
Temperature-Related Effects of Myocardial Protection Strategies in Swine Hearts after Prolonged Warm Ischemia

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setting, Tissue Biopsies and Experimental Groups
2.2. Tissue Samples Preparation
2.3. Estimation of the Total Amount of Proteins
2.4. Tissue Cytokines Analysis
2.5. Lipid Peroxidation Assay
2.6. Catalase Activity Assay
2.7. Selenium-Glutathione Peroxidase Activity
2.8. SOD Activity Assay
2.9. Transmission Electron Microscopy
2.10. Mitochondrial Analysis (%)
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
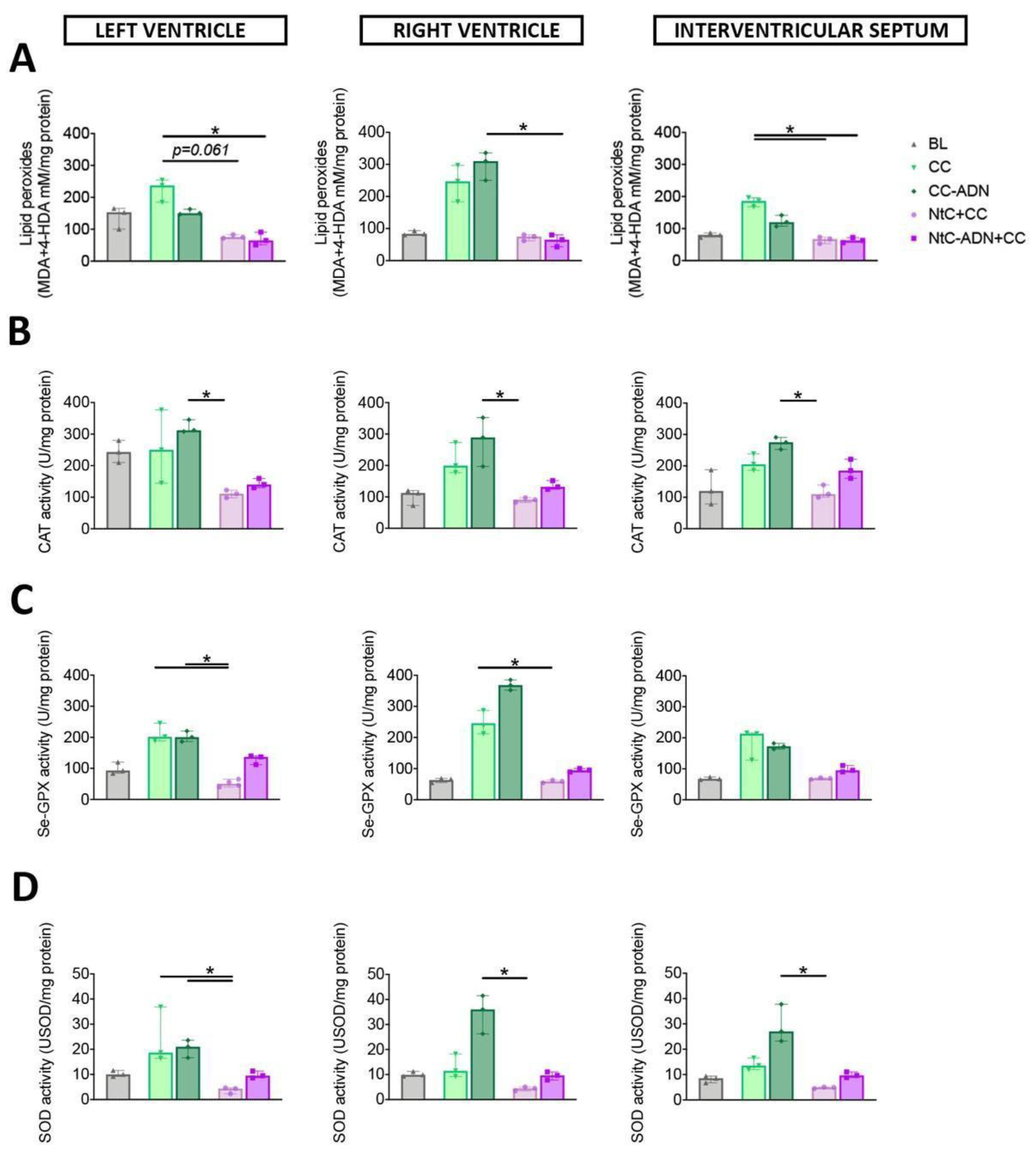
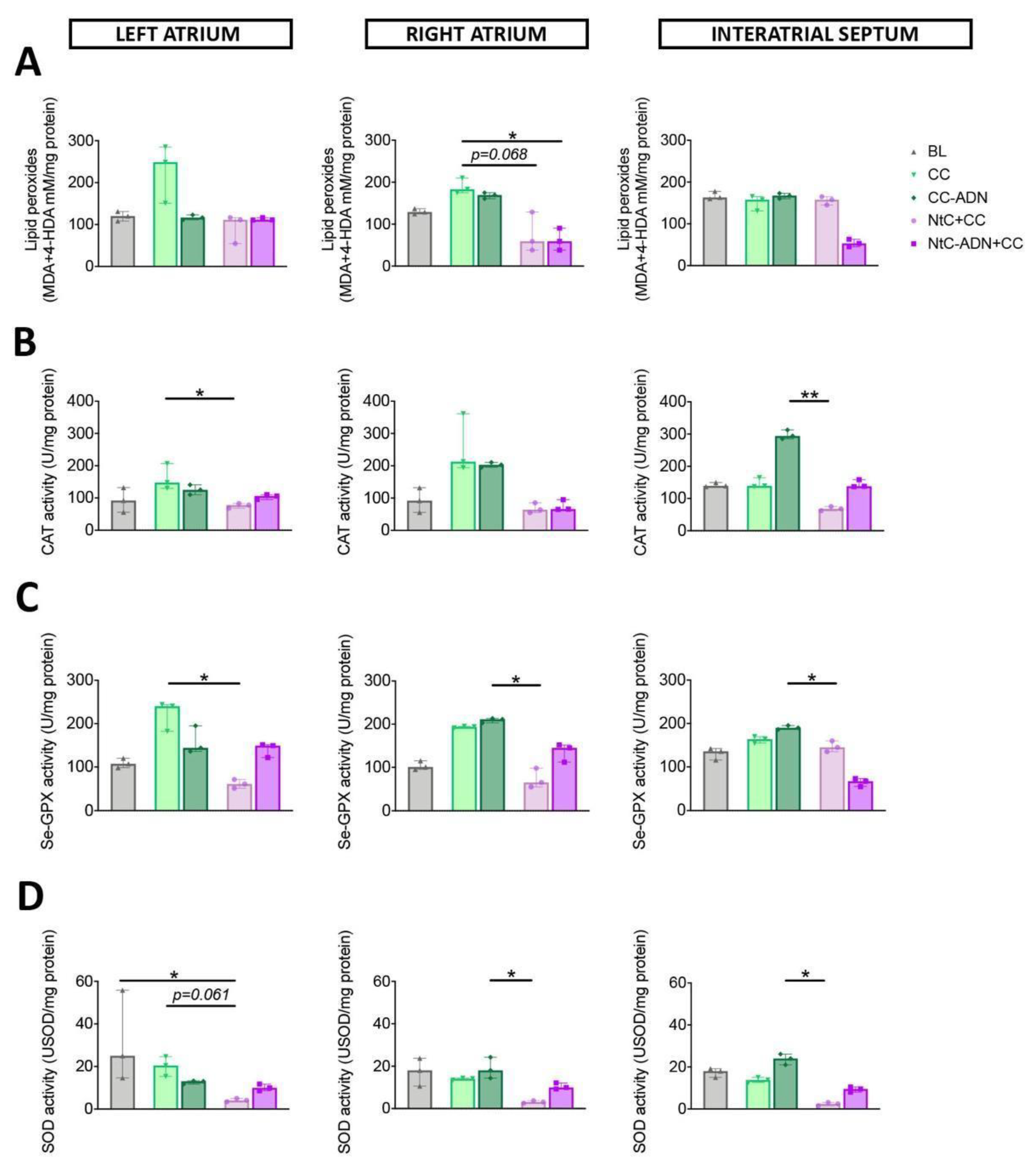
3.1. Combined Normothermic and Cold Cardioplegia Improves Myocardial Protection against Oxidative Stress
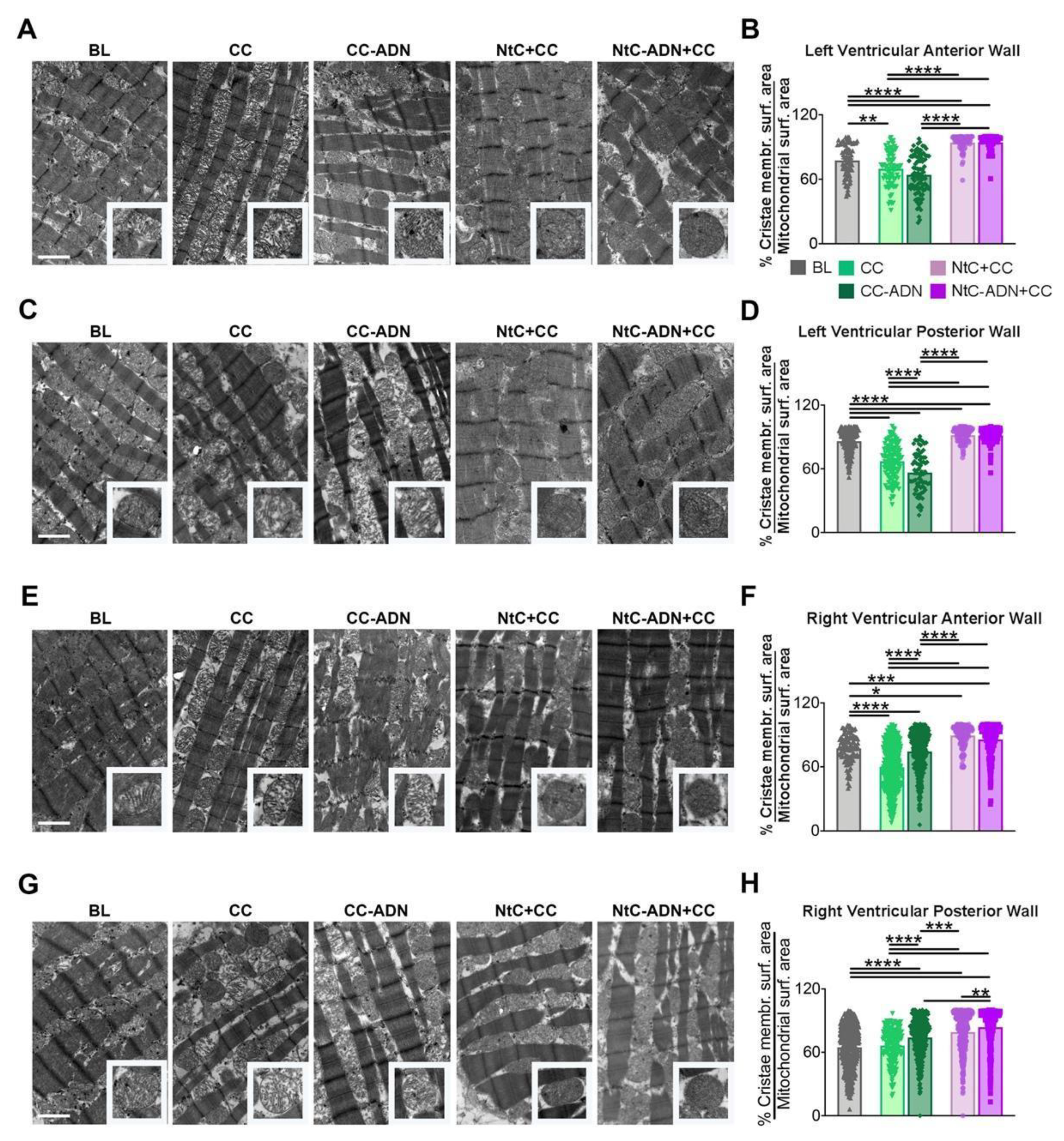
3.2. NtC + CC Treatment Preserves Mitochondrial Morphology and Cristae Structure
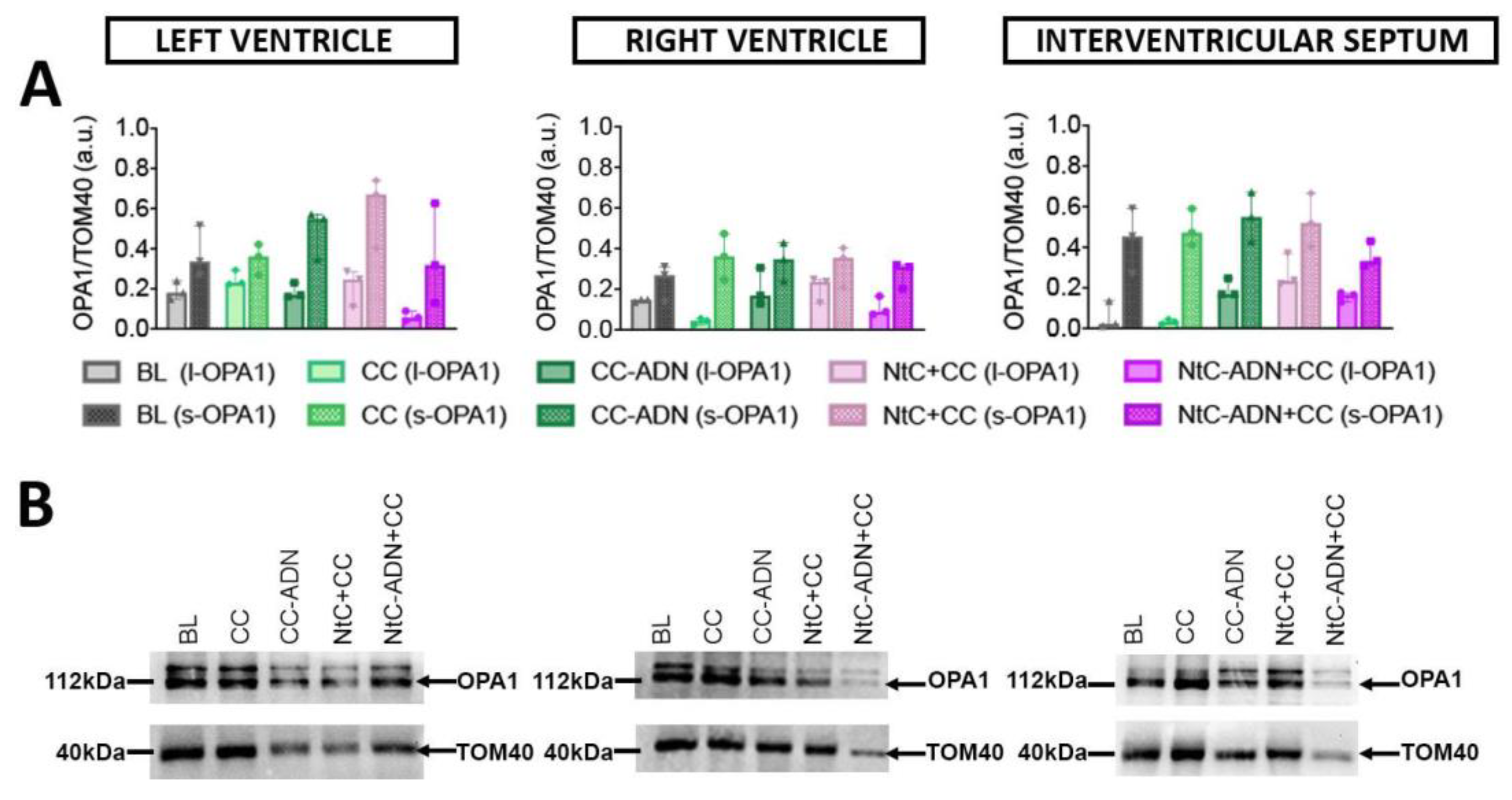
3.3. Different Protection Strategies Do Not Affect OPA1 Expression
3.4. Tissue Cytokines, Chemokines and Tissue Repair Mediators Have a Lower Expression after NtC + CC Treatment
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnard, C.N. The operation. A human cardiac transplant: An interim report of a successful operation performed at Groote Schuur Hospital, Cape Town. S. Afr. Med. J. 1967, 41, 1271–1274. [Google Scholar] [PubMed]
- Iyer, A.; Gao, L.; Doyle, A.; Rao, P.; Cropper, J.R.; Soto, C.; Dinale, A.; Kumarasinghe, G.; Jabbour, A.; Hicks, M.; et al. Normothermic Ex vivo perfusion provides superior organ preservation and enables viability assessment of hearts from DCD donors. Am. J. Transplant. 2015, 15, 371–380. [Google Scholar] [CrossRef] [PubMed]
- White, C.W.; Ali, A.; Hasanally, D.; Xiang, B.; Li, Y.; Mundt, P.; Lytwyn, M.; Colah, S.; Klein, J.; Ravandi, A.; et al. A cardioprotective preservation strategy employing ex vivo heart perfusion facilitates successful transplant of donor hearts after cardiocirculatory death. J. Hear. Lung Transplant. 2013, 32, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Chew, H.C.; Gao, L.; Villanueva, J.; Hicks, M.; Doyle, A.; Kumarasinghe, G.; Jabbour, A.; Jansz, P.C.; Feneley, M.P.; et al. Pathophysiological trends during withdrawal of life support: Implications for organ donation after circulatory death. Transplantation 2016, 100, 2621–2629. [Google Scholar] [CrossRef] [PubMed]
- Longnus, S.L.; Mathys, V.; Dornbierer, M.; Dick, F.; Carrel, T.P.; Tevaearai, H.T. Heart transplantation with donation after circulatory determination of death. Nat. Rev. Cardiol. 2014, 11, 354–363. [Google Scholar] [CrossRef]
- Zhou, T.; Prather, E.R.; Garrison, D.E.; Zuo, L. Interplay between ROS and antioxidants during ischemia-reperfusion injuries in cardiac and skeletal muscle. Int. J. Mol. Sci. 2018, 19, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görlach, A.; Dimova, E.Y.; Petry, A.; Martínez-Ruiz, A.; Hernansanz-Agustín, P.; Rolo, A.P.; Palmeira, C.M.; Kietzmann, T. Reactive oxygen species, nutrition, hypoxia and diseases: Problems solved? Redox Biol. 2015, 6, 372–385. [Google Scholar] [CrossRef]
- Ma, Y. Role of neutrophils in cardiac injury and repair following myocardial infarction. Cells 2021, 10, 1676. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Place, S.P.; Ferro, D.; Santovito, G. Peroxiredoxin 6 from the emerald rockcode Trematomus bernacchii: Molecular characterization and transcription studies in response to warming. J. Comp. Physiol. B 2016, 186, 59–71. [Google Scholar] [CrossRef]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Garofalo, F.; Pellegrino, D.; Gerdol, M.; Ferro, D.; Place, S.P.; Santovito, G. Molecular characterization of novel mitochondrial peroxiredoxins from the Antarctic emerald rockcod and their gene expression in response to environmental warming. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108580. [Google Scholar] [CrossRef] [PubMed]
- Sattin, G.; Bakiu, R.; Tolomeo, A.M.; Carraro, A.; Coppola, D.; Ferro, D.; Patarnello, T.; Santovito, G. Characterization and expression of a new cytoplasmic glutathione peroxidase 1 gene in the Antarctic fish Trematomus bernacchii. Hydrobiologia 2015, 761, 363–372. [Google Scholar] [CrossRef]
- Goyal, M.M.; Basak, A. Human catalase: Looking for complete identity. Protein Cell 2010, 1, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R. Glutathione peroxidases and redox-regulated transcription factors. Biol. Chem. 2006, 387, 1329–1335. [Google Scholar] [CrossRef]
- Iyer, A.; Gao, L.; Doyle, A.; Rao, P.; Jayewardene, D.; Wan, B.; Kumarasinghe, G.; Jabbour, A.; Hicks, M.; Jansz, P.C.; et al. Increasing the tolerance of DCD hearts to warm ischemia by pharmacological postconditioning. Am. J. Transplant. 2014, 14, 1744–1752. [Google Scholar] [CrossRef]
- Kotani, Y.; Ishino, K.; Osaki, S.; Honjo, O.; Suezawa, T.; Kanki, K.; Yutani, C.; Sano, S. Efficacy of MCI-186, a free-radical scavenger and antioxidant, for resuscitation of nonbeating donor hearts. J. Thorac. Cardiovasc. Surg. 2007, 133, 1626–1632. [Google Scholar] [CrossRef] [Green Version]
- Dhital, K.K.; Chew, H.C.; Macdonald, P.S. Donation after circulatory death heart transplantation. Curr. Opin. Organ. Transplant. 2017, 22, 189–197. [Google Scholar] [CrossRef]
- Larsen, T.; Solberg, S.; Johansen, R.; Jørgensen, L. Effect of cooling on the intracellular concentrations of Na+, K+ and Cl- in cultured human endothelial cells. Scand. J. Clin. Lab. Investig. 1988, 48, 565–571. [Google Scholar] [CrossRef]
- Parolari, A.; Rubini, P.; Cannata, A.; Bonati, L.; Alamanni, F.; Tremoli, E.; Biglioli, P. Endothelial damage during myocardial preservation and storage. Ann. Thorac. Surg. 2002, 73, 682–690. [Google Scholar] [CrossRef]
- Smulowitz, P.B.; Serna, D.L.; Beckham, G.E.; Milliken, J.C. Ex vivo cardiac allograft preservation by continuous perfusion techniques. ASAIO J. 2000, 46, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.; Otani, H.; Imamura, H.; Omori, K.; Inagaki, C. Role of sodium pump activity in warm induction of cardioplegia combined with reperfusion of oxygenated cardioplegic solution. J. Thorac. Cardiovasc. Surg. 1995, 110, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Stone, M.L.; Sharma, A.K.; Mas, V.R.; Gehrau, R.C.; Mulloy, D.P.; Zhao, Y.; Lau, C.L.; Kron, I.L.; Huerter, M.E.; Laubach, V.E. Ex vivo perfusion with adenosine A2A receptor agonist enhances rehabilitation of murine donor lungs after circulatory death. Transplantation 2015, 99, 2494–2503. [Google Scholar] [CrossRef] [Green Version]
- Leprince, P.; Popov, A.F.; Simon, A.R.; Benk, C.; Siepe, M. Ex vivo perfusion of the heart with the use of the organ care system. Eur. J. Cardio-Thorac. Surg. 2016, 49, 1318–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, M.B.E.; Coward, W.A.; Prentice, A.M.; Davies, P.S.W.; Strain, J.J.; McKenna, P.G.; Mahoney, C.A.; White, J.A.; Stewart, C.M.; Kerr, M.J.J. Daily energy expenditure in free-living children: Comparison of heart-rate monitoring with the doubly labeled water (2H2(18)O) method. Am. J. Clin. Nutr. 1992, 56, 343–352. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Conrad, N.; Judge, A.; Tran, J.; Mohseni, H.; Hedgecott, D.; Crespillo, A.P.; Allison, M.; Hemingway, H.; Cleland, J.G.; McMurray, J.J.V.; et al. Temporal trends and patterns in heart failure incidence: A population-based study of 4 million individuals. Lancet 2018, 391, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, G.M.; Meier, P.; White, S.K.; Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury: Looking beyond primary PCI. Eur. Heart J. 2013, 34, 1714–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arni, S.; Maeyashiki, T.; Citak, N.; Opitz, I.; Inci, I. Subnormothermic ex vivo lung perfusion temperature improves graft preservation in lung transplantation. Cells 2021, 10, 748. [Google Scholar] [CrossRef] [PubMed]
- White, C.W.; Messer, S.J.; Large, S.R.; Conway, J.; Kim, D.H.; Kutsogiannis, D.J.; Nagendran, J.; Freed, D.H. Transplantation of hearts donated after circulatory death. Front. Cardiovasc. Med. 2018, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Michel, S.G.; La Muraglia, G.M.; Madariaga, M.L.L.; Titus, J.S.; Selig, M.K.; Farkash, E.A.; Allan, J.S.; Anderson, L.M.; Madsen, J.C. Twelve-hour hypothermic machine perfusion for donor heart preservation leads to improved ultrastructural characteristics compared to conventional cold storage. Ann. Transplant. 2015, 20, 461–468. [Google Scholar] [CrossRef]
- Jing, L.; Yao, L.; Zhao, M.; Peng, L.P.; Liu, M. Organ preservation: From the past to the future. Acta Pharmacol. Sin. 2018, 39, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Pinnelas, R.; Kobashigawa, J.A. Ex vivo normothermic perfusion in heart transplantation: A review of the TransMedics® organ care system. Future Cardiol. 2021, 18, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Lu, Y.; Xin, L.; Gao, J.; Shang, C.; Jiang, Z.; Lin, H.; Fang, X.; Qu, Y.; Wang, Y.; et al. Role of oxidative stress in reperfusion following myocardial ischemia and its treatments. Oxid. Med. Cell. Longev. 2021, 2021, 6614009. [Google Scholar] [CrossRef] [PubMed]
- Becker, L.B. New concepts in reactive oxygen species and cardiovascular reperfusion physiology. Cardiovasc. Res. 2004, 61, 461–470. [Google Scholar] [CrossRef]
- Lodrini, A.M.; Goumans, M.-J. Cardiomyocytes cellular phenotypes after myocardial infarction. Front. Cardiovasc. Med. 2021, 8, 1629. [Google Scholar] [CrossRef]
- Hoppins, S.; Lackner, L.; Nunnari, J. The machines that divide and fuse mitochondria. Annu. Rev. Biochem. 2007, 76, 751–780. [Google Scholar] [CrossRef]
- Cipolat, S.; Rudka, T.; Hartmann, D.; Costa, V.; Serneels, L.; Craessaerts, K.; Metzger, K.; Frezza, C.; Annaert, W.; D’Adamio, L.; et al. Mitochondrial rhomboid PARL regulates cytochrome c release during apoptosis via OPA1-dependent cristae remodeling. Cell 2006, 126, 163–175. [Google Scholar] [CrossRef]
- Cogliati, S.; Frezza, C.; Soriano, M.E.; Varanita, T.; Quintana-Cabrera, R.; Corrado, M.; Cipolat, S.; Costa, V.; Casarin, A.; Gomes, L.C.; et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell 2013, 155, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermann, B. Mitochondrial fusion and fission in cell life and death. Nat. Rev. Mol. Cell Biol. 2010, 11, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.; Wai, T.; Baker, M.J.; Kladt, N.; Schauss, A.C.; Rugarli, E.; Langer, T. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell Biol. 2014, 204, 919–929. [Google Scholar] [CrossRef] [PubMed]
- MacVicar, T.; Langer, T. OPA1 processing in cell death and disease—The long and short of it. J. Cell Sci. 2016, 129, 2297–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olichon, A.; Baricault, L.; Gas, N.; Guillou, E.; Valette, A.; Belenguer, P.; Lenaers, G. Loss of OPA1 perturbates the mitochondrial inner membrane structure and integrity, leading to cytochrome c release and apoptosis. J. Biol. Chem. 2003, 278, 7743–7746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Carelli, V.; Manfredi, G.; Chan, D.C. Proteolytic cleavage of Opa1 stimulates mitochondrial inner membrane fusion and couples fusion to oxidative phosphorylation. Cell Metab. 2014, 19, 630–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilkerson, R.; De La Torre, P.; Vallier, S.S. Mitochondrial OMA1 and OPA1 as gatekeepers of organellar structure/function and cellular stress response. Front. Cell Dev. Biol. 2021, 9, 527. [Google Scholar] [CrossRef]
- Yao, W.; Li, H.; Luo, G.; Li, X.; Chen, C.; Yuan, D.; Chi, X.; Xia, Z.; Hei, Z. SERPINB1 ameliorates acute lung injury in liver transplantation through ERK1/2-mediated STAT3-dependent HO-1 induction. Free Radic. Biol. Med. 2017, 108, 542–553. [Google Scholar] [CrossRef]
- Lei, I.; Huang, W.; Ward, P.A.; Pober, J.S.; Tellides, G.; Ailawadi, G.; Pagani, F.D.; Landstrom, A.P.; Wang, Z.; Mortensen, R.M.; et al. Differential inflammatory responses of the native left and right ventricle associated with donor heart preservation. Physiol. Rep. 2021, 9, e15004. [Google Scholar] [CrossRef] [PubMed]
- Savio, L.E.B.; Leite-Aguiar, R.; Alves, V.S.; Coutinho-Silva, R.; Wyse, A.T.S. Purinergic signaling in the modulation of redox biology. Redox Biol. 2021, 47, 102137. [Google Scholar] [CrossRef]
- Liu, H.; Xia, Y. Beneficial and detrimental role of adenosine signaling in diseases and therapy. J. Appl. Physiol. 2015, 119, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, G.; Milano, G.; Moccetti, T. Role of mitogen-activated protein kinases in myocardial ischemia-reperfusion injury during heart transplantation. J. Transplant. 2012, 2012, 928954. [Google Scholar] [CrossRef] [Green Version]
- Rudd, D.M.; Dobson, G.P. Eight hours of cold static storage with adenosine and lidocaine (Adenocaine) heart preservation solutions: Toward therapeutic suspended animation. J. Thorac. Cardiovasc. Surg. 2011, 142, 1552–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlsson, A.; Sobrosa, C.; Kaijser, L.; Jansson, E.; Bomfim, V. Adenosine in cold blood cardioplegia—A placebo-controlled study. Interact. Cardiovasc. Thorac. Surg. 2012, 14, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, F.; Azarfarin, R.; Salahi, S.; Baharestani, B.; Ghavidel, A.A. The effects of cold blood cardioplegia with and without adenosine on hemodynamic status of patients undergoing mitral valve surgery. Multidiscip. Cardiovasc. Ann. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Francica, A.; Tonelli, F.; Rossetti, C.; Tropea, I.; Luciani, G.B.; Faggian, G.; Dobson, G.P.; Onorati, F. Cardioplegia between evolution and revolution: From depolarized to polarized cardiac arrest in adult cardiac surgery. J. Clin. Med. 2021, 10, 4485. [Google Scholar] [CrossRef]
- Giordano, P.; Scrascia, G.; D’Agostino, D.; Mastro, F.; Rotunno, C.; Conte, M.; Rociola, R.; Paparella, D. Myocardial damage following cardiac surgery: Comparison between single-dose Celsior cardioplegic solution and cold blood multi-dose cardioplegia. Perfusion 2013, 28, 496–503. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BL | CC | CC-ADN | NtC + CC | NtC-ADN + CC | ||
|---|---|---|---|---|---|---|
| GM-CSF | LV | 15.7 (3.4) | 17.1 (5.4) | 15.9 (1.5) | 5.6 (5.6) * | 9.7 (2.3) |
| RV | 12.25 (3.3) | 15.5 (2.1) | 12.9 (2.7) | 10.8 (1.6) | 8.1 (5.3) | |
| IVS | 15.9 (8.4) | 16.2 (2.8) | 18.5 (5.1) | 7.7 (5.4) * | 13.5 (3.8) | |
| TNF-a | LV | - | - | - | - | - |
| RV | - | - | - | - | - | |
| IVS | - | - | - | - | - | |
| INF-g | LV | - | - | - | - | - |
| RV | - | - | - | - | - | |
| IVS | - | - | - | - | - | |
| IL-1a | LV | - | 1.5 (0.8) | 0.5 (0.8) | 1.7 (2.9) | - |
| RV | - | - | - | - | - | |
| IVS | 0.5 (0.8) | - | - | - | - | |
| IL-1b | LV | 3.7 (3.7) | - | 7.5 (13.4) | - | - |
| RV | - | 6.5 (6.5) | 2.8 (4.9) | 1.5 (2.5) | 1.7 (3.1) | |
| IVS | - | - | - | - | - | |
| IL-1ra | LV | 15.1 (0.6) | 13.7 (1.1) | 14.9 (4.1) | 13.1 (3.9) | 12.7 (5.2) |
| RV | 13.3 (1.6) | 13.6 (3.8) | 11.8 (2.7) | 10.8 (0.9) | 13.7 (3.1) | |
| IVS | 15.4 (1.7) | 14.9 (1.4) | 16.5 (4.8) | 12.6 (2.2) | 13.8 (3.7) | |
| IL-2 | LV | 12.3 (2.5) | 9.5 (5.5) | 13.6 (2.3) | 6.4 (4.6) | 6.9 (4.2) |
| RV | 10.3 (1.7) | 18.6 (13.4) | 11.4 (1.5) | 8.2 (4.8) | 9.7 (5.9) | |
| IVS | 9.3 (5.5) | 12.4 (0.5) | 21.3 (11.8) | 6.5 (4.6) | 9.3 (5.7) | |
| IL-4 | LV | 10.2 (3.3) | 6.5 (1.2) | 9.1 (5.4) | 2.2 (5.4) * | 1.9 (2.1) * |
| RV | 10.5 (2.7) | 11.7 (4.3) | 8.1 (6.4) | 4.3 (5.2) * | 0.8 (1.2) * | |
| IVS | 6.1 (7.1) | 9.4 (3.2) | 10.7 (4.8) | 4.8 (8.2) | 4.1 (5.1) | |
| IL-6 | LV | - | - | 3.1 (5.1) | - | 2 (3.1) |
| RV | - | 2.5 (2.5) | 1.2 (1.4) | 1.5 (2.5) | 0.8 (1.3) | |
| IVS | - | - | 1.3 (1.9) | 1.9 (3.3) | 2.1 (3.4) | |
| IL-8 | LV | 28.9 (16.2) | 20.4 (10.7) | 17.4 (5.2) | 13.8 (7.7) | 4.8 (4.8) ** |
| RV | 19.7 (6.3) | 45.3 (8.9) | 35.2 (12.1) | 18.1 (0.8) * | 30.9 (6.9) | |
| IVS | 28.9 (13.2) | 37.6 (5.5) | 39.3 (14.4) | 21.7 (13.1) | 32.6 (19.7) | |
| IL-10 | LV | - | - | - | - | - |
| RV | 27.1 (14.1) | 40.8 (11.1) | 25.7 (13.3) | 35.1 (9.4) | 28.7 (15.5) | |
| IVS | 35.6 (7.1) | 28.9 (4.7) | 38.4 (6.1) | 47.6 (2.0) | 43.4 (6.8) | |
| IL-12 | LV | - | - | - | - | - |
| RV | - | - | - | - | - | |
| IVS | - | - | - | - | - | |
| IL-18 | LV | - | - | - | - | - |
| RV | - | - | - | - | - | |
| IVS | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tolomeo, A.M.; Fabozzo, A.; Malvicini, R.; De Lazzari, G.; Bisaccia, P.; Gaburro, G.; Arcidiacono, D.; Notarangelo, D.; Caicci, F.; Zanella, F.; et al. Temperature-Related Effects of Myocardial Protection Strategies in Swine Hearts after Prolonged Warm Ischemia. Antioxidants 2022, 11, 476. https://doi.org/10.3390/antiox11030476
Tolomeo AM, Fabozzo A, Malvicini R, De Lazzari G, Bisaccia P, Gaburro G, Arcidiacono D, Notarangelo D, Caicci F, Zanella F, et al. Temperature-Related Effects of Myocardial Protection Strategies in Swine Hearts after Prolonged Warm Ischemia. Antioxidants. 2022; 11(3):476. https://doi.org/10.3390/antiox11030476
Chicago/Turabian StyleTolomeo, Anna Maria, Assunta Fabozzo, Ricardo Malvicini, Giada De Lazzari, Paola Bisaccia, Gianluca Gaburro, Diletta Arcidiacono, Denni Notarangelo, Federico Caicci, Fabio Zanella, and et al. 2022. "Temperature-Related Effects of Myocardial Protection Strategies in Swine Hearts after Prolonged Warm Ischemia" Antioxidants 11, no. 3: 476. https://doi.org/10.3390/antiox11030476