.; Choi, H. Anti-obesity activities of fermented soygerm isoflavones by Bifidobacterium breve. Biofactors 2007, 29, 105–112. [Google Scholar] [CrossRef]
- Kwak, C.S.; Park, S.C.; Song, K.Y. Doenjang, a fermented soybean paste, decreased visceral fat accumulation and adipocyte size in rats fed with high fat diet more effectively than nonfermented soybeans. J. Med. Food 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Oh, H.G.; Kang, Y.R.; Lee, H.Y.; Kim, J.H.; Shin, E.H.; Lee, B.G.; Park, S.H.; Moon, D.I.; Kim, O.J.; Lee, I.A.; et al. Ameliorative effects of Monascus pilosus-fermented black soybean (Glycine max L. Merrill) on high-fat diet-induced obesity. J. Med. Food 2014, 17, 972–978. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kim, B.; Park, H.; Ji, Y.; Holzapfel, W.; Kim, D.Y.; Hyun, C.K. Long-term fermented soybean paste improves metabolic parameters associated with non-alcoholic fatty liver disease and insulin resistance in high-fat diet-induced obese mice. Biochem. Biophys. Res. Commun. 2018, 495, 1744–1751. [Google Scholar] [CrossRef]
- Kushida, M.; Okouchi, R.; Iwagaki, Y.; Asano, M.; Du, M.X.; Yamamoto, K.; Tsuduki, T. Fermented Soybean Suppresses Visceral Fat Accumulation in Mice. Mol. Nutr. Food Res. 2018, 62, e1701054. [Google Scholar] [CrossRef]
- Cha, Y.S.; Park, Y.; Lee, M.; Chae, S.W.; Park, K.; Kim, Y.; Lee, H.S. Doenjang, a Korean fermented soy food, exerts antiobesity and antioxidative activities in overweight subjects with the PPAR-γ2 C1431T polymorphism: 12-week, double-blind randomized clinical trial. J. Med. Food. 2014, 17, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Li, S.; Wang, S.; Ho, C.T. Chemistry and bioactivity of Gardenia jasminoides. J. Food Drug Anal. 2017, 25, 43–61. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.S.; Huh, Y.S. Effect of intake of gardenia fruits and combined exercise of middle-aged obese women on hormones regulating energy metabolism. J. Exerc. Nutr. Biochem. 2014, 18, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, K.; Shimada, T.; Nagareda, Y.; Watanabe, M.; Ishizaki, J.; Sai, Y.; Miyamoto, K.; Aburada, M. Preventive effect of geniposide on metabolic disease status in spontaneously obese type 2 diabetic mice and free fatty acid-treated HepG2 cells. Biol. Pharm. Bull. 2011, 34, 1613–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.G.; Kong, C.Y.; Song, P.; Zhang, X.; Yuan, Y.P.; Tang, Q.Z. Geniposide Protects against Obesity-Related Cardiac Injury through AMPKα- and Sirt1-Dependent Mechanisms. Oxid. Med. Cell Longev. 2018, 2018, 6053727. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Chen, K.; Feng, M.; Shao, W.; Wu, J.; Chen, K.; Liang, T.; Liu, C. Genipin alleviates high-fat diet-induced hyperlipidemia and hepatic lipid accumulation in mice via miR-142a-5p/SREBP-1c axis. Febs. J. 2018, 285, 501–517. [Google Scholar] [CrossRef]
- Guan, L.; Gong, D.; Yang, S.; Shen, N.; Zhang, S.; Li, Y.; Wu, Q.; Yuan, B.; Sun, Y.; Dai, N.; et al. Genipin ameliorates diet-induced obesity via promoting lipid mobilization and browning of white adipose tissue in rats. Phytother Res. 2018, 32, 723–732. [Google Scholar] [CrossRef]
- Wang, L.; Chen, G.; Wu, S.; Xu, Y.; Guo, C.; Wang, M.; Liang, T.; Guo, Z.; Di, H.J.; Hu, Z. Genipin improves lipid metabolism and sperm parametersin obese mice via regulation of miR-132 expression. Acta Biochim. Biophys. Sin. (Shanghai) 2022, 54, 1278–1288. [Google Scholar] [CrossRef]
- Wang, T.; Wang, Y.; Kontani, Y.; Kobayashi, Y.; Sato, Y.; Mori, N.; Yamashita, H. Evodiamine improves diet-induced obesity in a uncoupling protein-1-independent manner: Involvement of antiadipogenic mechanism and extracellularly regulated kinase/mitogen-activated protein kinase signaling. Endocrinology 2008, 149, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Fahmy, H.; Zjawiony, J.K.; Davies, G.E. Inhibitory effect and transcriptional impact of berberine and evodiamine on human white preadipocyte differentiation. Fitoterapia 2010, 81, 259–268. [Google Scholar] [CrossRef]
- Bak, E.J.; Park, H.G.; Kim, J.M.; Kim, J.M.; Yoo, Y.J.; Cha, J.H. Inhibitory effect of evodiamine alone and in combination with rosiglitazone on in vitro adipocyte differentiation and in vivo obesity related to diabetes. Int. J. Obes. (Lond.) 2010, 34, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Kusudo, T.; Takeuchi, T.; Yamashita, Y.; Kontani, Y.; Okamatsu, Y.; Saito, M.; Mori, N.; Yamashita, H. Evodiamine inhibits insulin-stimulated mTOR-S6K activation and IRS1 serine phosphorylation in adipocytes and improves glucose tolerance in obese/diabetic mice. PLoS ONE 2013, 8, e83264. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, H.; Li, B.; Meng, X.; Wang, J.; Zhang, Y.; Yao, S.; Ma, Q.; Jin, L.; Yang, J.; et al. Berberine activates thermogenesis in white and brown adipose tissue. Nat. Commun. 2014, 5, 5493. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Xia, M.; Duan, Y.; Zhang, L.; Jiang, H.; Hu, X.; Yan, H.; Zhang, Y.; Gu, Y.; Shi, H.; et al. Berberine promotes the recruitment and activation of brown adipose tissue in mice and humans. Cell Death Dis. 2019, 10, 468. [Google Scholar] [CrossRef] [Green Version]
- Boozer, C.N.; Daly, P.A.; Homel, P.; Solomon, J.L.; Blanchard, D.; Nasser, J.A.; Strauss, R.; Meredith, T. Herbal ephedra/caffeine for weight loss: A 6-month randomized safety and efficacy trial. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Shekelle, P.G.; Hardy, M.L.; Morton, S.C.; Maglione, M.; Mojica, W.A.; Suttorp, M.J.; Rhodes, S.L.; Jungvig, L.; Gagné, J. Efficacy and safety of ephedra and ephedrine for weight loss and athletic performance: A meta-analysis. JAMA 2003, 289, 1537–1545. [Google Scholar] [CrossRef] [Green Version]
- Song, M.K.; Um, J.Y.; Jang, H.J.; Lee, B.C. Beneficial effect of dietary Ephedra sinica on obesity and glucose intolerance in high-fat diet-fed mice. Exp. Ther. Med. 2012, 3, 707–712. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Lim, C.; Lim, S.; Lee, B.; Cho, S. Effect of Ephedrae Herba methanol extract on high-fat diet-induced hyperlipidaemic mice. Pharm. Biol. 2019, 57, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Kim, B.S.; Han, K.; Kim, H. Ephedra-Treated Donor-Derived Gut Microbiota Transplantation Ameliorates High Fat Diet-Induced Obesity in Rats. Int. J. Environ. Res. Public Health 2017, 14, 555. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Shon, D.H.; Ryu, Y.H.; Ko, Y. Extract of Ephedra sinica Stapf Induces Browning of Mouse and Human White Adipocytes. Foods 2022, 11, 1028. [Google Scholar] [CrossRef]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab. Syndr. Obes. 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Darlington, G.J.; Ross, S.E.; MacDougald, O.A. The role of C/EBP genes in adipocyte differentiation. J. Biol. Chem. 1998, 273, 30057–30060. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway. Genes. Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Elberg, G.; Gimble, J.M.; Tsai, S.Y. Modulation of the murine peroxisome proliferator-activated receptor gamma 2 promoter activity by CCAAT/enhancer-binding proteins. J. Biol. Chem. 2000, 275, 27815–27822. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.L.; Robinson, C.E.; Gimble, J.M. CAAT/enhancer binding proteins directly modulate transcription from the peroxisome proliferator-activated receptor gamma 2 promoter. Biochem. Biophys. Res. Commun. 1997, 240, 99–103. [Google Scholar] [CrossRef]
- Zuo, Y.; Qiang, L.; Farmer, S.R. Activation of CCAAT/enhancer-binding protein (C/EBP) alpha expression by C/EBP beta during adipogenesis requires a peroxisome proliferator-activated receptor-gamma-associated repression of HDAC1 at the C/ebp alpha gene promoter. J. Biol. Chem. 2006, 281, 7960–7967. [Google Scholar] [CrossRef] [Green Version]
- Kulterer, O.C.; Niederstaetter, L.; Herz, C.T.; Haug, A.R.; Bileck, A.; Pils, D.; Kautzky-Willer, A.; Gerner, C.; Kiefer, F.W. The Presence of Active Brown Adipose Tissue Determines Cold-Induced Energy Expenditure and Oxylipin Profiles in Humans. J. Clin. Endocrinol. Metab. 2020, 105, 2203–2216. [Google Scholar] [CrossRef]
- Blondin, D.P.; Daoud, A.; Taylor, T.; Tingelstad, H.C.; Bézaire, V.; Richard, D.; Carpentier, A.C.; Taylor, A.W.; Harper, M.E.; Aguer, C.; et al. Four-week cold acclimation in adult humans shifts uncoupling thermogenesis from skeletal muscles to brown adipose tissue. J. Physiol. 2017, 595, 2099–2113. [Google Scholar] [CrossRef] [Green Version]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerbäck, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Matsushita, M.; Yoneshiro, T.; Aita, S.; Kameya, T.; Sugie, H.; Saito, M. Impact of brown adipose tissue on body fatness and glucose metabolism in healthy humans. Int. J. Obes. (Lond.) 2014, 38, 812–817. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.I.; Middelbeek, R.J.; Goodyear, L.J. Exercise Effects on White Adipose Tissue: Beiging and Metabolic Adaptations. Diabetes 2015, 64, 2361–2368. [Google Scholar] [CrossRef] [Green Version]
- Enerbäck, S.; Jacobsson, A.; Simpson, E.M.; Guerra, C.; Yamashita, H.; Harper, M.E.; Kozak, L.P. Mice lacking mitochondrial uncoupling protein are cold-sensitive but not obese. Nature 1997, 387, 90–94. [Google Scholar] [CrossRef]
- Golozoubova, V.; Cannon, B.; Nedergaard, J. UCP1 is essential for adaptive adrenergic nonshivering thermogenesis. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E350–E357. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, H.M.; Golozoubova, V.; Cannon, B.; Nedergaard, J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab. 2009, 9, 203–209. [Google Scholar] [CrossRef]
- Uldry, M.; Yang, W.; St-Pierre, J.; Lin, J.; Seale, P.; Spiegelman, B.M. Complementary action of the PGC-1 coactivators in mitochondrial biogenesis and brown fat differentiation. Cell Metab. 2006, 3, 333–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, R.H. A NOTE ON THE ABSORPTION OF FAT. Am. J. Physiol.-Leg. Content 1909, 24, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Sepa-Kishi, D.M.; Jani, S.; Da Eira, D.; Ceddia, R.B. Cold acclimation enhances UCP1 content, lipolysis, and triacylglycerol resynthesis, but not mitochondrial uncoupling and fat oxidation, in rat white adipocytes. Am. J. Physiol. Cell Physiol. 2019, 316, C365–C376. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, H.; Zechner, R.; Zimmermann, R. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 2006, 281, 40236–40241. [Google Scholar] [CrossRef] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef]
- Shin, H.; Ma, Y.; Chanturiya, T.; Cao, Q.; Wang, Y.; Kadegowda, A.K.G.; Jackson, R.; Rumore, D.; Xue, B.; Shi, H.; et al. Lipolysis in Brown Adipocytes Is Not Essential for Cold-Induced Thermogenesis in Mice. Cell Metab. 2017, 26, 764–777.e765. [Google Scholar] [CrossRef] [Green Version]
- Daval, M.; Diot-Dupuy, F.; Bazin, R.; Hainault, I.; Viollet, B.; Vaulont, S.; Hajduch, E.; Ferré, P.; Foufelle, F. Anti-lipolytic action of AMP-activated protein kinase in rodent adipocytes. J. Biol. Chem. 2005, 280, 25250–25257. [Google Scholar] [CrossRef] [Green Version]
- van der Vaart, J.I.; Boon, M.R.; Houtkooper, R.H. The Role of AMPK Signaling in Brown Adipose Tissue Activation. Cells 2021, 10, 1122. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. AMP-Activated Protein Kinase (AMPK) Regulates Energy Metabolism through Modulating Thermogenesis in Adipose Tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kazak, L.; Jedrychowski, M.P.; Lu, G.Z.; Erickson, B.K.; Szpyt, J.; Pierce, K.A.; Laznik-Bogoslavski, D.; Vetrivelan, R.; Clish, C.B.; et al. Mitochondrial ROS regulate thermogenic energy expenditure and sulfenylation of UCP1. Nature 2016, 532, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [Green Version]
- Lefranc, C.; Friederich-Persson, M.; Palacios-Ramirez, R.; Nguyen Dinh Cat, A. Mitochondrial oxidative stress in obesity: Role of the mineralocorticoid receptor. J. Endocrinol. 2018, 238, R143–R159. [Google Scholar] [CrossRef]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef]
- Jaiswal, A.K. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic. Biol. Med. 2004, 36, 1199–1207. [Google Scholar] [CrossRef]
- Bellner, L.; Lebovics, N.B.; Rubinstein, R.; Buchen, Y.D.; Sinatra, E.; Sinatra, G.; Abraham, N.G.; McClung, J.A.; Thompson, E.A. Heme Oxygenase-1 Upregulation: A Novel Approach in the Treatment of Cardiovascular Disease. Antioxid. Redox. Signal. 2020, 32, 1045–1060. [Google Scholar] [CrossRef]
- Singh, S.P.; Schragenheim, J.; Cao, J.; Falck, J.R.; Abraham, N.G.; Bellner, L. PGC-1 alpha regulates HO-1 expression, mitochondrial dynamics and biogenesis: Role of epoxyeicosatrienoic acid. Prostaglandins Other Lipid Mediat. 2016, 125, 8–18. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula | ND | HFD |
|---|---|---|
| Casein | 200.0 | 265.0 |
| L-Cystine | 3.0 | 4.0 |
| Corn Starch | 397.486 | - |
| Maltodextrin | 132.0 | 160.0 |
| Sucrose | 100.0 | 90.0 |
| Lard | - | 310.0 |
| Soybean Oil | 70.0 | 30.0 |
| Cellulose | 50.0 | 65.5 |
| Mineral Mix a | 35.0 | 48.0 |
| Calcium Phosphate | - | 3.4 |
| Vitamin Mix b | 10.0 | 21.0 |
| Choline Bitartrate | 2.5 | 3.0 |
| TBHQ, antioxidant c | 0.014 | - |
| Blue Food Color | - | 0.1 |
| Primary Antibody | Manufacturer | Catalog No. | Molecular Weight |
|---|---|---|---|
| PPARγ | Cell Signaling Technology | 2435s | 53, 57 |
| C/EBPα | Santa Cruz Biotechnology | sc-61 | 42, 30 |
| ATGL | Abcam | EPR19650 | 55 |
| p-HSL | Cell Signaling Technology | 4139S | 81, 83 |
| HSL | Abcam | ab45422 | 90 |
| p-AMPK | Cell Signaling Technology | 2535s | 62 |
| AMPK | Cell Signaling Technology | 2532s | 62 |
| UCP1 | GeneTex | GTX112784 | 33 |
| PGC1α | Thermo Fischer Scientific | PA5-38021 | 90 |
| β-actin | Cell Signaling Technology | 3700S | 45 |
| GAPDH | Santa Cruz Biotechnology | sc-32233 | 37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, W.Y.; Song, G.; Boo, M.; Kim, H.I.; Park, J.Y.; Jung, S.J.; Choi, M.; Kim, B.; Kim, Y.D.; Kim, M.-H.; et al. Anmyungambi Decoction Ameliorates Obesity through Activation of Non-Shivering Thermogenesis in Brown and White Adipose Tissues. Antioxidants 2023, 12, 49. https://doi.org/10.3390/antiox12010049
Park WY, Song G, Boo M, Kim HI, Park JY, Jung SJ, Choi M, Kim B, Kim YD, Kim M-H, et al. Anmyungambi Decoction Ameliorates Obesity through Activation of Non-Shivering Thermogenesis in Brown and White Adipose Tissues. Antioxidants. 2023; 12(1):49. https://doi.org/10.3390/antiox12010049
Chicago/Turabian StylePark, Woo Yong, Gahee Song, Mina Boo, Hyo In Kim, Ja Yeon Park, Se Jin Jung, Minji Choi, Beomsu Kim, Young Doo Kim, Myung-Ho Kim, and et al. 2023. "Anmyungambi Decoction Ameliorates Obesity through Activation of Non-Shivering Thermogenesis in Brown and White Adipose Tissues" Antioxidants 12, no. 1: 49. https://doi.org/10.3390/antiox12010049