
An In Vitro Study of Saffron Carotenoids: The Effect of Crocin Extracts and Dimethylcrocetin on Cancer Cell Lines

 , , , , , , ,
, , , , , , ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Crocins and Dimethylcrocetin
2.2. Cell Culture and Reagents
2.3. Experimental Setup
2.4. Assessment of Cell Viability
2.5. RNA Extraction and cDNA Synthesis
2.6. Reverse Transcription Real-Time PCR (qRT-PCR)
2.7. Data Analysis and Statistics
3. Results
3.1. LC/Q-TOF/HRMS Analysis
3.2. The Biological Effects on Glioblastoma Cells (A172)
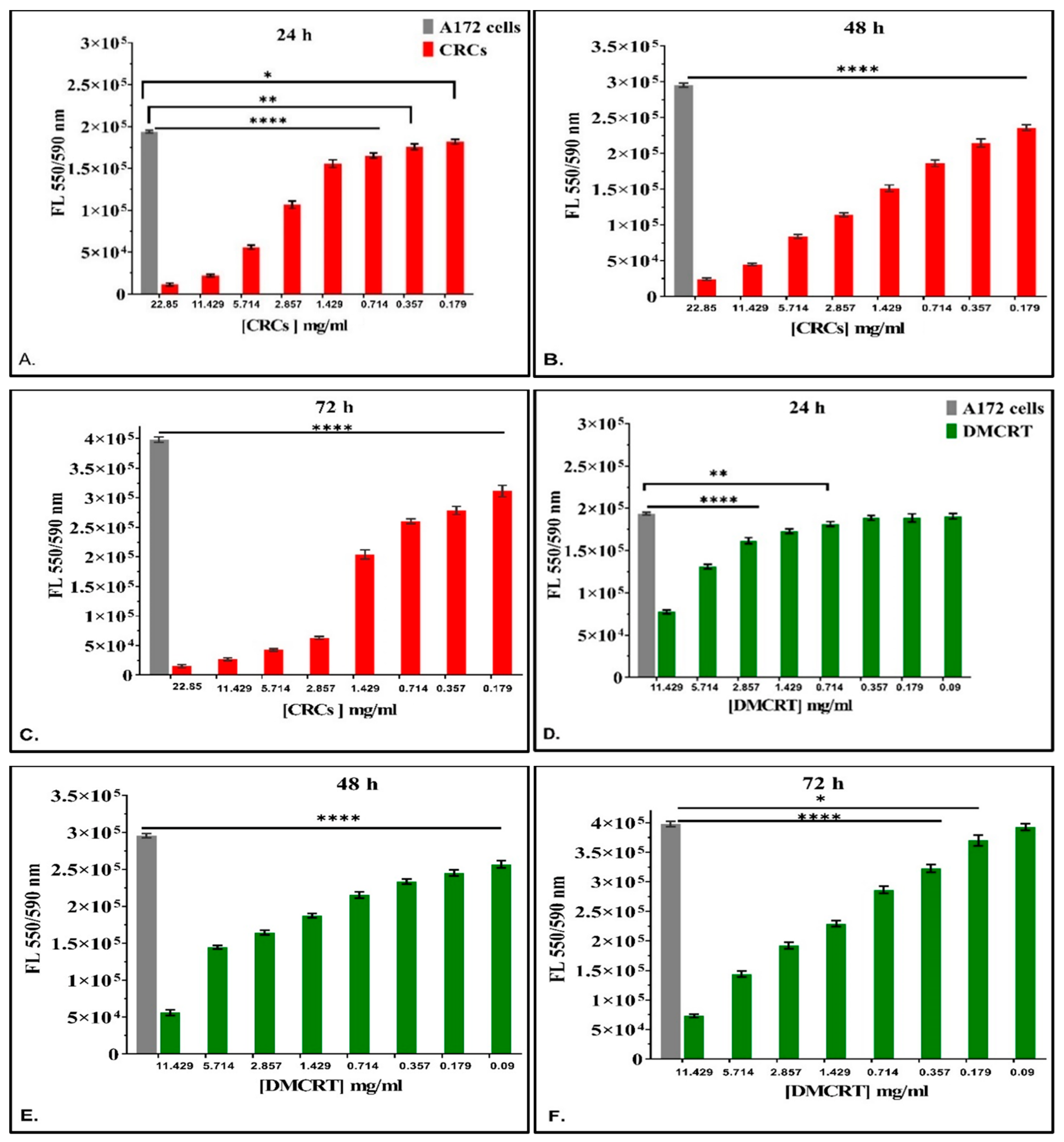
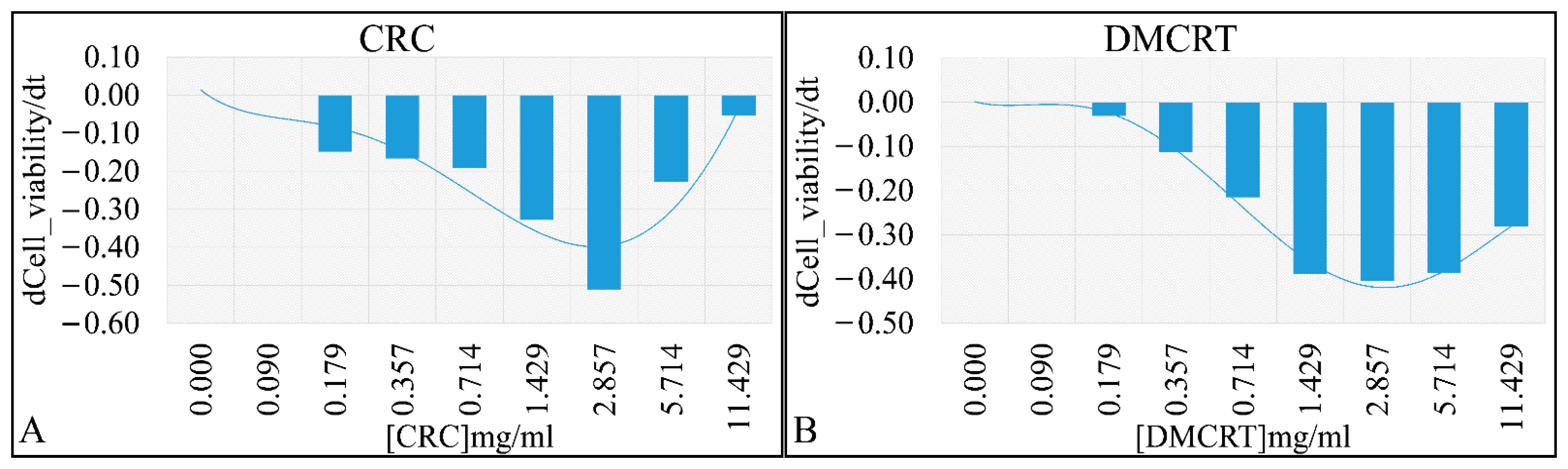
3.2.1. The Dose-Dependent Effect of CRCs and DMCRT on Glioblastoma Cells (A172)
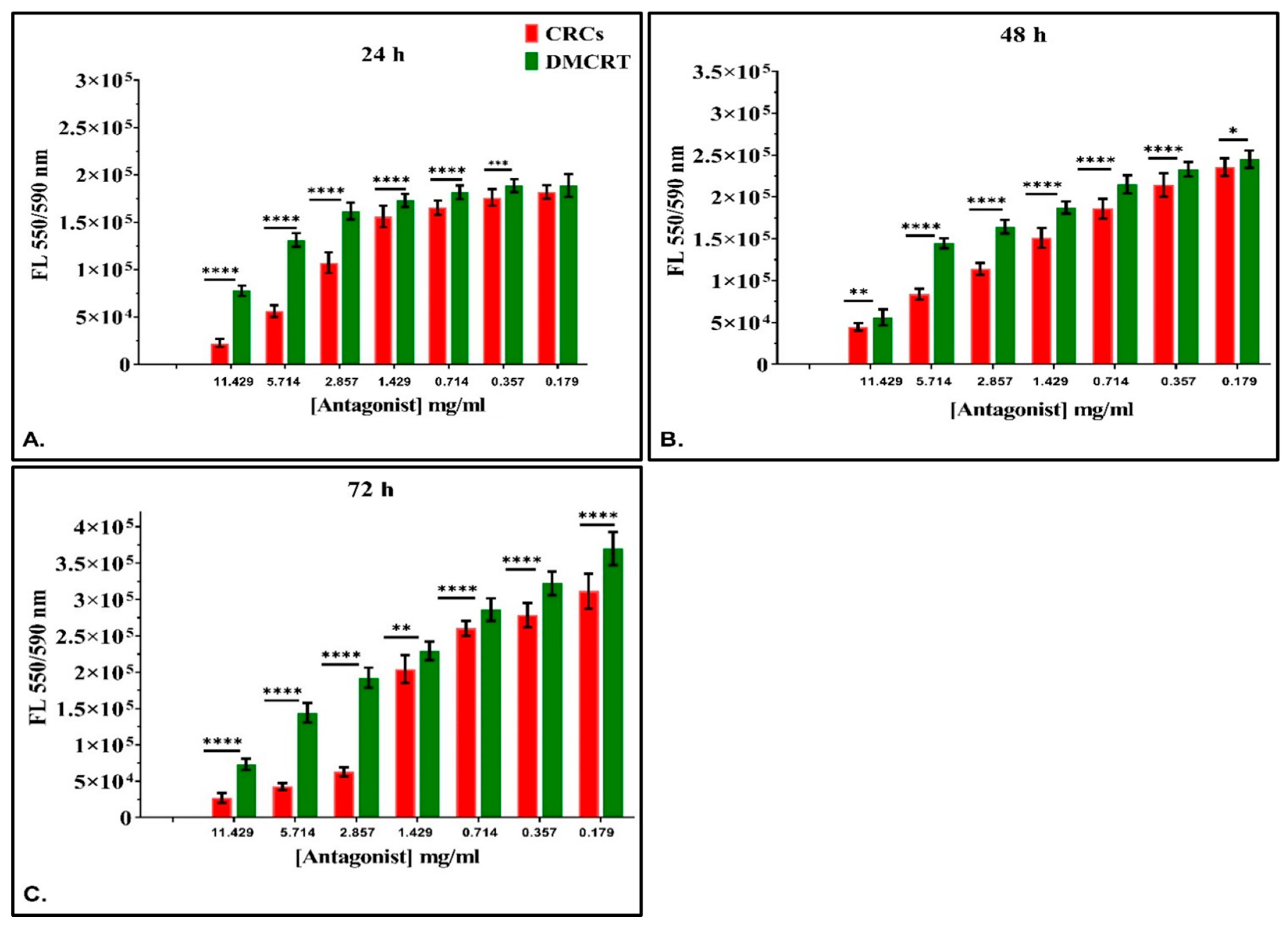
3.2.2. The Comparative Dose-Dependent Effect of CRCs and DMCRT on Glioblastoma Cells (A172)
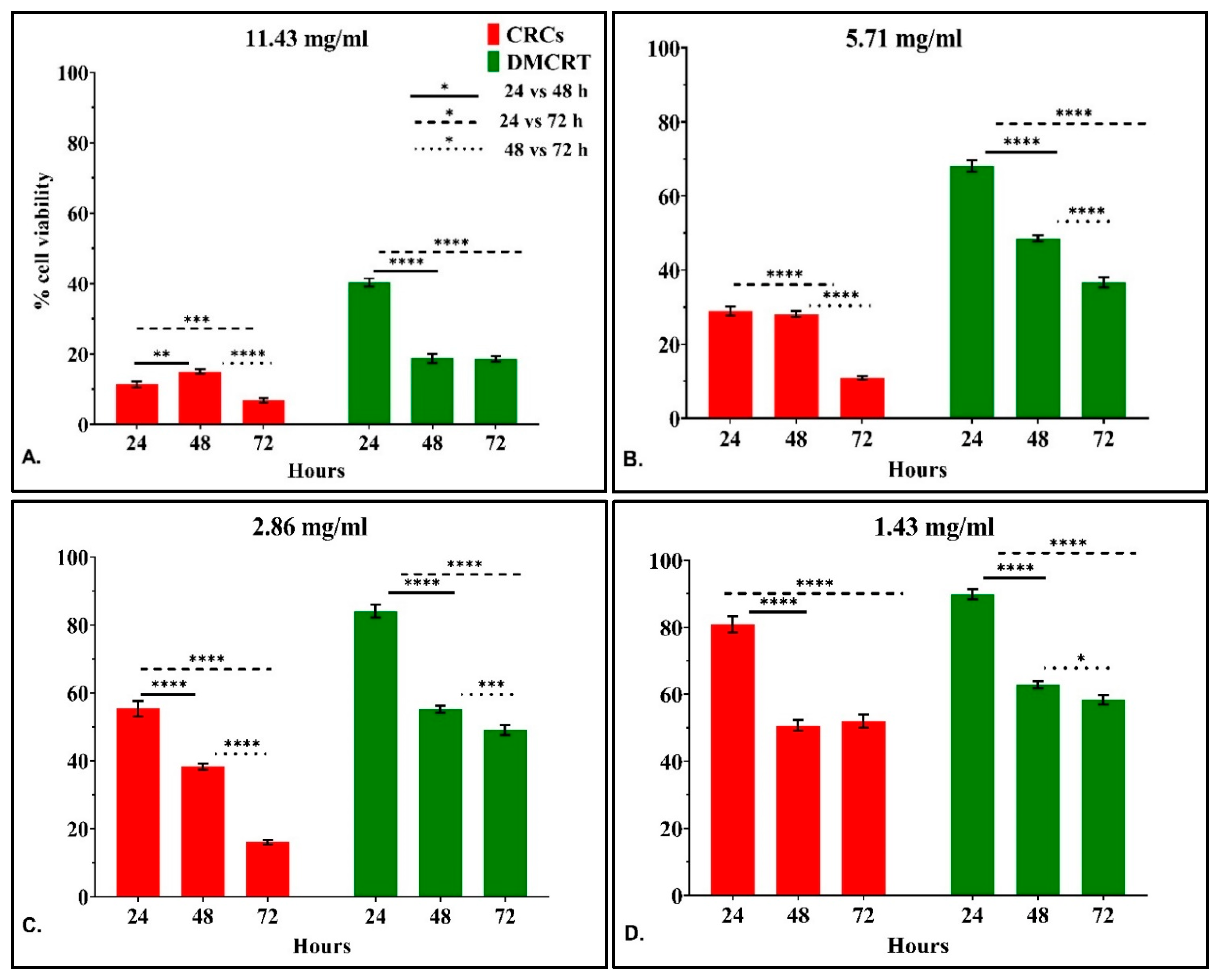
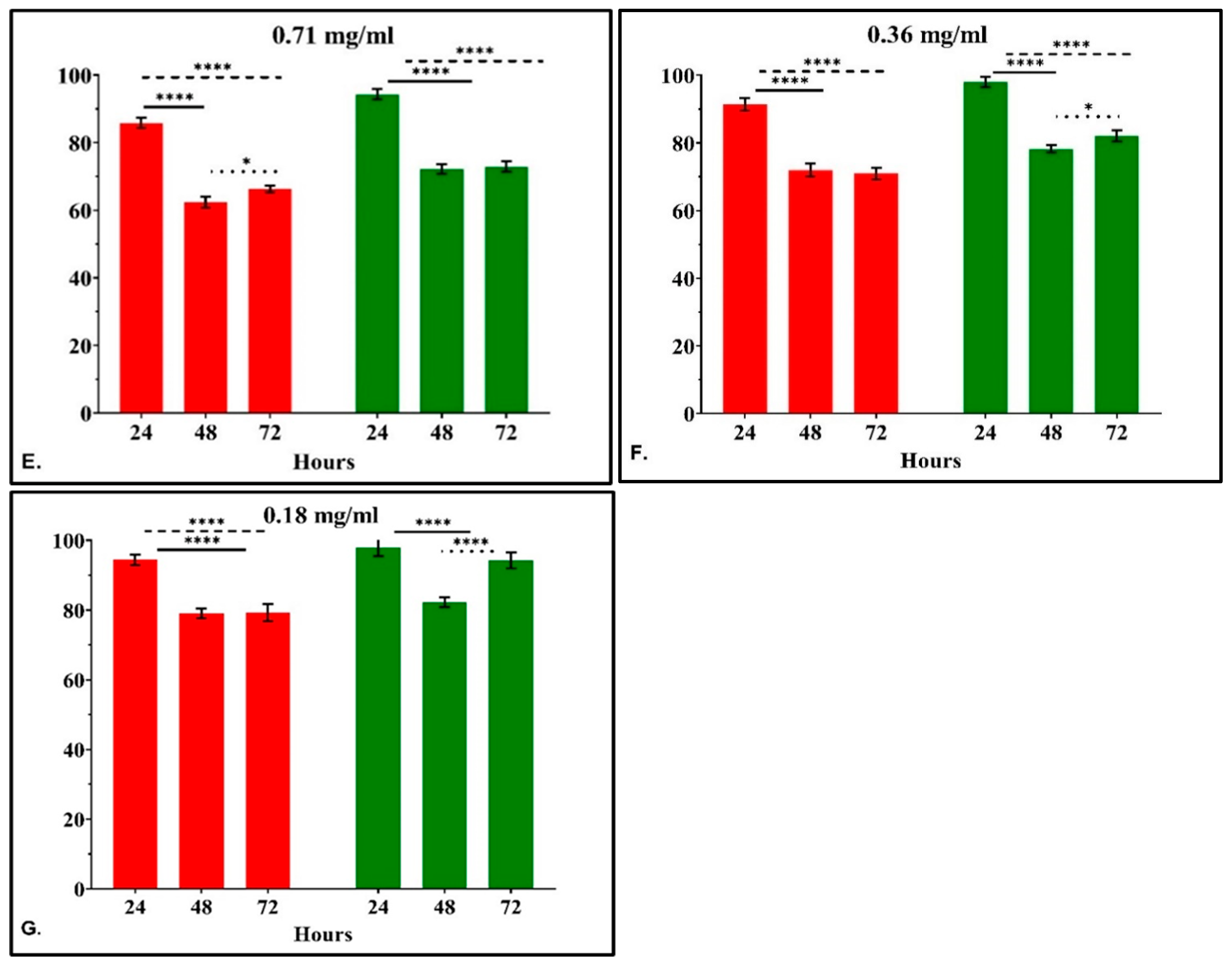
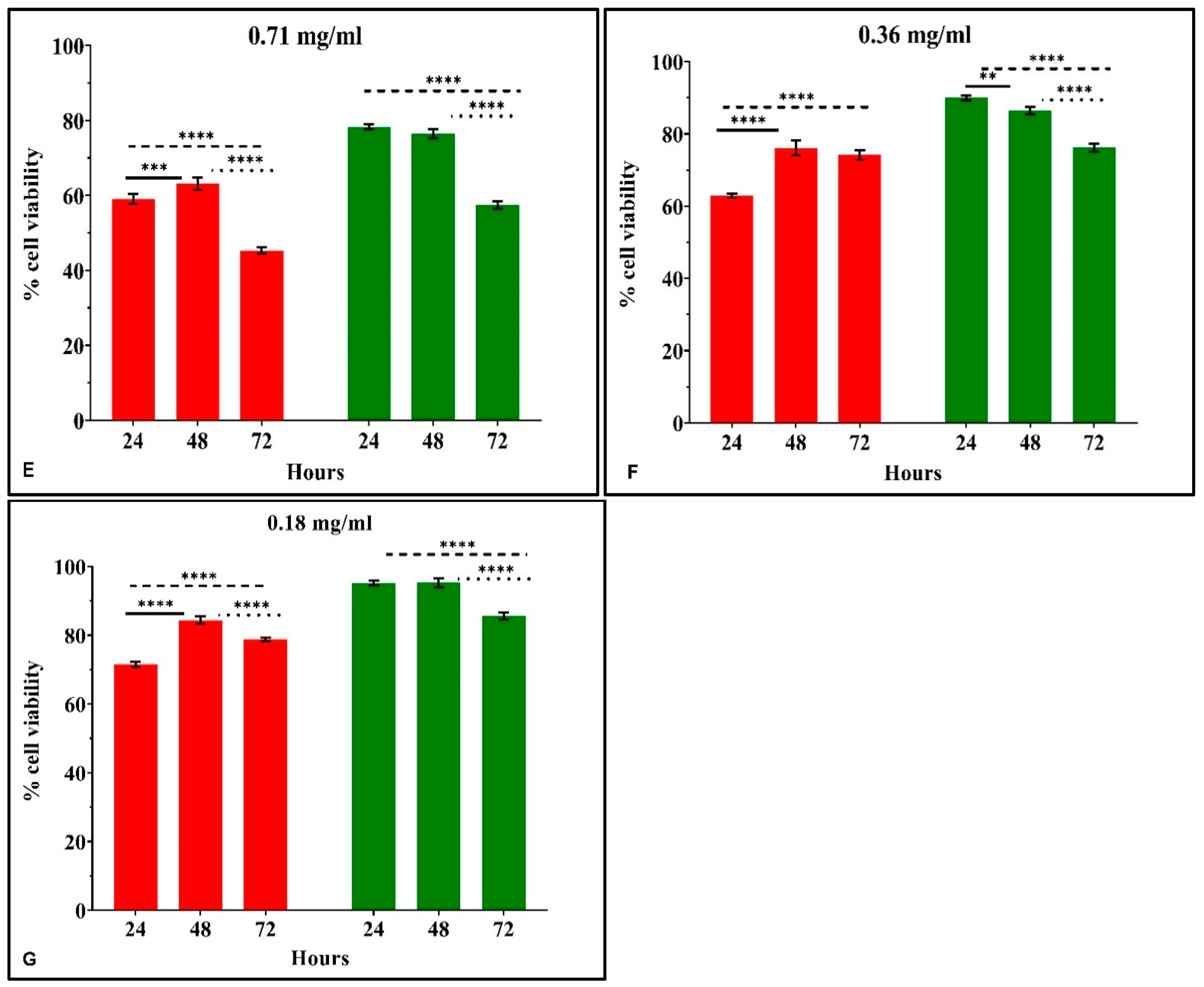
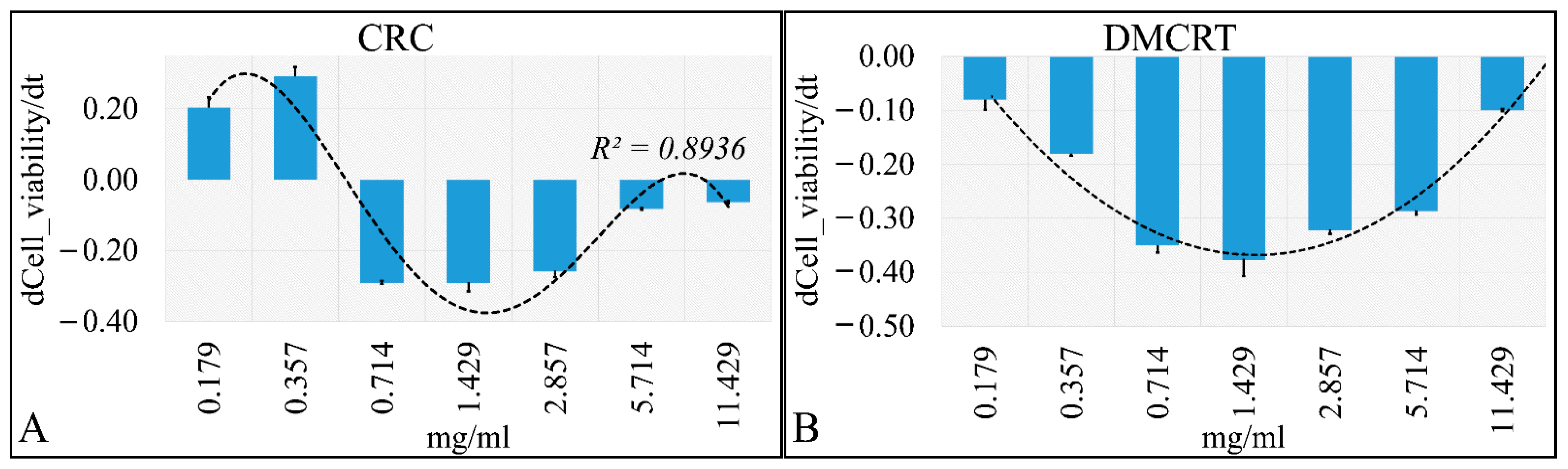
3.2.3. The Time-Dependent Effect of CRCs and DMCRT on Glioblastoma Cells (A172)
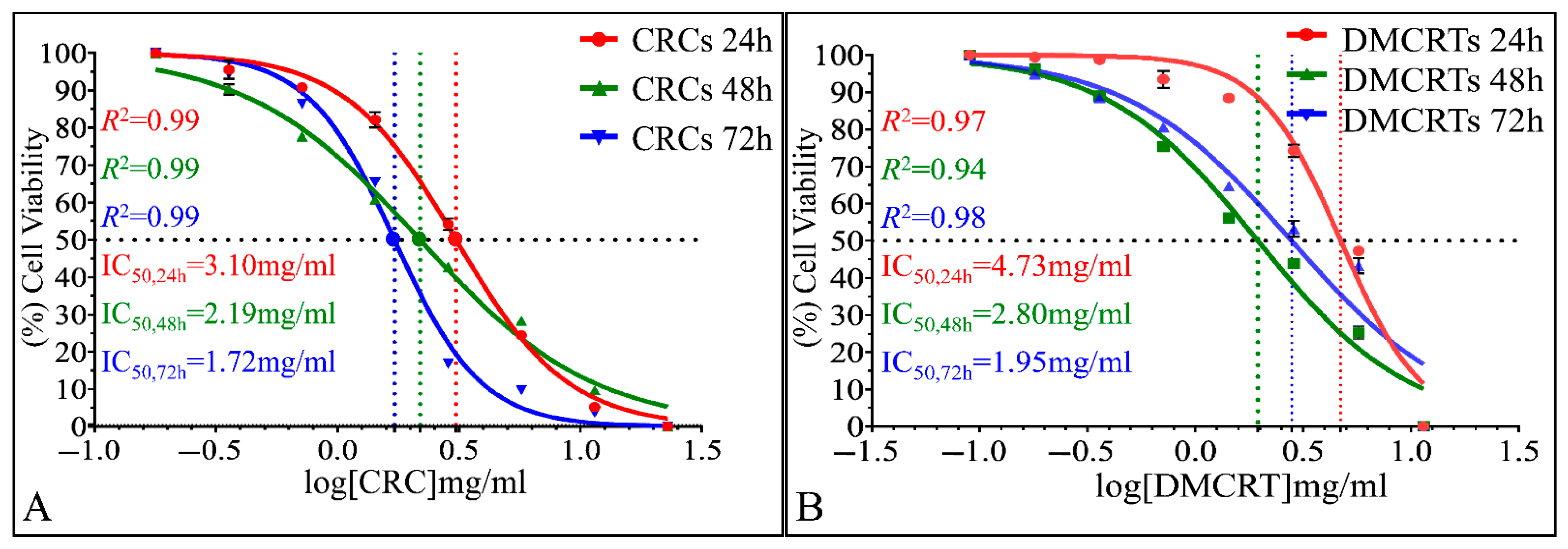
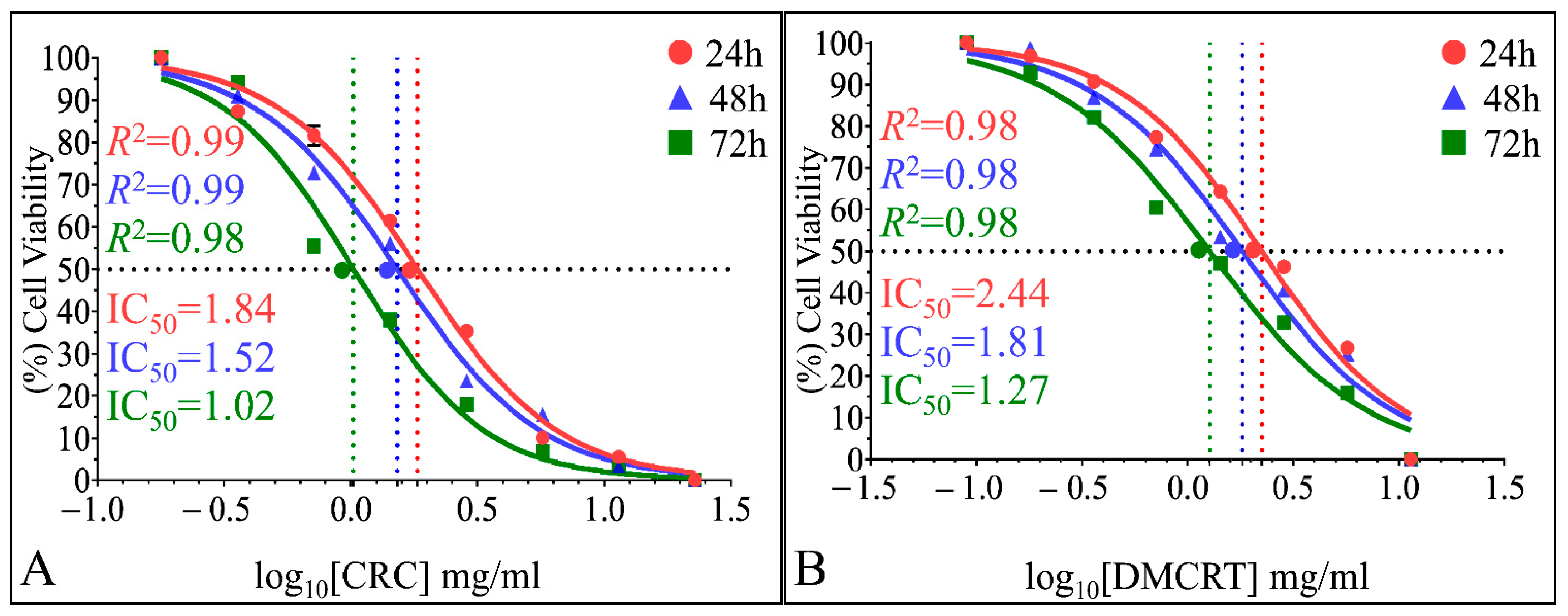
3.2.4. The IC50 Curves of CRCs and DMCRT on Glioblastoma Cells (A172)
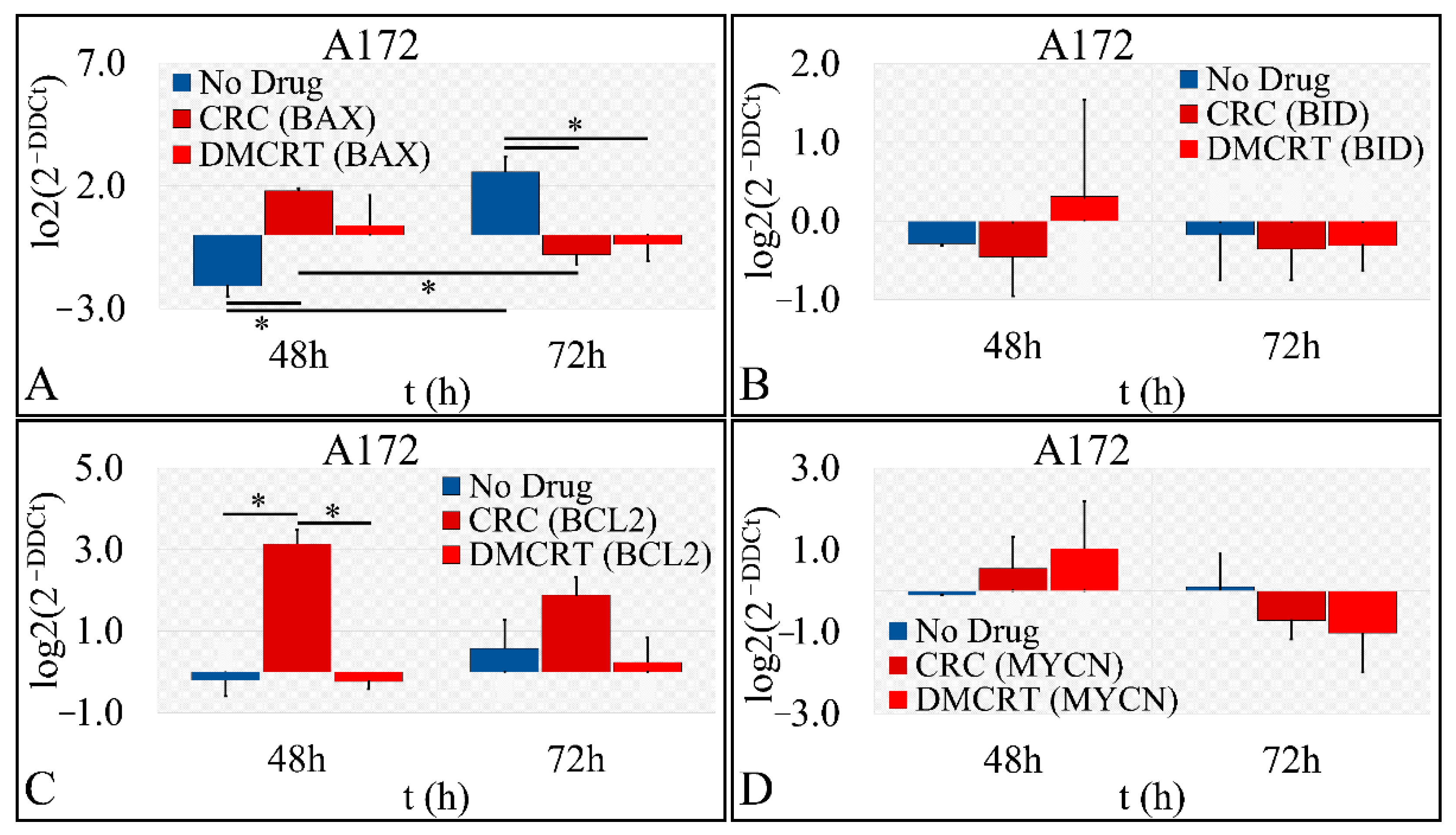
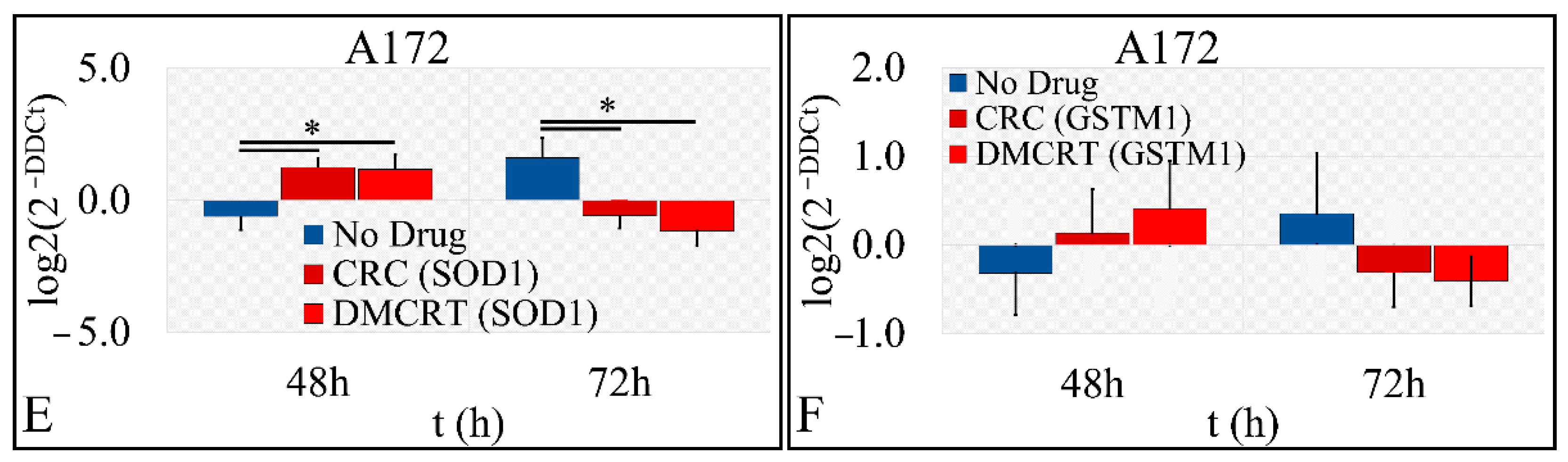
3.2.5. Gene Expression under CRCs and DMCRT on Glioblastoma Cells (A172)
3.3. The Biological Effects on Rhabdomyosarcoma Cells (TE671)
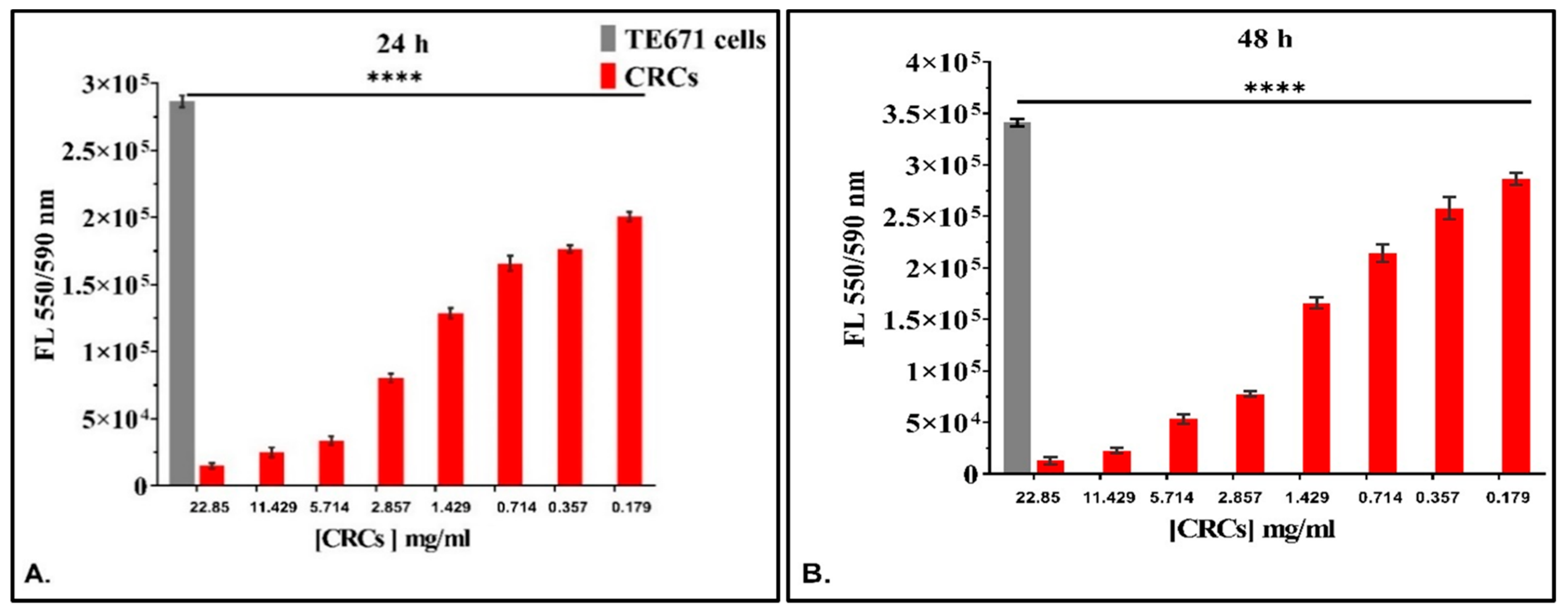
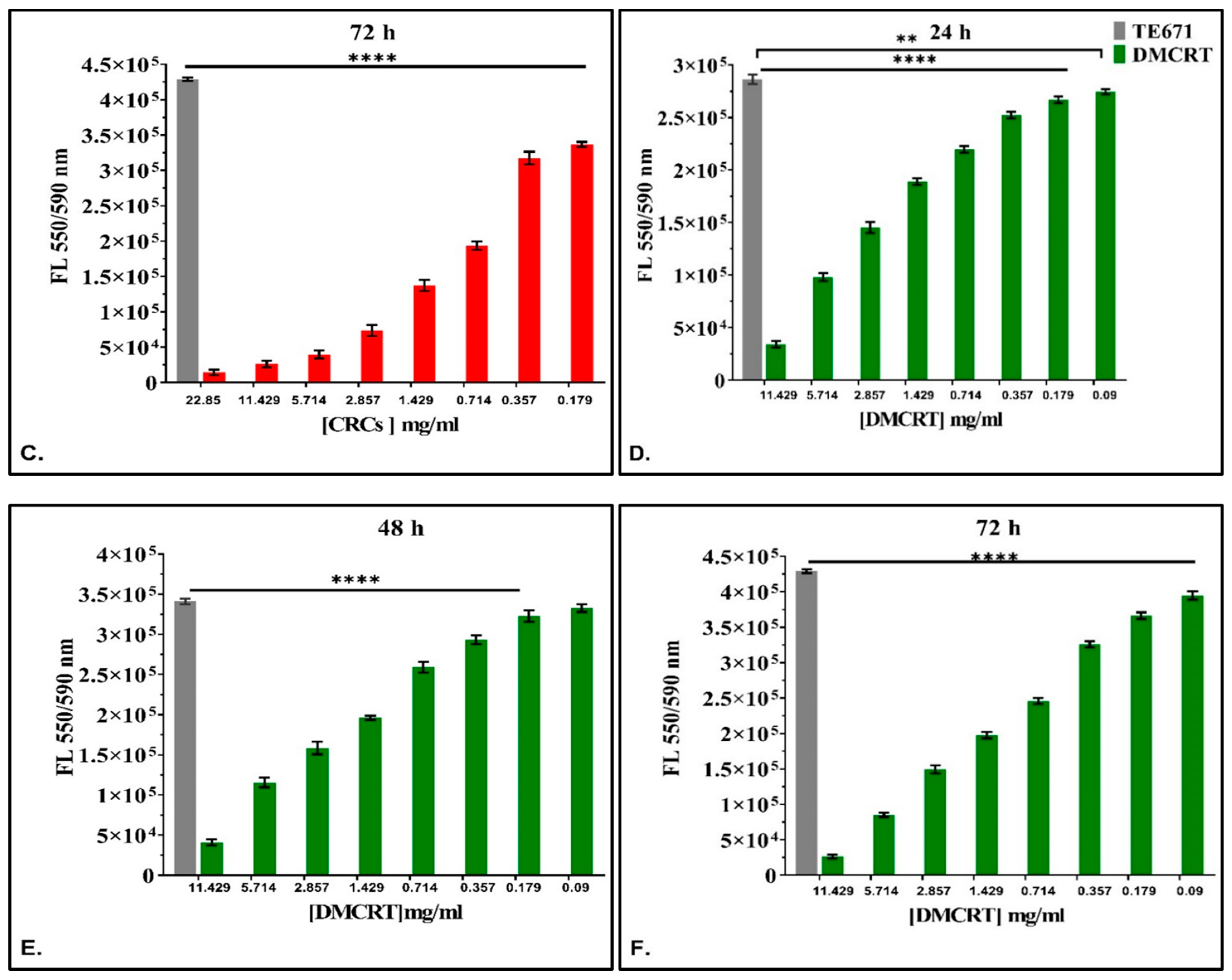
3.3.1. The Dose-Dependent Effect of CRCs and DMCRT on Rhabdomyosarcoma Cells (TE671)
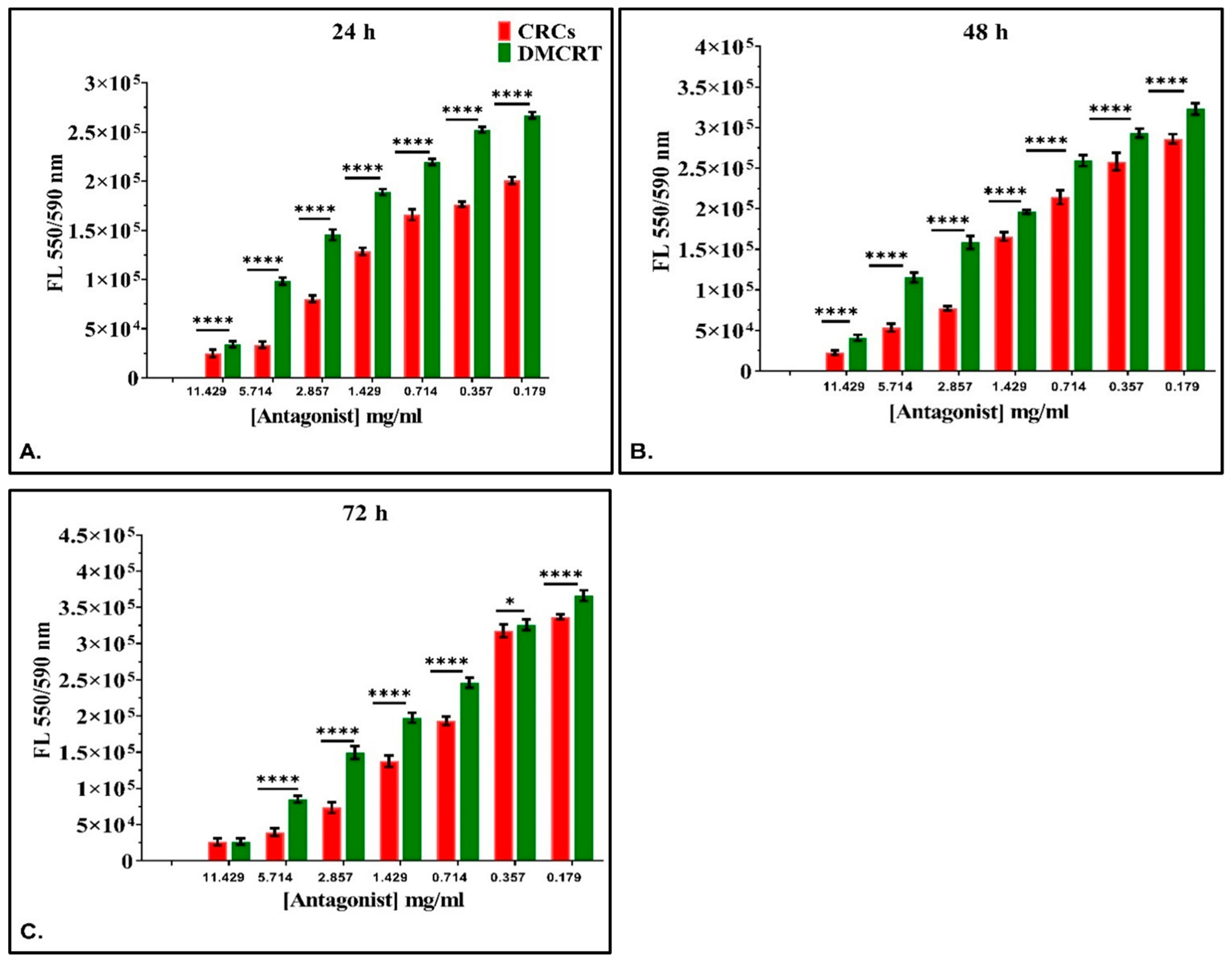
3.3.2. The Comparative Dose-Dependent Effect of CRCs and DMCRT on Rhabdomyosarcoma Cells (TE671)
3.3.3. The Time-Dependent Effect of CRCs and DMCRT on Rhabdomyosarcoma Cells (TE671)
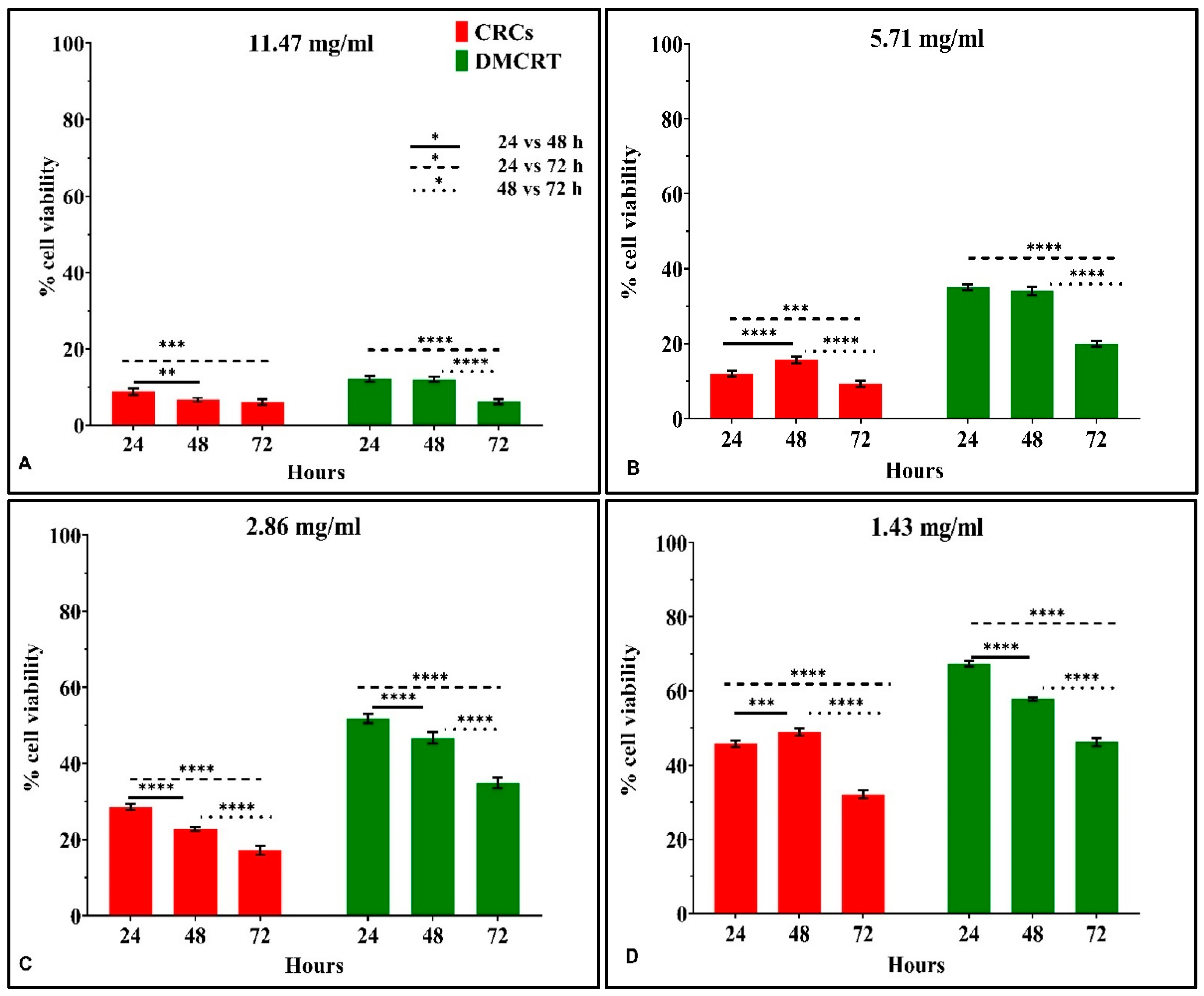
3.3.4. The IC50 Curves of CRCs and DMCRT on Rhabdomyosarcoma Cells (TE671)
3.3.5. Gene Expression under CRCs and DMCRT on Rhabdomyosarcoma Cells (TE671)
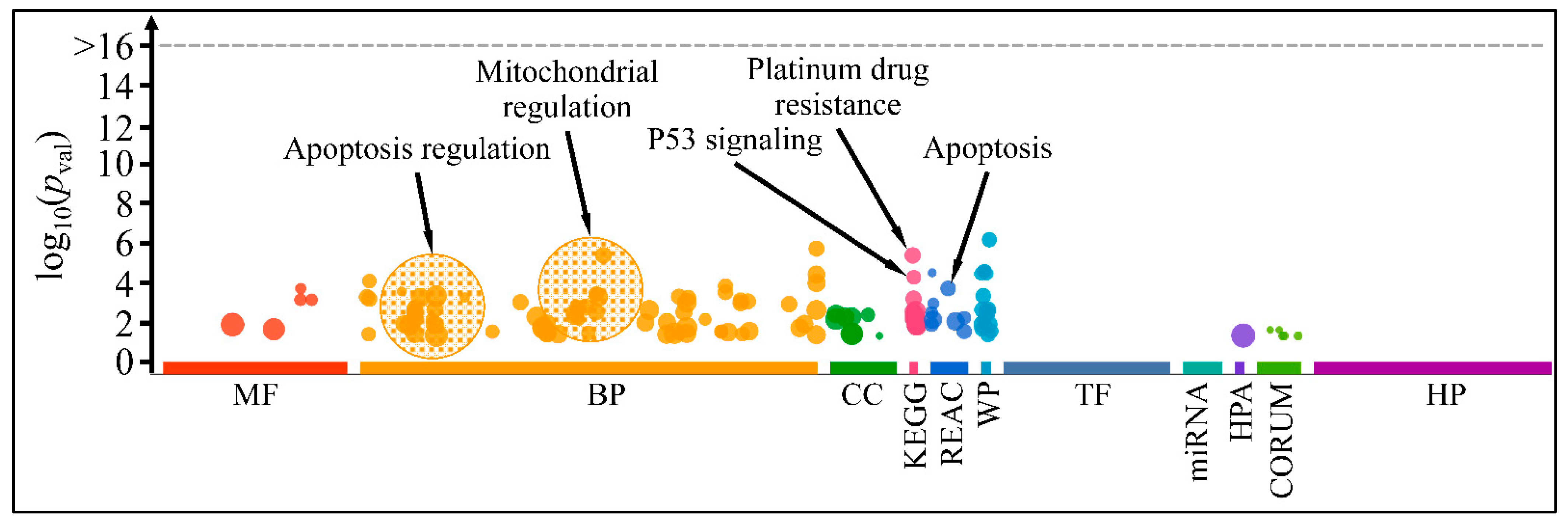
3.4. Functional Annotations of Selected Genes
4. Discussion
4.1. In Vitro Cytotoxicity of Saffron’s Metabolites
4.2. Gene Expression Alterations in Cancer Cell Lines in the Presence of Saffron’s Metabolites
5. Conclusions
Supplementary Materials
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namayandeh, S.M.; Khazaei, Z.; Lari Najafi, M.; Goodarzi, E.; Moslem, A. GLOBAL Leukemia in Children 0–14 Statistics 2018, Incidence and Mortality and Human Development Index (HDI): GLOBOCAN Sources and Methods. Asian Pac. J. Cancer Prev. 2020, 21, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Ferrucci, V.; Boffa, I.; De Masi, G.; Zollo, M. Natural compounds for pediatric cancer treatment. Naunyn-Schmiedeberg's Arch. Pharmacol. 2016, 389, 131–149. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-derived anticancer agents: A green anticancer approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Shoeb, M. Anticancer agents from medicinal plants. Bangladesh J. Pharmacol. 2008, 1, 35–41. [Google Scholar] [CrossRef]
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anti-cancer activity: From folklore to practice. Front. Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef]
- Christodoulou, E.; Kadoglou, N.P.; Kostomitsopoulos, N.; Valsami, G. Saffron: A natural product with potential pharmaceutical applications. J. Pharm. Pharmacol. 2015, 67, 1634–1649. [Google Scholar] [CrossRef]
- Hosseinzadeh, H. Saffron: A herbal medicine of third millennium. Jundishapur J. Nat. Pharm. Prod. 2014, 9, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Boskabady, M.H.; Davoodi, S. Use of in vitro assays to assess the potential antiproliferative and cytotoxic effects of saffron (Crocus sativus L.) in human lung cancer cell line. Pharm. Mag. 2010, 6, 309–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Betti, G.; Hensel, A. Saffron in phytotherapy: Pharmacology and clinical uses. Wien. Med. Wochenschr. 2007, 157, 315. [Google Scholar] [CrossRef] [PubMed]
- Alavizadeh, S.H.; Hosseinzadeh, H. Bioactivity assessment and toxicity of crocin: A comprehensive review. Food Chem. Toxicol. 2014, 64, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulou-Kyriakides, M.; Kyriakidis, D.A. Croscus sativus-biological active constitutents. Stud. Nat. Prod. Chem. 2002, 26, 293–312. [Google Scholar]
- Assimopoulou, A.N.; Sinakos, Z.; Papageorgiou, V.P. Radical scavenging activity of Crocus sativus L. extract and its bioactive constituents. Phytother. Res. 2005, 19, 997–1000. [Google Scholar] [CrossRef]
- Tarantilis, P.A.; Tsoupras, G.; Polissiou, M. Determination of saffron (Crocus sativus L.) components in crude plant extract using high-performance liquid chromatography-UV-visible photodiode-array detection-mass spectrometry. J. Chromatogr. 1995, 699, 107–118. [Google Scholar] [CrossRef]
- Caballero-Ortega, H.; Pereda-Miranda, R.; Abdullaev, F.I. HPLC quantification of major active components from 11 different saffron (Crocus sativus L.) sources. Food Chem. 2007, 100, 1126–1131. [Google Scholar] [CrossRef]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and natural carotenoids: Biochemical activities and anti-tumor effects. Biochim. Biophys. Acta 2014, 1845, 20–30. [Google Scholar] [CrossRef]
- Rahaiee, S.; Hashemi, M.; Shojaosadati, S.A.; Moini, S.; Razavi, S.H. Nanoparticles based on crocin loaded chitosan-alginate biopolymers: Antioxidant activities, bioavailability and anticancer properties. Int. J. Biol. Macromol. 2017, 99, 401–408. [Google Scholar] [CrossRef]
- Rahaiee, S.; Moini, S.; Hashemi, M.; Shojaosadati, S.A. Evaluation of antioxidant activities of bioactive compounds and various extracts obtained from saffron (Crocus sativus L.): A review. J. Food Sci. Technol. 2015, 52, 1881–1888. [Google Scholar] [CrossRef] [Green Version]
- Bathaie, S.Z.; Mousavi, S.Z. New applications and mechanisms of action of saffron and its important ingredients. Crit. Rev. Food Sci. Nutr. 2010, 50, 761–786. [Google Scholar] [CrossRef] [PubMed]
- Winterhalter, P.; Straubinger, M. Saffron-renewed interest in an ancient spice. Food Rev. Int. 2000, 16, 39–59. [Google Scholar] [CrossRef]
- Feizzadeh, B.; Afshari, J.T.; Rakhshandeh, H.; Rahimi, A.; Brook, A.; Doosti, H. Cytotoxic effect of saffron stigma aqueous extract on human transitional cell carcinoma and mouse fibroblast. Urol. J. 2008, 5, 161–167. [Google Scholar]
- Bostan, H.B.; Mehri, S.; Hosseinzadeh, H. Toxicology effects of saffron and its constituents: A review. Iran. J. Basic Med. Sci. 2017, 20, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Butnariu, M.; Quispe, C.; Herrera-Bravo, J.; Sharifi-Rad, J.; Singh, L.; Aborehab, N.M.; Bouyahya, A.; Venditti, A.; Sen, S.; Acharya, K.; et al. The Pharmacological Activities of Crocus sativus L.: A Review Based on the Mechanisms and Therapeutic Opportunities of its Phytoconstituents. Oxid. Med. Cell Longev. 2022, 2022, 8214821. [Google Scholar] [CrossRef]
- Mehri, S.; Razavi, B.-M.; Hosseinzadeh, H. Chapter 34—Safety and toxicity of saffron. In Saffron; Koocheki, A., Khajeh-Hosseini, M., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 517–530. [Google Scholar]
- Premkumar, K.; Abraham, S.K.; Santhiya, S.T.; Ramesh, A. Protective effects of saffron (Crocus sativus Linn.) on genotoxins-induced oxidative stress in Swiss albino mice. Phytother. Res. 2003, 17, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, M.A.; Tsachaki, M.; Efthimiopoulos, S.; Cordopatis, P.; Lamari, F.N.; Margarity, M. Memory enhancing effects of saffron in aged mice are correlated with antioxidant protection. Behav. Brain Res. 2011, 219, 197–204. [Google Scholar] [CrossRef]
- Linardaki, Z.I.; Orkoula, M.G.; Kokkosis, A.G.; Lamari, F.N.; Margarity, M. Investigation of the neuroprotective action of saffron (Crocus sativus L.) in aluminum-exposed adult mice through behavioral and neurobiochemical assessment. Food Chem. Toxicol. 2013, 52, 163–170. [Google Scholar] [CrossRef]
- Zhang, Y.; Shoyama, Y.; Sugiura, M.; Saito, H. Effects of Crocus sativus L. on the Ethanol-Induced Impairment of Passive Avoidance Performances in Mice. Biol. Pharm. Bull. 1994, 17, 217–221. [Google Scholar] [CrossRef]
- Pitsikas, N.; Sakellaridis, N. Crocus sativus L. extracts antagonize memory impairments in different behavioural tasks in the rat. Behav. Brain Res. 2006, 173, 112–115. [Google Scholar] [CrossRef]
- Sugiura, M.; Shoyama, Y.; Saito, H.; Nishiyama, N. Crocin Improves the Ethanol-induced Impairment of Learning Behaviors of Mice in Passive Avoidance Tasks. Proc. Jpn. Acad. Ser. B 1995, 71, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, H.; Shariaty, V.M.; Sameni, A.K.; Vahabzadeh, M. Acute and sub-acute toxicity of crocin, a constituent of Crocus sativus L. (saffron), in mice and rats. Pharmacologyonline 2010, 2, 943–951. [Google Scholar]
- Taheri, F.; Bathaie, S.Z.; Ashrafi, M.; Ghasemi, E. Assessment of crocin toxicity on the rat liver. Modares J. Med. Sci. Pathobiol. 2014, 17, 67–79. [Google Scholar]
- Razavi, B.M.; Hosseinzadeh, H.; Movassaghi, A.R.; Imenshahidi, M.; Abnous, K. Protective effect of crocin on diazinon induced cardiotoxicity in rats in subchronic exposure. Chem. Biol. Interact. 2013, 203, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Q.; Liu, J.X.; Wang, J.N.; Xu, L. Effects of crocin on reperfusion-induced oxidative/nitrative injury to cerebral microvessels after global cerebral ischemia. Brain Res. 2007, 1138, 86–94. [Google Scholar] [CrossRef]
- Salem, M.; Shaheen, M.; Tabbara, A.; Borjac, J. Saffron extract and crocin exert anti-inflammatory and anti-oxidative effects in a repetitive mild traumatic brain injury mouse model. Sci. Rep. 2022, 12, 5004. [Google Scholar] [CrossRef]
- Ghadrdoost, B.; Vafaei, A.A.; Rashidy-Pour, A.; Hajisoltani, R.; Bandegi, A.R.; Motamedi, F.; Haghighi, S.; Sameni, H.R.; Pahlvan, S. Protective effects of saffron extract and its active constituent crocin against oxidative stress and spatial learning and memory deficits induced by chronic stress in rats. Eur. J. Pharmacol. 2011, 667, 222–229. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, L.; Zhang, L.; Liu, W.-B.; Chen, X.-Y.; Yang, X.-G. Crocin prevents retinal ischaemia/reperfusion injury-induced apoptosis in retinal ganglion cells through the PI3K/AKT signalling pathway. Exp. Eye Res. 2013, 107, 44–51. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Sadeghnia, H.R.; Ziaee, T.; Danaee, A. Protective effect of aqueous saffron extract (Crocus sativus L.) and crocin, its active constituent, on renal ischemia-reperfusion-induced oxidative damage in rats. J. Pharm. Pharm. Sci. 2005, 8, 387–393. [Google Scholar]
- Mashmoul, M.; Azlan, A.; Yusof, B.N.M.; Khaza’ai, H.; Mohtarrudin, N.; Boroushaki, M.T. Effects of saffron extract and crocin on anthropometrical, nutritional and lipid profile parameters of rats fed a high fat diet. J. Funct. Foods 2014, 8, 180–187. [Google Scholar] [CrossRef]
- Jahromi, A.S.; Kargar, M.; Kafilzadeh, F.; Jamalidoust, M.; Moradzadeh, M. Crocin Promotes Apoptosis in Human EBV-Transformed B-Lymphocyte via Intrinsic Pathway. Mediterr J. Hematol. Infect Dis. 2021, 13, e2021049. [Google Scholar] [CrossRef] [PubMed]
- Ahmadnia, H.; Tavakkol Afshari, J.; Tabeshpour, J.; Younesi Rostami, M.; Mansourian, E.; Akhavan Rezayat, A.; Brook, A. Cytotoxic Effect of Saffron Stigma Aqueous Extract on Human Prostate Cancer and Mouse Fibroblast Cell Lines. Urol. J. 2020, 18, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Farrukh, A.; Murali, C.; Soleimani, A.; Praz, F.; Graziani, G.; Brim, H.; Ashktorab, H. Saffron and Its Major Ingredients’ Effect on Colon Cancer Cells with Mismatch Repair Deficiency and Microsatellite Instability. Molecules 2021, 26, 3855. [Google Scholar] [CrossRef] [PubMed]
- Bijani, F.; Zabihi, E.; Bijani, A.; Nouri, H.R.; Nafarzadeh, S.; Seyedmajidi, M. Evaluation of apoptotic effect of crocin, cisplatin, and their combination in human oral squamous cell carcinoma cell line HN5. Dent. Res. J. 2021, 18, 70. [Google Scholar]
- Colapietro, A.; Mancini, A.; Vitale, F.; Martellucci, S.; Angelucci, A.; Llorens, S.; Mattei, V.; Gravina, G.L.; Alonso, G.L.; Festuccia, C. Crocetin Extracted from Saffron Shows Antitumor Effects in Models of Human Glioblastoma. Int. J. Mol. Sci. 2020, 21, 423. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Park, B. Saffron carotenoids inhibit STAT3 activation and promote apoptotic progression in IL-6-stimulated liver cancer cells. Oncol. Rep. 2018, 39, 1883–1891. [Google Scholar] [CrossRef] [Green Version]
- Shariat Razavi, S.M.; Mahmoudzadeh Vaziri, R.; Karimi, G.; Arabzadeh, S.; Keyvani, V.; Behravan, J.; Kalalinia, F. Crocin Increases Gastric Cancer Cells’ Sensitivity to Doxorubicin. Asian Pac. J. Cancer Prev. 2020, 21, 1959–1967. [Google Scholar] [CrossRef]
- Ahmadabadi, F.; Saghebjoo, M.; Hedayati, M.; Hoshyar, R.; Huang, C.J. Treatment-induced tumor cell apoptosis following high-intensity interval training and saffron aqueous extract in mice with breast cancer. Physiol. Int. 2021, 108, 19–26. [Google Scholar] [CrossRef]
- Kakouri, E.; Agalou, A.; Kanakis, C.; Beis, D.; Tarantilis, P.A. Crocins from Crocus sativus L. in the Management of Hyperglycemia. In Vivo Evidence from Zebrafish. Molecules 2020, 25, 5223. [Google Scholar] [CrossRef]
- Carmona, M.; Zalacain, A.; Sanchez, A.M.; Novella, J.L.; Alonso, G.L. Crocetin esters, picrocrocin and its related compounds present in Crocus sativus stigmas and Gardenia jasminoides fruits. Tentative identification of seven new compounds by LC-ESI-MS. J. Agric. Food Chem. 2006, 54, 973–979. [Google Scholar] [CrossRef]
- Mohajeri, S.A.; Hosseinzadeh, H.; Keyhanfar, F.; Aghamohammadian, J. Extraction of crocin from saffron (Crocus sativus) using molecularly imprinted polymer solid-phase extraction. J. Sep. Sci. 2010, 33, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- Karkoula, E.; Angelis, A.; Koulakiotis, N.S.; Gikas, E.; Halabalaki, M.; Tsarbopoulos, A.; Skaltsounis, A.L. Rapid isolation and characterization of crocins, picrocrocin, and crocetin from saffron using centrifugal partition chromatography and LC-MS. J. Sep. Sci. 2018, 41, 4105–4114. [Google Scholar] [CrossRef] [PubMed]
- Assimiadis, M.K.; Tarantilis, P.A.; Polissiou, M.G. UV-Vis, FT-Raman, and 1H NMR Spectroscopies of cis-trans Carotenoids from Saffron (Crocus sativus L.). Appl. Spectrosc. 1998, 52, 519–522. [Google Scholar] [CrossRef]
- Tarantilis, P.A.; Beljebbar, A.; Manfait, M.; Polissiou, M. FT-IR, FT-Raman spectroscopic study of carotenoids from saffron (Crocus sativus L.) and some derivatives. Spectrochim Acta A 1998, 54, 651–657. [Google Scholar] [CrossRef]
- Hamid, R.; Rotshteyn, Y.; Rabadi, L.; Parikh, R.; Bullock, P. Comparison of alamar blue and MTT assays for high through-put screening. Toxicol. In Vitro 2004, 18, 703–710. [Google Scholar] [CrossRef]
- Petrenko, Y.A.; Gorokhova, N.A.; Tkachova, E.N.; Petrenko, A.Y. The reduction of Alamar Blue by peripheral blood lymphocytes and isolated mitochondria. Ukr Biokhim Zh (1999) 2005, 77, 100–105. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Schmoyer, D.; Kirov, S.; Snoddy, J. GOTree Machine (GOTM): A web-based platform for interpreting sets of interesting genes using Gene Ontology hierarchies. BMC Bioinform. 2004, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Vali, F.; Changizi, V.; Safa, M. Synergistic Apoptotic Effect of Crocin and Paclitaxel or Crocin and Radiation on MCF-7 Cells, a Type of Breast Cancer Cell Line. Int. J. Breast Cancer 2015, 2015, 139349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollaei, H.; Safaralizadeh, R.; Babaei, E.; Abedini, M.R.; Hoshyar, R. The anti-proliferative and apoptotic effects of crocin on chemosensitive and chemoresistant cervical cancer cells. Biomed. Pharmacother. 2017, 94, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Liu, B.B.; Qian, X.D.; Li, L.Q.; Cao, H.B.; Guo, Q.S.; Zhou, G.F. Crocin induces autophagic apoptosis in hepatocellular carcinoma by inhibiting Akt/mTOR activity. OncoTargets Ther. 2018, 11, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, T.; Jiang, G.; Gong, W.; Qian, H.; Zou, C. Synergistic apoptotic effect of crocin and cisplatin on osteosarcoma cells via caspase induced apoptosis. Toxicol. Lett. 2013, 221, 197–204. [Google Scholar] [CrossRef]
- Faridi, N.; Heidarzadeh, H.R.; Mohagheghi, M.A.; Bathaie, S.Z.J.B.; Research, C.C. BT-474 Breast Cancer Cell Apoptosis Induced by Crocin, a Saffron Carotenoid. Basic Clin. Cancer Res. 2019, 11, 5–15. [Google Scholar] [CrossRef]
- Amin, A.; Bajbouj, K.; Koch, A.; Gandesiri, M.; Schneider-Stock, R. Defective autophagosome formation in p53-null colorectal cancer reinforces crocin-induced apoptosis. Int. J. Mol. Sci. 2015, 16, 1544–1561. [Google Scholar] [CrossRef] [Green Version]
- Jabini, R.; Ehtesham-Gharaee, M.; Dalirsani, Z.; Mosaffa, F.; Delavarian, Z.; Behravan, J. Evaluation of the Cytotoxic Activity of Crocin and Safranal, Constituents of Saffron, in Oral Squamous Cell Carcinoma (KB Cell Line). Nutr. Cancer 2017, 69, 911–919. [Google Scholar] [CrossRef]
- Luo, Y.; Cui, S.; Tang, F.; Shen, C.; Qi, Y.; Lu, D.; Ma, L.; Yang, Y.; Li, Y.; Chen, R.; et al. The combination of crocin with cisplatin suppresses growth of gastric carcinoma cell line BGC-823 and promotes cell apoptosis. Pak. J. Pharm. Sci. 2017, 30, 1629–1634. [Google Scholar]
- Luo, Y.; Yu, P.; Zhao, J.; Guo, Q.; Fan, B.; Diao, Y.; Jin, Y.; Zhang, C. Pathogenesis and anti-proliferation mechanisms of Crocin in human gastric carcinoma cells. Int. J. Clin. Exp. Pathol. 2020, 13, 912–922. [Google Scholar]
- Noureini, S.K.; Wink, M. Antiproliferative effects of crocin in HepG2 cells by telomerase inhibition and hTERT down-regulation. Asian Pac. J. Cancer Prev. 2012, 13, 2305–2309. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Xu, H.J.; Zhao, Y.X.; Wang, L.Z.; Sun, L.R.; Wang, Z.; Sun, X.F. Crocin Exhibits Antitumor Effects on Human Leukemia HL-60 Cells In Vitro and In Vivo. Evid-Based Complement. Altern. Med. 2013, 2013, 690164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wang, Z.; Wang, L.; Wang, L.Z.; Zang, C.; Sun, L.R. The Effect and Mechanisms of Proliferative Inhibition of Crocin on Human Leukaemia Jurkat Cells. West Indian Med. J. 2015, 64, 473–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhao, S.; Wang, X.; Zhang, L.; Jiang, E.; Gu, Y.; Shangguan, A.J.; Zhao, H.; Lv, T.; Yu, Z. Crocin inhibits cell proliferation and enhances cisplatin and pemetrexed chemosensitivity in lung cancer cells. Transl. Lung Cancer Res. 2015, 4, 775–783. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.M.; Mancini, A.; Lizzi, A.R.; De Simone, A.; Marroccella, C.E.; Gravina, G.L.; Tatone, C.; Festuccia, C. Crocus sativus stigma extract and its major constituent crocin possess significant antiproliferative properties against human prostate cancer. Nutr. Cancer 2013, 65, 930–942. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, S.; Wang, K.; Huang, Y.; Yang, N.; Yang, Z.; Zheng, Z.; Wang, Y. Crocin induces autophagic cell death and inhibits cell invasion of cervical cancer SiHa cells through activation of PI3K/AKT. Ann. Transl. Med. 2020, 8, 1180. [Google Scholar] [CrossRef]
- Cragg, G.M.; Boyd, M.R.; Cardellina, J.H., 2nd; Newman, D.J.; Snader, K.M.; McCloud, T.G. Ethnobotany and drug discovery: The experience of the US National Cancer Institute. Ciba Found. Symp. 1994, 185, 178–190, discussion 190–176. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Frenkel, G.D. The effect of saffron on intracellular DNA, RNA and protein synthesis in malignant and non-malignant human cells. BioFactors 1992, 4, 43–45. [Google Scholar]
- Chryssanthi, D.G.; Dedes, P.G.; Karamanos, N.K.; Cordopatis, P.; Lamari, F.N. Crocetin inhibits invasiveness of MDA-MB-231 breast cancer cells via downregulation of matrix metalloproteinases. Planta Med. 2011, 77, 146–151. [Google Scholar] [CrossRef]
- Chryssanthi, D.G.; Lamari, F.N.; Iatrou, G.; Pylara, A.; Karamanos, N.K.; Cordopatis, P. Inhibition of breast cancer cell proliferation by style constituents of different Crocus species. Anticancer Res. 2007, 27, 357–362. [Google Scholar]
- Lu, P.; Lin, H.; Gu, Y.; Li, L.; Guo, H.; Wang, F.; Qiu, X. Antitumor effects of crocin on human breast cancer cells. Int. J. Clin. Exp. Med. 2015, 8, 20316–20322. [Google Scholar]
- Xia, D. Ovarian cancer HO-8910 cell apoptosis induced by crocin in vitro. Nat. Prod. Commun. 2015, 10, 249–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavakkol-Afshari, J.; Brook, A.; Mousavi, S.H. Study of cytotoxic and apoptogenic properties of saffron extract in human cancer cell lines. Food Chem. Toxicol. 2008, 46, 3443–3447. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Riverón-Negrete, L.; Caballero-Ortega, H.; Manuel Hernández, J.; Pérez-López, I.; Pereda-Miranda, R.; Espinosa-Aguirre, J.J. Use of in vitro assays to assess the potential antigenotoxic and cytotoxic effects of saffron (Crocus sativus L.). Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2003, 17, 731–736. [Google Scholar] [CrossRef]
- Abdullaev, F.I. Inhibitory effect of crocetin on intracellular nucleic acid and protein synthesis in malignant cells. Toxicol. Lett. 1994, 70, 243–251. [Google Scholar] [CrossRef]
- Dhar, A.; Mehta, S.; Dhar, G.; Dhar, K.; Banerjee, S.; Van Veldhuizen, P.; Campbell, D.R.; Banerjee, S.K. Crocetin inhibits pancreatic cancer cell proliferation and tumor progression in a xenograft mouse model. Mol. Cancer Ther. 2009, 8, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutheil, W.G.; Reed, G.; Ray, A.; Anant, S.; Dhar, A. Crocetin: An agent derived from saffron for prevention and therapy for cancer. Curr. Pharm. Biotechnol. 2012, 13, 173–179. [Google Scholar] [CrossRef]
- Ray, P.; Guha, D.; Chakraborty, J.; Banerjee, S.; Adhikary, A.; Chakraborty, S.; Das, T.; Sa, G. Crocetin exploits p53-induced death domain (PIDD) and FAS-associated death domain (FADD) proteins to induce apoptosis in colorectal cancer. Sci. Rep. 2016, 6, 32979. [Google Scholar] [CrossRef]
- An, B.; Ma, Y.; Xu, Y.; Liu, X.; Zhang, X.; Zhang, J.; Yang, C. Crocin regulates the proliferation and migration of neural stem cells after cerebral ischemia by activating the Notch1 pathway. Folia Neuropathol. 2020, 58, 201–212. [Google Scholar] [CrossRef]
- Xu, H.J.; Zhang, K.P.; Zhong, R.; Zhao, Y.X.; Li, X.R.; Lu, Y.; Song, A.Q.; Pang, X.Y.; Sun, L.R. Influence of crocin on proliferation in vitro and function of dendritic cells derived from bone marrow of children with acute leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2012, 20, 57–61. [Google Scholar]
- Nam, K.N.; Park, Y.-M.; Jung, H.-J.; Lee, J.Y.; Min, B.D.; Park, S.-U.; Jung, W.-S.; Cho, K.-H.; Park, J.-H.; Kang, I.; et al. Anti-inflammatory effects of crocin and crocetin in rat brain microglial cells. Eur. J. Pharmacol. 2010, 648, 110–116. [Google Scholar] [CrossRef]
- Du, J.; Li, Y.; Song, D.; Liu, J.; Huang, Q.; Li, J.; Li, B.; Li, L. Protective effects of crocin against endogenous Aβ-induced neurotoxicity in N2a/APP695swe cells. Psychopharmacology 2021, 238, 2839–2847. [Google Scholar] [CrossRef] [PubMed]
- Laabich, A.; Vissvesvaran, G.P.; Lieu, K.L.; Murata, K.; McGinn, T.E.; Manmoto, C.C.; Sinclair, J.R.; Karliga, I.; Leung, D.W.; Fawzi, A.; et al. Protective Effect of Crocin against Blue Light– and White Light–Mediated Photoreceptor Cell Death in Bovine and Primate Retinal Primary Cell Culture. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3156–3163. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Lin, S.; Tong, Z.; Chen, S.; Cao, Y.; Li, Q.; Jiang, Y.; Cai, W.; Tong, Y.; Zahra, B.S.; et al. Crocetin ameliorates non-alcoholic fatty liver disease by modulating mitochondrial dysfunction in L02 cells and zebrafish model. J. Ethnopharmacol. 2022, 285, 114873. [Google Scholar] [CrossRef] [PubMed]
- Fagot, D.; Pham, D.M.; Laboureau, J.; Planel, E.; Guerin, L.; Nègre, C.; Donovan, M.; Bernard, B.A. Crocin, a natural molecule with potentially beneficial effects against skin ageing. Int. J. Cosmet. Sci. 2018, 40, 388–400. [Google Scholar] [CrossRef]
- Hoshyar, R.; Bathaie, S.Z.; Sadeghizadeh, M. Crocin triggers the apoptosis through increasing the Bax/Bcl-2 ratio and caspase activation in human gastric adenocarcinoma, AGS, cells. DNA Cell Biol. 2013, 32, 50–57. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, M.; Krishna Mohan, S.; Hao, Z. Crocin treatment promotes the oxidative stress and apoptosis in human thyroid cancer cells FTC-133 through the inhibition of STAT/JAK signaling pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22608. [Google Scholar] [CrossRef]
- Xu, Q.; Yu, J.; Jia, G.; Li, Z.; Xiong, H. Crocin attenuates NF-κB-mediated inflammation and proliferation in breast cancer cells by down-regulating PRKCQ. Cytokine 2022, 154, 155888. [Google Scholar] [CrossRef]
- Xu, G.-L.; Qian, Z.-Y.; Yu, S.-Q.; Gong, Z.-N.; Shen, X.-C. Evidence of crocin against endothelial injury induced by hydrogen peroxide in vitro. J. Asian Nat. Prod. Res. 2006, 8, 79–85. [Google Scholar] [CrossRef]
- Xu, G.; Gong, Z.; Yu, W.; Gao, L.; He, S.; Qian, Z. Increased Expression Ratio of Bcl-2/Bax Is Associated with Crocin-Mediated Apoptosis in Bovine Aortic Endothelial Cells. Basic Clin. Pharmacol. Toxicol. 2007, 100, 31–35. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, S.; Chen, Q.; Wu, Y.; Zhang, J.; Li, N. Crocin inhibits RANKL-induced osteoclastogenesis by regulating JNK and NF-κB signaling pathways. Mol. Med. Rep. 2018, 17, 7947–7951. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Wang, L.; Si, S.; Sun, Y.; Pei, W.; Ming, Y.; Sun, L. Crocin improves the proliferation and cytotoxic function of T cells in children with acute lymphoblastic leukemia. Biomed. Pharmacother. 2018, 99, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Gezici, S. Comparative anticancer activity analysis of saffron extracts and a principle component, crocetin for prevention and treatment of human malignancies. J. Food Sci. Technol. 2019, 56, 5435–5443. [Google Scholar] [CrossRef] [PubMed]
- Aung, H.H.; Wang, C.Z.; Ni, M.; Fishbein, A.; Mehendale, S.R.; Xie, J.T.; Shoyama, C.Y.; Yuan, C.S. Crocin from Crocus sativus possesses significant anti-proliferation effects on human colorectal cancer cells. Exp. Oncol. 2007, 29, 175–180. [Google Scholar] [PubMed]
- Trujillo-Jiménez, F.; García-López, P.; Garcí-Carrancá, A.; Abdullaev, F.I. Effect of saffron on the viability of normal and malignant human cells in vitro. Acta Hortic. 2004, 650, 463–469. [Google Scholar] [CrossRef]
- See, S.J.; Levin, V.A.; Yung, W.K.; Hess, K.R.; Groves, M.D. 13-cis-retinoic acid in the treatment of recurrent glioblastoma multiforme. Neuro-Oncology 2004, 6, 253–258. [Google Scholar] [CrossRef]
- Mawson, A.R. Retinoids in the treatment of glioma: A new perspective. Cancer Manag. Res. 2012, 4, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.P.; Matthay, K.K.; Villablanca, J.G.; Maurer, B.J. Retinoid therapy of high-risk neuroblastoma. Cancer Lett. 2003, 197, 185–192. [Google Scholar] [CrossRef]
- Reynolds, C.P.; Lemons, R.S. Retinoid therapy of childhood cancer. Hematol. Oncol. Clin. N. Am. 2001, 15, 867–910. [Google Scholar] [CrossRef]
- Masetti, R.; Biagi, C.; Zama, D.; Vendemini, F.; Martoni, A.; Morello, W.; Gasperini, P.; Pession, A. Retinoids in pediatric onco-hematology: The model of acute promyelocytic leukemia and neuroblastoma. Adv. Ther. 2012, 29, 747–762. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for Prevention and Treatment of Cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef]
- Wang, H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Lee, J.H.; Kong, A.N. Plants vs. cancer: A review on natural phytochemicals in preventing and treating cancers and their druggability. Anti-Cancer Agents Med. Chem. 2012, 12, 1281–1305. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, B.; Wang, Y.; Han, S.; Wang, C. Crocin promotes apoptosis of human skin cancer cells by inhibiting the JAK/STAT pathway. Exp. Ther. Med. 2018, 16, 5079–5084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Luo, C.L.; Wu, X.H.; Hu, H.B.; Lv, C.F.; Ji, H.Y. Proliferation apoptotic influence of crocin on human bladder cancer T24 cell line. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 2008, 33, 1869–1873. [Google Scholar]
- Veisi, A.; Akbari, G.; Mard, S.A.; Badfar, G.; Zarezade, V.; Mirshekar, M.A. Role of crocin in several cancer cell lines: An updated review. Iran. J. Basic Med. Sci. 2020, 23, 3–12. [Google Scholar] [CrossRef]
- Medema, R.H.; Macurek, L. Checkpoint recovery in cells: How a molecular understanding can help in the fight against cancer. F1000 Biol. Rep. 2011, 3, 10. [Google Scholar] [CrossRef]
- Peng, A. Working hard for recovery: Mitotic kinases in the DNA damage checkpoint. Cell Biosci. 2013, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- van Vugt, M.A.; Medema, R.H. Checkpoint adaptation and recovery: Back with Polo after the break. Cell Cycle 2004, 3, 1383–1386. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, C.Z.; Wen, X.D.; Shoyama, Y.; Yuan, C.S. Role of saffron and its constituents on cancer chemoprevention. Pharm. Biol. 2013, 51, 920–924. [Google Scholar] [CrossRef] [Green Version]
- Heidarzadeh, H.R.; Bathaie, S.Z.; Abroun, S.; Mohagheghi, M.A. Evaluating the cytotoxic effect of crocin on MDA-MB-468 cell line based on apoptosis induction, ER stress, and autophagy markers. MJMS 2018, 20, 37–51. [Google Scholar]
- Hashemi, S.A.; Bathaie, S.Z.; Mohagheghi, M.A.J.A.J.o.P. Crocetin and crocin decreased cholesterol and triglyceride content of both breast cancer tumors and cell lines. Avicenna J. Phytomed. 2020, 10, 384–397. [Google Scholar]
- Mostafavinia, S.E.; Khorashadizadeh, M.; Hoshyar, R. Antiproliferative and Proapoptotic Effects of Crocin Combined with Hyperthermia on Human Breast Cancer Cells. DNA Cell Biol. 2016, 35, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Escribano, J.; Alonso, G.L.; Coca-Prados, M.; Fernandez, J.A. Crocin, safranal and picrocrocin from saffron (Crocus sativus L.) inhibit the growth of human cancer cells in vitro. Cancer Lett. 1996, 100, 23–30. [Google Scholar] [CrossRef]
- Jiang, Z.; Gu, M.; Liu, J.; Li, H.; Peng, J.; Zhang, Y. Anticancer activity of crocin against cervical carcinoma (HeLa cells): Bioassessment and toxicity evaluation of crocin in male albino rats. J. Photochem. Photobiology. B Biol. 2018, 180, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, J.M.; Kim, S.C.; Park, C.B.; Lee, P.C. Proposed cytotoxic mechanisms of the saffron carotenoids crocin and crocetin on cancer cell lines. Biochem. Cell Biol. 2014, 92, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Mollaei, H.; Abedini, M.R.; Hoshyar, R. Suppressive Effect of Crocin and Cisplatin on Pluripotency Genes Expression in Human Cervical Cancer Cells. Int. J. Cancer Manag. 2017, 10, e11152. [Google Scholar] [CrossRef] [Green Version]
- García-Olmo, D.C.; Riese, H.H.; Escribano, J.; Ontañón, J.; Fernandez, J.A.; Atiénzar, M.; García-Olmo, D. Effects of long-term treatment of colon adenocarcinoma with crocin, a carotenoid from saffron (Crocus sativus L.): An experimental study in the rat. Nutr. Cancer 1999, 35, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, H.A.; Quinn, G.A.; Nasef, M.M.; Mishra, V.; Aljabali, A.A.A.; El-Tanani, M.; Serrano-Aroca, Á.; Webba Da Silva, M.; McCarron, P.A.; Tambuwala, M.M. Crocin Inhibits Angiogenesis and Metastasis in Colon Cancer via TNF-α/NF-kB/VEGF Pathways. Cells 2022, 11, 1502. [Google Scholar] [CrossRef]
- Bi, X.; Jiang, Z.; Luan, Z.; Qiu, D. Crocin exerts anti-proliferative and apoptotic effects on cutaneous squamous cell carcinoma via miR-320a/ATG2B. Bioengineered 2021, 12, 4569–4580. [Google Scholar] [CrossRef]
- Li, S.; Shen, X.-Y.; Ouyang, T.; Qu, Y.; Luo, T.; Wang, H.-Q. Synergistic anticancer effect of combined crocetin and cisplatin on KYSE-150 cells via p53/p21 pathway. Cancer Cell Int. 2017, 17, 98. [Google Scholar] [CrossRef]
- Li, S.; Jiang, S.; Jiang, W.; Zhou, Y.; Shen, X.Y.; Luo, T.; Kong, L.P.; Wang, H.Q. Anticancer effects of crocetin in human esophageal squamous cell carcinoma KYSE-150 cells. Oncol. Lett. 2015, 9, 1254–1260. [Google Scholar] [CrossRef] [Green Version]
- Vazifedan, V.; Mousavi, S.H.; Sargolzaei, J.; Soleymanifard, S.; Fani Pakdel, A. Study of Crocin & Radiotherapy-induced Cytotoxicity and Apoptosis in the Head and Neck Cancer (HN-5) Cell Line. Iran. J. Pharm. Res. 2017, 16, 230–237. [Google Scholar] [PubMed]
- Luo, Y.; Yu, P.; Zhao, J.; Guo, Q.; Fan, B.; Diao, Y.; Jin, Y.; Wu, J.; Zhang, C. Inhibitory Effect of Crocin Against Gastric Carcinoma via Regulating TPM4 Gene. OncoTargets Ther. 2021, 14, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Tarantilis, P.A.; Morjani, H.; Polissiou, M.; Manfait, M. Inhibition of growth and induction of differentiation of promyelocytic leukemia (HL-60) by carotenoids from Crocus sativus L. Anticancer. Res. 1994, 14, 1913–1918. [Google Scholar] [PubMed]
- Hoshyar, R.; Mollaei, H. A comprehensive review on anticancer mechanisms of the main carotenoid of saffron, crocin. J. Pharm. Pharmacol. 2017, 69, 1419–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradzadeh, M.; Kalani, M.R.; Avan, A. The antileukemic effects of saffron (Crocus sativus L.) and its related molecular targets: A mini review. J. Cell. Biochem. 2019, 120, 4732–4738. [Google Scholar] [CrossRef]
- Geromichalos, G.D.; Papadopoulos, T.; Sahpazidou, D.; Sinakos, Z. Safranal, a Crocus sativus L constituent suppresses the growth of K-562 cells of chronic myelogenous leukemia. In silico and in vitro study. Food Chem. Toxicol. 2014, 74, 45–50. [Google Scholar] [CrossRef]
- Samarghandian, S.; Borji, A.; Farahmand, S.K.; Afshari, R.; Davoodi, S. Crocus sativus L. (saffron) stigma aqueous extract induces apoptosis in alveolar human lung cancer cells through caspase-dependent pathways activation. BioMed Res. Int. 2013, 2013, 417928. [Google Scholar] [CrossRef] [Green Version]
- Mahdizadeh, S.; Karimi, G.; Behravan, J.; Arabzadeh, S.; Lage, H.; Kalalinia, F. Crocin suppresses multidrug resistance in MRP overexpressing ovarian cancer cell line. Daru 2016, 24, 17. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, H.; Sam, S.; Rozati, R.; Sultan, P.; Islam, T.; Rathore, B.; Lone, Z.; Sharma, M.; Triphati, J.; Saxena, R.C. DNA fragmentation and cell cycle arrest: A hallmark of apoptosis induced by crocin from kashmiri saffron in a human pancreatic cancer cell line. Asian Pac. J. Cancer Prev. 2010, 11, 675–679. [Google Scholar]
- Deng, L.; Li, J.; Lu, S.; Su, Y. Crocin inhibits proliferation and induces apoptosis through suppressing MYCN expression in retinoblastoma. J. Biochem. Mol. Toxicol. 2019, 33, e22292. [Google Scholar] [CrossRef]
- Sun, J.; Xu, X.M.; Ni, C.Z.; Zhang, H.; Li, X.Y.; Zhang, C.L.; Liu, Y.R.; Li, S.F.; Zhou, Q.Z.; Zhou, H.M. Crocin Inhibits Proliferation and Nucleic Acid Synthesis and Induces Apoptosis in the Human Tongue Squamous Cell Carcinoma Cell Line Tca8113. Asian Pac. J. Cancer Prev. 2011, 12, 2679–2683. [Google Scholar] [PubMed]
- Jafarnia, N.; Ghorbani, Z.; Nokhostin, M.; Manayi, A.; Nourimajd, S.; Razeghi Jahromi, S. Effect of Saffron (Crocus sativus L.) as an Add-On Therapy to Sertraline in Mild to Moderate Generalized Anxiety Disorder: A Double Blind Randomized Controlled Trial. Arch. Neurosci. 2017, 4, e14332. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.A.; Ali Redha, A.; Snoeck, E.R.; Singh, S.; Simal-Gandara, J.; Ibrahim, S.A.; Jafari, S.M. Anti-Depressant Properties of Crocin Molecules in Saffron. Molecules 2022, 27, 2076. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.L.; Li, M.X.; Li, X.L.; Wang, P.; Wang, W.G.; Du, W.Z.; Yang, Z.Q.; Chen, S.F.; Wu, D.; Tian, X.Y. Crocetin: A Systematic Review. Front. Pharmacol. 2021, 12, 745683. [Google Scholar] [CrossRef]
- Ayatollahi, H.; Javan, A.O.; Khajedaluee, M.; Shahroodian, M.; Hosseinzadeh, H. Effect of Crocus sativus L. (Saffron) on Coagulation and Anticoagulation Systems in Healthy Volunteers. Phytother. Res. 2014, 28, 539–543. [Google Scholar] [CrossRef]
- Modaghegh, M.H.; Shahabian, M.; Esmaeili, H.A.; Rajbai, O.; Hosseinzadeh, H. Safety evaluation of saffron (Crocus sativus) tablets in healthy volunteers. Phytomedicine 2008, 15, 1032–1037. [Google Scholar] [CrossRef]
- Mohamadpour, A.H.; Ayati, Z.; Parizadeh, M.R.; Rajbai, O.; Hosseinzadeh, H. Safety evaluation of crocin (a constituent of saffron) tablets in healthy volunteers. Iran. J. Basic Med. Sci. 2013, 16, 39–46. [Google Scholar]
- Mazidi, M.; Shemshian, M.; Mousavi, S.H.; Norouzy, A.; Kermani, T.; Moghiman, T.; Sadeghi, A.; Mokhber, N.; Ghayour-Mobarhan, M.; Ferns, G.A. A double-blind, randomized and placebo-controlled trial of Saffron (Crocus sativus L.) in the treatment of anxiety and depression. J. Complement. Integr. Med. 2016, 13, 195–199. [Google Scholar] [CrossRef]
- Akhondzadeh Basti, A.; Moshiri, E.; Noorbala, A.A.; Jamshidi, A.H.; Abbasi, S.H.; Akhondzadeh, S. Comparison of petal of Crocus sativus L. and fluoxetine in the treatment of depressed outpatients: A pilot double-blind randomized trial. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2007, 31, 439–442. [Google Scholar] [CrossRef]
- Akhondzadeh, S.; Tahmacebi-Pour, N.; Noorbala, A.-A.; Amini, H.; Fallah-Pour, H.; Jamshidi, A.-H.; Khani, M. Crocus sativus L. in the treatment of mild to moderate depression: A double-blind, randomized and placebo-controlled trial. Phytother. Res. 2005, 19, 148–151. [Google Scholar] [CrossRef]
- Ghajar, A.; Neishabouri, S.M.; Velayati, N.; Jahangard, L.; Matinnia, N.; Haghighi, M.; Ghaleiha, A.; Afarideh, M.; Salimi, S.; Meysamie, A.; et al. Crocus sativus L. versus Citalopram in the Treatment of Major Depressive Disorder with Anxious Distress: A Double-Blind, Controlled Clinical Trial. Pharmacopsychiatry 2017, 50, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moshiri, E.; Basti, A.A.; Noorbala, A.-A.; Jamshidi, A.-H.; Hesameddin Abbasi, S.; Akhondzadeh, S. Crocus sativus L. (petal) in the treatment of mild-to-moderate depression: A double-blind, randomized and placebo-controlled trial. Phytomedicine 2006, 13, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Noorbala, A.A.; Akhondzadeh, S.; Tahmacebi-Pour, N.; Jamshidi, A.H. Hydro-alcoholic extract of Crocus sativus L. versus fluoxetine in the treatment of mild to moderate depression: A double-blind, randomized pilot trial. J. Ethnopharmacol. 2005, 97, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Shahmansouri, N.; Farokhnia, M.; Abbasi, S.-H.; Kassaian, S.E.; Noorbala Tafti, A.-A.; Gougol, A.; Yekehtaz, H.; Forghani, S.; Mahmoodian, M.; Saroukhani, S.; et al. A randomized, double-blind, clinical trial comparing the efficacy and safety of Crocus sativus L. with fluoxetine for improving mild to moderate depression in post percutaneous coronary intervention patients. J. Affect. Disord. 2014, 155, 216–222. [Google Scholar] [CrossRef]
- Mousavi, B.; Bathaie, S.Z.; Fadai, F.; Ashtari, Z.; Ali beigi, N.; Farhang, S.; Hashempour, S.; Shahhamzei, N.; Heidarzadeh, H. Safety evaluation of saffron stigma (Crocus sativus L.) aqueous extract and crocin in patients with schizophrenia. Avicenna J. Phytomed. 2015, 5, 413–419. [Google Scholar]
- Ghobadi, H.; Abdollahi, N.; Madani, H.; Aslani, M.R. Effect of Crocin From Saffron (Crocus sativus L.) Supplementation on Oxidant/Antioxidant Markers, Exercise Capacity, and Pulmonary Function Tests in COPD Patients: A Randomized, Double-Blind, Placebo-Controlled Trial. Front. Pharmacol. 2022, 13, 884710. [Google Scholar] [CrossRef]
- Rahimi, G.; Shams, S.; Aslani, M.R. Effects of crocin supplementation on inflammatory markers, lipid profiles, insulin and cardioprotective indices in women with PCOS: A randomized, double-blind, placebo-controlled trial. Phytother. Res. 2022. [Google Scholar] [CrossRef]
- Farokhnia, M.; Shafiee Sabet, M.; Iranpour, N.; Gougol, A.; Yekehtaz, H.; Alimardani, R.; Farsad, F.; Kamalipour, M.; Akhondzadeh, S. Comparing the efficacy and safety of Crocus sativus L. with memantine in patients with moderate to severe Alzheimer’s disease: A double-blind randomized clinical trial. Hum. Psychopharmacol. Clin. Exp. 2014, 29, 351–359. [Google Scholar] [CrossRef]
- Pitsikas, N. The Effect of Crocus sativus L. and Its Constituents on Memory: Basic Studies and Clinical Applications. Evid. Based Com. Altern. Med. 2015, 2015, 926284. [Google Scholar] [CrossRef] [Green Version]
- Akhondzadeh, S.; Sabet, M.S.; Harirchian, M.H.; Togha, M.; Cheraghmakani, H.; Razeghi, S.; Hejazi, S.S.; Yousefi, M.H.; Alimardani, R.; Jamshidi, A.; et al. Saffron in the treatment of patients with mild to moderate Alzheimer’s disease: A 16-week, randomized and placebo-controlled trial. J. Clin. Pharm. Ther. 2010, 35, 581–588. [Google Scholar] [CrossRef]
- Akhondzadeh, S.; Shafiee Sabet, M.; Harirchian, M.H.; Togha, M.; Cheraghmakani, H.; Razeghi, S.; Hejazi, S.S.; Yousefi, M.H.; Alimardani, R.; Jamshidi, A.; et al. A 22-week, multicenter, randomized, double-blind controlled trial of Crocus sativus in the treatment of mild-to-moderate Alzheimer’s disease. Psychopharmacology 2010, 207, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L.; Drummond, P.D.; Inarejos-García, A.M.; Prodanov, M. affron(®), a standardised extract from saffron (Crocus sativus L.) for the treatment of youth anxiety and depressive symptoms: A randomised, double-blind, placebo-controlled study. J. Affect Disord 2018, 232, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikbakht-Jam, I.; Khademi, M.; Nosrati, M.; Eslami, S.; Foroutan-Tanha, M.; Sahebkar, A.; Tavalaie, S.; Ghayour-Mobarhan, M.; Ferns, G.A.A.; Hadizadeh, F.; et al. Effect of crocin extracted from saffron on pro-oxidant–anti-oxidant balance in subjects with metabolic syndrome: A randomized, placebo-controlled clinical trial. Eur. J. Integr. Med. 2016, 8, 307–312. [Google Scholar] [CrossRef]
- Sepahi, S.; Mohajeri, S.A.; Hosseini, S.M.; Khodaverdi, E.; Shoeibi, N.; Namdari, M.; Tabassi, S.A.S. Effects of Crocin on Diabetic Maculopathy: A Placebo-Controlled Randomized Clinical Trial. Am. J. Ophthalmol. 2018, 190, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Razavi, B.M.; Hosseinzadeh, H. Pharmacokinetic Properties of Saffron and its Active Components. Eur. J. Drug Metab. Pharmacokinet. 2018, 43, 383–390. [Google Scholar] [CrossRef]
- Christodoulou, E.; Grafakou, M.E.; Skaltsa, E.; Kadoglou, N.; Kostomitsopoulos, N.; Valsami, G. Preparation, chemical characterization and determination of crocetin’s pharmacokinetics after oral and intravenous administration of saffron (Crocus sativus L.) aqueous extract to C57/BL6J mice. J. Pharm. Pharmacol. 2019, 71, 753–764. [Google Scholar] [CrossRef]
- Lautenschläger, M.; Sendker, J.; Hüwel, S.; Galla, H.J.; Brandt, S.; Düfer, M.; Riehemann, K.; Hensel, A. Intestinal formation of trans-crocetin from saffron extract (Crocus sativus L.) and in vitro permeation through intestinal and blood brain barrier. Phytomedicine 2015, 22, 36–44. [Google Scholar] [CrossRef]
- Almodóvar, P.; Briskey, D.; Rao, A.; Prodanov, M.; Inarejos-García, A.M. Bioaccessibility and Pharmacokinetics of a Commercial Saffron (Crocus sativus L.) Extract. Evid.-Based Complement. Altern. Med. eCAM 2020, 2020, 1575730. [Google Scholar] [CrossRef] [Green Version]
- Jafarisani, M.; Bathaie, S.Z.; Mousavi, M.F. Saffron carotenoids (crocin and crocetin) binding to human serum albumin as investigated by different spectroscopic methods and molecular docking. J. Biomol. Struct. Dyn. 2018, 36, 1681–1690. [Google Scholar] [CrossRef]
- Chryssanthi, D.G.; Lamari, F.N.; Georgakopoulos, C.D.; Cordopatis, P. A new validated SPE-HPLC method for monitoring crocetin in human plasma--application after saffron tea consumption. J. Pharm. Biomed. Anal. 2011, 55, 563–568. [Google Scholar] [CrossRef]
- Song, Y.-N.; Wang, Y.; Zheng, Y.-H.; Liu, T.-L.; Zhang, C. Crocins: A comprehensive review of structural characteristics, pharmacokinetics and therapeutic effects. Fitoterapia 2021, 153, 104969. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Qian, Z.; Du, P.; Fu, J. Pharmacokinetic properties of crocin (crocetin digentiobiose ester) following oral administration in rats. Phytomedicine 2007, 14, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Ajam, M.; Reyhani, T.; Roshanravan, V.; Zare, Z. Increased Miscarriage Rate in Female Farmers Working in Saffron Fields: A Possible Effect of Saffron Toxicity. Asia Pac. J. Med. Toxicol. 2014, 3, 73–75. [Google Scholar] [CrossRef]
- Abdullaev, F.I. Cancer Chemopreventive and Tumoricidal Properties of Saffron (Crocus sativus L.). Exp. Biol. Med. 2002, 227, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Karimi, G.; Taiebi, N.; Hosseinzadeh, H.; Shirzad, F. Evaluation of Subacute Toxicity of Aqueous Extract of Crocus sativus L. Stigma and Petal in Rats. Jmpir 2004, 3, 29–35. [Google Scholar]
- Ramadan, A.; Soliman, G.; Mahmoud, S.S.; Nofal, S.M.; Abdel-Rahman, R.F. Evaluation of the safety and antioxidant activities of Crocus sativus and Propolis ethanolic extracts. J. Saudi Chem. Soc. 2012, 16, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Babaei, A.; Arshami, J.; Haghparast, A.; Danesh Mesgaran, M. Effects of saffron (Crocus sativus) petal ethanolic extract on hematology, antibody response, and spleen histology in rats. Avicenna J. Phytomed. 2014, 4, 103–109. [Google Scholar]
- Hosseinzadeh, H.; Abootorabi, A.; Sadeghnia, H.R. Protective effect of Crocus sativus stigma extract and crocin (trans-crocin 4) on methyl methanesulfonate-induced DNA damage in mice organs. DNA Cell Biol. 2008, 27, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Skowroń, J.; Zapór, L.; Miranowicz-Dzierżawska, K. Classification of the Substances on the Basis of the Acute-Toxic-Class Method (ATC). Int. J. Occup. Saf. Ergon. 1998, 4, 107–116. [Google Scholar] [CrossRef]
- El Midaoui, A.; Ghzaiel, I.; Vervandier-Fasseur, D.; Ksila, M.; Zarrouk, A.; Nury, T.; Khallouki, F.; El Hessni, A.; Ibrahimi, S.O.; Latruffe, N.; et al. Saffron (Crocus sativus L.): A Source of Nutrients for Health and for the Treatment of Neuropsychiatric and Age-Related Diseases. Nutrients 2022, 14, 597. [Google Scholar] [CrossRef]
- Chan, S.-L.; Yu, V.C. Proteins of the Bcl-2 family in apoptosis signalling: From mechanistic insights to therapeutic opportunities. Clin. Exp. Pharmacol. Physiol. 2004, 31, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Azmi, A.S.; Wang, Z.; Philip, P.A.; Mohammad, R.M.; Sarkar, F.H. Emerging Bcl-2 inhibitors for the treatment of cancer. Expert Opin. Emerg. Drugs 2011, 16, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, H.A.; Hakkim, F.L.; Sam, S. Molecular Mechanism of Crocin Induced Caspase Mediated MCF-7 Cell Death: In Vivo Toxicity Profiling and Ex Vivo Macrophage Activation. Asian Pac. J. Cancer Prev. 2016, 17, 1499–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Yin, X.M.; Chao, D.T.; Milliman, C.L.; Korsmeyer, S.J. BID: A novel BH3 domain-only death agonist. Genes Dev. 1996, 10, 2859–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehri, S.; Abnous, K.; Mousavi, S.H.; Shariaty, V.M.; Hosseinzadeh, H. Neuroprotective effect of crocin on acrylamide-induced cytotoxicity in PC12 cells. Cell. Mol. Neurobiol. 2012, 32, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Soeda, S.; Ochiai, T.; Paopong, L.; Tanaka, H.; Shoyama, Y.; Shimeno, H. Crocin suppresses tumor necrosis factor-alpha-induced cell death of neuronally differentiated PC-12 cells. Life Sci. 2001, 69, 2887–2898. [Google Scholar] [CrossRef]
- Kim, B.; Lee, K.Y.; Park, B. Crocin Suppresses Constitutively Active STAT3 Through Induction of Protein Tyrosine Phosphatase SHP-1. J. Cell Biochem. 2017, 118, 3290–3298. [Google Scholar] [CrossRef]
- Beljebbar, A.; Sockalingum, G.D.; Morjani, H.; Angiboust, J.F.; Polissiou, M.; Manfait, M. Differential Interaction Modes of Dimethylcrocetin in K562 and HL60 Tumor Cells As Probed by Near Infrared FT-Raman Microspectroscopy. In Spectroscopy of Biological Molecules: 6th European Conference on the Spectroscopy of Biological Molecules, Villeneuve d’Ascq, France, 3–8 September 1995; Merlin, J.C., Turrell, S., Huvenne, J.P., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 475–476. [Google Scholar]
- Li, S.; Qu, Y.; Shen, X.Y.; Ouyang, T.; Fu, W.B.; Luo, T.; Wang, H.Q. Multiple Signal Pathways Involved in Crocetin-Induced Apoptosis in KYSE-150 Cells. Pharmacology 2019, 103, 263–272. [Google Scholar] [CrossRef]
- Bakshi, H.A.; Zoubi, M.S.A.; Hakkim, F.L.; Aljabali, A.A.A.; Rabi, F.A.; Hafiz, A.A.; Al-Batanyeh, K.M.; Al-Trad, B.; Ansari, P.; Nasef, M.M.; et al. Dietary Crocin is Protective in Pancreatic Cancer while Reducing Radiation-Induced Hepatic Oxidative Damage. Nutrients 2020, 12, 1901. [Google Scholar] [CrossRef]
- Schramm, A.; Lode, H. MYCN-targeting vaccines and immunotherapeutics. Hum. Vaccines Immunother. 2016, 12, 2257–2258. [Google Scholar] [CrossRef] [Green Version]
- Amin, B.; Abnous, K.; Motamedshariaty, V.; Hosseinzadeh, H. Attenuation of oxidative stress, inflammation and apoptosis by ethanolic and aqueous extracts of Crocus sativus L. stigma after chronic constriction injury of rats. An. Da Acad. Bras. Cienc. 2014, 86, 1821–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandegi, A.R.; Rashidy-Pour, A.; Vafaei, A.A.; Ghadrdoost, B. Protective Effects of Crocus sativus L. Extract and Crocin against Chronic-Stress Induced Oxidative Damage of Brain, Liver and Kidneys in Rats. Adv. Pharm. Bull. 2014, 4, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, G.K.; Chang, A.; Grigg, J.; McCluskey, P. Efficacy and Safety of Saffron Supplementation: Current Clinical Findings. Crit. Rev. Food Sci. Nutr. 2016, 56, 2767–2776. [Google Scholar] [CrossRef]
- Das, I.; Das, S.; Saha, T. Saffron suppresses oxidative stress in DMBA-induced skin carcinoma: A histopathological study. Acta Histochem. 2010, 112, 317–327. [Google Scholar] [CrossRef]
- Giaccio, M. Crocetin from saffron: An active component of an ancient spice. Crit. Rev. Food Sci. Nutr. 2004, 44, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Sadeghnia, H.R. Safranal, a constituent of Crocus sativus (saffron), attenuated cerebral ischemia induced oxidative damage in rat hippocampus. J. Pharm. Pharm. Sci. 2005, 8, 394–399. [Google Scholar]
- Kanakis, C.D.; Tarantilis, P.A.; Pappas, C.; Bariyanga, J.; Tajmir-Riahi, H.A.; Polissiou, M.G. An overview of structural features of DNA and RNA complexes with saffron compounds: Models and antioxidant activity. J. Photochem. Photobiol. B 2009, 95, 204–212. [Google Scholar] [CrossRef]
- Kanakis, C.D.; Tarantilis, P.A.; Tajmir-Riahi, H.A.; Polissiou, M.G. Crocetin, dimethylcrocetin, and safranal bind human serum albumin: Stability and antioxidative properties. J. Agric. Food Chem. 2007, 55, 970–977. [Google Scholar] [CrossRef]
- Mehri, S.; Abnous, K.; Khooei, A.; Mousavi, S.H.; Shariaty, V.M.; Hosseinzadeh, H. Crocin reduced acrylamide-induced neurotoxicity in Wistar rat through inhibition of oxidative stress. Iran. J. Basic Med. Sci. 2015, 18, 902–908. [Google Scholar]
- Naghizadeh, B.; Boroushaki, M.T.; Vahdati Mashhadian, N.; Mansouri, S.M.T. Protective Effects of Crocin against Cisplatin-Induced Acute Renal Failure and Oxidative Stress in Rats. Iran. Biomed. J. 2008, 12, 93–100. [Google Scholar]
- Serrano-Diaz, J.; Sanchez, A.M.; Maggi, L.; Martinez-Tome, M.; Garcia-Diz, L.; Murcia, M.A.; Alonso, G.L. Increasing the applications of Crocus sativus flowers as natural antioxidants. J. Food Sci. 2012, 77, C1162–C1168. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Bathaie, S.Z.; Abroun, S.; Azizian, M. Effect of Crocin on Cell Cycle Regulators in N-Nitroso-N-Methylurea-Induced Breast Cancer in Rats. DNA Cell Biol. 2015, 34, 684–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, X.; Zhang, Y.; Xue, Y.; Li, Z.; Shi, J.; Wang, H.; Chu, L. Crocin protects against cardiotoxicity induced by doxorubicin through TLR-2/NF-κB signal pathway in vivo and vitro. Int. Immunopharmacol. 2020, 84, 106548. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Cao, A.; Li, C.; Wang, L.; Wu, Q.; Li, X.; Lv, X.; Zhu, J.; Chun, H.; et al. Crocin Improves Endothelial Mitochondrial Dysfunction via GPx1/ROS/KCa3.1 Signal Axis in Diabetes. Front. Cell Dev. Biol. 2021, 9, 651434. [Google Scholar] [CrossRef] [PubMed]
- Nasimian, A.; Farzaneh, P.; Tamanoi, F.; Bathaie, S.Z. Cytosolic and mitochondrial ROS production resulted in apoptosis induction in breast cancer cells treated with Crocin: The role of FOXO3a, PTEN and AKT signaling. Biochem. Pharmacol. 2020, 177, 113999. [Google Scholar] [CrossRef]
- Tang, Y.; Yang, H.; Yu, J.; Li, Z.; Xu, Q.; Ding, B.; Jia, G. Crocin induces ROS-mediated papillary thyroid cancer cell apoptosis by modulating the miR-34a-5p/PTPN4 axis in vitro. Toxicol. Appl. Pharmacol. 2022, 437, 115892. [Google Scholar] [CrossRef]
- Nie, Z.; Deng, S.; Zhang, L.; Chen, S.; Lu, Q.; Peng, H. Crocin protects against dexamethasone-induced osteoblast apoptosis by inhibiting the ROS/Ca2+-mediated mitochondrial pathway. Mol. Med. Rep. 2019, 20, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Moradzadeh, M.; Ghorbani, A.; Erfanian, S.; Mohaddes, S.T.; Rahimi, H.; Karimiani, E.G.; Mashkani, B.; Chiang, S.C.; El-Khamisy, S.F.; Tabarraei, A.; et al. Study of the mechanisms of crocetin-induced differentiation and apoptosis in human acute promyelocytic leukemia cells. J. Cell Biochem. 2018, 120, 1943–1957. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Accession No | Primer F (5′-3′) | Primer R (5′-3′) |
|---|---|---|---|---|
| ACTB | actin, beta | NM_001101.3 | CTGTCCACCTTCCAGCAGATGT | AGCATTTGCGGTGGACGAT |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase | NM_001256799.2 | TTGCCCTCAACGACCACTTT | CACCCTGTTGCTGTAGCCAAA |
| BAX | BCL2 Associated X, Apoptosis Regulator | NM_001291428.1 | GGTTGTCGCCCTTTTCTA | CGGAGGAAGTCCAATGTC |
| BID | BH3 interacting domain death agonist | NM_001196.3 | TCCTTGCTCCGTGATGTCTTTC | AAGCTCCTCACGTAGGTGCGTA |
| BCL2 | BCL2 apoptosis regulator | NM_000633.2 | GATGTGATGCCTCTGCGAAG | CATGCTGATGTCTCTGGAATCT |
| MYCN | MYCN proto-oncogene, bHLH transcription factor | NM_001293228.1 | CCCCTGGGTCTGCCCCGTTT | GCCGAAGTAGAAGTCATCTT |
| SOD1 | superoxide dismutase 1 | NM_000454.4 | GGATGAAGAGAGGCATGTTGGA | TAGACACATCGGCCACACCAT |
| GSTM1 | glutathione S-transferase mu 1 | NM_000561.3 | ACTTGATTGATGGGGCTCAC | TCTCCAAAATGTCCACACGA |
| Cancer | Cell Line | IC50 | Range of Concentrations | Ref(s) |
|---|---|---|---|---|
| Bladder cancer | 5637 | 0.2 mg/mL | 0.05–4 mg/mL of SAE | [23,119] |
| Breast cancer | MDA-MB-231 | 0.5 mg/mL of SAE | 0.1–1 mg/mL of SAE | [25,62,66,80,81,98,105,120,121,122] |
| >0.195 mg/mL of trans-CRC-4 | 9.7696 μg/mL–0.976 mg/mL of trans-CRC-4 | |||
| BT-474 HER2+ | 3.5 mg/mL at 24 h of CRCs | 1–5 mg/mL of CRCs | ||
| MCF-7 | 0.35–0.78 mg/mL of SAE | 0.1–1 mg/mL of SAE | ||
| >0.195 mg/mL of trans-CRC-4 | 9.7696 μg/mL–0.976 mg/mL of trans-CRC-4 | |||
| 3.5 mg/mL at 48 h of treatment with CRCs | 1.5–6 mg/mL of CRCs | |||
| 0.05 mg/mL of CRCs | 0.01–0.2 mg/mL of CRCs | |||
| 3.5 mg/mL of CRCs | 2–5 mg/mL of CRCs | |||
| >4 mg/mL of CRCs | 1–4 mg/mL of CRCs | |||
| BT-549 | ~4 mg/mL of CRCs | 1–4 mg/mL of CRCs | ||
| MDA-MB-468 | 3–4 mg/mL of CRCs | 2–5 mg/mL of CRCs | ||
| 3 mg/mL of CRCs at 24 h 2.5 mg/mL of CRCs at 48 h 1.5 mg/mL of CRCs at 72 h | 1–5 mg/mL of CRCs | |||
| Cervical cancer | HeLa | 2.3 mg/mL of saffron ethanolic extract | 1–5 mg/mL of saffron ethanolic extract | [63,76,105,123,124,125,126] |
| 1.92 mg/mL of saffron extracts | 0.25–4 mg/mL of saffron extracts | |||
| 3–3.5 mg/mL of CRCs | 0.976–9.7696 mg/mL of CRCs | |||
| 0.072248 mg/mL of CRT | 0.328–1.31 mg/mL of CRT | |||
| SiHa | 3.9078 mg/mL of CRCs | 0.0587–15.613 mg/mL of CRCs | ||
| Sensitive human cervical cancer cell line OV2008 cells | 3 mg/mL of CRCs at 24 h 1.5–2.7 mg/mL of CRCs at 48 h 1–1.5 mg/mL of CRCs at 72 h | 1–5 mg/mL of CRCs | ||
| Resistant human cervical cancer cell line C13 | 7 mg/mL of CRCs at 24 h 5 mg/mL of CRCs at 48 h 2.5 mg/mL of CRCs at 72 h | 1–4 mg/mL of CRCs | ||
| Colorectal cancer | SW480 | >1.0 mg/mL of Crocus sativus extract | 0.25–3 mg/mL of Crocus sativus extract | [67,104,127,128] |
| 0.977 mg/mL of CRCs | 0.029–0.977 mg/mL of CRCs | |||
| HCT116 | 1.0 mg/mL of Crocus sativus extract | 0.25–3 mg/mL of Crocus sativus extract | ||
| 1.94 mg/mL of CRCs | 0.977–4 mg/mL of CRCs | |||
| >0.293 mg/mL of CRCs | 0.029–0.977 mg/mL of CRCs | |||
| 4.89–7.82 mg/mL of CRCs | 0.488–14.65 mg/mL of CRCs | |||
| 0.052 mg/mL of CRT | 0.328–1.31 mg/mL CRT | |||
| HT-29 human colon adenocarcinoma cells | >1.0 mg/mL of Crocus sativus extract | 0.25–3 mg/mL of Crocus sativus extract | ||
| >0.293 mg/mL of CRCs | 0.029–0.977 mg/mL of CRCs | |||
| 0.39 mg/mL of CRCs | 4.88 μg/mL–1.9539 mg/mL of CRCs | |||
| >40 μg/mL of CRCs | 10–40 μg/mL of CRCs | |||
| Caco-2 | ~40 μg/mL of CRCs | 10–40 μg/mL of CRCs | ||
| DHD/K12-PROb rat colon adenocarcinoma cells | 0.97696 mg/mL of CRCs | 4.88 μg/mL–1.9539 mg/mL of CRCs | ||
| Cutaneous squamous cell carcinoma | A431 | 3.9078 mg/mL of CRCs | 0.977–3.9078 mg/mL of CRCs | [129] |
| SCL-1 | 3.9078 mg/mL of CRCs | 0.977–3.9078 mg/mL of CRCs | ||
| Esophageal squamous carcinoma | KYSE-150 | 65.68 μg/mL of CRT | 4.105–65.68 μg/mL of CRT | [130,131] |
| Gastric cancer | AGS | 2.026–4 mg/mL of CRCs | 0.003–19.68 mg/mL of CRCs | [48,69,70,132,133] |
| 2 mg/mL of CRCs | 2–6 mg/mL of CRCs | |||
| 2.405 mg/mL of CRCs | 0.003–19.68 mg/mL of CRCs | |||
| SGC-7901 | 2.527 mg/mL of CRCs | 0.003–19.68 mg/mL of CRCs | ||
| HGC-27 | 2–4 mg/mL of CRCs | 2–6 mg/mL of CRCs | ||
| BGC-823 | 2.321 mg/mL of CRCs | 8–16 mg/mL of CRCs | ||
| EPG85-257 | ~78.15 μg/mL | 9.77–97.7 μg/mL | ||
| Head and neck | HN5 | 0.58 mg/mL of CRCs at 48 h 0.48 mg/mL of CRCs at 72 h 12.5–50 µg/mL of CRCs for 24, 48 and 72 h had no inhibitory effects on cell proliferation | 12.5–1000 μg/mL of CRCs | [45,132] |
| Hepatocellular carcinoma | HepG2 | 3 mg/mL of CRCs | 0.977–5 mg/mL of CRCs | [64,71,125] |
| 2.75–3.25 mg/mL of CRCs at 48 h | 0–10 mg/mL of CRCs | |||
| 0.2 mg/mL CRT | 0.328–1.31 mg/mL of CRT | |||
| HCCLM3 | 3 mg/mL of CRCs | 3–5 mg/mL | ||
| Leukemia | HL60 | 5 mg/mL of CRCs at 24 h | 0.625–10 mg/mL of CRCs | [25,72,73,134,135,136,137] |
| 3 mg/mL of CRCs at 48 h | 0.625–10 mg/mL of CRCs | |||
| >0.0328 mg/mL of CRT | 0.0016–0.0328 mg/mL of CRT | |||
| 11–39 mg/mL of CRCs | Non applicable | |||
| 7–30 mg/mL of DMCRT | Non applicable | |||
| MOLT-4 human T-cell leukemia cell line | >0.488 mg/mL of CRCs | 0.0488–0.488 mg/mL of CRCs | ||
| Jurkat cells | 2.5 mg/mL of CRCs at 24 h 1.25 mg/mL at 48 h of CRCs | 0.625–10 mg/mL of CRCs | ||
| CO 88BV59-1 EBV-transformed B-lymphocyte | 0.17 mg/mL of CRCs at 24 h 0.109 mg/mL of CRCs at 48 h 0.0774 mg/mL of CRCs at 72 h | 0.195 μg/mL–0.195 mg/mL of CRCs | [42] | |
| Lung adenocarcinoma | A549 | 170–380 μg/mL of SAE | 100–800 μg/mL of SAE | [11,74,125,138] |
| 1.5 mg/mL of saffron ethanolic extract at 24 h 0.565 mg/mL of saffron ethanolic extract at 48 h | 0.5–2 mg/mL of saffron ethanolic extract | |||
| 5.3537 mg/mL of CRCs | 0.977–4 mg/mL of CRCs | |||
| 4–5 mg/mL | 1–16 mg/mL of CRCs | |||
| 0.134 mg/mL CRT | 0.328–1.31 mg/mL of CRT | |||
| 4–5 mg/mL | 1–16 mg/mL | |||
| SPC-A1 | 4–5 mg/mL | 1–16 mg/mL | ||
| Neuroblastoma | SKNSH | 1.66 mg/mL of saffron extracts | 0.25–4 mg/mL of saffron extracts | [105] |
| Ovarian cancer | SK-OV-3 | 3.3 mg/mL of CRCs | 0.977–4 mg/mL of CRCs | [125,139] |
| 0.0623 mg/mL CRT | 0.328–1.31 mg/mL of CRT | |||
| A2780 | >0.0781 mg/mL of CRCs | 0.00977–0.0977 mg/mL of CRCs | ||
| Oral squamous cell carcinoma | KB | 1.9246 mg/mL of CRCs | 0.0488–3.9078 mg/mL of CRCs | [68] |
| Osteosarcoma | MG63 | 1.95–3.9 mg/mL of CRCs | 0.488–3.9 mg/mL of CRCs | [65] |
| OS732 | ||||
| Pancreatic cancer | BxPC-3 | >10 mg/mL of CRCs | 1–10 mg/mL of CRCs | [140] |
| Prostate cancer | Hormone-sensitive LAPC-4, CWR22 and LnCaP cells | 4.2-> 8 mg/mL of aqueous saffron extracts | 0.1–8 mg/mL of aqueous and alcoholic saffron extracts | |
| Hormone-insensitive 22rv1, C4–2B, DU145, PC3 cells | 0.8–7.9 mg/mL of alcoholic saffron extracts | 0.1–8 mg/mL of aqueous and alcoholic saffron extracts | [75] | |
| 0.254–0.928 mg/mL of CRCs | 0.0977–3.91 mg/mL of CRCs | |||
| Retinoblastoma | Y-79 | 0.0195–0.07815 mg/mL of CRCs | 0.00488–0.07815 mg/mL of CRCs | [141] |
| WERI-Rb-1 | 0.0195–0.07815 mg/mL of CRCs | 0.00488–0.07815 mg/mL of CRCs | ||
| Tongue Squamous Cell Carcinoma | Tca8113 | 0.218 mg/mL of CRCs | 0.098–0.781 mg/mL of CRCs | [142] |
| Cell Line | BCL2 | Effect | Ref. |
|---|---|---|---|
| Normal human liver cell L02 | ↑ | Anti-apoptotic | [94] |
| Zebrafish NAFLD (Non-Alcoholic Fatty Liver Disease) model | ↑ | Anti-apoptotic | [94] |
| MC3T3-E1 osteoblasts | ↑ | Anti-apoptotic (after treatment with dexamethasone) | [209] |
| PC12 cells rat pheochromocytoma | ↑ | Anti-apoptotic (after treatment with acrylamide) | [186] |
| Retinal ganglion cells (RGCs) | ↑ | Anti-apoptotic (after retinal ischemia/reperfusion-IR injury) | [39] |
| Rat neural stem cells | ↑ | Anti-apoptotic (after glucose deprivation or IR injury) | [89] |
| Bovine aortic endothelial cells | ↑ | Anti-apoptotic (after cells are exposed to H2O2) | [100] |
| T24 cell of transitional cell carcinoma of bladder (TCCB) | ↓ | Pro-apoptotic | [114] |
| EBV-Transformed B-Lymphocytes | ↓ | Pro-apoptotic | [42] |
| BXPC3 and Capan-2 pancreatic adenocarcinoma | ↓ | Pro-apoptotic | [191] |
| KYSE-150 cells esophageal squamous cell carcinoma | ↓ | Pro-apoptotic | [190] |
| Acute promyelocytic leukemia cells HL60, NB4 and primary APL cells | ↓ | Pro-apoptotic | [210] |
| HL60 acute promyelocytic leukemia cells | ↓ | Pro-apoptotic | [72] |
| Hep3B and HepG2 hepatoblastoma | ↓ | Pro-apoptotic | [47] |
| Gastric adenocarcinoma (AGS) cells | ↓ | Pro-apoptotic | [96] |
| Human multiple myeloma cells | ↓ | Pro-apoptotic | [188] |
| MCF-7 breast cancer cells | ↓ | Pro-apoptotic | [184] |
| Human prostate cancer cells (LAPC-4, PC3) | ↓ | Pro-apoptotic | [75] |
| Lung adenocarcinoma A549 and SPC-A1 | ↓ | Pro-apoptotic | [74] |
| Normal human liver cell L02 | ↓ | Anti-apoptotic | [94] |
| Zebrafish | ↓ | Anti-apoptotic | [94] |
| MC3T3-E1 osteoblasts | ↓ | Anti-apoptotic | [209] |
| PC12 cells rat pheochromocytoma | ↓ | Anti-apoptotic (after treatment with acrylamide) | [186] |
| Retinal ganglion cells (RGCs) | ↓ | Anti-apoptotic (after retinal ischemia/reperfusion-IR injury) | [39] |
| Rat neural stem cells | ↓ | Anti-apoptotic (after glucose deprivation or IR injury) | [89] |
| Bovine aortic endothelial cells | ↓ | Anti-apoptotic (after cells are exposed to H2O2) | [100] |
| EBV-Transformed B-Lymphocytes | ↑ | Pro-apoptotic | [42] |
| KYSE-150 cells esophageal squamous cell carcinoma | ↑ | Pro-apoptotic | [190] |
| Acute promyelocytic leukemia cells HL60, NB4 and primary APL cells | ↑ | Pro-apoptotic | [210] |
| HL60 acute promyelocytic leukemia cells | ↑ | Pro-apoptotic | [72,210] |
| Hep3B and HepG2 hepatoblastoma | ↑ | Pro-apoptotic | [47] |
| Gastric adenocarcinoma (AGS) cells | ↑ | Pro-apoptotic | [96] |
| MCF-7 breast cancer cells | ↑ | Pro-apoptotic | [184] |
| Human prostate cancer cells (LAPC-4, PC3) | ↑ | Pro-apoptotic | [75] |
| T24 cell of transitional cell carcinoma of bladder (TCCB) | ↑ | Pro-apoptotic | [114] |
| HL-60 Human Leukemia Cells | ↑ | Pro-apoptotic | [72] |
| Lung adenocarcinoma A549 and SPC-A1 | ↑ | Pro-apoptotic | [74] |
| Human multiple myeloma cells | ↑ | Pro-apoptotic | [188] |
| BXPC3 and Capan-2 pancreatic adenocarcinoma | ↑ | Pro-apoptotic | [191] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatziagapiou, K.; Nikola, O.; Marka, S.; Koniari, E.; Kakouri, E.; Zografaki, M.-E.; Mavrikou, S.S.; Kanakis, C.; Flemetakis, E.; Chrousos, G.P.; et al. An In Vitro Study of Saffron Carotenoids: The Effect of Crocin Extracts and Dimethylcrocetin on Cancer Cell Lines. Antioxidants 2022, 11, 1074. https://doi.org/10.3390/antiox11061074
Hatziagapiou K, Nikola O, Marka S, Koniari E, Kakouri E, Zografaki M-E, Mavrikou SS, Kanakis C, Flemetakis E, Chrousos GP, et al. An In Vitro Study of Saffron Carotenoids: The Effect of Crocin Extracts and Dimethylcrocetin on Cancer Cell Lines. Antioxidants. 2022; 11(6):1074. https://doi.org/10.3390/antiox11061074
Chicago/Turabian StyleHatziagapiou, Kyriaki, Olti Nikola, Sofia Marka, Eleni Koniari, Eleni Kakouri, Maria-Eleftheria Zografaki, Sophie S. Mavrikou, Charalabos Kanakis, Emmanouil Flemetakis, George P. Chrousos, and et al. 2022. "An In Vitro Study of Saffron Carotenoids: The Effect of Crocin Extracts and Dimethylcrocetin on Cancer Cell Lines" Antioxidants 11, no. 6: 1074. https://doi.org/10.3390/antiox11061074