
Prevention of Testicular Damage by Indole Derivative MMINA via Upregulated StAR and CatSper Channels with Coincident Suppression of Oxidative Stress and Inflammation: In Silico and In Vivo Validation

 , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Synthesis of MMINA
2.3. Drug Preparation
2.4. Acute Toxicity Testing of DMFM
2.5. Animals
2.6. Experimental Design and Drug Administration
- Group 1: Control group, received isotonic saline (i.p.);
- Group 2: DMSO group, received 1% DMSO (i.p.);
- Group 3: CDDP group, received a single dose of CDDP (12 mg/kg, i.p.) on day 8 of the experiment;
- Group 4: MMINA + CDDP group received MMINA (25 mg/kg, i.p.) daily dose for 2 weeks and a single dose of cisplatin (12 mg/kg, i.p.) on the 8th day of the experiment, 1 h after the dose of MMINA;
- Group 5: MMINA group, received MMINA (25 mg/kg, i.p.) for 14 days.
2.7. Sample Collection and Preparation
2.8. Daily Sperm Count and Motility
2.9. Hormone Analysis
2.10. Antioxidant Status and Oxidative Stress Markers
2.11. Gene Expression Analysis
2.12. Extraction of Protein and Western Blot Analysis
2.13. Histopathology
2.14. Morphometry and Planimetry
2.15. Molecular Docking
2.16. Pathway Construction
2.17. Statistics
3. Results
3.1. Compound Synthesis
3.2. Testis and Epididymis Weight after Comparative Treatment Using MMINA and CDDP
3.3. Daily Sperm Production and Sperm Number in Various Parts of Epididymis
3.4. CDDP Induced Deregulation of Sex Hormones and Testosterone Biosynthesis Pathway
3.5. Sperm Motility and Epididymis CatSper Channel Activation
3.6. MMINA Attenuated CDDP-Induced Oxidative Stress by Triggering Antioxidant Production
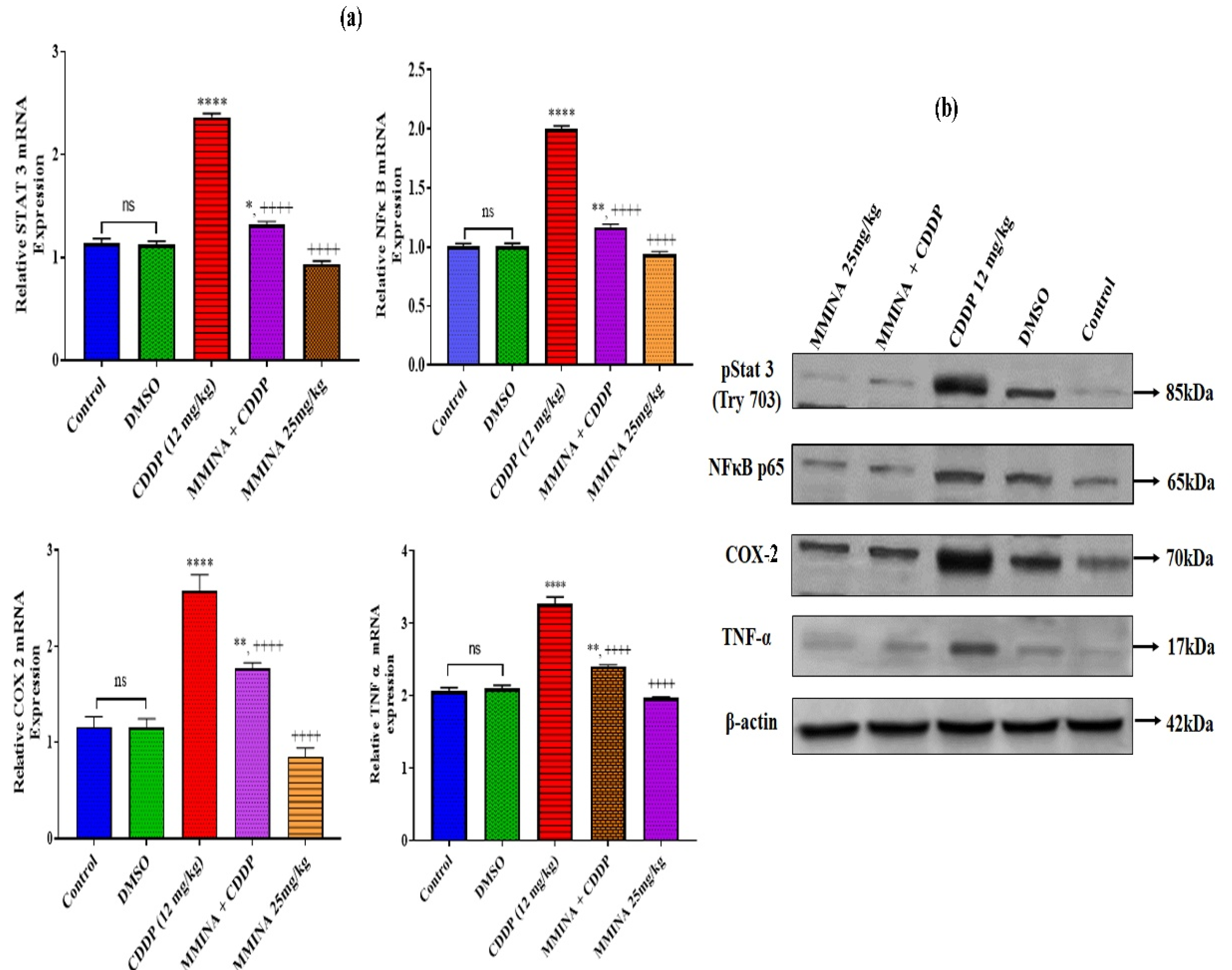
3.7. MMINA Inhibited Activation of Inflammatory Pathways in CDDP-Exposed Testicular Tissue
3.8. MMINA Attenuated CDDP-Induced Drop in Spermatogenesis and Testicular Impairments
3.9. Molecular Interaction of TNF-α, STAT3, COX-2, and NF-κβ with MMINA and Cisplatin
3.10. MMINA Modulated Molecular Interactions in Reducing Testicular Inflammation
3.11. Molecular Interaction of 3β-HSD, CatSper, CYP20A1, and StAR with MMINA and CDDP
3.12. MMINA Modulated Molecular Interactions in Promoting Testicular Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santabarbara, G.; Maione, P.; Rossi, A.; Gridelli, C. Pharmacotherapeutic options for treating adverse effects of Cisplatin chemotherapy. Expert Opin. Pharmacother. 2016, 17, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Quintanilha, J.C.F.; De Sousa, V.M.; Visacri, M.B.; Amaral, L.S.; Santos, R.M.M.; Zambrano, T.; Salazar, L.A.; Moriel, P. Involvement of cytochrome P450 in cisplatin treatment: Implications for toxicity. Cancer Chemother. Pharmacol. 2017, 80, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Cherry, S.M.; Hunt, P.A.; Hassold, T.J. Cisplatin disrupts mammalian spermatogenesis, but does not affect recombination or chromosome segregation. Mutat. Res. 2004, 564, 115–128. [Google Scholar] [CrossRef]
- Türk, G.; Ateşşahin, A.; Sönmez, M.; Çeribaşi, A.O.; Yüce, A. Improvement of cisplatin-induced injuries to sperm quality, the oxidant-antioxidant system, and the histologic structure of the rat testis by ellagic acid. Fertil. Steril. 2008, 89, 1474–1481. [Google Scholar] [CrossRef] [Green Version]
- Howell, S.J.; Shalet, S.M. Spermatogenesis after cancer treatment: Damage and recovery. JNCi Monogr. 2005, 2005, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Schilsky, R.L.; Lewis, B.J.; Sherins, R.J.; Young, R.C. Gonadal Dysfunction in Patients Receiving Chemotherapy for Cancer. Ann. Intern. Med. 1980, 93, 109. [Google Scholar] [CrossRef]
- Garcia, J.M.; Chen, J.-A.; Guillory, B.; Donehower, L.A.; Smith, R.G.; Lamb, D.J. Ghrelin Prevents Cisplatin-Induced Testicular Damage by Facilitating Repair of DNA Double Strand Breaks Through Activation of p53 in Mice. Biol. Reprod. 2015, 93, 24. [Google Scholar] [CrossRef]
- Martin, R.H.; Ernst, S.; Rademaker, A.; Barclay, L.; Ko, E.; Summers, N. Analysis of Sperm Chromosome Complements before, during, and after Chemotherapy. Cancer Genet. Cytogenet. 1999, 108, 133–136. [Google Scholar] [CrossRef]
- Kohsaka, T.; Minagawa, I.; Morimoto, M.; Yoshida, T.; Sasanami, T.; Yoneda, Y.; Ikegaya, N.; Sasada, H. Efficacy of relaxin for cisplatin-induced testicular dysfunction and epididymal spermatotoxicity. Basic Clin. Androl. 2020, 30, 3–13. [Google Scholar] [CrossRef]
- Sherwood, O.D. Relaxin’s Physiological Roles and Other Diverse Actions. Endocr. Rev. 2004, 25, 205–234. [Google Scholar] [CrossRef] [Green Version]
- Stocco, D.M. Clinical disorders associated with abnormal cholesterol transport: Mutations in the steroidogenic acute regulatory protein. Mol. Cell. Endocrinol. 2002, 191, 19–25. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenic enzymes. In Disorders of the Human Adrenal Cortex; Karger: Basel, Switzerland, 2008; Volume 13, pp. 1–18. [Google Scholar]
- Liu, P.Y.; Veldhuis, J.D. Hypothalamo-pituitary unit, testis, and male accessory organs. In Yen and Jaffe’s Reproductive Endocrinology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 285–300.e8. [Google Scholar]
- Tian, M.; Liu, F.; Liu, H.; Zhang, Q.; Li, L.; Hou, X.; Zhao, J.; Li, S.; Chang, X.; Sun, Y. Grape seed procyanidins extract attenuates Cisplatin-induced oxidative stress and testosterone synthase inhibition in rat testes. Syst. Biol. Reprod. Med. 2018, 64, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Favareto, A.P.A.; Fernandez, C.D.B.; da Silva, D.A.F.; Anselmo-Franci, J.A.; Kempinas, W.D.G. Persistent impairment of testicular histology and sperm motility in adult rats treated with cisplatin at peri-puberty. Basic Clin. Pharmacol. Toxicol. 2011, 109, 85–96. [Google Scholar] [CrossRef]
- Wang, T.-E.; Lai, Y.-H.; Yang, K.-C.; Lin, S.-J.; Chen, C.-L.; Tsai, P.-S. Counteracting Cisplatin-Induced Testicular Damages by Natural Polyphenol Constituent Honokiol. Antioxidants 2020, 9, 723. [Google Scholar] [CrossRef]
- Strünker, T.; Goodwin, N.; Brenker, C.; Kashikar, N.D.; Weyand, I.; Seifert, R.; Kaupp, U.B. The CatSper channel mediates progesterone-induced Ca2+ influx in human sperm. Nature 2011, 471, 382–386. [Google Scholar] [CrossRef]
- Sun, X.-H.; Zhu, Y.-Y.; Wang, L.; Liu, H.-L.; Ling, Y.; Li, Z.-L.; Sun, L.-B. The Catsper channel and its roles in male fertility: A systematic review. Reprod. Biol. Endocrinol. 2017, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.P.; Rajender, S. CatSper channel, sperm function and male fertility. Reprod. Biomed. Online 2015, 30, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Burchenal, J.H.; Kalaher, K.; Dew, K.; Lokys, L. Rationale for development of platinum analogs. Cancer Treat. Rep. 1979, 63, 1493–1498. [Google Scholar]
- Afsar, T.; Razak, S.; Aldisi, D.; Shabbir, M.; Almajwal, A.; Al Kheraif, A.A.; Arshad, M. Acacia hydaspica R. Parker ethyl-acetate extract abrogates cisplatin-induced nephrotoxicity by targeting ROS and inflammatory cytokines. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Ijaz, M.U.; Ashraf, A.; Ahmed, A.; Ismail, H.; Muzzamil, S.; Samad, A.; Al-Ghanim, K.; Al-Misned, F.; Ahmed, Z.; Mahboob, S. Remedial effects of casticin as an antioxidant on cisplatin induced oxidative damage in rat liver. J. King Saud Univ. Sci. 2020, 32, 1100–1105. [Google Scholar] [CrossRef]
- Harakeh, S.; Qari, Y.; Tashkandi, H.; Almuhayawi, M.; Saber, S.H.; Aljahdali, E.; El-Shitany, N.; Shaker, S.; Lucas, F.; Al Amri, T.; et al. Thymoquinone nanoparticles protect against cisplatin-induced nephrotoxicity in Ehrlich carcinoma model without compromising cisplatin anti-cancer efficacy. J. King Saud Univ. Sci. 2022, 34, 101675. [Google Scholar] [CrossRef]
- Afsar, T.; Razak, S.; Khan, M.R.; Almajwal, A. Acacia hydaspica ethyl acetate extract protects against cisplatin-induced DNA damage, oxidative stress and testicular injuries in adult male rats. BMC Cancer 2017, 17, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, C.K.; Indalkar, K.S.; Patil, C.R.; Goyal, S.N.; Chaturbhuj, G.U. Novel 2-phenyl-4, 5, 6, 7-tetrahydro [b] benzothiophene analogues as selective COX-2 inhibitors: Design, synthesis, anti-inflammatory evaluation, and molecular docking studies. Med. Chem. Lett. 2017, 27, 1721–1726. [Google Scholar] [CrossRef] [PubMed]
- Lamie, P.F.; Ali, W.A.; Bazgier, V.; Rárová, L. Novel N-substituted indole Schiff bases as dual inhibitors of cyclooxygenase-2 and 5-lipoxygenase enzymes: Synthesis, biological activities in vitro and docking study. Eur. J. Med. Chem. 2016, 123, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Abdellatif, K.R.A.; Lamie, P.F.; Omar, H. 3-Methyl-2-phenyl-1-substituted-indole derivatives as indomethacin analogs: Design, synthesis and biological evaluation as potential anti-inflammatory and analgesic agents. J. Enzyme Inhib. Med. Chem. 2016, 31, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.E. Pharmacology of penile erection. Pharmacol. Rev. 2001, 53, 417–450. [Google Scholar]
- Bhat, M.A.; Al-Omar, M.A.; Alsaif, N.A.; Almehizia, A.A.; Naglah, A.M.; Razak, S.; Khan, A.A.; Ashraf, N.M.J. Novel sulindac derivatives: Synthesis, characterisation, evaluation of antioxidant, analgesic, anti-inflammatory, ulcerogenic and COX-2 inhibition activity. J. Enzym. Inhib. Med. Chem. 2020, 35, 921–934. [Google Scholar] [CrossRef]
- Razak, S.; Afsar, T.; Bibi, N.; Abulmeaty, M.; Qamar, W.; Almajwal, A.; Inam, A.; Al Disi, D.; Shabbir, M.; Bhat, M.A. Molecular docking, pharmacokinetic studies, and in vivo pharmacological study of indole derivative 2-(5-methoxy-2-methyl-1H-indole-3-yl)-N′-[(E)-(3-nitrophenyl) methylidene] acetohydrazide as a promising chemoprotective agent against cisplatin induced organ damage. Sci. Rep. 2021, 11, 6245. [Google Scholar]
- Bhat, M.A.; Al-Omar, M.A.; Raish, M.; Ansari, M.A.; Abuelizz, H.A.; Bakheit, A.H.; Naglah, A.M. Indole Derivatives as Cyclooxygenase Inhibitors: Synthesis, Biological Evaluation and Docking Studies. Molecules 2018, 23, 1250. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.; Mahmoud-Ghoneim, D.; I Syam, M.; Daoud, S. Neural network assessment of herbal protection against chemotherapeutic-induced reproductive toxicity. Theor. Biol. Med Model. 2012, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Coşkun, N.; Hatipoğlu, M.T.; Özoğul, C.; Korkmaz, C.; Akyol, S.N.; Mıcılı, S.C.; Arık, G.S.; Erdoğan, D.J. The protective effects of acetyl L-carnitine on testis gonadotoxicity induced by cisplatin in rats. Balk. Med. J. 2013, 30, 235. [Google Scholar] [CrossRef]
- OECD. Test No. 425: Acute Oral Toxicity: Up-and-Down Procedure. In OECD Guidelines for the Testing of Chemicals; Section 4; OECD Publishing: Paris, France, 2001; Volume 2, pp. 12–16. [Google Scholar]
- Molina, A.; Moyano, M.; Serrano-Rodríguez, J.M.; Ayala, N.; Lora, A.; Serrano-Caballero, J. Analyses of anaesthesia with ketamine combined with different sedatives in rats. Vet. Med. 2015, 60, 368–375. [Google Scholar] [CrossRef]
- Ullah, H.; Ambreen, A.; Ahsan, N.; Jahan, S. Bisphenol S induces oxidative stress and DNA damage in rat spermatozoa in vitro and disrupts daily sperm production in vivo. Toxicol. Environ. Chem. 2017, 99, 953–965. [Google Scholar] [CrossRef]
- Soni, K.K.; Kim, H.K.; Choi, B.R.; Karna, K.K.; You, J.H.; Cha, J.S.; Shin, Y.S.; Lee, S.W.; Kim, C.Y.; Park, J.K. Dose-dependent effects of cisplatin on the severity of testicular injury in Sprague Dawley rats: Reactive oxygen species and endoplasmic reticulum stress. Drug Des. Dev. Ther. 2016, 10, 3959–3968. [Google Scholar] [CrossRef] [Green Version]
- Afsar, T.; Razak, S.; Almajwal, A.; Khan, M.R. Acacia hydaspica R. Parker ameliorates cisplatin induced oxidative stress, DNA damage and morphological alterations in rat pulmonary tissue. BMC Complement. Altern. Med. 2018, 18, 49. [Google Scholar] [CrossRef] [Green Version]
- Afsar, T.; Razak, S.; Almajwal, A. Effect of Acacia hydaspica R. Parker extract on lipid peroxidation, antioxidant status, liver function test and histopathology in doxorubicin treated rats. Lipids Heal. Dis. 2019, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Kruger, N.J. The Bradford method for protein quantitation. In Basic Protein and Peptide Protocols; Springer: Berlin/Heidelberg, Germany, 1994; pp. 9–15. [Google Scholar]
- Falvo, S.; Baccaria, G.C.; Spaziano, G.; Rosati, L.; Venditti, M.; Di Fiore, M.M.; Santillo, A. StAR protein and steroidogenic enzyme expressions in the rat Harderian gland. Comptes Rendus. Biol. 2018, 341, 160–166. [Google Scholar] [CrossRef]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/β-catenin signaling pathway. BMC Cancer 2018, 18, 1043. [Google Scholar] [CrossRef] [Green Version]
- Afsar, T.; Trembley, J.; Salomon, C.; Razak, S.; Khan, M.R.; Ahmed, K. Growth inhibition and apoptosis in cancer cells induced by polyphenolic compounds of Acacia hydaspica: Involvement of multiple signal transduction pathways. Sci. Rep. 2016, 6, 23077. [Google Scholar] [CrossRef] [Green Version]
- Trembley, J.H.; Unger, G.M.; Tobolt, D.K.; Korman, V.L.; Wang, G.; Ahmad, K.A.; Slaton, J.W.; Kren, B.T.; Ahmed, K. Systemic administration of antisense oligonucleotides simultaneously targeting CK2α and α′ subunits reduces orthotopic xenograft prostate tumors in mice. Mol. Cell. Biochem. 2011, 356, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef]
- David, A.; Islam, S.; Tankhilevich, E.; Sternberg, M.J. The AlphaFold Database of Protein Structures: A Biologist’s Guide. J. Mol. Biol. 2022, 434, 167336. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Kerwin, S.M. ChemBioOffice Ultra 2010 Suite; ACS Publications: Washington, DC, USA, 2010. [Google Scholar]
- Liu, Y.; Grimm, M.; Dai, W.-T.; Hou, M.-C.; Xiao, Z.-X.; Cao, Y. CB-Dock: A web server for cavity detection-guided protein–ligand blind docking. Acta Pharmacol. Sin. 2020, 41, 138–144. [Google Scholar] [CrossRef]
- Zhang, Y.-H.; Zeng, T.; Chen, L.; Huang, T.; Cai, Y.-D. Determining protein–protein functional associations by functional rules based on gene ontology and KEGG pathway. Biochim. et Biophys. Acta (BBA) Proteins Proteom. 2021, 1869, 140621. [Google Scholar] [CrossRef]
- Jiao, X.; Sherman, B.T.; Huang, D.W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A stateful web service to facilitate gene/protein list analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Oualline, S.; Oualline, G. Drawing with Inkscape. In Practical Free Alternatives to Commercial Software; Springer: Berlin/Heidelberg, Germany, 2018; pp. 187–219. [Google Scholar]
- Sabaawy, H.E.; Ryan, B.M.; Khiabanian, H.; Pine, S.R. JAK/STAT of all trades: Linking inflammation with cancer development, tumor progression and therapy resistance. Carcinogenesis 2021, 42, 1411–1419. [Google Scholar] [CrossRef]
- Yang, L.; Liu, R.; Fang, Y.; He, J. Anti-inflammatory effect of phenylpropanoids from Dendropanax dentiger in TNF-α-induced MH7A cells via inhibition of NF-κB, Akt and JNK signaling pathways. Int. Immunopharmacol. 2021, 94, 107463. [Google Scholar] [CrossRef]
- Ren, X.; Han, L.; Li, Y.; Zhao, H.; Zhang, Z.; Zhuang, Y.; Zhong, M.; Wang, Q.; Ma, W.; Wang, Y. Isorhamnetin attenuates TNF-α-induced inflammation, proliferation, and migration in human bronchial epithelial cells via MAPK and NF-κB pathways. Anat. Rec. 2021, 304, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Durdiakova, J.; Ostatnikova, D.; Celec, P. Testosterone and its metabolites—Modulators of brain functions. Acta Neurobiol. Exp. 2011, 71, 434–454. [Google Scholar]
- Dalian, D.; Haiyan, J.; Yong, F.; Salvi, R.; Someya, S.; Tanokura, M. Ototoxic effects of carboplatin in organotypic cultures in chinchillas and rats. J. Otol. 2012, 7, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Almeer, R.S.; Abdel Moneim, A.E.J. Evaluation of the protective effect of olive leaf extract on cisplatin-induced testicular damage in rats. Oxidative Med. Cell. Longev. 2018, 2018, 8487248. [Google Scholar] [CrossRef]
- Adejuwon, S.; Femi-Akinlosotu, O.; Omirinde, J.J. Cisplatin-induced testicular dysfunction and its amelioration by Launaea taraxacifolia leaf extract. Andrologia 2015, 47, 553–559. [Google Scholar] [CrossRef]
- Leblanc, G.A.; Kantoff, P.W.; Ng, S.F.; Frei, E., III; Waxman, D.J.J. Hormonal perturbations in patients with testicular cancer treated with cisplatin. Cancer 1992, 69, 2306–2310. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Dai, D.-Z.; Zhang, C.; Dai, Y. Apocynin and raisanberine alleviate intermittent hypoxia induced abnormal StAR and 3β-HSD and low testosterone by suppressing endoplasmic reticulum stress and activated p66Shc in rat testes. Reprod. Toxicol. 2013, 36, 60–70. [Google Scholar] [CrossRef]
- García, M.M.S.; Acquier, A.; Suarez, G.; Gomez, N.V.; Gorostizaga, A.; Mendez, C.F.; Paz, C. Cisplatin inhibits testosterone synthesis by a mechanism that includes the action of reactive oxygen species (ROS) at the level of P450scc. Chem. Interactions 2012, 199, 185–191. [Google Scholar] [CrossRef]
- Eid, A.H.; Abdelkader, N.F.; El-Raouf, A.; Ola, M.; Fawzy, H.M.; El-Denshary, E.-E.-D.S.J. Carvedilol alleviates testicular and spermatological damage induced by cisplatin in rats via modulation of oxidative stress and inflammation. Arch. Pharm. Res. 2016, 39, 1693–1702. [Google Scholar] [CrossRef]
- Iman, M.; Araghi, M.; Heidari, T.; Mohammadi, V.J. Melissa officinalis and vitamin E as the potential therapeutic candidates for reproductive toxicity caused by anti-cancer drug, cisplatin, in male rats. Recent Pat. Anticancer Drug Discov. 2017, 12, 73–80. [Google Scholar] [CrossRef]
- Zhu, J.; Li, S.; Zhang, Y.; Ding, G.; Zhu, C.; Huang, S.; Zhang, A.; Jia, Z.; Li, M. COX-2 contributes to LPS-induced Stat3 activation and IL-6 production in microglial cells. Am. J. Transl. Res. 2018, 10, 966–974. [Google Scholar]
- Wang, X.; Stocco, D.M.J. The decline in testosterone biosynthesis during male aging: A consequence of multiple alterations. Mol. Cell. Endocrinol. 2005, 238, 1–7. [Google Scholar] [CrossRef]
- Hassen, M.; Mohamed, H.; Montaser, M.; El-Sharnouby, M.; Awad, N.; Ebiya, R. Molecular, Immunomodulatory, and Histopathological Role of Mesenchymal Stem Cells and Beetroot Extract on Cisplatin Induced Testicular Damage in Albino Rats. Animals 2021, 11, 1142. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Miao, Z.; Yu, F.; Ren, Y.; Yang, J. d,l-Sulforaphane Induces ROS-Dependent Apoptosis in Human Gliomablastoma Cells by Inactivating STAT3 Signaling Pathway. Int. J. Mol. Sci. 2017, 18, 72. [Google Scholar] [CrossRef]
- Sachdeva, H.; Mathur, J.; Guleria, A. Indole derivatives as potential anticancer agents: A review. J. Chil. Chem. Soc. 2020, 65, 4900–4907. [Google Scholar] [CrossRef]
- Amin, A.; Abraham, C.; Hamza, A.A.; Abdalla, Z.A.; Al-Shamsi, S.B.; Harethi, S.S.; Daoud, S. A Standardized Extract of Ginkgo biloba Neutralizes Cisplatin-Mediated Reproductive Toxicity in Rats. J. Biomed. Biotechnol. 2012, 2012, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TREATMENTS | DAILY SPERM PRODUCTION (×106) | CAPUT/CORPUS EPIDIDYMIS SPERM NUMBER (×106/G ORGAN) | CAUDA EPIDIDYMIS SPERM NUMBER (×106/G ORGAN) |
|---|---|---|---|
| CONTROL | 50.65 ± 1.11 | 315.58 ± 5.53 | 546.35 ± 7.06 |
| DMSO VEHICLE | 50.71 ± 1.20 | 316.01 ± 5.82 | 546.90 ± 7.13 |
| CDDP (12 MG/KG) | 27.50 ± 0.98 ** | 138.18 ± 3.83 **** | 217.17 ± 4.45 **** |
| MMINA + CDDP | 47.54 ± 1.01 + | 291.17 ± 4.17 **, ++++ | 520.31 ± 5.97 **, ++++ |
| MMINA (25 MG/KG) | 51.37 ± 1.14 ++ | 319.01 ± 5.91 ++++ | 550.76 ± 6.81 ++++ |
| GROUPS | TBARS (NG/MG) | NO (NG/MG) | SOD (U/G) | GPX (NM/MIN/G) | TAC (URIC ACID EQUIVALENT MM) |
|---|---|---|---|---|---|
| CONTROL | 43.51 ± 2.16 | 21.13 ± 1.01 | 49.67 ±1.20 | 58.25 ± 1.20 | 0.51 ± 0.024 |
| DMSO VEHICLE | 43.98 ± 2.01 | 21.08 ± 1.14 | 49.65 ± 1.09 | 58.17 ± 1.19 | 0.51 ± 0.032 |
| CDDP (12 MG/KG) | 151.50 ± 3.46 **** | 98.54 ± 2.41 **** | 6.01 ± 0.41 **** | 6.75 ± 0.56 **** | 0.30 ± 0.022 **** |
| CDDP + MMINA | 57.67 ± 2.81 *,++++ | 32.69± 2.11 *,++++ | 39.99 ± 1.01 ++++ | 46.01 ±1.05 ++++ | 0.44 ± 0.027 *,+++ |
| MMINA (25 MG/KG) | 42.02 ± 2.37 ++++ | 20.92 ± 1.07 ++++ | 48.69 ± 1.23 ++++ | 59.35 ± 1.11 ++++ | 0.53 ± 0.041 ++++ |
| DRUG | PROTEIN | VINA SCORE |
|---|---|---|
| MMINA | COX-2 | −9.1 |
| TNF-α | −7.6 | |
| NF-κB | −7.3 | |
| STAT3 | −8.6 |
| Drug | Protein | Vina Score |
|---|---|---|
| MMINA | 3β-HSD | −9.6 |
| CatSper | −7.7 | |
| CYP20A1 | −7.6 | |
| StAR | −9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afsar, T.; Razak, S.; Trembley, J.H.; Khan, K.; Shabbir, M.; Almajwal, A.; Alruwaili, N.W.; Ijaz, M.U. Prevention of Testicular Damage by Indole Derivative MMINA via Upregulated StAR and CatSper Channels with Coincident Suppression of Oxidative Stress and Inflammation: In Silico and In Vivo Validation. Antioxidants 2022, 11, 2063. https://doi.org/10.3390/antiox11102063
Afsar T, Razak S, Trembley JH, Khan K, Shabbir M, Almajwal A, Alruwaili NW, Ijaz MU. Prevention of Testicular Damage by Indole Derivative MMINA via Upregulated StAR and CatSper Channels with Coincident Suppression of Oxidative Stress and Inflammation: In Silico and In Vivo Validation. Antioxidants. 2022; 11(10):2063. https://doi.org/10.3390/antiox11102063
Chicago/Turabian StyleAfsar, Tayyaba, Suhail Razak, Janeen H. Trembley, Khushbukhat Khan, Maria Shabbir, Ali Almajwal, Nawaf W. Alruwaili, and Muhammad Umar Ijaz. 2022. "Prevention of Testicular Damage by Indole Derivative MMINA via Upregulated StAR and CatSper Channels with Coincident Suppression of Oxidative Stress and Inflammation: In Silico and In Vivo Validation" Antioxidants 11, no. 10: 2063. https://doi.org/10.3390/antiox11102063