
A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective


1
CAS Key Laboratory of Biofuels, and Shandong Provincial Key Laboratory of Synthetic Biology, Qingdao Institute of Bioenergy and Bioprocess Technology, Chinese Academy of Sciences, Qingdao 266101, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Antioxidants 2022, 11(3), 529; https://doi.org/10.3390/antiox11030529
Submission received: 16 February 2022
/
Revised: 6 March 2022
/
Accepted: 7 March 2022
/
Published: 10 March 2022
(This article belongs to the Special Issue Dream Peroxygenases)
Abstract
:Cytochrome P450 monooxygenases (P450s) are promising versatile oxidative biocatalysts. However, the practical use of P450s in vitro is limited by their dependence on the co-enzyme NAD(P)H and the complex electron transport system. Using H2O2 simplifies the catalytic cycle of P450s; however, most P450s are inactive in the presence of H2O2. By mimicking the molecular structure and catalytic mechanism of natural peroxygenases and peroxidases, an artificial P450 peroxygenase system has been designed with the assistance of a dual-functional small molecule (DFSM). DFSMs, such as N-(ω-imidazolyl fatty acyl)-l-amino acids, use an acyl amino acid as an anchoring group to bind the enzyme, and the imidazolyl group at the other end functions as a general acid-base catalyst in the activation of H2O2. In combination with protein engineering, the DFSM-facilitated P450 peroxygenase system has been used in various oxidation reactions of non-native substrates, such as alkene epoxidation, thioanisole sulfoxidation, and alkanes and aromatic hydroxylation, which showed unique activities and selectivity. Moreover, the DFSM-facilitated P450 peroxygenase system can switch to the peroxidase mode by mechanism-guided protein engineering. In this short review, the design, mechanism, evolution, application, and perspective of these novel non-natural P450 peroxygenases for the oxidation of non-native substrates are discussed.
1. Introduction
Cytochrome P450s (CYPs or P450s), a broad class of heme-containing enzymes, play important roles in drug metabolism, detoxification of xenobiotics, and steroid biosynthesis [1]. These enzymes are ubiquitous in nature, being found in animals, plants, bacteria, fungi, and other organisms [2]. P450s have potential use in the catalytic monooxygenation of various organic substrates, including aliphatic and aromatic compounds, alkenes, and compounds containing heteroatoms such as nitrogen and sulfur [3,4,5,6,7,8]. In particular, P450s can regio- and stereoselectively oxidize inert C–H bonds, thus acting as an attractive enzyme class in the development of practical biocatalysts for organic synthesis [9,10,11].
A variety of approaches have been developed to solve the intrinsic drawbacks of P450s, e.g., poor enzyme stability, low turnover rates, narrow substrate scope, and the need for expensive cofactors (NAD(P)H). Protein engineering, including rational design and directed evolution, represents a first choice for solving most of these issues [12,13,14]. Moreover, effective strategies have emerged to overcome some specific problems during P450-driven catalysis [15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33]. For example, researchers have constructed a substrate engineering approach to improve the acceptance and/or the stereo-/regioselectivity of non-native substrates of P450s by introducing protecting/anchoring/directing groups to the substrate [15,16,17,18,19,20,21,22,23,24]. Watanabe and co-workers used a dummy co-substrate (so-called decoy molecule) to modulate substrate promiscuity of P450s, enabling wild-type P450s to hydroxylate various small molecules that are not accepted in the absence of the decoy molecule (such as gas alkanes and benzene) [25,26,27]. Biological and chemical regeneration of NAD(P)H has been used widely to support catalysis by P450s [28,29]. In addition, the surrogate oxidants (e.g., hydrogen peroxide, tert-butyl hydroperoxide, and iodosylbenzene) are also used to drive P450 instead of molecular oxygen and reduced NAD(P)H [30,31,32,33]. Despite being useful supplements to protein engineering, these strategies often play a role in combination with protein engineering. There have been many reviews discussing the abovementioned topics [34,35,36,37,38,39,40,41]. Herein, we focus on a unique H2O2-dependent P450 peroxygenase system facilitated by a dual functional small molecule (DFSM). The design, construction, mechanism, and catalytic application of the DFSM-facilitated P450 peroxygenases are reviewed, and current issues and future perspectives are also discussed.
2. Proof-of-Concept of the DFSM-Facilitated P450 Peroxygenase
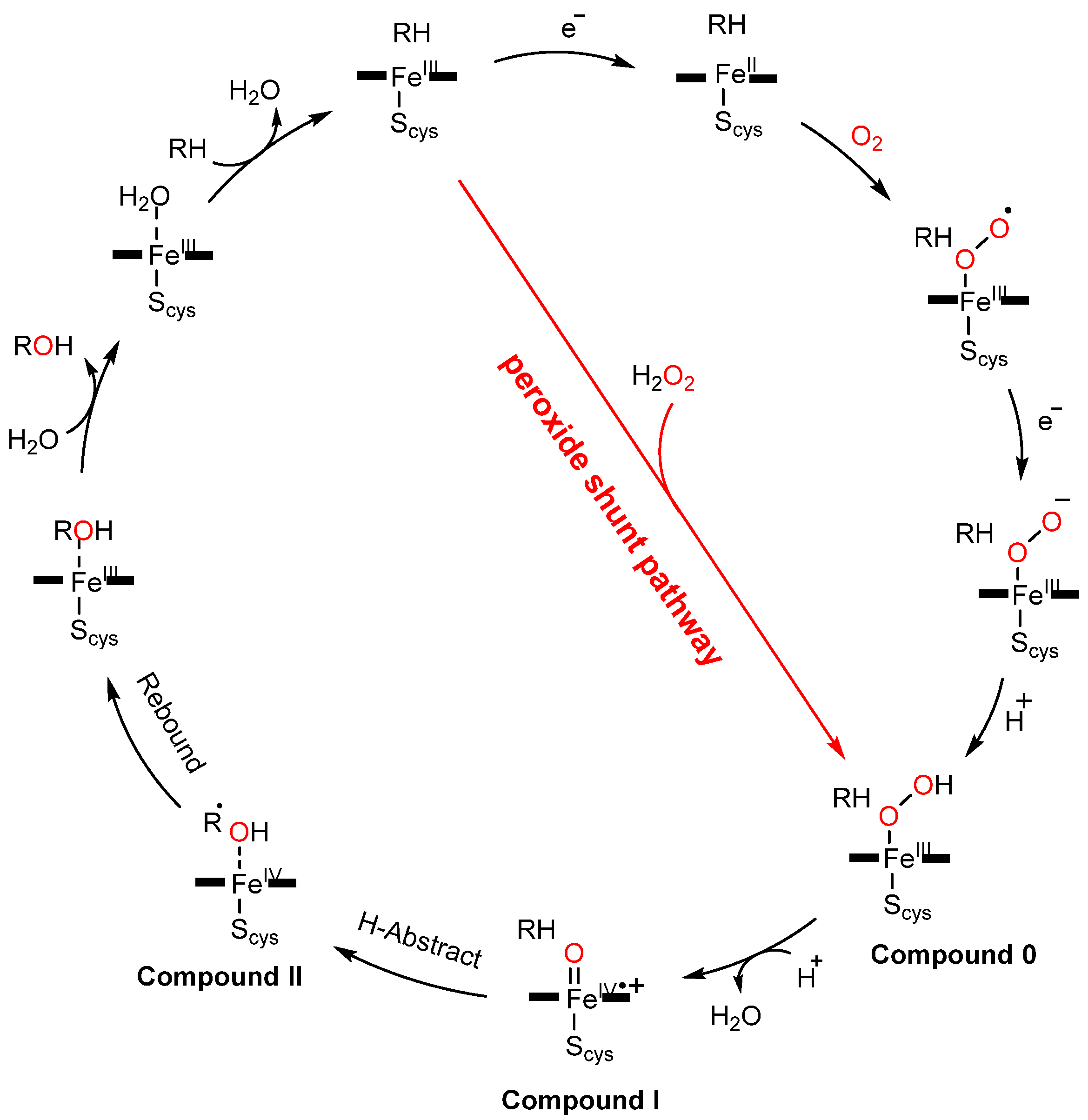
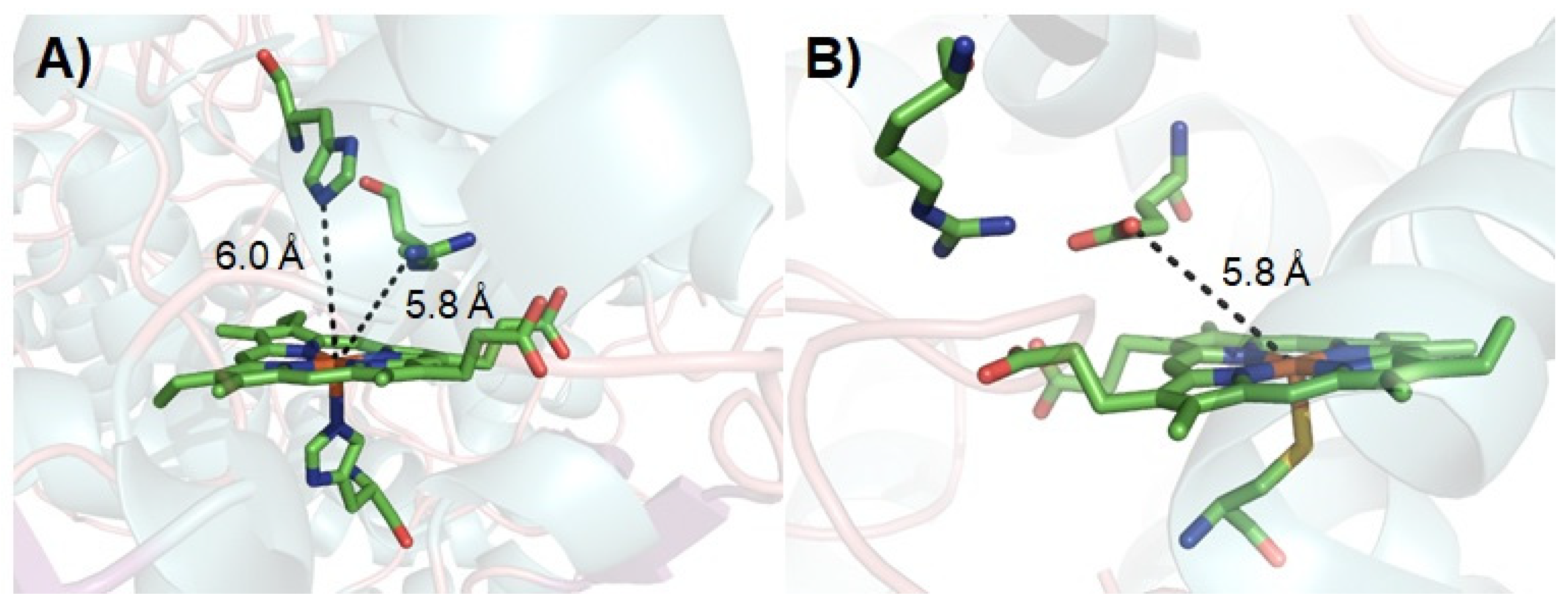
The complex catalytic cycle of P450s needs reduced co-enzyme NAD(P)H and a redox partner to support the activation of molecular oxygen. Thus, it had been suggested that surrogate peroxide species can be used to drive P450 catalysis through its shunt pathway (Figure 1), with low-cost H2O2 being one of the best choices. However, only a few native P450 peroxygenases (e.g., CYP 152 family) can use the unique substrate-assisted mechanism to activate H2O2 successfully [42,43,44,45,46,47], with most P450s examined (e.g., rat liver microsomal P450, human P450s such as CYP1A2 and 3A4, thermophilic archaea CYP119, CYP175A1, and P450cam) generally showing low efficiency for the H2O2-dependent reaction (shunt pathway in Figure 1) [48,49,50,51,52,53]. Although the peroxygenase and peroxidase activity of P450s can be partially improved by directed evolution, the catalytic efficiency of the evolved P450 variants is still not comparable to natural NAD(P)H-dependent P450s [54,55,56,57]. This may be caused by the inherent structural characteristics of P450s. Indeed, those enzymes that make good use of H2O2 in nature have acid-base amino acid residue pairs that play the role of an acid–base catalyst in their active site (Figure 2) [58,59]. In contrast, the crystal structures of other P450s have revealed that such amino acid residues are not present on the distal side of their heme centers. Previous reports have suggested that the introduction of a basic residue can modify myoglobin into a peroxidase through site-directed mutagenesis [60,61,62]. Similar strategies have been applied to improve the peroxygenase/peroxidase activity of P450s [63,64]; however, the catalytic efficiency was not always satisfactory. Crystal structure studies have provided hints for the poor activity in some cases, namely, the basic group on the side chain of the introduced residue is distal from the heme center such that this residue cannot efficiently activate H2O2 as the general acid–base catalyst [63].
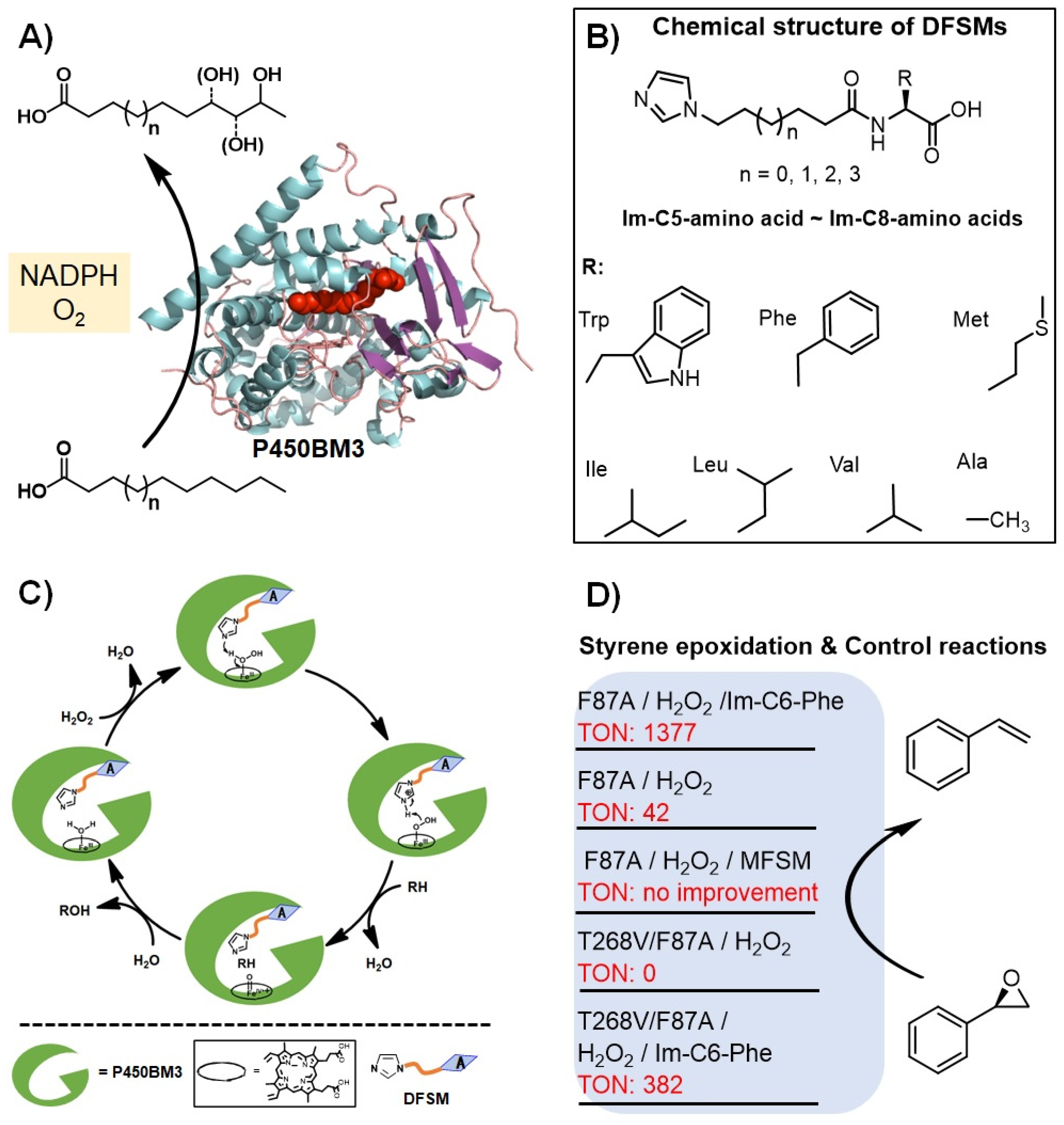
Based on previous reports, it has become clear that to use the shunt pathway of P450s efficiently, two points should be met: (1) a basic group located on the distal side of the heme center is necessary; and (2) the basic group should be placed at a suitable position to ensure that this residue plays the role as an acid-base catalyst efficiently. To this end, Ma et al. designed a DFSM approach to modify cytochrome P450BM3 monooxygenase from Bacillus megaterium into its peroxygenase mode (Figure 3) [65,66,67]. Typical structures of DFSMs, such as N-(ω-imidazolyl)-fatty acyl-l-amino acid (Im-Cn-AA), are shown in Figure 3B [66]. These DFSMs have an acyl amino acid moiety at one end as an anchoring group to bind with the enzyme, and an imidazolyl group at the other end as a basic group to assist the activation of H2O2. Moreover, the position of imidazolyl can be optionally tuned by changing the chain length of a flexible spacer having various CH2 numbers, while the introduced basic residues by site-directed mutagenesis often can’t extend sufficiently into a suitable catalytic site [63]. Ma et al. reasoned that the DFSM-facilitated P450BM3-H2O2 system was capable of running smoothly with a catalytic cycle that was similar to the native UPO peroxygenase under ideal conditions (Figure 3C) [68,69].
This concept was firstly validated by the H2O2-dependent epoxidation of styrene catalyzed by the P450BM3_F87A mutant. The presence of the best DFSM, N-(ω-imidazolyl)-hexanoyl-l-phenylalanine (Im-C6-Phe), increased the catalytic turnover number (TON) more than 30-fold than that of the F87A alone. The roles of the DFSMs were further demonstrated by using mono-functional small molecules (MFSMs) without the terminal imidazolyl group or acyl amino acid group, the latter didn’t improve TON and even inhibit the reactions. The ability of the DFSMs to generate peroxygenase activity was further demonstrated by using the double mutant F87A/T268V. The authors found that mutating the highly conserved T268 [69,70,71,72,73] abolished the H2O2 activity of the enzyme, which can be recovered upon the addition of DFSM. This discovery provides a unique choice of protein engineering sites for developing catalytic promiscuity of the current peroxygenase system (will be discussed below by combination with other results).
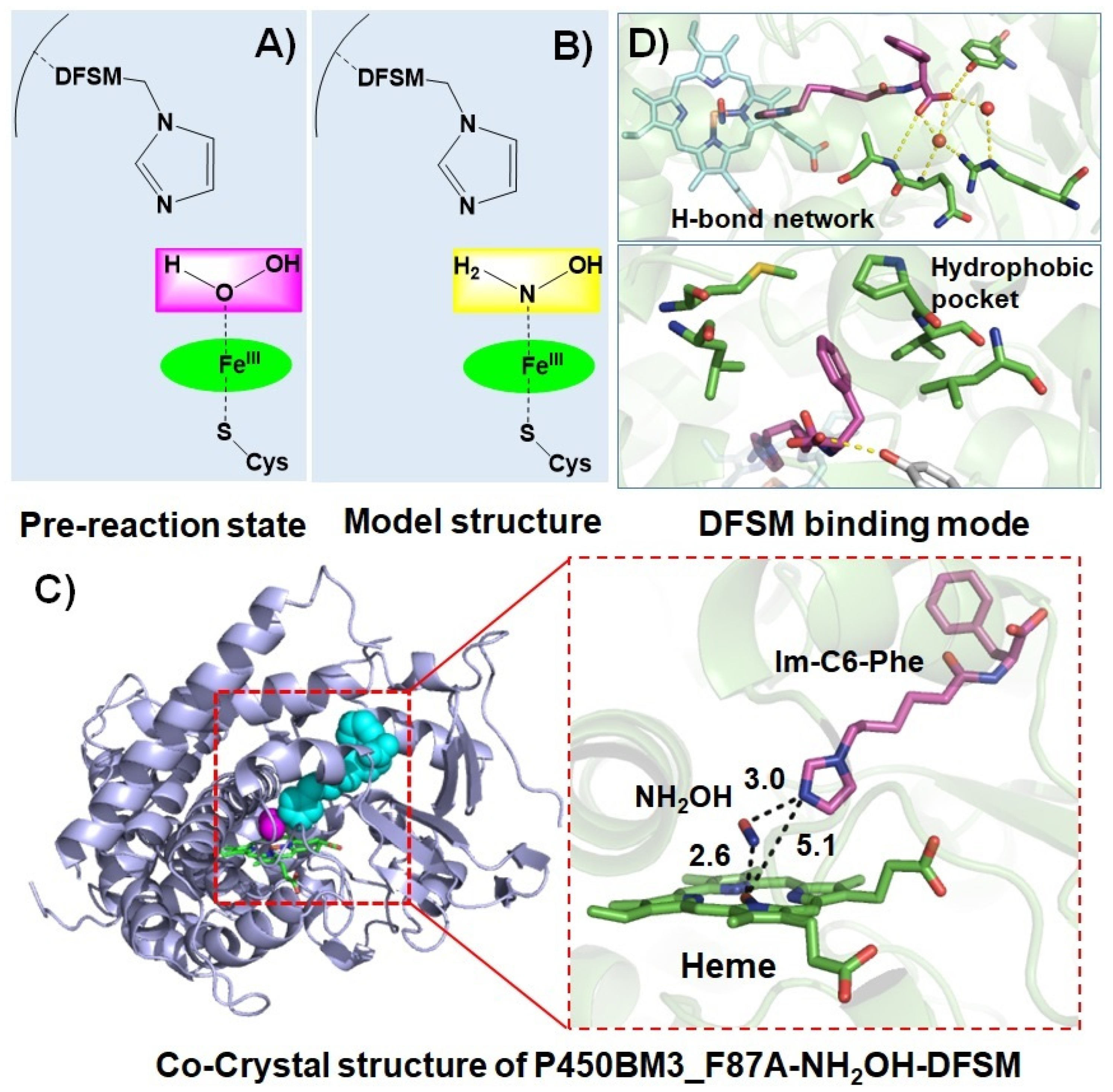
The catalytic role and mechanism of DFSMs have been further disclosed by combining structural biology and computational chemistry [74]. To mimic the pre-reaction state of P450-bounded H2O2 and avoid the H2O2-initiated reaction, Jiang et al. skillfully adopted the NH2OH molecule as the analog of H2O2 to prepare the co-crystal (Figure 4A,B). As a result, they successfully reported the first X-ray structure of the P450BM3 heme domain F87A mutant in complex with the DFSM, N-(ω-imidazolyl)-hexanoyl-l-phenylalanine (Im-C6-Phe) and NH2OH at 2.70 Å resolution (PDB ID: 7EGN, Figure 4C). The crystal structure clearly shows that Im-C6-Phe bound to P450BM3 through an H-bond network formed by interactions of its terminal carboxyl group with Arg47 and Tyr51, and hydrophobic interactions between its benzyl moiety and a hydrophobic pocket composed of Pro25, Val26, Leu29, Met185, and Leu188 (Figure 4D). The unique binding mode that involves additional hydrophobic interactions is distinct from those observed in the co-crystals of P450BM3 in complex with fatty acids (native substrates) or perfluoroacyl amino acids (decoy molecules) [75,76,77]. This binding mode plays a crucial role in positioning the imidazolyl group of the DFSM above the heme center, where the distance between the heme iron atom and the terminal nitrogen atom of the imidazolyl group is ~5 Å, indicating the imidazolyl group of the DFSM may act as a general acid–base catalyst in H2O2 activation, consistent with the original hypothesis by Ma et al. [66].
The mechanism for H2O2 activation was further elucidated by QM-MM computational investigations. These computational chemistry results revealed the crucial role of DFSM in promoting a heterolytic O–O cleavage to favor Cpd I formation [74]. The DFSM facilitates the formation of a proton channel between the imidazolyl group of the DFSM and proximal H of H2O2, thus enabling a heterolytic O–O cleavage and Cpd I formation, which is similar to the proposed mechanism for H2O2 activation in natural peroxygenases (e.g., UPO) or peroxidases (e.g., HRP). In contrast, the formation of Cpd I is apparently sluggish via the O–O homolysis mechanism in the absence of the DFSM. Similar results were also observed in the theoretical simulation of H2O2 activation by the P450cam T252A mutant [78], indicating weak H2O2 activation by NADPH-dependent P450s.
3. Catalytic Applications of the DFSM-Facilitated P450 Peroxygenase
In recent years, peroxygenase UPO has attracted considerable attention because of its versatile oxidation functions and potential in synthetic applications [79,80,81,82,83,84,85,86]. Moreover, peroxygenase that uses green and economic H2O2 to circumvent the use of expensive NADPH and the complex electron transfer system (redox partner proteins) has become a promising practical bio-oxidative catalyst when compared with using NAD(P)H-dependent P450 monooxygenases [79]. Despite concerns about the potential damage of H2O2 to enzymes, the use of a controlled fed-batch reactor or in situ-generating H2O2 has been demonstrated to enhance effectively the stability of peroxygenases through control of the H2O2 concentration in the reaction system, resulting in high catalytic turnovers [87,88,89,90,91]. Therefore, developing the catalytic potential of the artificial P450 peroxygenase is not only expected to expand the chemical space of P450 enzymes but also act as a beneficial supplement to the relatively scarce natural peroxygenase resources in nature. In fact, the DFSM-facilitated P450BM3-H2O2 system has shown versatile unique catalytic activity towards the peroxygenation reaction of various non-native substrates, such as epoxidation, hydroxylation, and sulfoxidation [66,92,93,94,95].
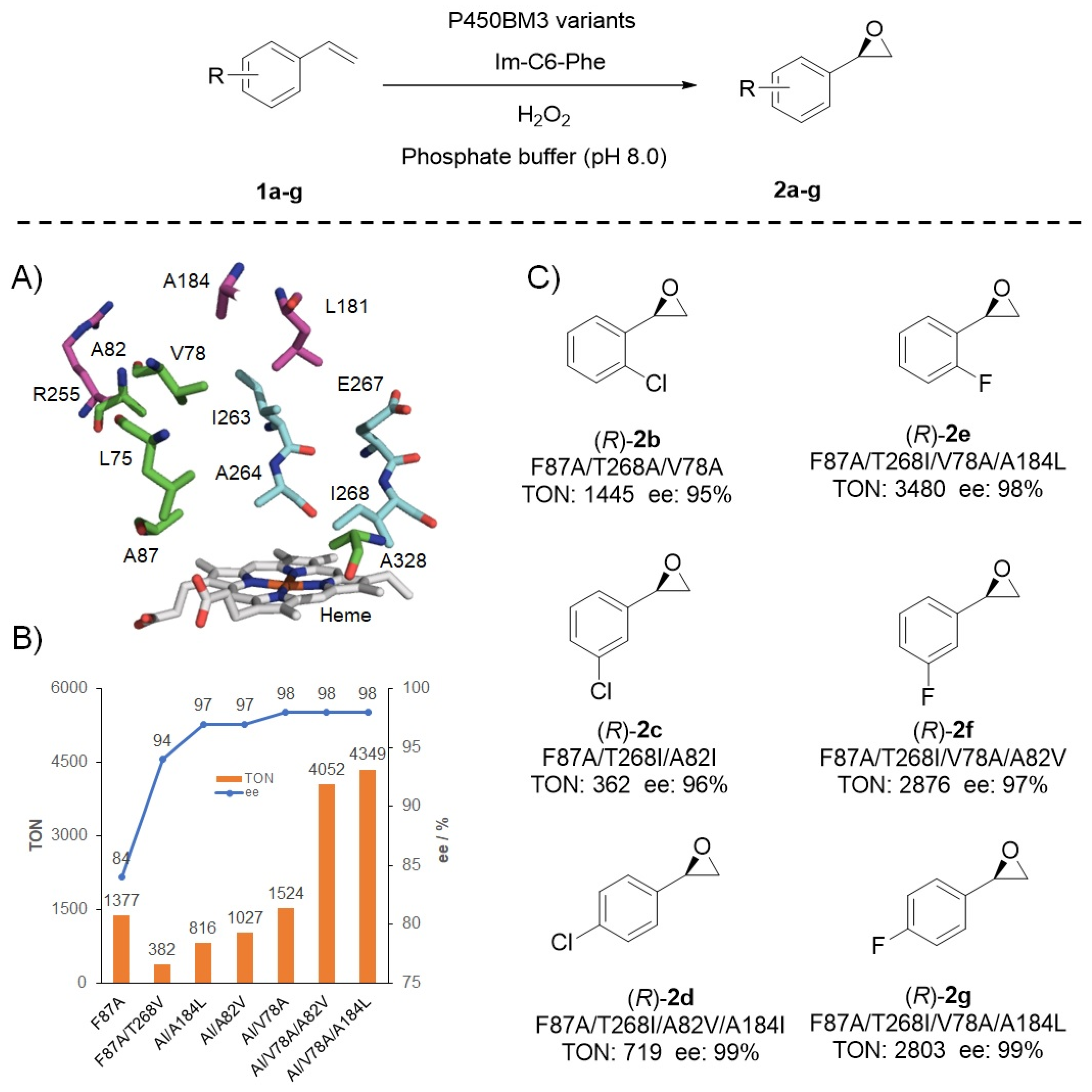
Asymmetric epoxidation of unfunctionalized olefins represents an important organic transformation to prepare optically pure epoxides [96,97,98,99]; however, the (R)-enantioselective epoxidation of styrene seems more difficult to achieve than the (S)-enantioselective reaction through either synthetic molecular catalysts or natural enzymatic bio-catalysts [100,101,102,103,104,105,106,107,108]. DFSM-facilitated P450BM3 peroxygenase enabled access to (R)-enantioselective epoxidation of unfunctionalized styrene and its derivatives (Figure 5). In view of the potential of the double mutant F87A/T268V in the (R)-enantioselective epoxidation of styrene in the presence of Im-C6-Phe, Zhao et al. systematically evaluated the effect of T268 residue and disclosed the roles of the T268 mutation in tuning activity and enantioselectivity of the NAD(P)H- and H2O2-dependent P450BM3 system, respectively [45]. Based on the more selective, but lower activity profile of the double mutant F87A/T268I (97% ee, TON = 335), a mutant library was constructed by introducing additional mutations at ten key residues around the substrate-binding pocket (Figure 5A). Two beneficial mutants were determined to give high (R)-enantioselective epoxidation of styrene (98% ee) with >4000 TONs (Figure 5B). This approach also gave modest to very good TONs (362–3480) and high (R)-enantioselectivities (95–99% ee) for the epoxidation of various styrene derivatives (Figure 5C), being comparable with the best (R)-enantioselective styrene monooxygenases, such as SeStyA from Streptomyces exfoliatus, AaStyA from Amycolatopsis albispora, and PbStyA from Pseudonocardiaceae reported recently [109,110]. The further semi-preparative scale experiments suggest its potential application in styrene epoxidation [92].
The direct hydroxylation of small alkanes to alcohols is a long-standing challenge because of the higher bond dissociation energies (BDE) of their C–H bonds when compared with that of the corresponding hydroxylated products, the latter easily leads to overoxidation [111,112]. Natural oxidizing enzymes, such as methane monooxygenase, soluble butane monooxygenase (sBMO), fungal peroxygenase (AaeUPO), and engineered P450s, are promising biocatalysts for the selective hydroxylation of small alkanes [76,77,93,113,114,115,116,117,118,119,120,121,122,123]. Recently, Chen et al. reported the peroxide-driven hydroxylation of small alkanes (C3–C6) by using engineered P450BM3 variants assisted by DFSMs [93]. Compared with some main results through enzymatic hydroxylation of small alkanes [116,117,118,119,120,121,122,123,124], DFSM-facilitated P450BM3 peroxygenase showed unique features and catalytic activities (Table 1). The hydrophobic mutation of T268 residue substantially improved the hydroxylation activities of small alkanes, which is distinct from NADPH-dependent P450 enzymes [94]. Here, the presence of the DFSM was critical for accomplishing the catalytic functions of engineered P450BM3 variants because the activity is completely lost in the absence of the DFSM. Two triple-mutants BM3_F87A/T268I/A184I and BM3_F87A/T268I/A82T showed the highest total turnover numbers (TTN) for the hydroxylation of propane and n-Butane (Entries 1–2 in Table 1), respectively, with better activity than AaeUPO, the only known H2O2-dependent native hydroxylase for small alkanes (entries 20–21 in Table 1) [116], and comparable activity to the P450BM3 decoy system (entries 3–8 in Table 1) [117,118,119,120], but far lower than P450PMOR1 and P450PMOR2, two evolved NADPH-dependent propane monooxygenases (entries 13–14 in Table 1) [121]. Notably, the product formation rates (PFR) for 2-propanol and 2-butanol of the current artificial P450 peroxygenase are far better than all reported natural or engineered enzyme systems. The contradiction between high PFR and low TTN suggests that the DFSM-facilitated P450 peroxygenase may be unstable. Nonetheless, reducing instability should yield an efficient biocatalyst for the direct hydroxylation of small alkanes. In addition, this peroxygenase system is unavailable for the hydroxylation of smaller alkanes (e.g., ethane and methane), which has been achieved by natural methane monooxygenase (MMO) or other enzymes (entries 9, 12, 16, 17 in Table 1) [76,77,118,122,123]. Anyhow, Ciuffetti et al. reported that CYP52L1 from Graphium sp. ATCC 58,400 can oxidize propane, but without any turnover numbers or catalytic constants mentioned [124]. This may be the only known P450 enzyme that uses gaseous alkanes as natural substrates, suggesting that P450 has a weak preference for small alkanes. Therefore, further protein engineering may be necessary for the DFSM-facilitated P450BM3 peroxygenase to access the direct hydroxylation of methane or ethane.
The O-demethylation of aromatic ethers is of important reaction to produce value-added phenolic compounds, which is also involved in aromatic ring-opening reactions of coniferyl and sinapyl lignin derivatives [125,126]. Various powerful oxidative enzymes, such as peroxidases from white-rot, soft-rot, and brown-rot fungi, as well as some bacteria, can catalyze demethylation of lignin-derived compounds and their model compounds [127,128,129,130]. A few P450 enzymes also show promise as an O-demethylase for lignin-derived aromatic ethers [131,132,133,134,135,136]. Recently, Jiang et al. successfully applied the DFSM-facilitated P450BM3 peroxygenase system to perform O-demethylation of various aromatic ether substrates (Table 2) [94]. These reactions show excellent regioselectivity toward the hydroxylation of the methoxy of aromatic ethers to give the demethylation product after automatically releasing formaldehyde. A suitable combination of the beneficial mutant and DFSM is important for controlling good regioselectivity. For example, some combinations examined still give aromatic hydroxylation as the main product. Although the DFSM-facilitated P450BM3 peroxygenase appears to open a new avenue for the key demethylation step in the bioconversion of lignin, it is still restricted by low TONs and narrow substrate scopes.
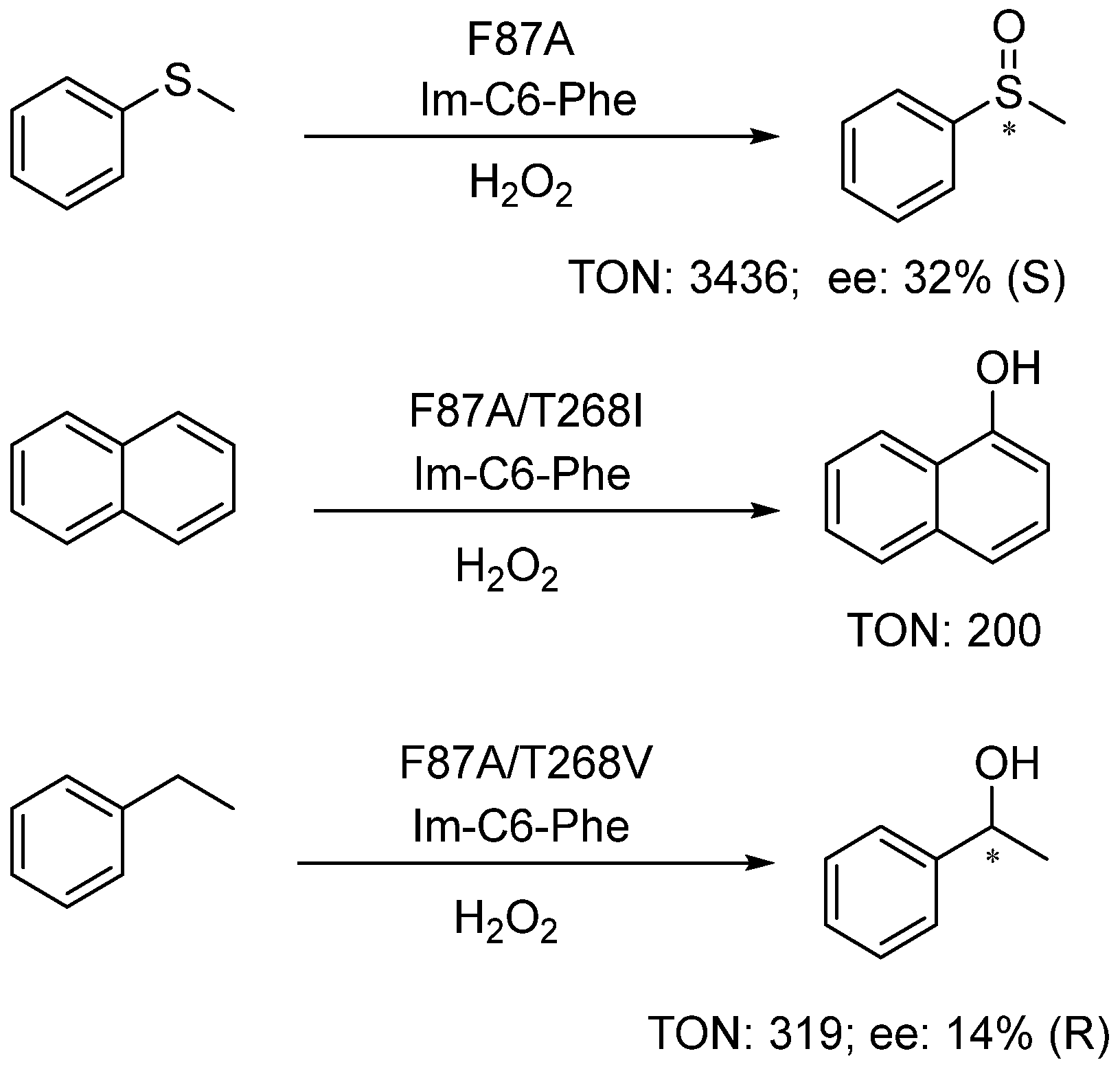
The DFSM-facilitated P450BM3 peroxygenase is also capable of catalyzing other types of reactions, including thioanisole sulfoxidation, aromatic hydroxylation of naphthalene, and the benzylic hydroxylation of ethylbenzene (Figure 6) [66,95]. The F87A mutant exclusively yielded sulfoxide with a PFR of 888 µmol·min−1·(µmol P450)−1 and a catalytic TON of 3436 in the presence of the DFSM, 35-fold higher than that without the DFSM. Interestingly, T268 mutations drastically affected the hydroxylation activity of the system, e.g., the double mutants F87G/T268V and F87A/T268V increased the TONs for the formation of 1-naphthol and 1-phenylethanol to 200 and 319, being around 15-fold and 8-fold to the single mutant F87G and F87A, respectively [95].
4. Switching Peroxidase Activity of the DFSM-Facilitated P450 Peroxygenase
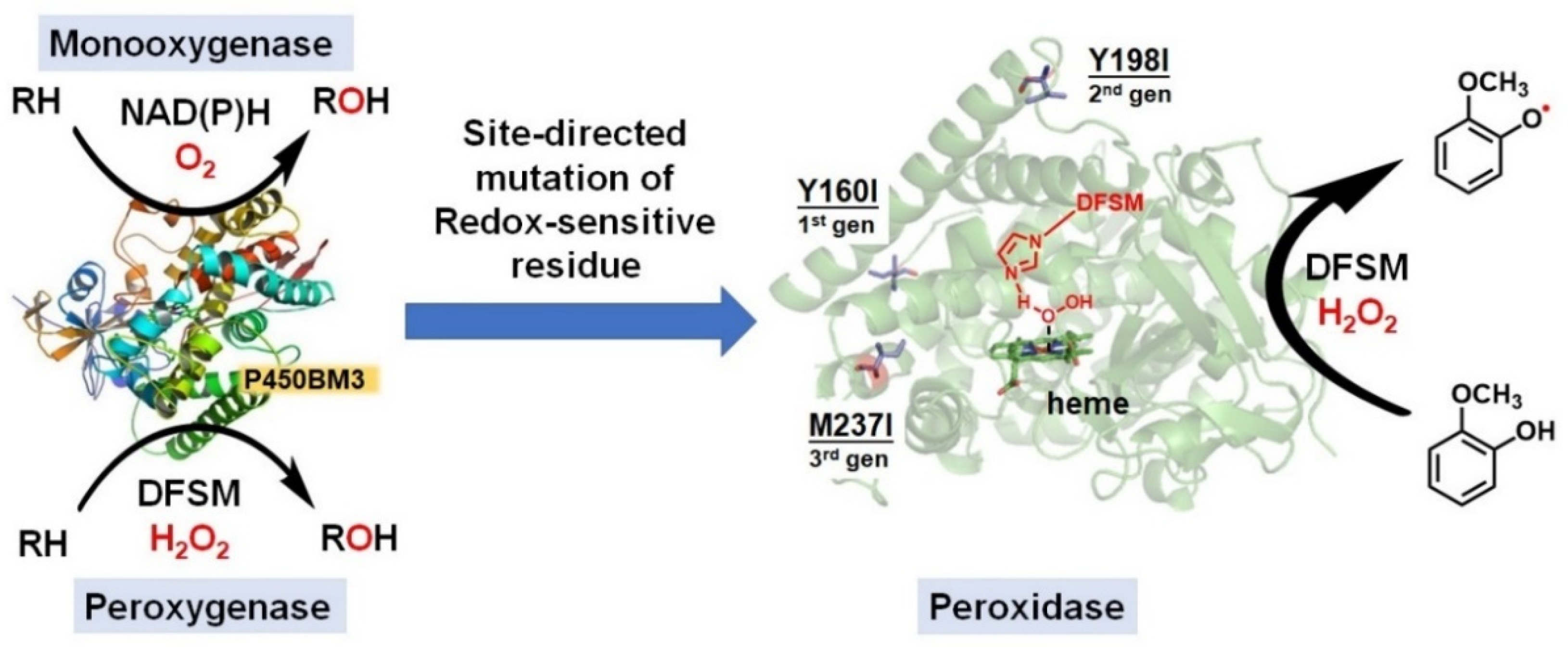
The catalytic promiscuity of enzymes is a fascinating topic for the biochemistry, synthetic biology, and chemical biology communities [137,138,139]. P450s have been well documented to carry out multiple catalytic functions such as monooxygenase, peroxygenase, and peroxidase activity [140]. However, research interest has focused on the monooxygenase and peroxygenase activities of P450s, and only a handful of studies have examined the catalytic peroxidase functionality of P450s. The non-natural DFSM-facilitated P450-H2O2 system described above mainly catalyzes various per-oxygenation reactions, including epoxidation, hydroxylation, and sulfoxidation [66,92,93,94,95]. Interestingly, the oxidation of guaiacol, a classical substrate of peroxidases [141,142,143,144], catalyzed by the DFSM-facilitated P450BM3-H2O2 system yielded demethylated catechol as a major product, suggesting it mainly functioned as a peroxygenase but not as a peroxidase [94]. After carefully analyzing the catalytic mechanism of the potential competitive oxidation pathways in the DFSM-facilitated P450BM3-H2O2 system, Ma et al. hypothesized that mutation of redox-sensitive residues may enable switching of peroxygenase activity to peroxidase activity [145]. Using a semi-rational design approach, similar to FRISM (focused rational iterative site-specific mutagenesis) named by Reetz and Wu [146,147], Ma et al. identified mutations of three key redox-sensitive tyrosine residues that are located on the surface of P450. Screening for activity-enhanced peroxidase mutants yielded a mutant that efficiently catalyzed one-electron oxidation of guaiacol through combination with other redox-sensitive residues located in the electron transfer pathway. The engineered system also exhibits favorable one-electron oxidation activity toward other peroxidase substrates, including 2,6-dimethoxyphenol, o-phenylenediamine, and p-phenylenediamine, and almost without peroxygenase activity for these substrates. Notably, this system attains the best peroxidase activity of any P450 reported [56,148], and rivals most natural peroxidases [149,150,151,152,153], suggesting significant potential for catalytic promiscuity of the DFSM-facilitated P450BM3-H2O2 system (Figure 7). Future efforts should explore the functional applications of the DFSM-facilitated P450 peroxidase in synthetic chemistry.
5. Summary and Perspectives
In summary, although only a few natural P450s, such as CYP152 peroxygenases from Sphingomonas paucimobilis, Bacillus subtilis, and Clostridium acetobutylicum can directly use an oxygen atom from peroxides for oxidation reactions [42,43,44,45,46,47], the engineered artificial P450 peroxygenases have significantly expanded the substrate scope and reaction types of P450-catalyzed per-oxygenation reactions. Therefore, it is no exaggeration to state that H2O2-driven P450 peroxygenases are emerging as powerful bio-oxidation catalysts. Among these, the DFSM-facilitated P450 peroxygenases provide a novel and unique solution for the efficient use of H2O2 by P450s, which exhibit much higher H2O2 activities in various reactions when compared with those P450 peroxygenases that have been engineered through site-directed mutagenesis and directed evolution [26,27,28,56,63,64,77,152,153]. Moreover, the DFSM-facilitated P450 peroxygenases may offer better opportunities for enhancing the regio- and enantioselectivity in oxidation reactions of non-natural substrates. On the one hand, the introduced DFSMs can influence the orientation of substrates through interaction with each other to modulate reaction selectivity, besides its role in the activation of H2O2, which still requires further experimental characterization. On the other hand, the highly conserved T268 residue can be optionally mutated in the DFSM-facilitated P450 peroxygenase system. In contrast, the mutation of T268 is not favorable in NADPH-dependent P450BM3 oxidation because this residue is located on the proximal side of the heme center and is thought to play multiple roles in NADPH-dependent catalysis [70,71,72,73,154]. In fact, successful examples of the DFSM-facilitated P450 peroxygenase system have demonstrated that mutation of T268 has a significant influence on regulating the substrate pocket space when employing a protein engineering strategy [45,46,47,48]. This suggests that protein engineering of the DFSM-facilitated P450 peroxygenase system may also have its own unique advantages for controlling reaction selectivity in comparison with natural NADPH-dependent P450s. In addition, the high peroxidase activity of the DFSM-facilitated P450-system developed recently expanded the catalytic promiscuity of the system [145], whose further application in organic transformation is expected.
The unique selectivity and activity of the DFSM-facilitated P450 peroxygenase system have shown its potential to be as a promising bio-oxidative catalyst; however, it is worth noting that there are still some drawbacks to hamper its further industrially utilization: (1) despite high efficiency, the introduction of DFSM undoubtedly increases the cost of the catalytic reaction, especially when a large excess is required; (2) oxidative damage of P450 caused by the presence of a large amount of H2O2; (3) the uncertainty associated with applying this strategy to other P450s; (4) the complex structures of DFSMs lead to an increase in the threshold of popularization and use; (5) the full catalytic mechanism still needs to be elucidated. In conclusion, the DFSM-facilitated P450 peroxygenase system simultaneously faces opportunities and challenges. Maximizing the potential of the system and answering the above issues will open new avenues for developing P450-based biocatalysts.
Author Contributions
Conceptualization and funding acquisition, Z.C.; writing—original draft preparation, review and editing, S.D., S.F., F.J. and Z.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 21977104 and 21778060; the Qingdao Innovative Leading Talent Project, grant number 18-1-2-9-zhc; QIBEBT, grant number I201901 and Y872361901, and Director Innovation Fund of Key Laboratory of Biofuels, Chinese Academy of Sciences, grant number Y872131901.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ortiz de Montellano, P.R.O. Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Kluwer Academic: New York, NY, USA; Plenum Publishers: New York, NY, USA, 2005. [Google Scholar]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta. Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R.; Urlacher, V.B. Cytochromes P450 as promising catalysts for biotechnological application: Chances and limitations. Appl. Microbiol. Biotech. 2014, 98, 6185–6203. [Google Scholar] [CrossRef] [PubMed]
- Fasan, R. Tuning P450 Enzymes as Oxidation Catalysts. ACS Catal. 2012, 2, 647–666. [Google Scholar] [CrossRef]
- de Montellano, P.R.O. Hydrocarbon Hydroxylation by Cytochrome P450 Enzymes. Chem. Rev. 2010, 110, 932–948. [Google Scholar] [CrossRef] [Green Version]
- Auclair, K.; Polic, V. Monoxygenase, Peroxidase and Peroxygenase Properties and Mechanisms of Cytochrome P450; Hrycay, E.G., Bandiera, S.M., Eds.; Springer: New YorK, NY, USA, 2015; pp. 209–228. [Google Scholar]
- Nastri, F.; Chino, M.; Maglio, O.; Bhagi-Damodaran, A.; Lu, Y.; Lombardi, A. Design and engineering of artificial oxygen-activating metalloenzymes. Chem. Soc. Rev. 2016, 45, 5020–5054. [Google Scholar] [CrossRef]
- Wang, T.T.; Fan, X.T.; Hou, C.X.; Liu, J.Q. Design of artificial enzymes by supramolecular strategies. Curr. Opin. Struct. Biol. 2018, 51, 19–27. [Google Scholar] [CrossRef]
- Wei, Y.F.; Ang, E.L.; Zhao, H.M. Recent developments in the application of P450 based biocatalysts. Curr. Opin. Chem. Biol. 2018, 43, 1–7. [Google Scholar] [CrossRef]
- McLntosh, J.A.; Farwell, C.C.; Arnold, F.H. Expanding P450 catalytic reaction space through evolution and engineering. Curr. Opin. Chem. Biol. 2014, 19, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Li, S.Y. Expansion of chemical space for natural products by uncommon P450 reactions. Nat. Prod. Rep. 2017, 34, 1061–1089. [Google Scholar] [CrossRef]
- Kumar, S.; Zhao, Y.; Sun, L.; Negi, S.S.; Halpert, J.R.; Muralidhara, B.K. Rational engineering of human cytochrome P450 2B6 for enhanced expression and stability: Importance of a Leu264 -> Phe substitution. Mol. Pharmacol. 2007, 72, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Rocha, C.G.; Gamble, C.G.; Lonsdale, R.; Li, A.; Nett, N.; Hoebenreich, S.; Lingnau, J.B.; Wirtz, C.; Fares, C.; Hinrichs, H.; et al. P450-Catalyzed Regio- and Diastereoselective Steroid Hydroxylation: Efficient Directed Evolution Enabled by Mutability Landscaping. ACS Catal. 2018, 8, 3395–3410. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Peng, Y.; Zhao, J.; Li, Q.; Yu, X.; Acevedo-Rocha, C.G.; Li, A. Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 2020, 7, 2. [Google Scholar] [CrossRef]
- Braunegg, G.; de Raadt, A.; Feichtenhofer, S.; Griengl, H.; Kopper, I.I.; Lehmann, A.; Weber, H.J. The Concept of Docking/Protecting Groups in Biohydroxylation. Angew. Chem. 1999, 38, 2763–2766. [Google Scholar] [CrossRef]
- de Raadt, A.; Griengl, H.; Weber, H. The concept of docking and protecting groups in biohydroxylation. Chem. Eur. J. 2001, 7, 27–31. [Google Scholar] [CrossRef]
- Munzer, D.F.; Meinhold, P.; Peters, M.W.; Feichtenhofer, S.; Griengl, H.; Arnold, F.H.; Glieder, A.; de Raadt, A. Stereoselective hydroxylation of an achiral cyclopentanecarboxylic acid derivative using engineered P450s BM-3. Chem. Commun. 2005, 20, 2597–2599. [Google Scholar] [CrossRef] [Green Version]
- Landwehr, M.; Hochrein, L.; Otey, C.R.; Kasrayan, A.; Bäckvall, J.-E.; Arnold, F.H. Enantioselective α-Hydroxylation of 2-Arylacetic Acid Derivatives and Buspirone Catalyzed by Engineered Cytochrome P450 BM-3. J. Am. Chem. Soc. 2006, 128, 6058–6059. [Google Scholar] [CrossRef] [Green Version]
- Narayan, A.R.H.; Jimenez-Oses, G.; Liu, P.; Negretti, S.; Zhao, W.X.; Gilbert, M.M.; Ramabhadran, R.O.; Yang, Y.F.; Furan, L.R.; Li, Z.; et al. Enzymatic hydroxylation of an unactivated methylene C-H bond guided by molecular dynamics simulations. Nat. Chem. 2015, 7, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Chaulagain, M.R.; Knauff, A.R.; Podust, L.M.; Montgomery, J.; Sherman, D.H. Selective oxidation of carbolide C-H bonds by an engineered macrolide P450 mono-oxygenase. Proc. Natl. Acad. Sci. USA 2009, 106, 18463–18468. [Google Scholar] [CrossRef] [Green Version]
- Negretti, S.; Narayan, A.R.H.; Chiou, K.C.; Kells, P.M.; Stachowski, J.L.; Hansen, D.A.; Podust, L.M.; Montgomery, J.; Sherman, D.H. Directing Group-Controlled Regioselectivity in an Enzymatic C-H Bond Oxygenation. J. Am. Chem. Soc. 2014, 136, 4901–4904. [Google Scholar] [CrossRef]
- Larsen, A.T.; May, E.M.; Auclair, K. Predictable Stereoselective and Chemoselective Hydroxylations and Epoxidations with P450 3A4. J. Am. Chem. Soc. 2011, 133, 7853–7858. [Google Scholar] [CrossRef]
- Polic, V.; Cheong, K.J.; Hammerer, F.; Auclair, K. Regioselective Epoxidations by Cytochrome P450 3A4 Using a Theobromine Chemical Auxiliary to Predictably Produce N-Protected β- or γ-Amino Epoxides. Adv. Synth. Catal. 2017, 359, 3983–3989. [Google Scholar] [CrossRef]
- Polic, V.; Auclair, K. Controlling substrate specificity and product regio- and stereo-selectivities of P450 enzymes without mutagenesis. Bioorg. Med. Chem. 2014, 22, 5547–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, O.; Watanabe, Y. Bringing out the Potential of Wild-type Cytochrome P450s using Decoy Molecules: Oxygenation of Nonnative Substrates by Bacterial Cytochrome P450s. Isr. J. Chem. 2015, 55, 32–39. [Google Scholar] [CrossRef]
- Shoji, O.; Watanabe, Y. Monooxygenation of Nonnative Substrates Catalyzed by Bacterial Cytochrome P450s Facilitated by Decoy Molecules. Chem. Lett. 2017, 46, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Shoji, O.; Aiba, Y.; Watanabe, Y. Hoodwinking Cytochrome P450BM3 into Hydroxylating Non-Native Substrates by Exploiting Its Substrate Misrecognition. Acc. Chem. Res. 2019, 52, 925–934. [Google Scholar] [CrossRef]
- Hara, M.; Ohkawa, H.; Narato, M.; Shirai, M.; Asada, Y.; Karube, I.; Miyake, J. Regeneration of NADPH by cactus chloroplasts: Coupling reaction with P450 monooxygenase. J. Ferment. Bioeng. 1997, 84, 324–329. [Google Scholar] [CrossRef]
- Lee, S.H.; Kwon, Y.C.; Kim, D.M.; Park, C.B. Cytochrome P450-catalyzed O-dealkylation coupled with photochemical NADPH regeneration. Biotechnol. Bioeng. 2013, 110, 383–390. [Google Scholar] [CrossRef]
- Cirino, P.C.; Arnold, F.H. Regioselectivity and activity of cytochrome P450 BM-3 and mutant F87A in reactions driven by hydrogen peroxide. Adv. Synth. Catal. 2002, 344, 932–937. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Zhou, L.; Cong, Z. Progress in cytochrome P450 monooxygenase driven by hydrogen peroxide. Biot. Resour. 2017, 39, 75–84. [Google Scholar]
- Hiroya, K.; Murakami, Y.; Shimizu, T.; Hatano, M.; Demontellano, P.R.O. Differential roles of Glu318 and Thr319 in cytochrome P450 1A2 catalysis supported by NADPH-cytochrome P450 reductase and tert-butyl hydroperoxide. Arch. Biochem. Biophys. 1994, 310, 397–401. [Google Scholar] [CrossRef]
- Cho, K.B.; Moreau, Y.; Kumar, D.; Rock, D.A.; Jones, J.P.; Shaik, S. Formation of the active species of cytochrome p450 by using iodosylbenzene: A case for spin-selective reactivity. Chem. Eur. J. 2007, 13, 4103–4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunham, N.P.; Arnold, F.H. Nature’s Machinery, Repurposed: Expanding the Repertoire of Iron-Dependent Oxygenases. ACS Catal. 2020, 10, 12239–12255. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, C.; Cong, Z. Strategies for Substrate-Regulated P450 Catalysis: From Substrate Engineering to Co-catalysis. Chem. Eur. J. 2019, 25, 6853–6863. [Google Scholar] [CrossRef]
- Shoji, O.; Watanabe, Y. Peroxygenase reactions catalyzed by cytochromes P450. J. Biol. Inorg. Chem. 2014, 19, 529–539. [Google Scholar] [CrossRef]
- Zhang, L.B.; Wang, Q. Harnessing P450 Enzyme for Biotechnology and Synthetic Biology. Chembiochem 2022, 23, e202100439. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.B.; Girhard, M. Cytochrome P450 Monooxygenases in Biotechnology and Synthetic Biology. Trends Biotechnol. 2019, 37, 882–897. [Google Scholar] [CrossRef]
- Munro, A.W.; McLean, K.J.; Grant, J.L.; Makris, T.M. Structure and function of the cytochrome P450 peroxygenase enzymes. Biochem. Soc. Trans. 2018, 46, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Bornscheuer, U.T.; Pohl, M. Improved biocatalysts by directed evolution and rational protein design. Curr. Opin. Chem. Biol. 2001, 5, 137–143. [Google Scholar] [CrossRef]
- Wang, X.D.; Saba, T.; Yiu, H.H.P.; Howe, R.F.; Anderson, J.A.; Shi, J.F. Cofactor NAD(P)H Regeneration Inspired by Heterogeneous Pathways. Chem 2017, 2, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Yamada, A.; Sugimoto, H.; Matsunaga, I.; Ogura, H.; Ichihara, K.; Adachi, S.; Park, S.Y.; Shiro, Y. Substrate recognition and molecular mechanism of fatty acid hydroxylation by cytochrome P450 from Bacillus subtilis. Crystallographic, spectroscopic, and mutational studies. J. Biol. Chem. 2003, 278, 9761–9767. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, I.; Yokotani, N.; Gotoh, O.; Kusunose, E.; Yamada, M.; Ichihara, K. Molecular Cloning and Expression of Fatty Acid α-Hydroxylase from Sphingomonas paucimobilis. J. Biol. Chem. 1997, 272, 23592–23596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujishiro, T.; Shoji, O.; Nagano, S.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Crystal structure of H2O2-dependent cytochrome P450SPalpha with its bound fatty acid substrate: Insight into the regioselective hydroxylation of fatty acids at the alpha position. J. Biol. Chem. 2011, 286, 29941–29950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rude, M.A.; Baron, T.S.; Brubaker, S.; Alibhai, M.; Del Cardayre, S.B.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel p450 fatty acid decarboxylase from Jeotgalicoccus species. Appl. Environ. Microb. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markel, U.; Lanvers, P.; Sauer, D.F.; Wittwer, M.; Dhoke, G.V.; Davari, M.D.; Schiffels, J.; Schwaneberg, U. A Photoclick-Based High-Throughput Screening for the Directed Evolution of Decarboxylase OleT. Chem. Eur. J. 2021, 27, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Girhard, M.; Schuster, S.; Dietrich, M.; Duerre, P.; Urlacher, V.B. Cytochrome P450 monooxygenase from Clostridium acetobutylicum: A new alpha-fatty acid hydroxylase. Biochem. Biophys. Res. Commun. 2007, 362, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Hrycay, E.G.; Gustafsson, J.A.; Ingelmansundberg, M.; Ernster, L. Sodium periodate, sodium chlorite, organic hydroperoxides, and H2O2 as hydroxylating agents in steroid hydroxylation reactions catalyzed by partially purified cytochrome P-450. Biochem. Biophys. Res. Commun. 1975, 66, 209–216. [Google Scholar] [CrossRef]
- Anari, M.R.; Josephy, P.D.; Henry, T.; Obrien, P.J. Hydrogen peroxide supports human and rat cytochrome P450 1A2-catalyzed 2-amino-3-methylimidazo 4,5-f quinoline bioactivation to mutagenic metabolites: Significance of cytochrome P450 peroxygenase. Chem. Res. Toxicol. 1997, 10, 582–588. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Li, Y.; Stearns, R.A.; de Montellano, P.R.O.; Baillie, T.A.; Tang, W. Cytochrome p450 3A4-mediated oxidative conversion of a cyano to an amide group in the metabolism of pinacidil. Biochemistry 2002, 41, 2712–2718. [Google Scholar] [CrossRef]
- Rabe, K.S.; Kiko, K.; Niemeyer, C.M. Characterization of the peroxidase activity of CYP119, a thermostable P450 from Sulfolobus acidocaldarius. Chembiochem 2008, 9, 420–425. [Google Scholar] [CrossRef]
- Goyal, S.; Banerjee, S.; Mazumdar, S. Oxygenation of monoenoic fatty acids by CYP175A1, an orphan cytochrome P450 from Thermus thermophilus HB27. Biochemistry 2012, 51, 7880–7890. [Google Scholar] [CrossRef]
- Gelb, M.H.; Heimbrook, D.C.; Malkonen, P.; Sligar, S.G. Stereochemistry and deuterium isotope effects in camphor hydroxylation by the cytochrome P450cam monoxygenase system. Biochemistry 1982, 21, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Cirino, P.C.; Arnold, F.H. A Self-Sufficient Peroxide-Driven Hydroxylation Biocatalyst. Angew. Chem. Int. Ed. 2003, 42, 3299–3301. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Chen, C.S.; Waxman, D.J.; Halpert, J.R. Directed evolution of mammalian cytochrome P4502B1. J. Biol. Chem. 2005, 280, 19569–19575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, R.K.; Goyal, S.; Mazumdar, S. Modification of the heme active site to increase the peroxidase activity of thermophilic cytochrome P450: A rational approach. J. Inorg. Biochem. 2010, 104, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Lin, Z.L.; Arnold, F.H. Laboratory evolution of peroxide-mediated cytochrome P450 hydroxylation. Nature 1999, 399, 670–673. [Google Scholar] [CrossRef]
- Gajhede, M.; Schuller, D.J.; Henriksen, A.; Smith, A.T.; Poulos, T.L. Crystal structure of horseradish peroxidase C at 2.15 angstrom resolution. Nat. Struct. Biol. 1997, 4, 1032–1038. [Google Scholar] [CrossRef]
- Piontek, K.; Strittmatter, E.; Ullrich, R.; Grobe, G.; Pecyna, M.J.; Kluge, M.; Scheibner, K.; Hofrichter, M.; Plattner, D.A. Structural basis of substrate conversion in a new aromatic peroxygenase: Cytochrome P450 functionality with benefits. J. Biol. Chem. 2013, 288, 34767–34776. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, S.I.; Roach, M.P.; Matsui, T.; Watanabe, Y. Investigations of the roles of the distal heme environment and the proximal heme iron ligand in peroxide activation by heme enzymes via molecular engineering of myoglobin. Acc. Chem. Res. 2001, 34, 818–825. [Google Scholar] [CrossRef]
- Watanabe, Y.; Ueno, T. Introduction of P450, Peroxidase, and Catalase Activities into Myoglobin by Site-Directed Mutagenesis: Diverse Reactivities of Compound I. Bull. Chem. Soc. Jpn. 2003, 76, 1309–1322. [Google Scholar] [CrossRef]
- Adachi, S.; Nagano, S.; Ishimori, K.; Watanabe, Y.; Morishima, I.; Egawa, T.; Kitagawa, T.; Makino, R. Roles of proximal ligand in heme proteins: Replacement of proximal histidine of human myoglobin with cysteine and tyrosine by site-directed mutagenesis as models for P-450, chloroperoxidase, and catalase. Biochemistry 1993, 32, 241–252. [Google Scholar] [CrossRef]
- Shoji, O.; Fujishiro, T.; Nishio, K.; Kano, Y.; Kimoto, H.; Chien, S.-C.; Onoda, H.; Muramatsu, A.; Tanaka, S.; Hori, A.; et al. A substrate-binding-state mimic of H2O2-dependent cytochrome P450 produced by one-point mutagenesis and peroxygenation of non-native substrates. J. Inorg. Biochem. 2016, 6, 5806–5811. [Google Scholar] [CrossRef] [Green Version]
- Podgorski, M.N.; Harbort, J.S.; Lee, J.H.Z.; Nguyen, G.T.H.; Bruning, J.B.; Donald, W.A.; Bernhardt, P.V.; Harmer, J.R.; Bell, S.G. An Altered Heme Environment in an Engineered Cytochrome P450 Enzyme Enables the Switch from Monooxygenase to Peroxygenase Activity. ACS Catal. 2022, 12, 1614–1625. [Google Scholar] [CrossRef]
- Whitehouse, C.J.C.; Bell, S.G.; Wong, L.-L. P450BM3 (CYP102A1): Connecting the dots. Chem. Soc. Rev. 2012, 41, 1218–1260. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Chen, Z.; Chen, J.; Chen, J.; Wang, C.; Zhou, H.; Yao, L.; Shoji, O.; Watanabe, Y.; Cong, Z. Dual-Functional Small Molecules for Generating an Efficient Cytochrome P450BM3 Peroxygenase. Angew. Chem. Int. Ed. 2018, 57, 7628–7633. [Google Scholar] [CrossRef]
- Willot, S.J.P.; Tieves, F.; Girhard, M.; Urlacher, V.B.; Hollmann, F.; de Gonzalo, G. P450BM3-Catalyzed Oxidations Employing Dual Functional Small Molecules. Catalysts 2019, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lan, D.; Durrani, R.; Hollmann, F. Peroxygenases en route to becoming dream catalysts. What are the opportunities and challenges? Curr. Opin. Chem. Biol. 2017, 37, 1–9. [Google Scholar] [CrossRef]
- Olmedo, A.; Aranda, C.; del Río, J.C.; Kiebist, J.; Scheibner, K.; Martínez, A.T.; Gutiérrez, A. From Alkanes to Carboxylic Acids: Terminal Oxygenation by a Fungal Peroxygenase. Angew. Chem. Int. Ed. 2016, 55, 12248–12251. [Google Scholar] [CrossRef]
- Imai, M.; Shimada, H.; Watanabe, Y.; Matsushima-Hibiya, Y.; Makino, R.; Koga, H.; Horiuchi, T.; Ishimura, Y. Uncoupling of the cytochrome P-450cam monooxygenase reaction by a single mutation, threonine-252 to alanine or valine: Possible role of the hydroxy amino acid in oxygen activation. Proc. Natl. Acad. Sci. USA 1989, 86, 7823–7827. [Google Scholar] [CrossRef] [Green Version]
- Martinis, S.A.; Atkins, W.M.; Stayton, P.S.; Sligar, S.G. A Conserved Residue of Cytochrome-P-450 Is Involved in Heme-Oxygen Stability and Activation. J. Am. Chem. Soc. 1989, 111, 9252–9253. [Google Scholar] [CrossRef]
- Truan, G.; Peterson, J.A. Thr268 in Substrate Binding and Catalysis in P450BM-31. Arch. Biochem. Biophy. 1998, 349, 53–64. [Google Scholar] [CrossRef]
- Rittle, J.; Green, M.T. Cytochrome P450 compound I: Capture, characterization, and C-H bond activation kinetics. Science 2010, 330, 933–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Jiang, Y.; Chen, Q.; Dong, S.; Feng, Y.; Cong, Z.; Shaik, S.; Wang, B. H-Bonding Networks Dictate the Molecular Mechanism of H2O2 Activation by P450. ACS Catal. 2021, 11, 8774–8785. [Google Scholar] [CrossRef]
- Haines, D.C.; Tomchick, D.R.; Machius, M.; Peterson, J.A. Pivotal role of water in the mechanism of P450BM-3. Biochemistry 2001, 40, 13456–13465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, Z.; Shoji, O.; Kasai, C.; Kawakami, N.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Activation of Wild-Type Cytochrome P450BM3 by the Next Generation of Decoy Molecules: Enhanced Hydroxylation of Gaseous Alkanes and Crystallographic Evidence. ACS Catal. 2014, 5, 150–156. [Google Scholar] [CrossRef]
- Shoji, O.; Yanagisawa, S.; Stanfield, J.K.; Suzuki, K.; Cong, Z.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Direct Hydroxylation of Benzene to Phenol by Cytochrome P450BM3 Triggered by Amino Acid Derivatives. Angew. Chem. Int. Ed. 2017, 56, 10324–10329. [Google Scholar] [CrossRef]
- Wang, B.; Li, C.; Dubey, K.D.; Shaik, S. Quantum mechanical/molecular mechanical calculated reactivity networks reveal how cytochrome P450cam and Its T252A mutant select their oxidation pathways. J. Am. Chem. Soc. 2015, 137, 7379–7390. [Google Scholar] [CrossRef]
- Ramirez-Escudero, M.; Molina-Espeja, P.; Gomez de Santos, P.; Hofrichter, M.; Sanz-Aparicio, J.; Alcalde, M. Structural Insights into the Substrate Promiscuity of a Laboratory-Evolved Peroxygenase. ACS Chem. Biol. 2018, 13, 3259–3268. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Micro. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef]
- Sigmund, M.-C.; Poelarends, G.J. Current state and future perspectives of engineered and artificial peroxygenases for the oxyfunctionalization of organic molecules. Nat. Catal. 2020, 3, 690–702. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R. Oxidations catalyzed by fungal peroxygenases. Curr. Opin. Chem. Biol. 2014, 19, 116–125. [Google Scholar] [CrossRef]
- Bormann, S.; Gomez Baraibar, A.; Ni, Y.; Holtmann, D.; Hollmann, F. Specific oxyfunctionalisations catalysed by peroxygenases: Opportunities, challenges and solutions. Catal. Sci. Technol. 2015, 5, 2038–2052. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, R.; Poraj-Kobielska, M.; Scholze, S.; Halbout, C.; Sandvoss, M.; Pecyna, M.J.; Scheibner, K.; Hofrichter, M. Side chain removal from corticosteroids by unspecific peroxygenase. J. Inorg. Biochem. 2018, 183, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Molina-Espeja, P.; Canellas, M.; Plou, F.J.; Hofrichter, M.; Lucas, F.; Guallar, V.; Alcalde, M. Synthesis of 1-Naphthol by a Natural Peroxygenase Engineered by Directed Evolution. ChemBioChem 2016, 17, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Gomez de Santos, P.; Cañellas, M.; Tieves, F.; Younes, S.H.H.; Molina-Espeja, P.; Hofrichter, M.; Hollmann, F.; Guallar, V.; Alcalde, M. Selective Synthesis of the Human Drug Metabolite 5′-Hydroxypropranolol by an Evolved Self-Sufficient Peroxygenase. ACS Catal. 2018, 8, 4789–4799. [Google Scholar] [CrossRef] [Green Version]
- Hagstrom, A.E.; Tornvall, U.; Nordblad, M.; Hatti-Kaul, R.; Woodley, J.M. Chemo-enzymatic epoxidation-process options for improving biocatalytic productivity. Biotechnol. Prog. 2011, 27, 67–76. [Google Scholar] [CrossRef]
- Wunschik, D.S.; Ingenbosch, K.N.; Suss, P.; Liebelt, U.; Quint, S.; Dyllick-Brenzinger, M.; Zuhse, R.; Menyes, U.; Hoffmann-Jacobsen, K.; Opwis, K.; et al. Enzymatic epoxidation of cyclohexene by peroxidase immobilization on a textile and an adapted reactor design. Enzyme. Microb. Technol. 2020, 136, 109512. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Fernandez-Fueyo, E.; Gomez Baraibar, A.; Ullrich, R.; Hofrichter, M.; Yanase, H.; Alcalde, M.; van Berkel, W.J.; Hollmann, F. Peroxygenase-Catalyzed Oxyfunctionalization Reactions Promoted by the Complete Oxidation of Methanol. Angew. Chem. Int. Ed. 2016, 55, 798–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.S.; Lee, H.; Tieves, F.; Lee, Y.W.; Son, E.J.; Zhang, W.; Shin, B.; Hollmann, F.; Park, C.B. Bias-Free in situ H2O2 Generation in a Photovoltaic-Photoelectrochemical Tandem Cell for Biocatalytic Oxyfunctionalization. ACS Catal. 2019, 9, 10562–10566. [Google Scholar] [CrossRef]
- Yayci, A.; Baraibar, A.G.; Krewing, M.; Fueyo, E.F.; Hollmann, F.; Alcalde, M.; Kourist, R.; Bandow, J.E. Plasma-Driven in situ Production of Hydrogen Peroxide for Biocatalysis. ChemSusChem 2020, 13, 2072–2079. [Google Scholar] [CrossRef]
- Zhao, P.; Chen, J.; Ma, N.; Chen, J.; Qin, X.; Liu, C.; Yao, F.; Yao, L.; Jin, L.; Cong, Z. Enabling highly (R)-enantioselective epoxidation of styrene by engineering unique non-natural P450 peroxygenases. Chem. Sci. 2021, 12, 6307–6314. [Google Scholar] [CrossRef]
- Chen, J.; Kong, F.; Ma, N.; Zhao, P.; Liu, C.; Wang, X.; Cong, Z. Peroxide-Driven Hydroxylation of Small Alkanes Catalyzed by an Artificial P450BM3 Peroxygenase System. ACS Catal. 2019, 9, 7350–7355. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, C.; Ma, N.; Chen, J.; Liu, C.; Wang, F.; Xu, J.; Cong, Z. Regioselective aromatic O-demethylation with an artificial P450BM3 peroxygenase system. Catal. Sci. Technol. 2020, 10, 1219–1223. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, J.; Ma, N.; Zhou, H.; Cong, Z. Selective hydroxylation of naphthalene using the H2O2-dependent engineered P450BM3 driven by dual-functional small molecules. J. Porphyr. Phthalocyanines. 2018, 22, 831–836. [Google Scholar] [CrossRef]
- Xu, G.; Mahmood, Q.; Lv, C.; Yang, R.; Zhou, L.; Wang, Q. Asymmetric kinetic resolution polymerization. Coord. Chem. Rev. 2020, 414, 213296. [Google Scholar] [CrossRef]
- Ren, W.-M.; Yue, T.-J.; Zhang, X.; Gu, G.-G.; Liu, Y.; Lu, X.-B. Stereoregular CO2 Copolymers from Epoxides with an Electron-Withdrawing Group: Crystallization and Unexpected Stereocomplexation. Macromolecules 2017, 50, 7062–7069. [Google Scholar] [CrossRef]
- Wu, G.-P.; Wei, S.-H.; Ren, W.-M.; Lu, X.-B.; Li, B.; Zu, Y.-P.; Darensbourg, D.J. Alternating copolymerization of CO2 and styrene oxide with Co(iii)-based catalyst systems: Differences between styrene oxide and propylene oxide. Energy Environ. Sci. 2011, 4, 5084–5092. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Li, Z. Biocatalytic selective functionalisation of alkenes via single-step and one-pot multi-step reactions. Chem. Commun. 2019, 55, 883–896. [Google Scholar] [CrossRef]
- Matsumoto, K.; Oguma, T.; Katsuki, T. Highly enantioselective epoxidation of styrenes catalyzed by proline-derived C1-symmetric titanium(salan) complexes. Angew. Chem. Int. Ed. 2009, 48, 7432–7435. [Google Scholar] [CrossRef]
- Wang, C.; Yamamoto, H. Asymmetric Epoxidation Using Hydrogen Peroxide as Oxidant. Chem.–Asian J. 2015, 10, 2056–2068. [Google Scholar] [CrossRef]
- Lin, H.; Xu, M.-Y.; Liu, Y.; Wu, Z.-L. Biocatalytic Epoxidation for Green Synthesis. In Green Biocatalysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 351–372. [Google Scholar]
- Wang, L.; Wei, S.; Pan, X.; Liu, P.; Du, X.; Zhang, C.; Pu, L.; Wang, Q. Enhanced Turnover for the P450 119 Peroxygenase-Catalyzed Asymmetric Epoxidation of Styrenes by Random Mutagenesis. Chem. Eur. J. 2018, 24, 2741–2749. [Google Scholar] [CrossRef]
- Li, A.; Liu, J.; Pham, S.Q.; Li, Z. Engineered P450pyr monooxygenase for asymmetric epoxidation of alkenes with unique and high enantioselectivity. Chem. Commun. 2013, 49, 11572–11574. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Wang, J.B.; Reetz, M.T. Exploiting Designed Oxidase-Peroxygenase Mutual Benefit System for Asymmetric Cascade Reactions. J. Am. Chem. Soc. 2019, 141, 5655–5658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, T.; Scholtissek, A.; Hofmann, S.; Koch, R.; Tischler, D. Accessing Enantiopure Epoxides and Sulfoxides: Related Flavin-Dependent Monooxygenases Provide Reversed Enantioselectivity. ChemCatChem 2020, 12, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Tang, Y.; Dong, S.; Lang, R.; Chen, H. A new monooxygenase from Herbaspirillum huttiense catalyzed highly enantioselective epoxidation of allylbenzenes and allylic alcohols. Catal. Sci. Technol. 2020, 10, 2145–2151. [Google Scholar] [CrossRef]
- van Schie, M.; Paul, C.E.; Arends, I.; Hollmann, F. Photoenzymatic epoxidation of styrenes. Chem. Commun. 2019, 55, 1790–1792. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Guo, C.; Lin, H.; Ding, Z.Y.; Wu, Z.L. Functional characterization of an (R)-selective styrene monooxygenase from streptomyces sp. NRRL S-31. Enzym. Microb. Technol. 2019, 132, 109391. [Google Scholar] [CrossRef]
- Xiao, H.; Dong, S.; Liu, Y.; Pei, X.-Q.; Lin, H.; Wu, Z.-L. A new clade of styrene monooxygenases for (R)-selective epoxidation. Catal. Sci. Technol. 2021, 11, 2195–2201. [Google Scholar] [CrossRef]
- Nesterov, D.S.; Nesterova, O.V.; Pombeiro, A.J.L. Homo- and heterometallic polynuclear transition metal catalysts for alkane CH bonds oxidative functionalization: Recent advances. Coord. Chem. Rev. 2018, 355, 199–222. [Google Scholar] [CrossRef]
- Soussan, L.; Pen, N.; Belleville, M.-P.; Marcano, J.S.; Paolucci-Jeanjean, D. Alkane biohydroxylation: Interests, constraints and future developments. J. Biotechnol. 2016, 222, 117–142. [Google Scholar] [CrossRef]
- Banerj Ee, R.; Proshlyakov, Y.; Lipscomb, J.D.; Proshlyakov, D.A. Structure of the key species in the enzymatic oxidation of methane to methanol. Nature 2015, 518, 431. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.R. Handbook of Bond Dissociation Energies in Organic Compounds; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Cooley, R.B.; Dubbel, B.L.; Sayavedra-Soto, L.A.; Bottomley, P.J.; Arp, D.J. Kinetic characterization of the soluble butane monooxygenase from Thauera butanivorans, formerly ‘Pseudomonas butanovora’. Microbiology 2009, 155, 2086–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, S.; Kinne, M.; Wang, X.S.; Ullrich, R.; Kayser, G.; Groves, J.T.; Hofrichter, M. Selective hydroxylation of alkanes by an extracellular fungal peroxygenase. FEBS J. 2011, 278, 3667–3675. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, N.; Shoji, O.; Watanabe, Y. Use of Perfluorocarboxylic Acids to Trick Cytochrome P450BM3 into Initiating the Hydroxylation of Gaseous Alkanes. Angew. Chem. Int. Ed. 2011, 50, 5315–5318. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, K.; Ariyasu, S.; Stanfield, J.K.; Suzuki, K.; Onoda, H.; Kasai, C.; Sugimoto, H.; Aiba, Y.; Watanabe, Y.; Shoji, O. Systematic Evolution of Decoy Molecules for the Highly Efficient Hydroxylation of Benzene and Small Alkanes Catalyzed by Wild-Type Cytochrome P450BM3. ACS Catal. 2020, 10, 9136–9144. [Google Scholar] [CrossRef]
- Ariyasu, S.; Kodama, Y.; Kasai, C.; Cong, Z.; Stanfield, J.K.; Aiba, Y.; Watanabe, Y.; Shoji, O. Development of a High-Pressure Reactor Based on Liquid-Flow Pressurisation to Facilitate Enzymatic Hydroxylation of Gaseous Alkanes. ChemCatChem 2019, 11, 4709–4714. [Google Scholar] [CrossRef]
- Zilly, F.E.; Acevedo, J.P.; Augustyniak, W.; Deege, A.; Haeusig, U.W.; Reetz, M.T. Tuning a P450 Enzyme for Methane Oxidation. Angew. Chem. Int. Ed. 2011, 50, 2720–2724. [Google Scholar] [CrossRef]
- Fasan, R.; Chen, M.M.; Crook, N.C.; Arnold, F.H. Engineered alkane-hydroxylating cytochrome P450(BM3) exhibiting nativelike catalytic properties. Angew. Chem. Int. Ed. 2007, 46, 8414–8418. [Google Scholar] [CrossRef]
- Patel, R.N.; Hou, C.T.; Laskin, A.I.; Felix, A. Microbial Oxidation of Hydrocarbons: Properties of a Soluble Methane Monooxygenase from a Facultative Methane-Utilizing Organism, Methylobacterium sp. Strain CRL-26. Appl. Environ. Microbiol. 1982, 44, 1130–1137. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Bell, S.G.; Lednik, J.; Insley, A.; Rao, Z.H.; Wong, L.L. The heme monooxygenase cytochrome P450(cam) can be engineered to oxidize ethane to ethanol. Angew. Chem. Int. Ed. 2005, 44, 4029–4032. [Google Scholar] [CrossRef]
- Trippe, K.M.; Wolpert, T.J.; Hyman, M.R.; Ciuffetti, L.M. RNAi silencing of a cytochrome P450 monoxygenase disrupts the ability of a filamentous fungus, Graphium sp., to grow on short-chain gaseous alkanes and ethers. Biodegradation 2014, 25, 137–151. [Google Scholar] [CrossRef]
- Gazi, S. Valorization of wood biomass-lignin via selective bond scission: A minireview. Appl. Catal. B Environ. 2019, 257, 117936. [Google Scholar] [CrossRef]
- Kamimura, N.; Takahashi, K.; Mori, K.; Araki, T.; Fujita, M.; Higuchi, Y.; Masai, E. Bacterial catabolism of lignin-derived aromatics: New findings in a recent decade: Update on bacterial lignin catabolism. Environ. Microbiol. Rep. 2017, 9, 679–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.C.; Paice, M.; Zhang, X. Enzymatic Oxidation of Lignin: Challenges and Barriers toward Practical Applications. ChemCatChem 2020, 12, 401–425. [Google Scholar] [CrossRef]
- Venkatesagowda, B. Enzymatic demethylation of lignin for potential biobased polymer applications. Fungal Biol. Rev. 2019, 33, 190–224. [Google Scholar] [CrossRef]
- Richter, N.; Zepeck, F.; Kroutil, W. Cobalamin-dependent enzymatic O-, N-, and S-demethylation. Trends Biotechnol. 2015, 33, 371–373. [Google Scholar] [CrossRef]
- Lanfranchi, E.; Trajkovic, M.; Barta, K.; de Vries, J.G.; Janssen, D.B. Exploring the Selective Demethylation of Aryl Methyl Ethers with a Pseudomonas Rieske Monooxygenase. ChemBioChem 2019, 20, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Mallinson, S.J.B.; Machovina, M.M.; Silveira, R.L.; Garcia-Borras, M.; Gallup, N.; Johnson, C.W.; Allen, M.D.; Skaf, M.S.; Crowley, M.F.; Neidle, E.L.; et al. A promiscuous cytochrome P450 aromatic O-demethylase for lignin bioconversion. Nat. Commun. 2018, 9, 2487. [Google Scholar] [CrossRef] [Green Version]
- Machovina, M.M.; Mallinson, S.J.B.; Knott, B.C.; Meyers, A.W.; Garcia-Borras, M.; Bu, L.; Gado, J.E.; Oliver, A.; Schmidt, G.P.; Hinchen, D.J.; et al. Enabling microbial syringol conversion through structure-guided protein engineering. Proc. Natl. Acad. Sci. USA 2019, 16, 13970–13976. [Google Scholar] [CrossRef] [Green Version]
- Ellis, E.S.; Hinchen, D.J.; Bleem, A.; Bu, L.; Mallinson, S.J.B.; Allen, M.D.; Streit, B.R.; Machovina, M.M.; Doolin, Q.V.; Michener, W.E.; et al. Engineering a Cytochrome P450 for Demethylation of Lignin-Derived Aromatic Aldehydes. JACS Au 2021, 1, 252–261. [Google Scholar] [CrossRef]
- Dardas, A.; Gal, D.; Barrelle, M.; Sauretignazi, G.; Sterjiades, R.; Pelmont, J. The demethylation of guaiacol by a new bacterial cytochrome P-450. Arch. Biochem. Biophys. 1985, 236, 585–592. [Google Scholar] [CrossRef]
- Eltis, L.D.; Karlson, U.; Timmis, K.N. Purification and characterization of cytochrome P450RR1 from Rhodococcus rhodochrous. Eur. J. Biochem. 1993, 213, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.G.; Zhou, R.; Yang, W.; Tan, A.B.H.; Gentleman, A.S.; Wong, L.-L.; Zhou, W. Investigation of the Substrate Range of CYP199A4: Modification of the Partition between Hydroxylation and Desaturation Activities by Substrate and Protein Engineering. Chem. Eur. J. 2012, 18, 16677–16688. [Google Scholar] [CrossRef] [PubMed]
- Leveson-Gower, R.B.; Mayer, C.; Roelfes, G. The importance of catalytic promiscuity for enzyme design and evolution. Nat. Rev. Chem. 2019, 3, 687–705. [Google Scholar] [CrossRef]
- Sandoval, B.A.; Hyster, T.K. Emerging strategies for expanding the toolbox of enzymes in biocatalysis. Curr. Opin. Chem. Biol. 2020, 55, 45–51. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Kazlauskas, R.J. Catalytic promiscuity in biocatalysis: Using old enzymes to form new bonds and follow new pathways. Angew. Chem. Int. Ed. 2004, 43, 6032–6040. [Google Scholar] [CrossRef]
- Hrycay, E.G.; Bandiera, S.M. The monooxygenase, peroxidase, and peroxygenase properties of cytochrome P450. Arch. Biochem. Biophys. 2012, 522, 71–89. [Google Scholar] [CrossRef]
- Depillis, G.D.; Sishta, B.P.; Mauk, A.G.; Demontellano, P.R.O. Small substrates and cytochrome c are oxidized at different sites of cytochrome c peroxidase. J. Biol. Chem. 1991, 266, 19334–19341. [Google Scholar] [CrossRef]
- Zhang, P.; Yuan, H.; Xu, J.K.; Wang, X.J.; Gao, S.Q.; Tan, X.S.; Lin, Y.W. A Catalytic Binding Site Together with a Distal Tyr in Myoglobin Affords Catalytic Efficiencies Similar to Natural Peroxidases. ACS Catal. 2020, 10, 891–896. [Google Scholar] [CrossRef]
- Yamada, M.; Hashimoto, Y.; Kumano, T.; Tsujimura, S.; Kobayashi, M. New function of aldoxime dehydratase: Redox catalysis and the formation of an expected product. PLoS ONE 2017, 12, e0175846. [Google Scholar] [CrossRef] [Green Version]
- Chien, S.C.; Shoji, O.; Morimoto, Y.; Watanabe, Y. Use of apomyoglobin to gently remove heme from a H2O2-dependent cytochrome P450 and allow its reconstitution. New J. Chem. 2017, 41, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.N.; Fang, W.H.; Liu, C.A.F.; Qin, X.Q.; Wang, X.L.; Jin, L.Y.; Wang, B.J.; Cong, Z.Q. Switching an Artificial P450 Peroxygenase into Peroxidase via Mechanism-Guided Protein Engineering. ACS Catal. 2021, 11, 8449–8455. [Google Scholar] [CrossRef]
- Xu, J.; Cen, Y.X.; Singh, W.; Fan, J.J.; Wu, L.; Lin, X.F.; Zhou, J.H.; Huang, M.L.; Reetz, M.T.; Wu, Q. Stereodivergent Protein Engineering of a Lipase To Access All Possible Stereoisomers of Chiral Esters with Two Stereocenters. J. Am. Chem. Soc. 2019, 141, 7934–7945. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Wu, Q.; Reetz, M.T. Focused rational iterative site-specific mutagenesis (FRISM). In Enzyme Engineering and Evolution: General Methods; Methods in Enzymology; Tawfik, D.S., Ed.; Academic Press: New York, NY, USA, 2020; Volume 643, pp. 225–242. [Google Scholar]
- Vidal-Limon, A.; Aguila, S.; Ayala, M.; Batista, C.V.; Vazquez-Duhalt, R. Peroxidase activity stabilization of cytochrome P450BM3 by rational analysis of intramolecular electron transfer. J. Inorg. Biochem. 2013, 122, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Mate, M.J.; Romero, A.; Martinez, M.J.; Martinez, A.T.; Ruiz-Duenas, F.J. Two oxidation sites for low redox potential substrates: A directed mutagenesis, kinetic, and crystallographic study on Pleurotus eryngii versatile peroxidase. J. Biol. Chem. 2012, 287, 41053–410675. [Google Scholar] [CrossRef] [Green Version]
- Mandelman, D.; Jamal, J.; Poulos, T.L. Identification of two electron-transfer sites in ascorbate peroxidase using chemical modification, enzyme kinetics, and crystallography. Biochemistry 1998, 37, 17610–17617. [Google Scholar] [CrossRef]
- Murphy, E.J.; Metcalfe, C.L.; Nnamchi, C.; Moody, P.C.E.; Raven, E.L. Crystal structure of guaiacol and phenol bound to a heme peroxidase. FEBS J. 2012, 279, 1632–1639. [Google Scholar] [CrossRef]
- Li, Q.S.; Ogawa, J.; Shimizu, S. Critical role of the residue size at position 87 in H2O2-dependent substrate hydroxylation activity and H2O2 inactivation of cytochrome P450BM-3. Biochem. Biophys. Res. Commun. 2001, 280, 1258–1261. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Makris, T.M. Expanding the substrate scope and reactivity of cytochrome P450 OleT. Biochem. Biophys. Res. Commun. 2016, 476, 462–466. [Google Scholar] [CrossRef]
- Roberts, A.G.; Katayama, J.; Kaspera, R.; Ledwitch, K.V.; Trong, I.L.; Stenkamp, R.E.; Thompson, J.A.; Totah, R.A. The role of cytochrome P450 BM3 phenylalanine-87 and threonine-268 in binding organic hydroperoxides. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2016, 1860, 669–677. [Google Scholar] [CrossRef]
Figure 1.
The catalytic cycle of cytochrome P450 monooxygenase and the peroxide-shunt pathway.

Figure 2.
Active site structures of HRP ((A), PDB ID: 1ATJ) [58] and unspecific peroxygenase ((B), PDB ID: 2YOR) [59].

Figure 3.
Proof-of-concept of the dual-functional small molecule (DFSM)-facilitated P450 peroxygenase. (A) The NADPH-dependent P450BM3 monooxygenase. (B) Proposed catalytic cycle of the DFSM-facilitated P450 peroxygenase. (C) Chemical structures of the DFSM molecules. (D) Styrene epoxidation in the presence of the DFSM and control experiments.
Figure 3.
Proof-of-concept of the dual-functional small molecule (DFSM)-facilitated P450 peroxygenase. (A) The NADPH-dependent P450BM3 monooxygenase. (B) Proposed catalytic cycle of the DFSM-facilitated P450 peroxygenase. (C) Chemical structures of the DFSM molecules. (D) Styrene epoxidation in the presence of the DFSM and control experiments.

Figure 4.
Structural basis of the DFSM-facilitated P450 peroxygenase. (A) Proposed pre-reaction state of P450BM3 in the presence of H2O2 and DFSM. (B) The model structure with NH2OH instead of H2O2. (C) The co-crystal structure of P450BM3_F87A in complex with NH2OH and Im-C6-Phe. (D) The binding interactions of Im-C6-Phe with P450BM3.
Figure 4.
Structural basis of the DFSM-facilitated P450 peroxygenase. (A) Proposed pre-reaction state of P450BM3 in the presence of H2O2 and DFSM. (B) The model structure with NH2OH instead of H2O2. (C) The co-crystal structure of P450BM3_F87A in complex with NH2OH and Im-C6-Phe. (D) The binding interactions of Im-C6-Phe with P450BM3.

Figure 5.
Protein engineering of the DFSM-facilitated P450BM3 peroxygenase for catalyzing (R)-enantioselective epoxidation of styrene and its derivatives. (A) Key residues around the substrate-binding pocket of P450BM3; (B) protein engineering for styrene epoxidation; (C) the epoxidation of styrene derivatives by the DFSM-facilitated P450 peroxygenases.
Figure 5.
Protein engineering of the DFSM-facilitated P450BM3 peroxygenase for catalyzing (R)-enantioselective epoxidation of styrene and its derivatives. (A) Key residues around the substrate-binding pocket of P450BM3; (B) protein engineering for styrene epoxidation; (C) the epoxidation of styrene derivatives by the DFSM-facilitated P450 peroxygenases.

Figure 6.
DFSM-facilitated P450 peroxygenases catalyzed sulfoxidation and hydroxylation.

Figure 7.
Native monooxygenase activity. DFSM-facilitated peroxygenase activity and switching to peroxidase activity by mechanism-guided protein engineering.
Figure 7.
Native monooxygenase activity. DFSM-facilitated peroxygenase activity and switching to peroxidase activity by mechanism-guided protein engineering.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Catalytic hydroxylation of small alkanes by various enzymes in literature.
| Entry | Enzyme | Alkanes | Final Product | PFR a | TTN b | Ref. |
|---|---|---|---|---|---|---|
| 1 | BM3_F87A/T268I/A184I/Im-C6-Phe | Propane | 2-Propanol | 630 | 1775 | [93] |
| 2 | BM3_F87A/T268I/A82T/Im-C6-Phe | n-Butane | 2-Butanol | 1042 | 2253 | [93] |
| 3 | BM3/PFC10 | Propane | 2-Propanol | 70 | 700 | [117] |
| 4 | BM3/PFC9-L-Leu | Propane | 2-Propanol | 256 | 2560 | [76] |
| 5 | BM3/3CCPA-Pip-Phe | Propane | 2-Propanol | 615 | - | [118] |
| 6 | BM3/PFC9 | n-Butane | 2-Butanol | 110 | 1100 | [120] |
| 7 | BM3/PFC11 | Propane | 2-Propanol | - | 1021 | [120] |
| 8 | BM3/PFC7 | n-Butane | 2-Butanol | - | 3632 | [120] |
| 9 | BM3/C7AM-Pip-Phe | Ethane | Ethanol | 82.7 | - | [118] |
| 10 | P450cam_EB | n-Butane | 2-Butanol | 520 | - | [123] |
| 11 | P450cam_EB_L294M/T185M/L1358P/G248A | Propane | 2-Propanol | 505 | - | [123] |
| 12 | P450cam_EB_L294M/T185M/L1358P/G248A | Ethane | Ethanol | 78.2 | - | [123] |
| 13 | P450PMOR1 | Propane | 2-Propanol | 455 | 35,600 | [121] |
| 14 | P450PMOR2 | Propane | 2-Propanol | 370 | 45,800 | [121] |
| 15 c | CYP52L1 | Propane | 1-Propanol | - | - | [124] |
| 16 | sMMO | Methane | Methanol | 78 | - | [122] |
| 17 | sMMO | Ethane | Ethanol | 45.6 | - | [122] |
| 18 | sMMO | Propane | 2-Propanol | 33–58.8 | - | [122] |
| 19 | sMMO | n-Butane | 2-Butanol | 7.2–28.8 | - | [122] |
| 20 | AaeUPO | Propane | 2-Propanol | 17 | 999 | [116] |
| 21 | AaeUPO | n-Butane | 2-Butanol | 21 | 1258 | [116] |
a PFR: product formation rate in µmol·min−1·(µmol P450)−1. b TTN: total turnover number. c There is no catalytic turnover data reported.
Table 2.
Regioselective aromatic O-dealkylation by the DFSM-facilitated P450 peroxygenases.
 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Substrate | Enzyme System | Product | TON | |||||
| No. | R1 | R2 | R3 | No. | R2 | R3 | ||
| 3a | Me | H | H | BM3_F87A/T268I/Im-C5-Phe | 4a | H | H | 486 |
| 3b | Me | Me | H | BM3_F87A/T268I/Im-C6-Phe | 4b | Me | H | 356 |
| 3c | Me | OH | H | BM3_F87A/Im-C6-Phe | 4c | OH | H | 539 |
| 3d | Et | OH | H | BM3_F87G/T268G/Im-C5-Phe | 4d | OH | H | 99 |
| 3e | Me | OMe | H | BM3_F87A/T268I/Im-C6-Phe | 4e | OMe | H | 287 |
| 3f | H | OMe | OMe | BM3_F87G/T268V/Im-C5-Phe | 4f | OH | OMe | 165 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Di, S.; Fan, S.; Jiang, F.; Cong, Z. A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective. Antioxidants 2022, 11, 529. https://doi.org/10.3390/antiox11030529
AMA Style
Di S, Fan S, Jiang F, Cong Z. A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective. Antioxidants. 2022; 11(3):529. https://doi.org/10.3390/antiox11030529
Chicago/Turabian StyleDi, Siyu, Shengxian Fan, Fengjie Jiang, and Zhiqi Cong. 2022. "A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective" Antioxidants 11, no. 3: 529. https://doi.org/10.3390/antiox11030529
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.