
The Synthetic Flavonoid Hidrosmin Improves Endothelial Dysfunction and Atherosclerotic Lesions in Diabetic Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Design of the Experimental Mouse Models of Diabetes
2.3. Functional and Structural Vascular Studies
2.4. Pressure Myography Studies
2.5. Histological and Immunohistochemical Analysis of Atherosclerotic Lesions
2.6. In Vitro Experiments
2.7. mRNA Expression Analysis
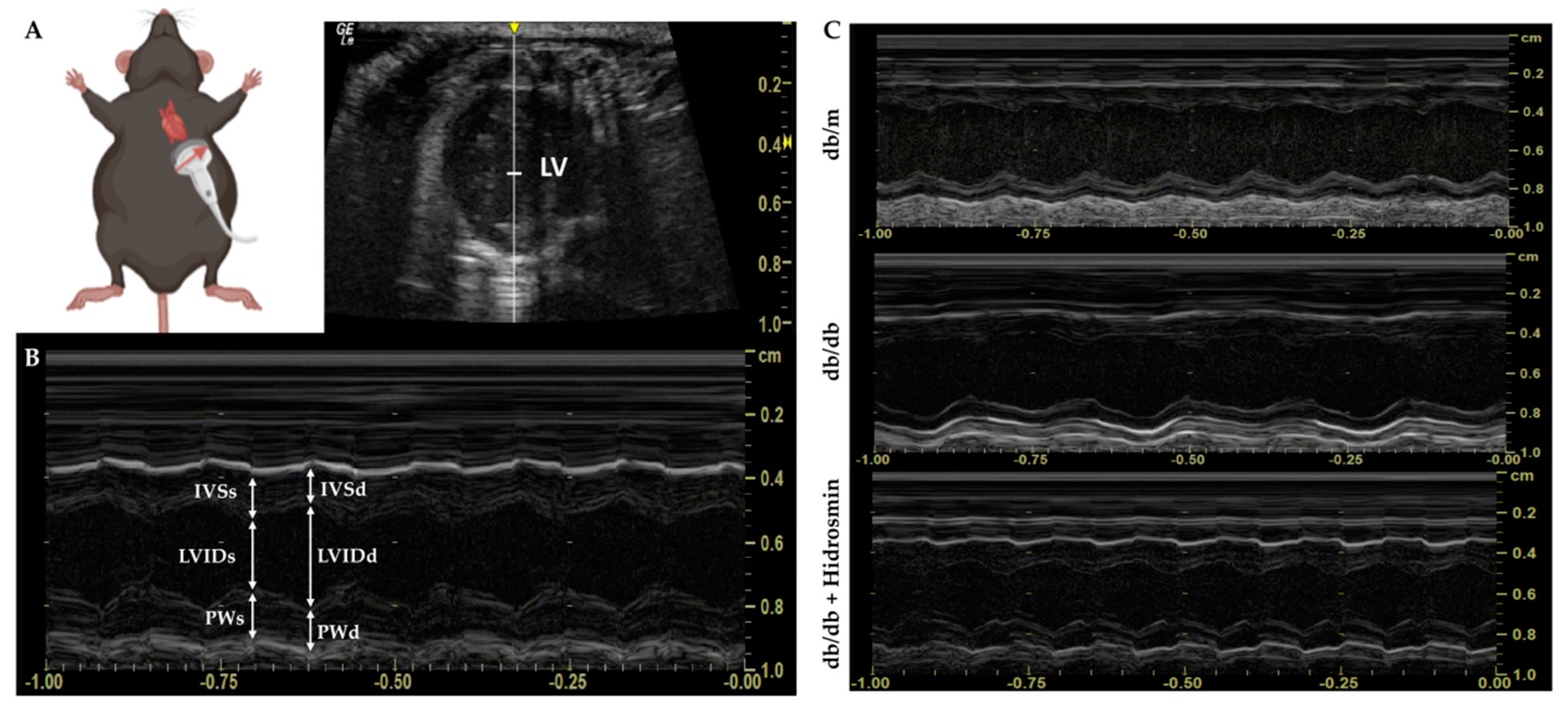
2.8. Echocardiography Assessment
2.9. Statistical Analysis
3. Results
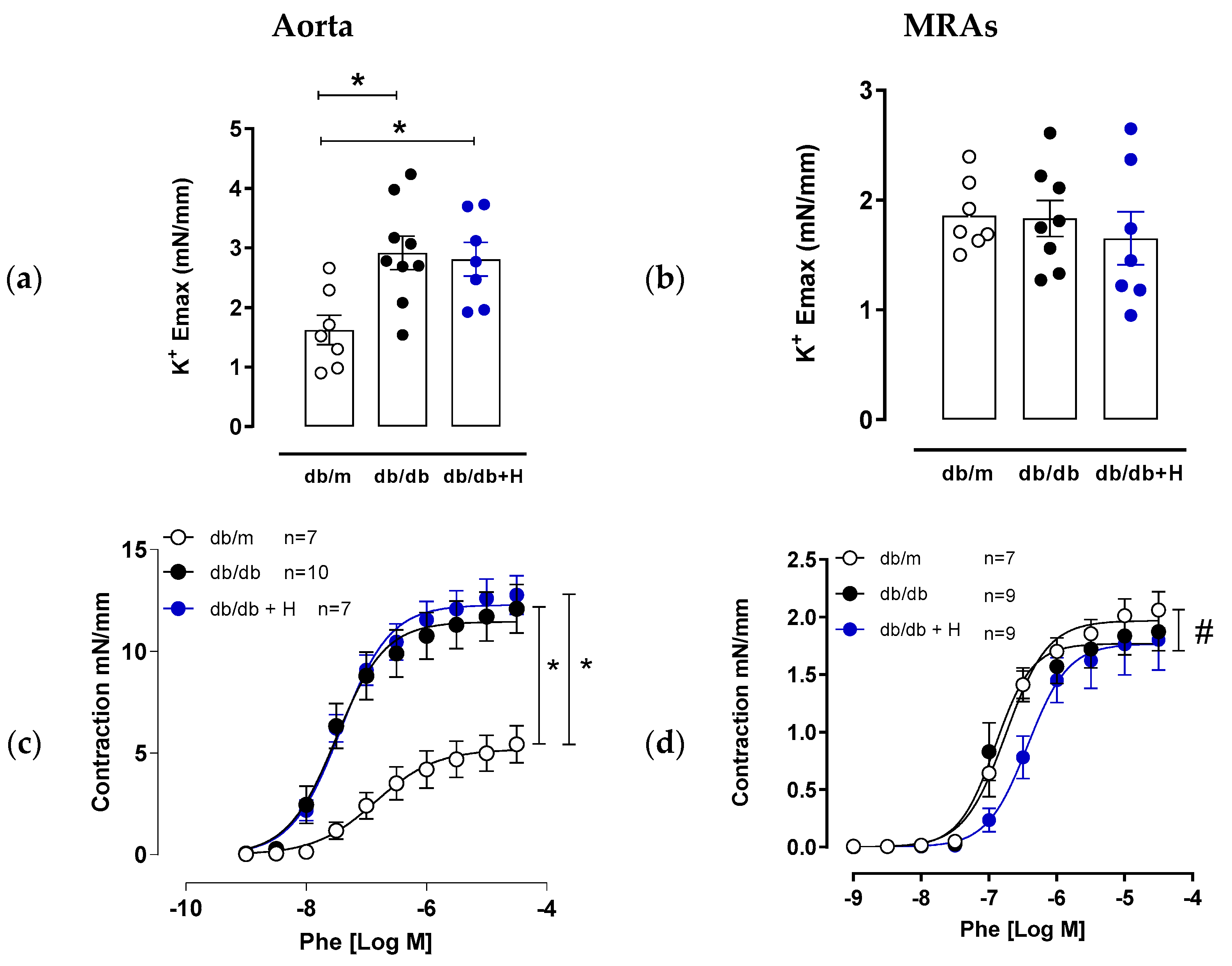
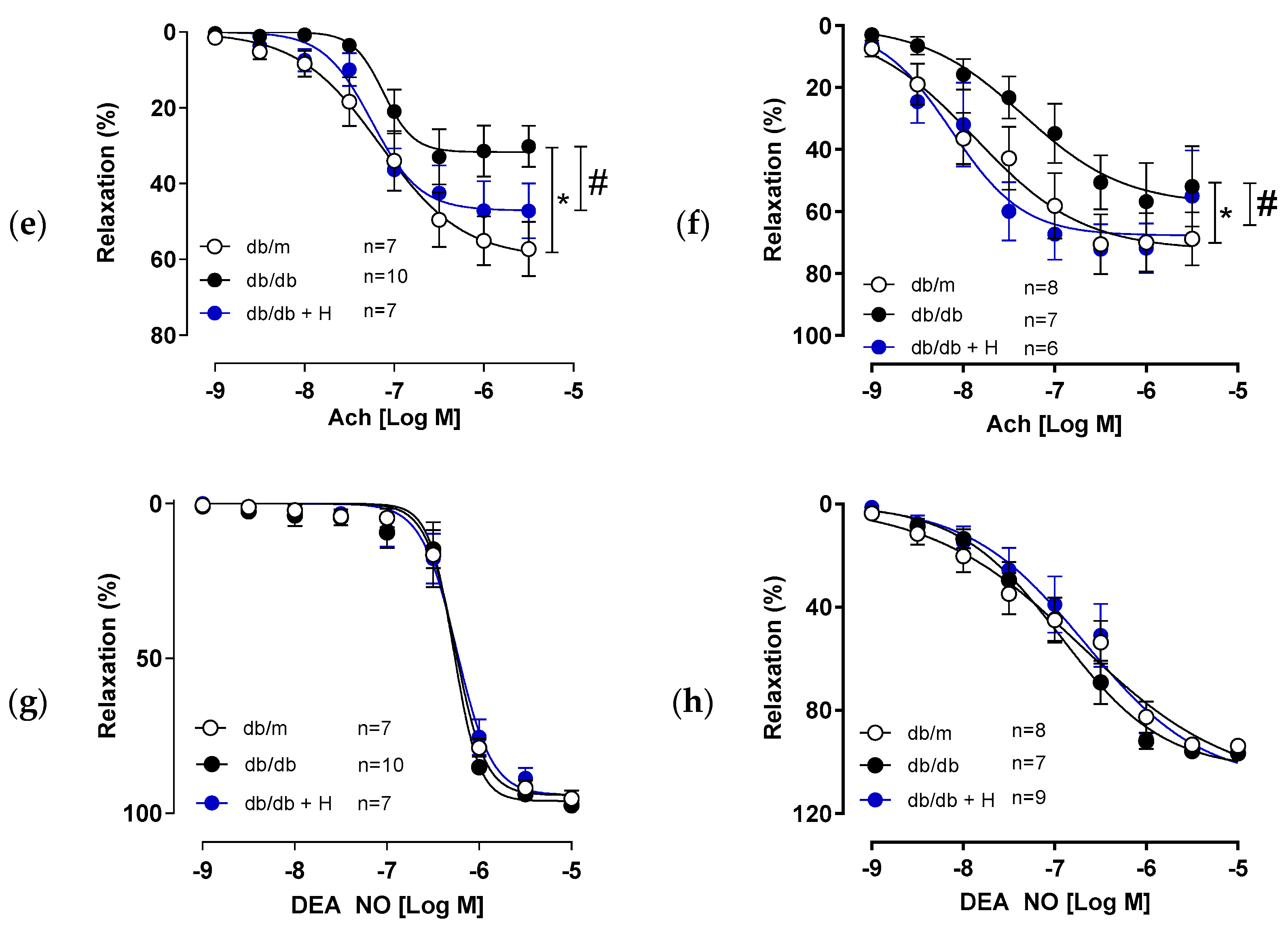
3.1. Hidrosmin Treatment Improves Endothelial Function in Aorta and Mesenteric Arteries
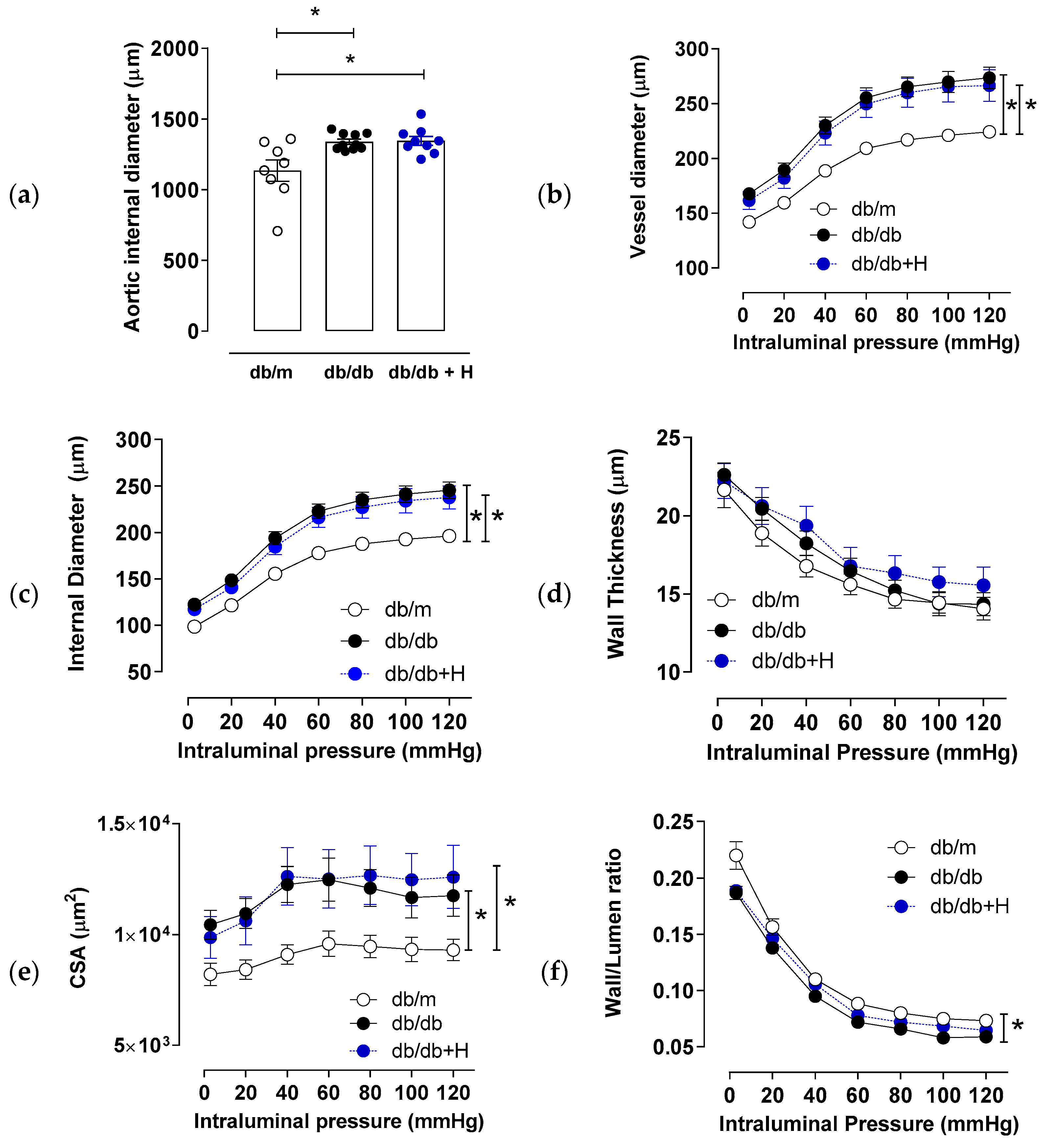
3.2. Hidrosmin Treatment Does Not Affect Structural Properties of Aorta and Mesenteric Arteries
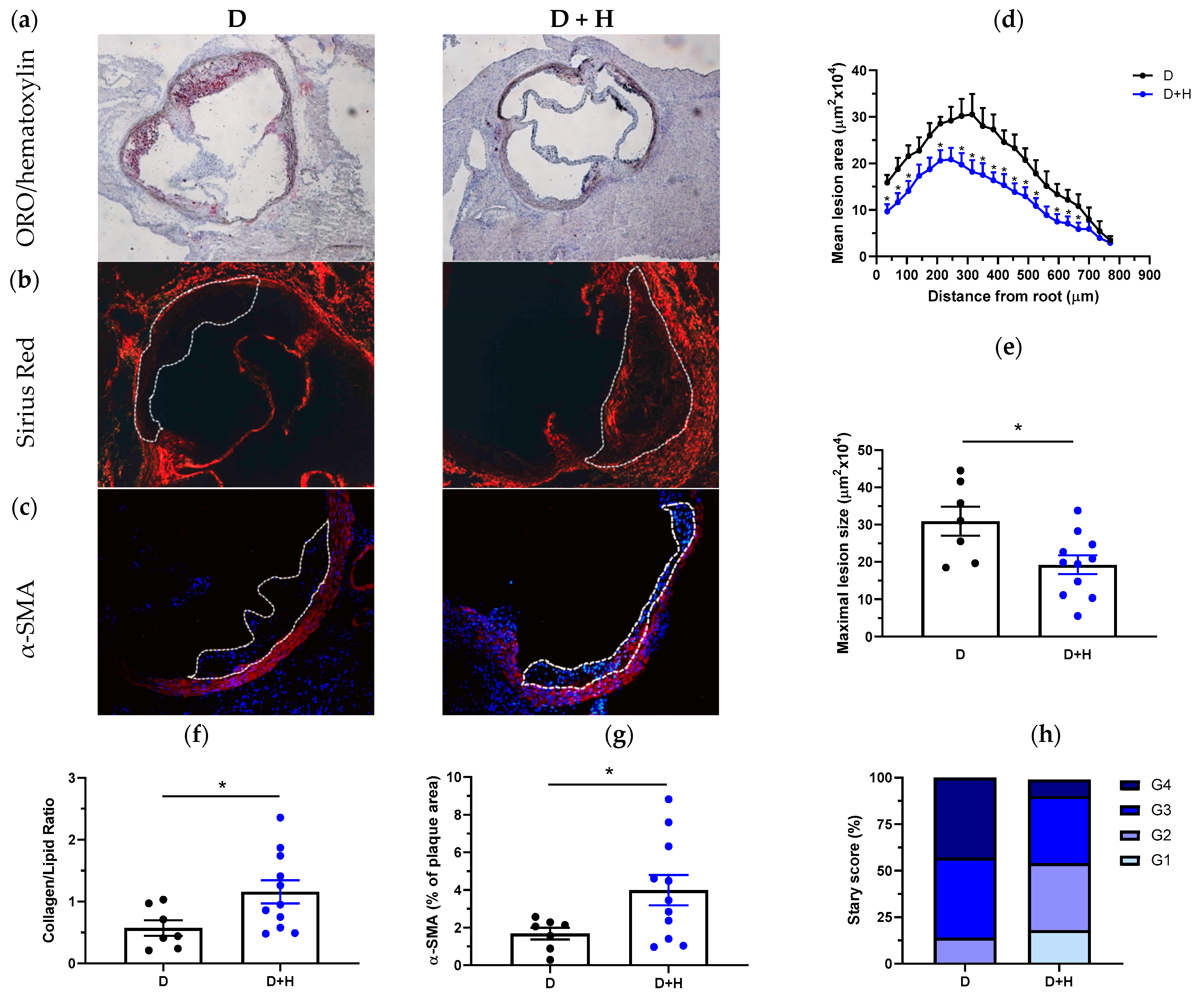
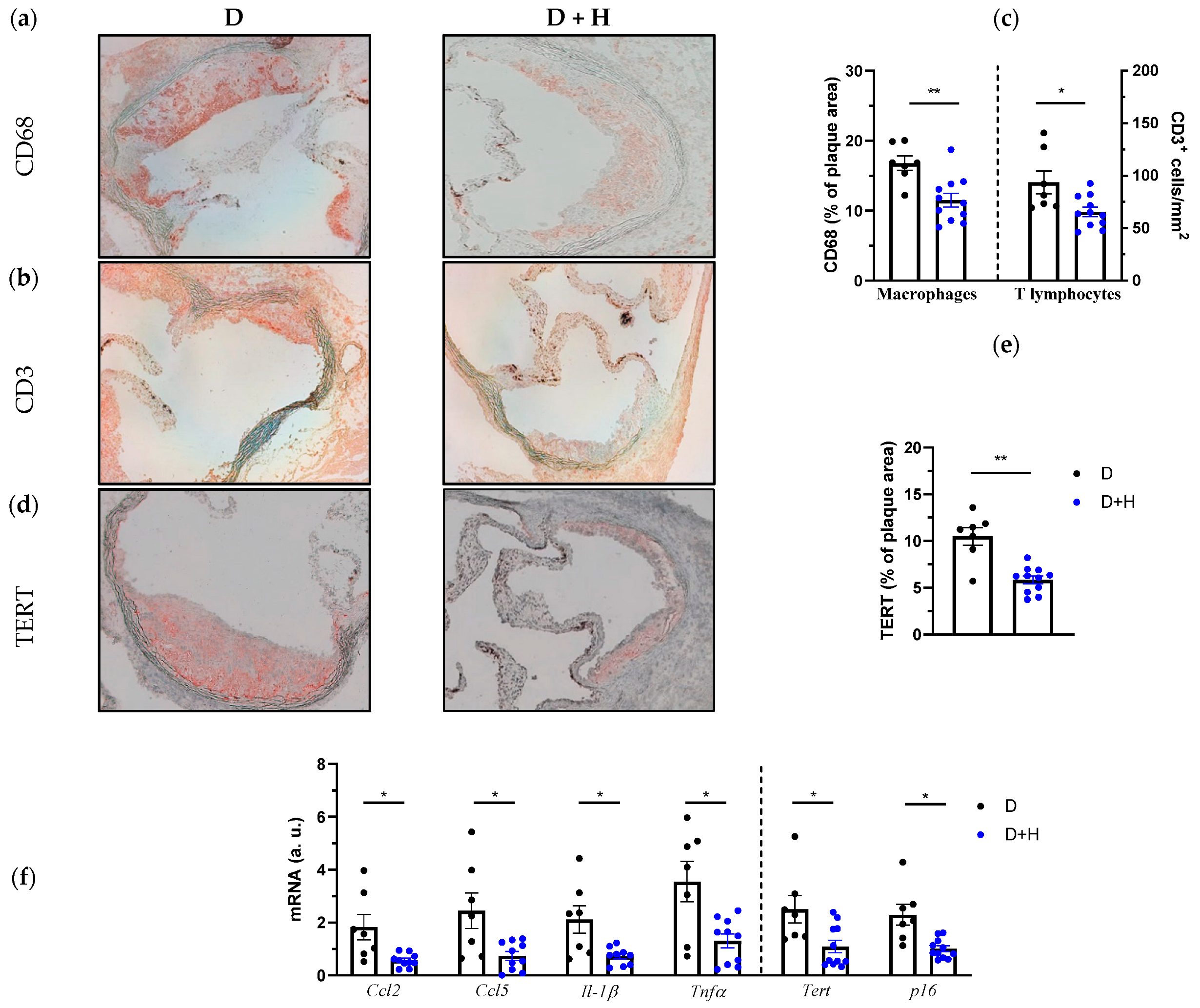
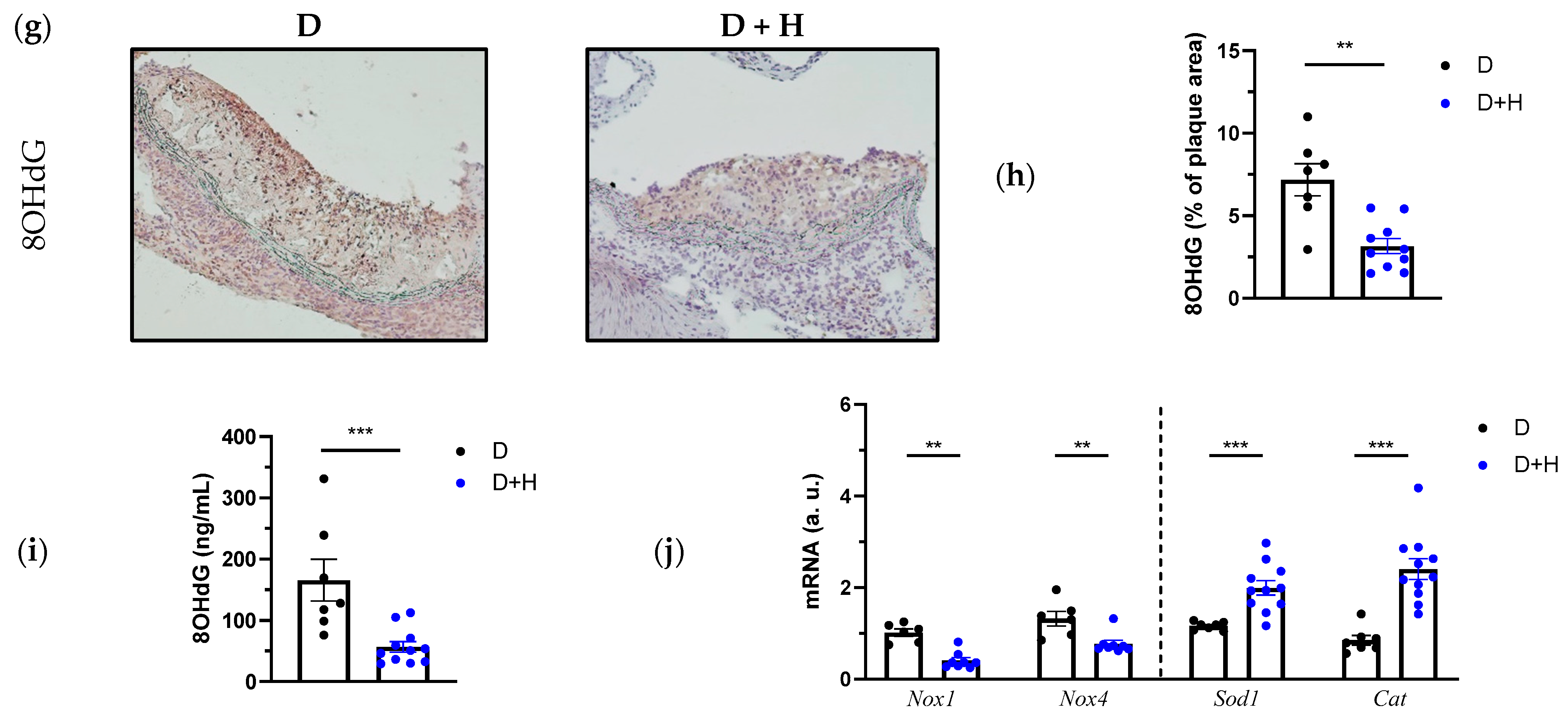
3.3. Hidrosmin Prevents Atherosclerosis Development and Partially Reverts Features of Plaque Instability, Inflammation and Oxidative Stress in Diabetic Mice
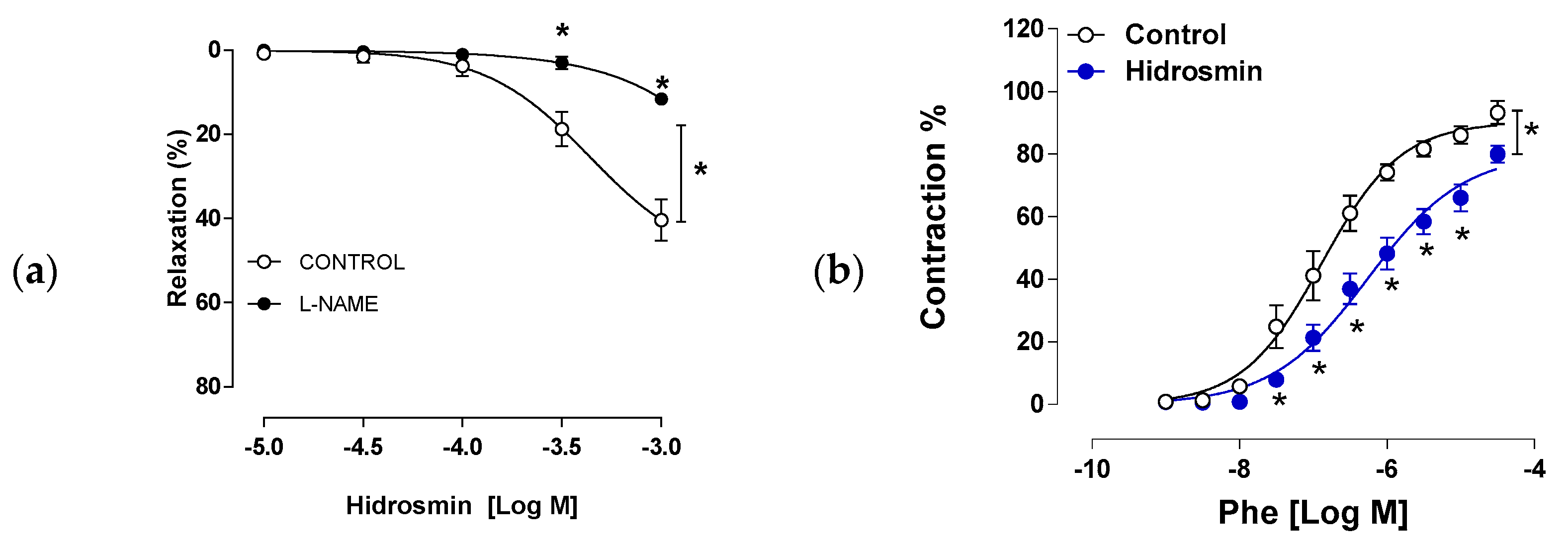
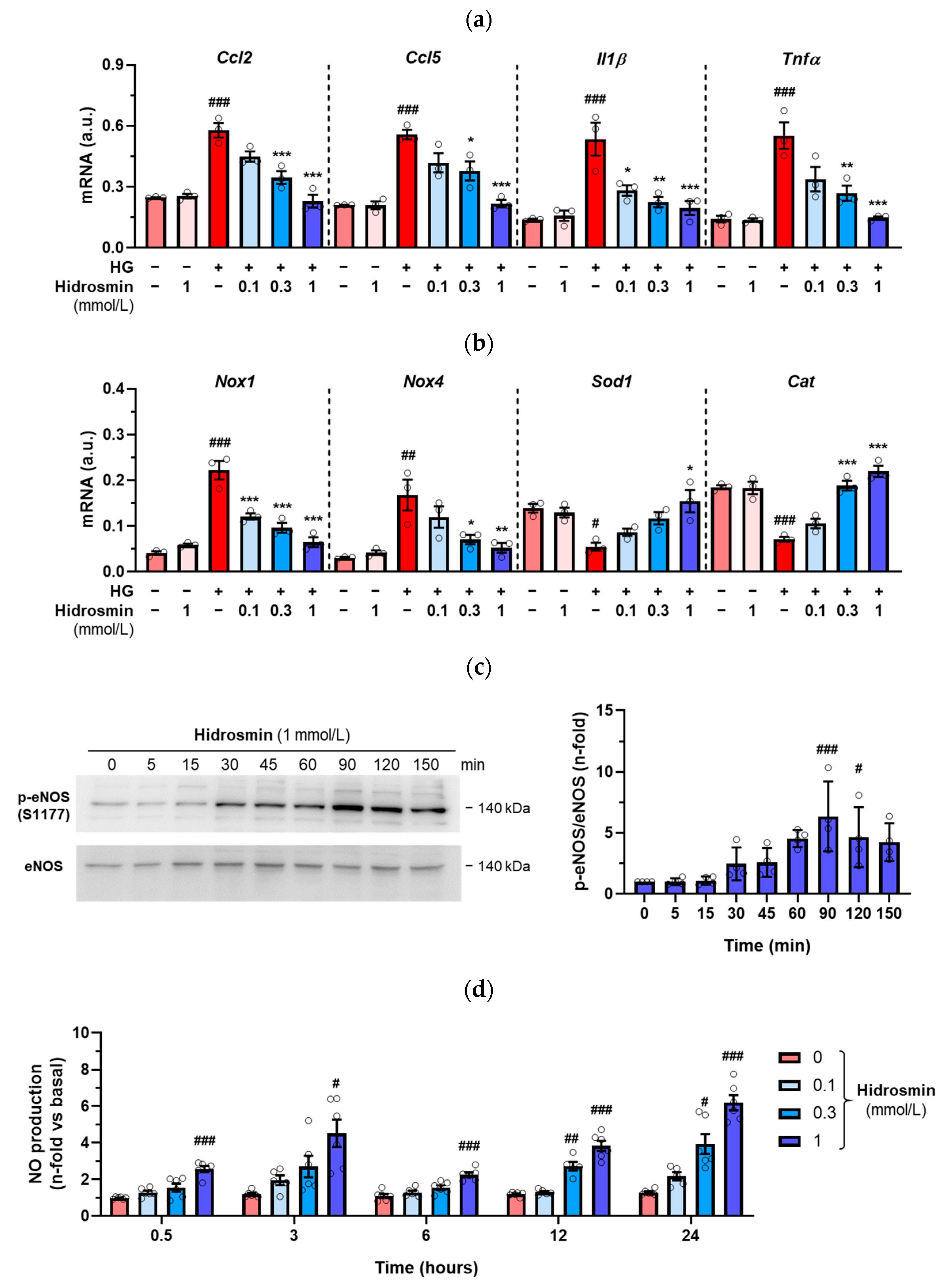
3.4. Effects of Hidrosmin on Cultured Vascular Cells
3.5. Hidrosmin Treatment Does Not Affect the Structure and Function of Hearts from Diabetic Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lind, L.; Sundström, J.; Ärnlöv, J.; Risérus, U.; Lampa, E. A longitudinal study over 40 years to study the metabolic syndrome as a risk factor for cardiovascular diseases. Sci. Rep. 2021, 11, 2978. [Google Scholar] [CrossRef]
- Einarson, T.R.; Acs, A.; Ludwig, C.; Panton, U.H. Prevalence of cardiovascular disease in type 2 diabetes: A systematic literature review of scientific evidence from across the world in 2007–2017. Cardiovasc. Diabetol. 2010, 17, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjesdal, O.; Bluemke, D.A.; Lima, J.A. Cardiac remodeling at the population level--risk factors, screening, and outcomes. Nat. Rev. Cardiol. 2011, 8, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Low Wang, C.C.; Hess, C.N.; Hiatt, W.R.; Goldfine, A.B. Clinical Update: Cardiovascular Disease in Diabetes Mellitus: Atherosclerotic Cardiovascular Disease and Heart Failure in Type 2 Diabetes Mellitus—Mechanisms, Management, and Clinical Considerations. Circulation 2016, 133, 2459–2502. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, A.O.; Jacobs, D.R.; Sanchez, O.A.; Goff, D.C.; Reiner, A.P.; Gross, M.D. Oxidative stress, inflammation, endothelial dysfunction and incidence of type 2 diabetes. Cardiovasc. Diabetol. 2016, 15, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palombo, C.; Kozakova, M. Arterial stiffness, atherosclerosis and cardiovascular risk: Pathophysiologic mechanisms and emerging clinical indications. Vascul Pharmacol. 2016, 77, 1–7. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Kim, Y.S. The role of advanced glycation end products in diabetic vascular complications. Diabetes Metab. J. 2018, 42, 188–195. [Google Scholar] [CrossRef]
- Briones, A.M.; Aras-Lopez, R.; Alonso, M.J.; Salaices, M. Small artery remodeling in obesity and insulin resistance. Curr. Vasc. Pharmacol. 2014, 12, 427–437. [Google Scholar] [CrossRef]
- Bruder-Nascimento, T.; Callera, G.E.; Montezano, A.C.; He, Y.; Antunes, T.T.; Nguyen Dinh Cat, A.; Tostes, R.C.; Touyz, R.M. Vascular injury in diabetic db/db mice is ameliorated by atorvastatin: Role of Rac1/2-sensitive Nox-dependent pathways. Clin. Sci. 2015, 128, 411–423. [Google Scholar] [CrossRef]
- Bäck, M.; Yurdagul, A.; Tabas, I.; Öörni, K.; Kovanen, P.T. Inflammation and its resolution in atherosclerosis: Mediators and therapeutic opportunities. Nat. Rev. Cardiol. 2019, 16, 389–406. [Google Scholar] [CrossRef]
- Poznyak, A.; Grechko, A.V.; Poggio, P.; Myasoedova, V.A.; Alfieri, V.; Orekhov, A.N. The Diabetes Mellitus–Atherosclerosis Connection: The Role of Lipid and Glucose Metabolism and Chronic Inflammation. Int. J. Mol. Sci. 2020, 21, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, W.; Lobo, M.; Siniawski, D.; Molinero, G.; Masson, G.; Huerín, M.; Nogueira, J.P. Role of non-statin lipid-lowering therapy in coronary atherosclerosis regression: A meta-analysis and meta-regression. Lipids Health Dis. 2020, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, S.; Liu, B.; Yan, Y.; Du, J.; Li, Y.; Jing, X.; Liu, Y.; Wang, J.; Du, J.; et al. The effects of lipid-lowering therapy on coronary plaque regression: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 7999. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M. Anticytokine Agents: Targeting Interleukin Signaling Pathways for the Treatment of Atherothrombosis. Circ. Res. 2019, 124, 437–450. [Google Scholar] [CrossRef]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; CANTOS Trial Group; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Chen, Y.; Li, M.; Luo, W.; Liu, Y.; Fu, Y.; Xia, H.; Xu, C.; Jiang, Y.; et al. Colchicine may become a new cornerstone therapy for coronary artery disease: A meta-analysis of randomized controlled trials. Clin. Rheumatol. 2022, 41, 1873–1887. [Google Scholar] [CrossRef]
- Wiciński, M.; Górski, K.; Wódkiewicz, E.; Walczak, M.; Nowaczewska, M.; Malinowski, B. Vasculoprotective Effects of Vildagliptin. Focus on Atherogenesis. Int. J. Mol. Sci. 2020, 21, 2275. [Google Scholar] [CrossRef] [Green Version]
- Caro-Ordieres, T.; Marín-Royo, G.; Opazo-Ríos, L.; Jiménez-Castilla, L.; Moreno, J.A.; Gómez-Guerrero, C.; Egido, J. The Coming Age of Flavonoids in the Treatment of Diabetic Complications. J. Clin. Med. 2020, 9, 346. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Sajid, M.; Channakesavula, C.N.; Stone, S.R.; Kaur, P. Synthetic Biology towards Improved Flavonoid Pharmacokinetics. Biomolecules 2021, 11, 754. [Google Scholar] [CrossRef]
- Kakkos, S.K.; Nicolaides, A.N. Efficacy of micronized purified flavonoid fraction (Daflon®) on improving individual symptoms, signs and quality of life in patients with chronic venous disease: A systematic review and meta-analysis of randomized double-blind placebo-controlled trials. Int. Angiol. 2018, 37, 143–154. [Google Scholar] [CrossRef]
- Feldo, M.; Wójciak-Kosior, M.; Sowa, I.; Kocki, J.; Bogucki, J.; Zubilewicz, T.; Kęsik, J.; Bogucka-Kocka, A. Effect of Diosmin Administration in Patients with Chronic Venous Disorders on Selected Factors Affecting Angiogenesis. Molecules 2019, 24, 3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, T.M.; Abo-Salem, O.M.; El Esawy, B.H.; El Askary, A. The Potential Protective Effects of Diosmin on Streptozotocin-Induced Diabetic Cardiomyopathy in Rats. Am. J. Med. Sci. 2020, 359, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Chen, Z.; Wu, L. Metabolism and pharmacological activities of the natural health-benefiting compound diosmin. Food Funct. 2020, 11, 8472–8492. [Google Scholar] [CrossRef] [PubMed]
- Om, H.; El-Naggar, M.E.; El-Banna, M.; Fouda, M.M.G.; Othman, S.I.; Allam, A.A.; Morsy, O.M. Combating atherosclerosis with targeted Diosmin nanoparticles-treated experimental diabetes. Investig. New Drugs 2020, 38, 1303–1315. [Google Scholar] [CrossRef]
- Martinez-Zapata, M.J.; Vernooij, R.W.; Simancas-Racines, D.; Uriona Tuma, S.M.; Stein, A.T.; Moreno Carriles, R.M.M.; Vargas, E.; Bonfill Cosp, X. Phlebotonics for venous insufficiency. Cochrane Database Syst. Rev. 2020, 11, CD003229. [Google Scholar] [CrossRef]
- Gloviczki, P.; Comerota, A.J.; Dalsing, M.C.; Eklof, B.G.; Gillespie, D.L.; Gloviczki, M.L.; Lohr, J.M.; McLafferty, R.B.; Meissner, M.H.; Murad, M.H.; et al. The care of patients with varicose veins and associated chronic venous diseases: Clinical practice guidelines of the Society for Vascular Surgery and the American Venous Forum. J. Vasc. Surg. 2011, 53, 2S–48S. [Google Scholar] [CrossRef] [Green Version]
- Wittens, C.; Davies, A.H.; Bækgaard, N.; Broholm, R.; Cavezzi, A.; Chastanet, S.; de Wolf, M.; Eggen, C.; Giannoukas, A.; Gohel, M.; et al. Editor’s Choice—Management of Chronic Venous Disease: Clinical Practice Guidelines of the European Society for Vascular Surgery (ESVS). Eur. J. Vasc. Endovasc. Surg. 2015, 49, 678–737. [Google Scholar] [CrossRef] [Green Version]
- Paredes, M.D.; Romecín, P.; Atucha, N.M.; O’Valle, F.; Castillo, J.; Ortiz, M.C.; García-Estañ, J. Moderate Effect of Flavonoids on Vascular and Renal Function in Spontaneously Hypertensive Rats. Nutrients 2018, 10, 1107. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Castilla, L.; Marín-Royo, G.; Orejudo, M.; Opazo-Ríos, L.; Caro-Ordieres, T.; Artaiz, I.; Suárez-Cortés, T.; Zazpe, A.; Hernández, G.; Gómez-Guerrero, C.; et al. Nephroprotective Effects of Synthetic Flavonoid Hidrosmin in Experimental Diabetic Nephropathy. Antioxidants 2021, 10, 1920. [Google Scholar] [CrossRef]
- Hsueh, W.; Abel, E.D.; Breslow, J.L.; Maeda, N.; Davis, R.C.; Fisher, E.A.; Dansky, H.; McClain, D.A.; McIndoe, R.; Wassef, M.K.; et al. Recipes for creating animal models of diabetic cardiovascular disease. Circ. Res. 2007, 100, 1415–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaro, I.; Oguiza, A.; Recio, C.; Mallavia, B.; Madrigal-Matute, J.; Blanco, J.; Egido, J.; Martin-Ventura, J.L.; Gomez-Guerrero, C. Targeting HSP90 Ameliorates Nephropathy and Atherosclerosis Through Suppression of NF-κB and STAT Signaling Pathways in Diabetic Mice. Diabetes 2015, 64, 3600–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recio, C.; Oguiza, A.; Lazaro, I.; Mallavia, B.; Egido, J.; Gomez-Guerrero, C. Suppressor of cytokine signaling 1-derived peptide inhibits Janus kinase/signal transducers and activators of transcription pathway and improves inflammation and atherosclerosis in diabetic mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1953–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros-Martínez, C.; Rodrigues-Díez, R.; Beltrán, L.M.; Moreno-Carriles, R.; Martínez-Martínez, E.; González-Amor, M.; Martínez-González, J.; Rodríguez, C.; Cachofeiro, V.; Salaices, M.; et al. Microsomal prostaglandin E synthase-1 is involved in the metabolic and cardiovascular alterations associated with obesity. Br. J. Pharmacol. 2022, 179, 2733–2753. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Glagov, S.; Guyton, J.R.; Insull, W.; Rosenfeld, M.E.; Schaffer, S.A.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1994, 89, 2462–2478. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Ho, D.; Vatner, D.E.; Vatner, S.F. Echocardiography in Mice. Curr. Protoc. Mouse Biol. 2011, 1, 71–83. [Google Scholar] [CrossRef]
- Nguyen Dinh Cat, A.; Callera, G.E.; Friederich-Persson, M.; Sanchez, A.; Dulak-Lis, M.G.; Tsiropoulou, S.; Montezano, A.C.; He, Y.; Briones, A.M.; Jaisser, F.; et al. Vascular dysfunction in obese diabetic db/db mice involves the interplay between aldosterone/mineralocorticoid receptor and Rho kinase signaling. Sci. Rep. 2018, 8, 2952. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, N.S.; Bruder-Nascimento, T.; Pereira, C.A.; Zanotto, C.Z.; Prado, D.S.; Silva, J.F.; Rassi, D.M.; Foss-Freitas, M.C.; Alves-Filho, J.C.; Carlos, D.; et al. NLRP3 Inflammasome and Mineralocorticoid Receptors Are Associated with Vascular Dysfunction in Type 2 Diabetes Mellitus. Cells 2019, 8, 1595. [Google Scholar] [CrossRef] [Green Version]
- Arildsen, L.; Andersen, J.V.; Waagepetersen, H.S.; Nissen, J.B.D.; Sheykhzade, M. Hypermetabolism and impaired endothelium-dependent vasodilation in mesenteric arteries of type 2 diabetes mellitus db/db mice. Diabetes Vasc. Dis. Res. 2019, 16, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Kerins, D.M.; Walther, T. Cardiovascular safety of anti-diabetic drugs. Eur. Heart J. Cardiovasc. Pharmacother. 2016, 2, 32–43. [Google Scholar] [CrossRef]
- Low Wang, C.C.; Everett, B.M.; Burman, K.D.; Wilson, P.W.F. Cardiovascular Safety Trials for All New Diabetes Mellitus Drugs? Circulation 2019, 139, 1741–1743. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Antras, J.; Picatoste, B.; Gomez-Hernandez, A.; Egido, J.; Tunon, J.; Lorenzo, O. Updating experimental models of diabetic cardiomyopathy. J. Diabetes Res. 2015, 2015, 656795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alex, L.; Russo, I.; Holoborodko, V.; Frangogiannis, N.G. Characterization of a mouse model of obesity-related fibrotic cardiomyopathy that recapitulates features of human heart failure with preserved ejection fraction. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H934–H949. [Google Scholar] [CrossRef] [Green Version]
- El-Fawal, R.; El Fayoumi, H.M.; Mahmoud, M.F. Effects of diosmin and crocin on metabolic syndrome-associated cardio-vascular complications in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1523–1536. [Google Scholar] [CrossRef]
- Li, K.X.; Diendéré, G.; Galanaud, J.P.; Mahjoub, N.; Kahn, S.R. Micronized purified flavonoid fraction for the treatment of chronic venous insufficiency, with a focus on postthrombotic syndrome: A narrative review. Res. Pract. Thromb. Haemost. 2021, 5, e12527. [Google Scholar] [CrossRef]
- Silambarasan, T.; Raja, B. Diosmin, a bioflavonoid reverses alterations in blood pressure, nitric oxide, lipid peroxides and antioxidant status in DOCA-salt induced hypertensive rats. Eur. J. Pharmacol. 2012, 679, 81–89. [Google Scholar] [CrossRef]
- Duarte, J.; Francisco, V.; Perez-Vizcaino, F. Modulation of nitric oxide by flavonoids. Food Funct. 2014, 5, 1653–1668. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Babu, P.V.A.; Symons, J.D.; Jalili, T. Metabolites of flavonoid compounds preserve indices of endothelial cell nitric oxide bioavailability under glucotoxic conditions. Nutr. Diabetes 2017, 7, e286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markus, M.R.P.; Rospleszcz, S.; Ittermann, T.; Baumeister, S.E.; Schipf, S.; Siewert-Markus, U.; Lorbeer, R.; Storz, C.; Ptushkina, V.; Peters, A.; et al. Glucose and insulin levels are associated with arterial stiffness and concentric remodeling of the heart. Cardiovasc. Diabetol. 2019, 18, 145. [Google Scholar] [CrossRef]
- Rizzoni, D.; Agabiti-Rosei, E. Endothelial factors and microvascular hypertensive disease. J. Cardiovasc. Pharmacol. 2001, 38, S15–S18. [Google Scholar] [CrossRef]
- Rizzoni, D.; Porteri, E.; Guelfi, D.; Muiesan, M.L.; Valentini, U.; Cimino, A.; Girelli, A.; Rodella, L.; Bianchi, R.; Sleiman, I.; et al. Structural alterations in subcutaneous small arteries of normotensive and hypertensive patients with non-insulin-dependent diabetes mellitus. Circulation 2001, 103, 1238–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza-Smith, F.M.; Katz, P.S.; Trask, A.J.; Stewart, J.A.; Lord, K.C.; Varner, K.J.; Vassallo, D.V.; Lucchesi, P.A. Mesenteric resistance arteries in type 2 diabetic db/db mice undergo outward remodeling. PLoS ONE 2011, 6, e23337. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Cau, S.B.; Lopes, R.A.; Manzato, C.P.; Neves, K.B.; Bruder-Nascimento, T.; Mestriner, F.L.; Montezano, A.C.; Nguyen Dinh Cat, A.; Touyz, R.M.; et al. Mineralocorticoid receptor blockade prevents vascular remodelling in a rodent model of type 2 diabetes mellitus. Clin. Sci. 2015, 129, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.M.; Zheng, L.; Wang, Q.; Hu, Y.W. The emerging role of cell senescence in atherosclerosis. Clin. Chem. Lab. Med. 2020, 59, 27–38. [Google Scholar] [CrossRef]
- Liu, S.C.; Wang, S.S.; Wu, M.Z.; Wu, D.C.; Yu, F.J.; Chen, W.J.; Chiang, F.T.; Yu, M.F. Activation of telomerase and expression of human telomerase reverse transcriptase in coronary atherosclerosis. Cardiovasc. Pathol. 2005, 14, 232–240. [Google Scholar] [CrossRef]
- Kang, G.G.; Francis, N.; Hill, R.; Waters, D.; Blanchard, C.; Santhakumar, A.B. Dietary Polyphenols and Gene Expression in Molecular Pathways Associated with Type 2 Diabetes Mellitus: A Review. Int. J. Mol. Sci. 2019, 21, 140. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Mundhe, N.; Borgohain, M.; Chowdhury, L.; Kwatra, M.; Bolshette, N.; Ahmed, A.; Lahkar, M. Diosmin Modulates the NF-kB Signal Transduction Pathways and Downregulation of Various Oxidative Stress Markers in Alloxan-Induced Diabetic Nephropathy. Inflammation 2016, 39, 1783–1797. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Db/m | db/db | db/db + H | |
|---|---|---|---|
| Blood Glucose (mg/dL) | 139.3 ± 5.1 | 396.4 ± 25.9 * | 385.3 ± 26.0 |
| Body Weight (g) | 32.2 ± 0.46 | 52.8 ± 1.4 * | 50.4 ± 1.9 |
| Triglycerides (mg/dL) | 84.3 ± 11.5 | 94.3 ± 13.5 | 91.8 ± 11.3 |
| Total Cholesterol (mg/dL) | 88.6 ± 2.1 | 171.6 ± 10.2 * | 165.6 ± 7.6 |
| LDL-C (mg/dL) | UD | 26.3 ± 3.3 | 24.3 ± 2.7 |
| HDL-C (mg/dL) | 76.3 ± 1.6 | 128.5 ± 6.4 * | 125.9 ± 4.7 |
| ASAT (UI/L) | 111.2 ± 31.2 | 135.8 ± 11.1 | 125.8 ± 8.9 |
| ALAT (UI/L) | 39.3 ± 4.1 | 150.4 ± 18.0 * | 148.6 ± 17.1 |
| Urea (mg/dL) | 36.6 ± 1.3 | 42.0 ± 2.4 | 43.0 ± 1.3 |
| BUN (mg/dL) | 17.3 ± 0.6 | 19.6 ± 1.1 | 20.0 ± 0.6 |
| D | D + H | |
|---|---|---|
| Body Weight (g) | 21.7 ± 0.8 | 21 ± 0.9 |
| Blood Glucose (mg/dL) | 489.9 ± 27.8 | 418.3 ± 34.0 |
| Triglycerides (mg/dL) | 241.1 ± 51.2 | 129.2 ± 17.7 * |
| Total Cholesterol (mg/dL) | 928.3 ± 82.4 | 669.25 ± 82.7 * |
| LDL-C (mg/dL) | 694.5 ± 116.7 | 565.8 ± 82.5 |
| HDL-C (mg/dL) | 95.7 ± 6.8 | 83.9 ± 7.3 |
| Variables | db/m | db/db | db/db + H |
|---|---|---|---|
| IVSd (μm) | 102.4 ±4.3 | 115.4 ± 15.2 | 109.6 ± 15.1 |
| LVIDd (μm) | 379.5 ± 4.8 | 336.2 ± 46.6 | 336.3 ± 37.1 |
| PWd (μm) | 99.0 ± 2.4 | 106.2 ± 12.9 | 94.2 ± 11.8 |
| IVSs (μm) | 144.8 ± 2.7 | 171.9 ±18.9 | 165.8 ± 18.0 |
| LVIDs (μm) | 260.0 ± 4.5 | 222.4 ± 36.7 | 219.2 ± 25.3 |
| PWs (μm) | 138.6 ±2.4 | 137.1 ± 13.9 | 120.4 ± 16.9 |
| LV mass (μg) | 148.6 ± 4.7 | 146.4 ± 19.5 | 125.3 ± 16.6 |
| LV mass/BW (μg/g) | 5.0 ± 0.2 | 2.6 ± 0.4 * | 2.4 ± 0.3 * |
| EF (%) | 67.1 ± 2.4 | 71.2 ± 9.9 | 72.1 ± 10.2 |
| FS (%) | 31.3 ± 1.6 | 34.2 ± 5.0 | 35.4 ± 5.4 |
| PWT (%) | 40.5 ± 5.1 | 28.8 ± 7.4 | 29.2 ± 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Castilla, L.; Opazo-Ríos, L.; Marin-Royo, G.; Orejudo, M.; Rodrigues-Diez, R.; Ballesteros-Martínez, C.; Soto-Catalán, M.; Caro-Ordieres, T.; Artaiz, I.; Suarez-Cortés, T.; et al. The Synthetic Flavonoid Hidrosmin Improves Endothelial Dysfunction and Atherosclerotic Lesions in Diabetic Mice. Antioxidants 2022, 11, 2499. https://doi.org/10.3390/antiox11122499
Jiménez-Castilla L, Opazo-Ríos L, Marin-Royo G, Orejudo M, Rodrigues-Diez R, Ballesteros-Martínez C, Soto-Catalán M, Caro-Ordieres T, Artaiz I, Suarez-Cortés T, et al. The Synthetic Flavonoid Hidrosmin Improves Endothelial Dysfunction and Atherosclerotic Lesions in Diabetic Mice. Antioxidants. 2022; 11(12):2499. https://doi.org/10.3390/antiox11122499
Chicago/Turabian StyleJiménez-Castilla, Luna, Lucas Opazo-Ríos, Gema Marin-Royo, Macarena Orejudo, Raquel Rodrigues-Diez, Constanza Ballesteros-Martínez, Manuel Soto-Catalán, Teresa Caro-Ordieres, Inés Artaiz, Tatiana Suarez-Cortés, and et al. 2022. "The Synthetic Flavonoid Hidrosmin Improves Endothelial Dysfunction and Atherosclerotic Lesions in Diabetic Mice" Antioxidants 11, no. 12: 2499. https://doi.org/10.3390/antiox11122499