
Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota



1
Programa de Doctorado en Ingeniería de Alimentos, Universidad del Bío-Bío, Av. Andrés Bello 720, Chillan 3780000, Chile
2
Laboratorio de Crioconcentración, Departamento de Ingeniería en Alimentos, Universidad del Bío-Bío, Av. Andrés Bello 720, Chillan 3780000, Chile
3
Departamento de Ingeniería en Alimentos, Facultad de Ingeniería, Universidad de La Serena, Av. Raúl Bitrán 1305, La Serena 1700000, Chile
4
Grupo de Crioconcentración de Alimentos y Procesos Relacionados, Universidad del Bío-Bío, Av. Andrés Bello 720, Chillan 3780000, Chile
*
Authors to whom correspondence should be addressed.
Antioxidants 2022, 11(1), 109; https://doi.org/10.3390/antiox11010109
Submission received: 21 November 2021
/
Revised: 28 December 2021
/
Accepted: 30 December 2021
/
Published: 4 January 2022
(This article belongs to the Special Issue Application of Antioxidants and Bioactive Compounds in Food from Agriculture to Health Benefits)
Abstract
:Helicobacter pylori (H. pylori) infection affects more than half of the world’s population, and thus, about 10 to 20% of people with H. pylori suffer from peptic ulcers, which may ultimately lead to gastric cancer. The increase in antibiotic resistance and susceptibility has encouraged the search for new alternative therapies to eradicate this pathogen. Several plant species are essential sources of polyphenols, and these bioactive compounds have demonstrated health-promoting properties, such as the gut microbiota stimulation, inflammation reduction, and bactericidal effect. Therefore, this review aims to discuss the potential effect of plant-based polyphenols against H. pylori and their role in the gut microbiota improvement.
1. Introduction
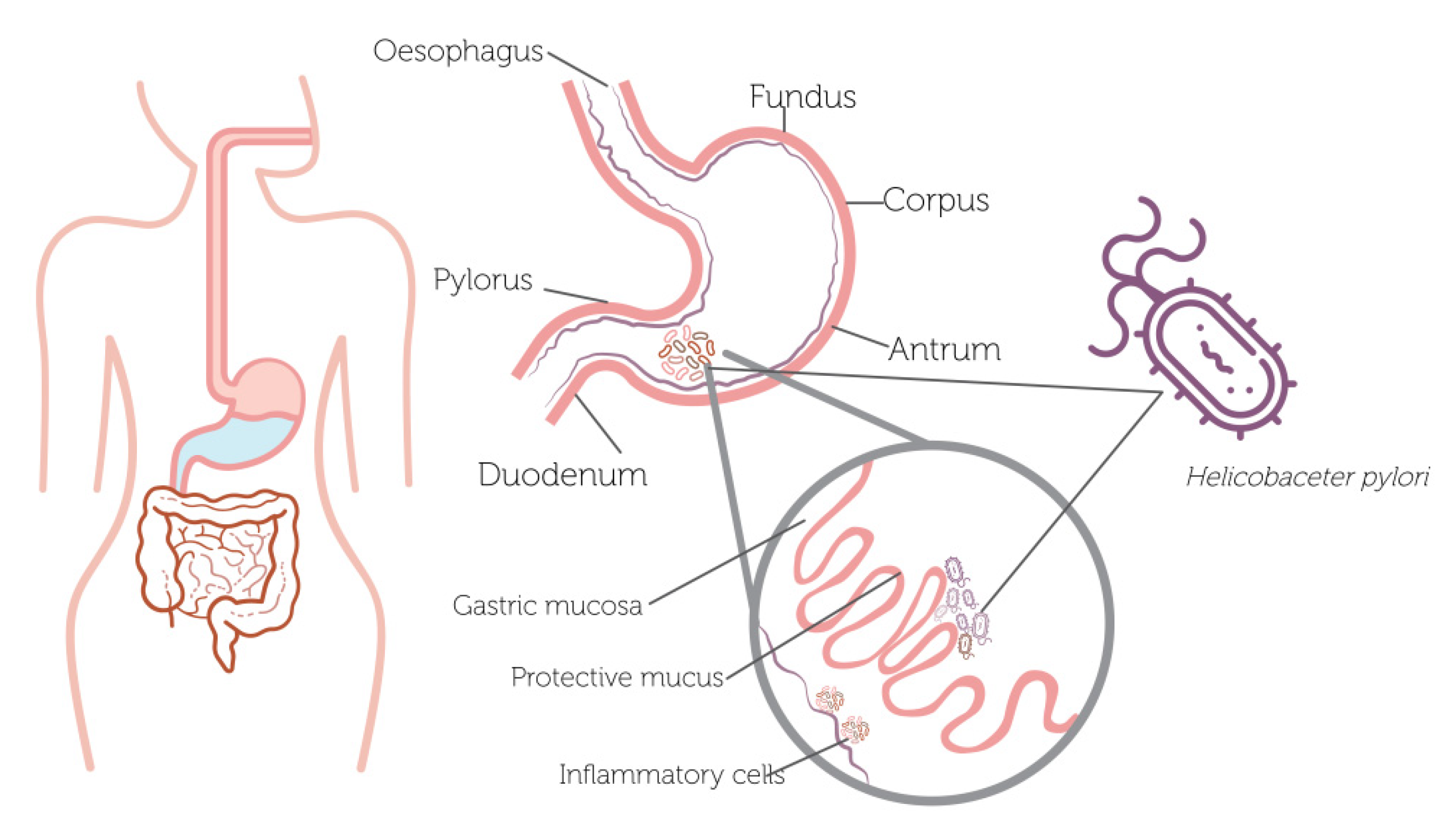
Helicobacter pylori (H. pylori) is a Gram-negative spiral bacterium that has long been recognized by its ability to colonize chronically the stomach lining and the upper section of the intestine, generating gastritis, gastroduodenal ulcers and gastric carcinoma [1]. Specifically, H. pylori establishes the optimal environmental conditions to survive short acidic exposures, since it produces two enzymes, urease and carbonic anhydrase (CA), where the urease converts the host urea into ammonia and carbon dioxide (CO2), while CA catalyzes the transformation of CO2 into bicarbonate (HCO3-). Hence, both reactions allow to increase the pH levels. Thereby, the host’s immune response is affected by the constant inflammation of the epithelial cells, causing the adaption and survival of the H. pylori over time in the local environmental conditions [2,3]. Furthermore, H. pylori continuously moves along the host tissues through chemotaxis (directed motility) by controlling flagellar rotation, where the bacterial cells travel toward advantageous environments or away from disadvantageous environments [4,5,6,7], being this movement key to colonize from the lumen to the mucus layer, overlying the epithelium due to less acidic conditions [8]. Moreover, another factor that may influence H. pylori colonization is the configuration of the stomach, since the human stomach is a heterogeneous and complex system, composing of various kinds of regions that differ in cellular composition and conditions, where the human stomach has three glandular regions: fundus, corpus, and antrum [9].
H. pylori infection affects more than half of the world population, and it plays a crucial role in gastric diseases [10]. This pathogen causes gastroduodenal inflammation that can lead to atrophic gastritis, gastric ulcers, and, finally, gastric cancer (one of the major causes of mortality worldwide, according to the National Comprehensive Cancer Network) [11]. As shown in Figure 1, H. pylori generally proliferates in the upper section of the intestine and gastric mucosa, triggering an inflammatory process.
According to Sjomin et al. [12], the global prevalence of H. pylori is similar between genders, with 42.7% in females and 46.3% in males [13], increasing with risks factors on eating habits and lifestyle behaviors such as poor hand hygiene, low frequency of hand-washing practice, low consumption of fruits and vegetables, and high consumption of fried foods [12]. Hence, around 20% of the world population infected with H. pylori suffers peptic ulcers, which ultimately produce gastric cancer, and thus, studies have indicated that 75% of gastric cancer can be attributed to H. pylori infection [4,14].
In the same way, the most common treatment against H. pylori infection is the combination of two antibiotics and a proton pump inhibitor administered twice a day, and it presents high effectivity (≈90%). However, the treatment produces metabolic side effects such as mitochondrial damage and hypoprothrombinemia [15,16]. In addition, the human genetic susceptibility to antibiotics, the various mechanisms of H. pylori to resist gastric acidity conditions, and the globally observed increase in antibiotic resistance of H. pylori have led researchers to seek and propose effective and novel alternative therapies against this pathogen through the use of different bioactive components.
Thus, in recent times, polyphenols and phytochemicals from plants, fruits, and vegetables, have demonstrated interesting antimicrobial properties, suggesting the possibility that bioactive compounds can assist in the prevention and treatment of microbial pathogens. Thereby, several studies have assessed the role of bioactive compounds as antibacterial properties, and ways to modulate the gut microecosystem to promote the population of probiotics, and thus, favoring effective prevention and treatments against bacterial infections [17,18,19].
Therefore, this review aims to discuss the therapeutic effects of plant-based polyphenols against H. pylori and the potential role to improve the gut microbiota in human metabolic health.
2. Healthy Effects of Polyphenols and Antioxidants
Polyphenols are natural bioactive compounds derived from plants, fruits, and vegetables. These bioactive compounds have different phenolic hydroxyl groups in their structure, and thus, it is possible to identify phenolic acids, tannins, carotenoids, flavonoids, anthocyanins, and proanthocyanidins, among others, where the molecules present a wide variety of concentrations in the foods [20]. Thereby, polyphenols display a myriad of health-beneficial properties on the human body, since these components have demonstrated different medicinal and nutraceutical properties, helping in the gastrointestinal digestion [21], reducing the blood lipid levels [22], improving the body immunity [23], and in addition, polyphenols present neuroprotective, cardioprotective, anti-inflammatory, anti-diabetic, anti-carcinogenic, and anti-ageing effects, among other health improvements [24,25,26]. Nonetheless, despite all advantages of polyphenols, these components are very bitter and astringent tastes, making it difficult to incorporate into food or model foods [27], and also, polyphenols are unstable under certain conditions such as exposure to light, presence of oxygen and enzymes, high temperatures, and pH changes, weakening their activity [28,29,30].
An important point, studies on the potential prebiotic effect of polyphenols on the composition and function of the gut microbiota, gut permeability, gut-derived proinflammatory stimuli, have increased considerably, and the results have been linked to the high production of anti-inflammatory molecules such as interleukin-4 (IL-4), interleukin-10 (IL-10), interleukin-13 (IL-13), and adiponectin [31,32,33,34]. Moreover, dietary polyphenols can contribute substantially to reduce the frequency of oxidative stress-induced damage triggered by inflammation or infectious process, and thus, it plays a vital role in chemoprevention through different action mechanisms such as nuclear factor erythroid 2-related factor 2 (Nrf2) and Nrf2-antioxidant response element (ARE) pathway, and phase I and phase II detoxifying enzymes [35]. Specifically, Nrf2-Are is an essential intermediary, since it induces cellular response against oxidative stress, a condition due to either a mechanism of aggressive cytotoxicity level, producing more reactive oxygen species (ROS) or the cellular incapacity to remove the reaction of reactive oxidant [36]. Besides, for phase I detoxifying enzymes, it interacts with nucleophilic groups, causing mutagenic gashes. Then, the products of phase I are purified through phase II detoxifying enzymes, inactivating ROS and detoxifying carcinogens. Thus, phase II enzymes catalyze electrophiles conjugation along with metabolites (xenobiotics and endogenous) of oxidative stress-induced lipid peroxidation [35]. Thereby, there is an association between diets with high polyphenol and antioxidant contents, allowing minimal toxicity of healthy tissues due to the reduction of oxidative stress and the protection of extracellular and intracellular DNA [37,38].
During the process of colonization of the intestinal epithelium, H. pylori triggers oxidant-sensitive transcription factors that cause tissue damage at the molecular and cellular levels [39], and consequently the activation of nuclear factor kappa-light chain-cancer-activated B cells (NF-kB), which induces the expression of the interleukin-8 (IL-8) gene [40,41,42]. In addition, H. pylori also activate nicotinamide adenine dinucleotide dinucleotide phosphate (NADPH) oxidase, which is a major source of ROS, also the cyclooxygenase (COX), nitric oxide (NO) and reactive nitrogen species (RNS) [43]. Mitochondria are susceptible to attack by ROS, and thus, these dysfunctional mitochondria drive the production of inflammatory cytokines [44].
The main attribute of polyphenols and their metabolites is their antioxidant action by targeting immune cells activating different signaling pathways that alter interleukins, cyclooxygenase, nitric oxide synthase and other inflammatory responses [45] caused by diseases and infections, such as H. pylori. Some polyphenols affect the NF-kB pathway by inhibiting the phosphorylation of kinases; preventing the translocation and transcription of proinflammatory mediators, also hindering the interaction between NF-kB and its predicted DNA [46]. Each type of polyphenol has a particular antioxidant mechanism; it is believed that the antioxidant activity of phenolic acids lies in their ability to scavenge radicals by donating hydrogen atoms [47]. In the case of stilbenoids, their antioxidant action is attributed to the presence of a hydroxyl in the ortho-position, which allows them to scavenge superoxide radicals [48]. Likewise, flavonoids have the ability to counteract the NF-kB signaling pathway and prevent the inflammatory effects of NO [46]; the inhibitory effects of flavonoids on the colonic expression of one of the COX isoforms (inducible COX-2) have also been described [49]. Nrf2 and NF-kB signaling pathways can be modulated by anthocyanins, catechins and ellagitannins; also anthocyanins can reduce COX-2 and NO and ellagitannins decrease IL-8 secretion [46]. Therefore, the action of polyphones in the management of H. pylori infection is by modulating inflammatory signals and factors that induce oxidative stress and thus the appearance of free radicals associated with further tissue damage.
Specifically, the studies on bioactive compounds, derived from foods, have grown exponentially since the mid-1990s, due to their beneficial effects as antimicrobial agents, and in addition, bioactive compounds have received growing attention due to the antibiotic resistance of pathogenic bacteria [50]. For example, a meta-analysis study was conducted to indicate the beneficial effects on human health of polyphenol-rich foods such as apples, wine, tea, and berries. This study emphasized that the doses of polyphenol intake affects the abundance of human gut microbiota, where the intake of polyphenols changed the gut microbiota of human subjects, since it stimulated the presence of Lactobacillus and Bifidobacterium species, and in turn, the polyphenols limited the presence Clostridium species. Thus, the polyphenols intake between 396–540 mg/day maintains a positive balance in the gut microbiota composition due to the increase in health-promoting species [51]. Moreover, recent works have emphasized the influence of dietary patterns on polyphenols daily intake and its effect on the gut microbiota composition, since a study in a U.S. cohort concluded that the diet presents between 498–662 mg/day [52], while Mediterranean dietary pattern has an average of 1171 mg/day [53], and Atlantic Diet provides 1011–1284 mg/day of polyphenols [54]. Therefore, each diet has a different effect in the body’s immune system against infectious agents such as viruses and bacteria (H. pylori) [55].
In this context, authors have indicated that H. pylori infection affects oxidative stress status of the host [56,57,58]. However, the consumption of dietary antioxidants can be effective to reduce negative aspects of H. pylori [59]. In this way, Judaki et al. [60] reported a significant reduction in the amount of oxidative DNA damage and apoptosis in H. pylori positive individuals after consuming curcumin (700 mg three times daily) during three months. Also, Jones et al. [61] demonstrated that capsaicin inhibits the growth of H. pylori, since capsaicin has been recognized as a potential anti-inflammatory agent due to the production of IL-8 in the gastric epithelium of H. pylori-positive individuals [62]. Similarly, Yanagawa et al. [63] showed that epigallocatechin-3-gallate (from green tea) increased the antibacterial activity of an antibiotic therapy, with high efficiency against Helicobacter pylori growth in vitro.
However, despite these advantages of polyphenols for human health, there are some concerns about polyphenol consumption, since the fortification and supplementation from polyphenol extracts can substitute fruit and vegetable intake, but these extracts may not have the same health benefits as the fresh fruits and vegetables due to different interactions between compounds after the extraction methods [64]. Silva and Pogačnik [65] have recognized the need to study any relationship between the overconsumption of polyphenol intake and the effect on the gut microbiota due to the accumulation of different molecules in the organism.
3. Polyphenols and Gut Microbiota
3.1. Biotransformation of Polyphenols by Gut Microbiota
Polyphenols present a low absorption rate in the human gut, where the colon has highest percentage of polyphenols absorption [66]. From this, multiple investigations have evaluated the role of phenolic compounds to regulate the intestinal microecosystem and to promote the growth of healthy bacteria such as Akkermansia, Faecalibacterium, Lactobacillus, Bifidobacterium and Enterococcus spp., since these microorganisms have been recognized for their anti-inflammatory and immuno-regulating effects, and thus, these species can be useful against infectious pathogens [67,68,69].
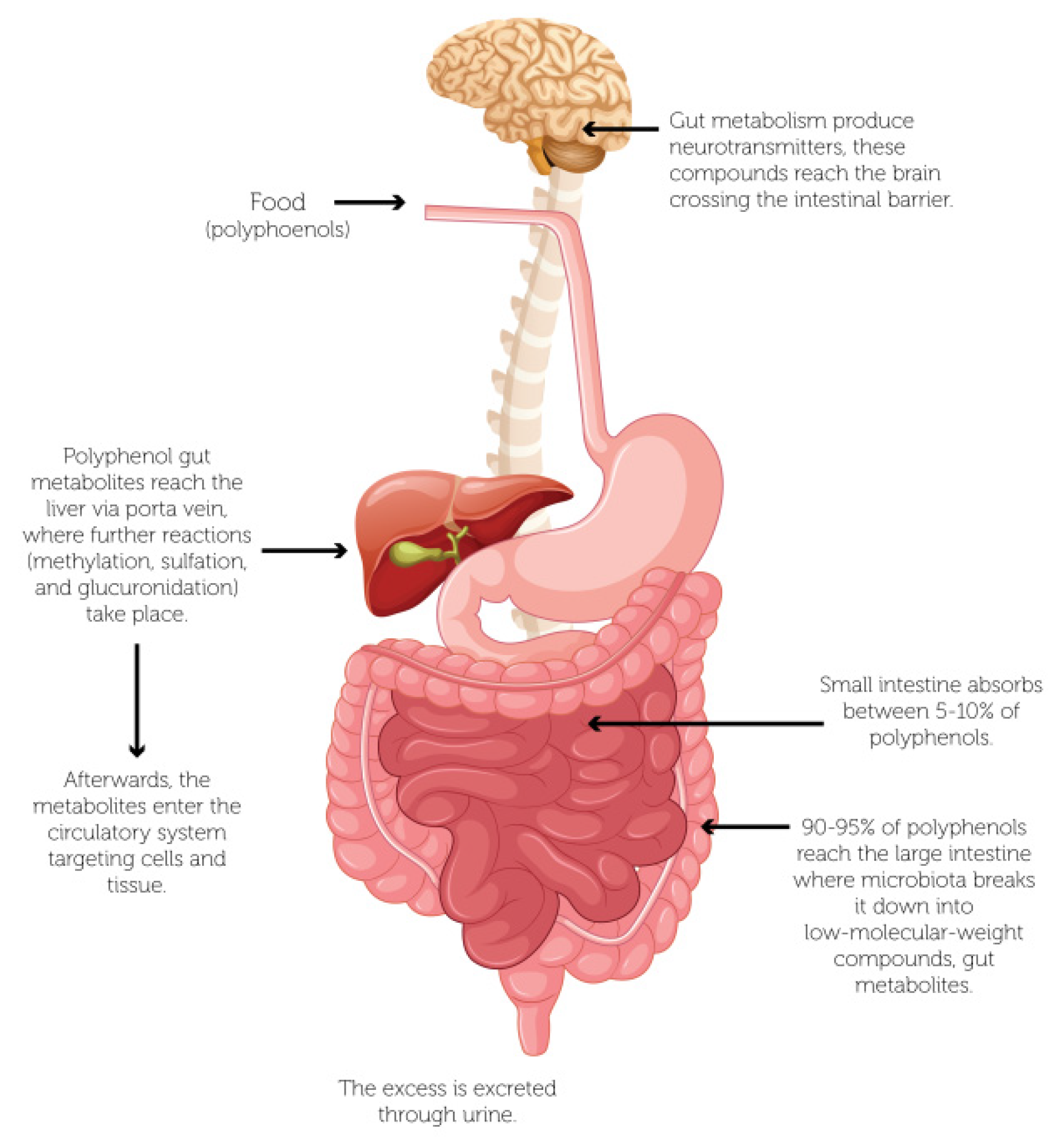
As mentioned above, polyphenols can be hydrolyzed by enzymes or metabolized by the gut microbiota. Thereby, these compounds differ from those in fresh or processed foods, since the polyphenols hydrolyzed (or metabolized) can reach the bloodstream, tissues, and brain, of the host [70]. In the same way, the chemical structure of polyphenols can be related to their bioavailability. Thus, polyphenols commonly consumed in the diet are esters, glycosides, or polymers, but these compounds cannot be absorbed in their original form [64]. Therefore, gut microbiota plays a vital role in the biotransformation of bioactive compounds, especially polyphenols, where 5–10% can be absorbed in the small intestine, and the remaining percentage can accumulate in the large intestine [71,72]. Hence, the microbiota produces decomposition of polyphenol structures into low-molecular-weight phenolic compounds, and they all have health positive effects on consumers, but the biotransformation varies significantly among individuals and populations [73]. Concretely, the bioaccessibility of phenolic compounds can vary significantly. For example, the anthocyanins have a bioaccessibility close to 5–10%, i.e., approximately 90–95% of polyphenols reach the colon [74]. Thus, the biotransformation by gut microbiota mainly involves the interaction of Bacteroides, Bifidobacterium, Clostridium, Fusobacterium, Lactobacillus and Peptostreptococus species [75]. Table 1 displays the biotransformation of polyphenols by gut microbiota and main metabolites.
3.2. Metabolism of Dietary Polyphenols: Role of the Microbiota
Polyphenols can influence the function and composition of the microbiota, since the microbiota generates metabolites (mainly aromatic and phenolic acids) through interaction with polyphenols, and the biotransformation process occurs mainly in the large intestine [63]. Figure 2 shows the biotransformation of polyphenols from food in the gastrointestinal tract, where the microbiota breaks the polyphenols into low-molecular-weight compounds through hydroxylation, isomerization, dehydrogenation, glycosylation decarboxylation, and methylation, among other chemical reactions. Then, the metabolites are absorbed in the small intestine and reach the liver through the portal vein, where other reactions are carried out by several methods such as methylation, sulfation, and glucuronidation. Hence, these metabolites enter the circulatory system and transferred to remote cells and tissues [72,77,78].
In vitro assays indicate that polyphenols may exert prebiotic-like effects on the intestinal microflora, promoting the growth [79]. Specifically, an in vitro study on tea polyphenols, it revealed the potential role of polyphenols to improve the profusion of Lactobacillus, Enterococcus and Bifidobacterium spp. [80,81]. Tomás-Barberán et al. [18] concluded that polyphenols could modulate the growth of Roseburia, Akkermansia and Faecalibacterium spp. Thereby, there is synergy between dietary polyphenols and gut microbiota, and thus, the gut microbiota can transform polyphenols into secondary metabolites, making them more bioavailable, and with highly beneficial effects to the consumers. Therefore, while polyphenols can modify the microbiota, the growth of pathogens can be prevented [82].
4. Anti-Helicobacter pylori Effects of Plant-Based Polyphenols
Various studies have highlighted the potential role of polyphenols against the adverse consequences of H. pylori infection [83]. However, the polyphenols do not totally eradicate H. pylori from the host, but these components can significantly reduce the negative effects (bacterial colonization, inflammatory responses and mucosal atrophy) of the infection, and in turn, the polyphenols allow to reduce the dose of antibiotics given to the patient, since the interaction polyphenols/antibiotics have demonstrated synergistic effects on H. pylori action [84]. Thus, ellagic acid has shown promising effects, since Chung [85] demonstrated excellent results with increased dose of ellagic acid on the arylamine N-acetyltransferase (NAT) activity in H. pylori cytosols, with different percentages of 2-aminofluorene and p-aminobenzoic acid acetylation in relation to the concentration of ellagic acid. Similarly, Martini [86] showed that isolated polyphenols such as ellagic acid, kaempferol, gallic acid and quercetin 3-O-β-D-glucopyranoside had different bactericidal effects against two H. pylori strains (CagA+ strain 10 K and CagA− strain G21), suggesting that the factors of time of exposure and concentration are responsible for a correct saturation of the CagA strains, and in addition, the individual polyphenols inactivated the H. pylori ion pumps (the flux of copper and metal cations through membranes regulated by enzymes). Zhang et al. [87] showed the inhibition of H. pylori infection through resveratrol, since this component decreased the H. pylori-induced mRNA transcription and protein expression levels of inducible nitric oxide synthase (iNOS) and IL-8, and it decreased the urease activity by ≈90%. Moreover, resveratrol increased the activity of heme oxygenase-1 (HO-1), an antioxidant enzyme with therapeutic target against oxidative stress and gastrointestinal diseases, and Nrf2, a transcriptional regulator of HO-1, and in turn, a cellular sensor of oxidative stress and it induces expression of cytoprotective genes, reducing the damage of ROS. Lee et al. [88] indicated that epigallocatechin inhibited the glycosylation of H. pylori-induced toll-like receptor 4 (TLR-4), and also, the component inhibited the urease enzyme, diminishing the damage on DNA and gastric mucosa cytotoxicity of epithelial cells by H. pylori.
In the same way, Mandalari et al. [17] recapitulated existing knowledge on citrus fruits and their bioactive compounds against H. pylori, and thus, the study indicated that β-Myrcene, hesperetin-7-O-glucoside, boropinic acid, sudachitin, 3′-demethoxysudachitin, auraptene, and bergamottin reduced the viability of H. pylori to 90%. Thereby, the polyphenols of citrus fruits and their derivatives produce an effective inhibition and eradication of H. pylori, and these positive effects can be individually or in combination with various antibiotics. In the same way, Takeuchi et al. [89] reported the constituents derived from nature to counteract the negative effects of H. pylori specie. Thus, the authors mentioned the potential of bioactive compounds found in foods and their derived products such as bovine milk (lactoferrin), green tea (catechin), broccoli sprout (sulforaphane), garlic (allicin), ginger (6-shogaol, 6-gingerol, 8-gingerol, and 10-gingerol), and apple peel (quercetin glycosides), among others, as alterative agents instead of antibiotics, and in turn, these foods can be more effective and safe to the consumers, helping in the use of a low dose of antibiotics. Specifically, Escandón et al. [90] reported that kaempferol and (-)-epicatechin had a significant suppression effect on the growth of H. pylori in a concentration-dependent manner. The results also proved that the inhibitory effect of H. pylori depends on the time of exposure to polyphenol accumulation. Thus, a mixture of low doses of kaempferol and (-)-epicatechin exhibited antibacterial activity. Chua et al. [91] explored the effects of blueberry and grape seed extracts for H. pylori eradication, showing no significant difference between the extracts and placebo group to eradicate the pathogen. However, the study emphasizes that the procyanidins from grape seed can help as an anti-inflammatory adipokine on low-grade inflammatory diseases. Cardoso et al. [92] studied Wild strawberry leaves (Fragaria vesca L.) and Agrimonia eupatoria L. (plant of the Rosaceae family) extracts, and an ellagitannin-enriched fraction of wild strawberry leaves against different clinical isolates of H. pylori, where all the fractions presented anti-H. pylori activity, but the results revealed an intense activity of ellagitannin-enriched fraction of wild strawberry leaves to prevent the growth of H. pylori infection, since it inhibited 67% of the pathogen, suggesting potential of plant extracts as bioactive agents in association with antibiotic therapy. Park et al. [93] used red raspberry (Rubus crataegifolius) and elm tree (Ulmus macrocarpa) to counteract the presence of H. pylori in the human body, and the study showed that the ellagic acid of red raspberry and the catequin of elm tree achieved excellent results against the pathogen, since these bioactive compounds prevented (separately or in combination) the growth of H. pylori. Thereby, the combination of red raspberry (75 μg/mL) and elm tree (75 μg/mL) proved to have a synergetic effect against this pathogen. Thereby, the combination of samples could be an alternative as a new and safe herbal product against anti-H. pylori infection. Moreover, Torres et al. [94] demonstrated that proanthocyanidins of avocado significantly decreased inflammatory signals of gastric adenocarcinoma cells infected with H. pylori, since it decreased the adherence of H. pylori infection in the organism. Similarly, Spósito et al. [95] indicated that Caseria sylvestris leaves derivatives displayed an important anti-H. pylori activity, where terpenes (diterpenes and sesquiterpenes), phenolic compounds presented high synergism against H. pylori through in vitro and in vivo assays. Betoret et al. [96] developed a low moisture apple snack impregnated with mandarin and pineapple/grape juices, and inoculated with a high microbial content of Lactobacillus salivarius spp. salivarius. Thus, the snacks showed positive effects against H. pylori, since the polyphenols and pH of the juices impeded the growth of the pathogen, and in turn, this combination allowed the growth of the probiotic. Barrera et al. [97] evaluated the effects of phenolic compounds (vitamin C, total phenols and flavonoids) and antioxidant content (ABTS-TEAC and DPPH assays) of clementine juice inoculated with Lactobacillus salivarius spp. salivarius on microbial counts and survival to in vitro digestion, where the results indicated that the inhibition of H. pylori growth was produced in all juices. Recently, Gao et al. [98] studied the effect of cranberry juice consumption (240 and 480 mL of juice, and 480 mL of placebo) on oxidative stress biomarkers and patients affected by H. pylori infection. The blood results indicated that cranberry juice remodeled the gut microbiota of the hospitalized patients, since the juice decreased the growth of H. pylori. Furthermore, Jahan et al. [99] revealed the opportunity to use pongamol extract, a flavonoid derivative present in Pongamia pinnata and Tephrosia purea against H. pylori, since this herbal exhibits diverse pharmacological activities, and thus, pongamol extract can be considered as an herbal medicine. Mishra et al. [100] demonstrated the inhibition of H. pylori infection through rosmaric acid of Asian Basil (Ocimum spp.), and other studies mentioned a relationship between H. pylori and the consumption of vegetables, since a lifestyle without dietary vegetables such as sprouted peas, Chinese chive (Allium tuberosum), and olive oil increase the susceptibility to contract H. pylori infection [39,101,102]. Table 2 shows the most relevant studies on juices, plant extracts, and certain polyphenols with anti-H. pylori properties.
5. Impact of Polyphenols on the Composition of Gut Microbiota
The gut microbiota is a complex microbial ecosystem that interacts with the human host. Gut microbiota plays an important role in nutrient regulation and various metabolic pathways and in maintaining the immune system and general health condition of the host [103]. The age, diet as well as the use of antibiotics and consumption of pre-and probiotics can affect the gut microbiota composition. Increasing evidence strongly suggests that besides dietary fibers, other foods like polyphenols are also considered as prebiotics [104].
In the last decade, it has been established that some polyphenols influence the gut microbiota [105], inhibiting the growth of some bacteria, while other bacteria can become activated [106]. Therefore, Morais et al. [107] reported that the consumption of polyphenols improved the growth and establishment of the Bifidobacteriaceae and Lactobacillaceae families and inhibited pathogenic bacteria such as E. coli, Clostridium perfringens, and H. pylori. The interaction between polyphenols and the gut microbiota can be approached both from the perspective of how they are metabolized by the microbiota and how they can modulate the microbiota [108,109]. Therefore, evidence indicates that the consumption of polyphenols contributes to maintaining intestinal health through their stimulating effects on the growth of beneficial bacteria and the inhibition of pathogenic bacteria, similar to prebiotics [110].
On the other hand, the consumption of polyphenols in a normal diet is accompanied by other food components that influence the composition of gut microbiota, such as fat and fiber [72]. In this context, Havlik et al. [111] reported a possible beneficial effect on health in the interaction of dietary fibers (pectin, psyllium and resistant maltodextrin) with rutin (a flavonoid), changing the levels of short-chain fatty acids and phenolic acids.
As mentioned above, the composition of the intestinal microbiota influences the health of the host, so that unfavorable changes can allow the colonization of abnormal microorganisms in the intestine that can lead to dysbiosis. This can cause different-nature immune and inflammatory responses, low-grade inflammation, weight gain, adiposity, and other metabolic problems [21,107]. Thus, Kemperman et al. [106] reported that black tea polyphenols produced a decrease in the concentration of beneficial Bifidobacteria and an increase in pathogenic bacteria such as Klebsiella, a change that slowed the host’s overall metabolism. However, most researchers show that the consumption of polyphenols causes favorable changes in the gut microbiota, increasing the acidolactic bacteria mentioned above. This modulation of the composition of the gut microbiota produces an increase in the composition of short-chain fatty acids, a decrease in obesity, inflammation, and the metabolism of adipogenesis and lipogenesis [72].
In general, it is complex to determine the direct effects of polyphenols on the intestinal microbiota, since they are highly variable depending on the source, chemical nature, and dose of the polyphenol [110]. However, Ma and Chen [51] recommended a daily intake of 396 mg of polyphenols for optimal human health. From a scientific point of view, it is not yet possible to report the precise mechanisms by which polyphenols modulate the intestinal microbiota, mainly due to the enormous chemical diversity of polyphenolic compounds [108,109].
6. Concluding Remarks and Future Perspectives
H. pilory infection is one of the main causes of gastric cancer and other diseases, and it has become increasingly vital research to treat due to antibiotic resistance. Consequently, polyphenols may contribute to the gastrointestinal health, since these bioactive compounds can prevent the proliferation of H. pylori. Also, polyphenols can modulate the gut microbiota and decrease oxidative stress, and thus, polyphenols improve the body’s immune system. The microbiota impacts the biotransformation of polyphenols into low-molecular-weight, allowing a high absorption, transport and delivery to the epithelium, ending in a correct distribution in the bloodstream, tissues, and brain, of the host. Thus, a stable synergy between the microbiota and polyphenols is essential to maintain the balance. Non-traditional therapies are required to address the pathogens antibiotic resistance, especially for H. pilory infection. Therefore, it is essential to continue the evaluation of dietary polyphenols from natural sources as a new and novel approach, especially endemic berries fruits with high amount of polyphenols content and antioxidant activity such as blueberry, calafate, murta, and maqui, among others, allowing to improve the microbiota and immune response against infections. Many studies have focused on a polyphenol or a mixture of a number of bioactive compounds, with anti-H. pylori properties. Thereby, the bioactive components of fruits, vegetables, and plants (stems, leaves, seeds, etc.), should be isolated, identified, and tested separately to understand the action mechanism in the human body as well in pathogens, and in turn, these components could be used as a new and safe product, and novel alternative therapies to eradicate the H. pylori infection.
Author Contributions
Conceptualization, M.G.-V., P.O.-P. and G.P.; writing—original draft preparation, M.G.-V.; writing—review and editing, P.O.-P. and G.P.; supervision, P.O.-P. and G.P.; project administration, P.O.-P. and G.P.; funding acquisition, P.O.-P. and G.P. All authors have read and agreed to the published version of the manuscript.
Funding
Patricio Orellana-Palma acknowledges the financial support of ANID-Chile (Agencia Nacional de Investigación y Desarrollo de Chile) through the FONDECYT Postdoctoral Grant 2019 (Folio 3190420).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge to Renata Guerra for the figure design. M.G.-V. thanks at the Universidad del Bío-Bío for the doctoral scholarship (2018–2022) and for the “Beca de Investigación”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ghobadi, E.; Ghanbarimasir, Z.; Emami, S. A review on the structures and biological activities of anti-Helicobacter pylori agents. Eur. J. Med. Chem. 2021, 223, 113669. [Google Scholar] [CrossRef]
- Baltas, N.; Karaoglu, S.A.; Tarakci, C.; Kolayli, S. Effect of propolis in gastric disorders: Inhibition studies on the growth of Helicobacter pylori and production of its urease. J. Enzyme Inhib. Med. Chem. 2016, 31, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Baranauskienė, L.; Matulis, D. Overview of human carbonic anhydrases. In Carbonic Anhydrase as Drug Target, 1st ed.; Matulis, D., Ed.; Springer: Cham, Switzerland, 2019; pp. 3–14. [Google Scholar]
- Howitt, M.R.; Lee, J.Y.; Lertsethtakarn, P.; Vogelmann, R.; Joubert, L.M.; Ottemann, K.M.; Amieva, M.R. ChePep controls Helicobacter pylori infection of the gastric glands and chemotaxis in the Epsilonproteobacteria. MBio 2011, 2, e00098-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolig, A.S.; Shanks, J.; Carter, J.E.; Ottemann, K.M. Helicobacter pylori requires TlpD-driven chemotaxis to proliferate in the antrum. Infect. Immun. 2012, 80, 3713–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amieva, M.; Peek, R.M. Pathobiology of Helicobacter pylori–induced gastric cancer. Gastroenterology 2016, 150, 64–78. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.S.; Ottemann, K.M. Colonization, localization, and inflammation: The roles of H. pylori chemotaxis in vivo. Curr. Opin. Microbiol. 2018, 41, 51–57. [Google Scholar] [CrossRef]
- Keilberg, D.; Ottemann, K.M. How Helicobacter pylori senses, targets and interacts with the gastric epithelium. Environ. Microbiol. 2016, 18, 791–806. [Google Scholar] [CrossRef]
- McCracken, K.W.; Aihara, E.; Martin, B.; Crawford, C.M.; Broda, T.; Treguier, J.; Wells, J.M. Wnt/β-catenin promotes gastric fundus specification in mice and humans. Nature 2017, 541, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garza-González, E.; Perez-Perez, G.I.; Maldonado-Garza, H.J.; Bosques-Padilla, F.J. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J. Gastroenterol. 2014, 20, 1438–1449. [Google Scholar] [CrossRef]
- Suzuki, H.; Matsuzaki, J. Gastric cancer: Evidence boosts Helicobacter pylori eradication. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 458–460. [Google Scholar] [CrossRef]
- Sjomina, O.; Pavlova, J.; Niv, Y.; Leja, M. Epidemiology of Helicobacter pylori infection. Helicobacter 2018, 23, e12514. [Google Scholar] [CrossRef] [Green Version]
- Ghasemian, A.; Fattahi, A.; Shokouhi Mostafavi, S.K.; Almarzoqi, A.H.; Memariani, M.; Ben Braiek, O.; Mirforughi, S.A. Herbal medicine as an auspicious therapeutic approach for the eradication of Helicobacter pylori infection: A concise review. J. Cell. Physiol. 2019, 234, 16847–16860. [Google Scholar] [CrossRef]
- Parreira, P.; Soares, B.I.; Freire, C.S.; Silvestre, A.J.; Reis, C.A.; Martins, M.C.L.; Duarte, M.F. Eucalyptus spp. outer bark extracts inhibit Helicobacter pylori growth: In vitro studies. Ind. Crops Prod. 2017, 105, 207–214. [Google Scholar] [CrossRef]
- Singh, R.; Sripada, L.; Singh, R. Side effects of antibiotics during bacterial infection: Mitochondria, the main target in host cell. Mitochondrion 2014, 16, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Qian, W.; Qin, Y.Y.; He, J.; Zhou, Y.H. Probiotics in Helicobacter pylori eradication therapy: A systematic review and meta-analysis. World J. Gastroenterol. 2015, 21, 4345–4357. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; Cirmi, S.; Navarra, M. Effectiveness of citrus fruits on Helicobacter pylori. Evid. Based Complement. Altern. Med. 2017, 2017, 379262. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.Y. Beneficial effects of dietary polyphenols on high-fat diet-induced obesity linking with modulation of gut microbiota. J. Agric. Food Chem. 2019, 68, 33–47. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Oreopoulou, V. Classification of phenolic compounds in plants. In Polyphenols in Plants: Isolation, Purification and Extract Preparation, 2nd ed.; Watson, R.R., Ed.; Academic Press: New York, NY, USA, 2019; pp. 263–281. [Google Scholar]
- Aravind, S.M.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of dietary polyphenols on gut microbiota, their metabolites and health benefits. Food Res. Int. 2021, 142, 110189. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Rocha, S.; de Freitas, V. Going “Green” in the prevention and management of atherothrombotic diseases: The role of dietary polyphenols. J. Clin. Med. 2021, 10, 1490. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Sureda, A.; Orhan, I.E.; Battino, M.; Arnone, A.; Jiménez-García, M.; Capó, X.; Cabot, J.; Sanadgol, N.; Giampieri, F.; et al. The neuroprotective effects of polyphenols, their role in innate immunity and the interplay with the microbiota. Neurosci. Biobehav. Rev. 2021, 128, 437–453. [Google Scholar] [CrossRef]
- Prabhu, S.; Molath, A.; Choksi, H.; Kumar, S.; Mehra, R. Classifications of polyphenols and their potential application in human health and diseases. Int. J. Physiol. Nutr. Phys. Educ. 2021, 6, 293–301. [Google Scholar] [CrossRef]
- Zhang, L.; Han, Z.; Granato, D. Polyphenols in foods: Classification, methods of identification, and nutritional aspects in human health. In Advances in Food and Nutrition Research Series 98: Application of Polyphenols in Foods and Food Models, 1st ed.; Granato, D., Ed.; Academic Press: New York, NY, USA, 2021; pp. 1–33. [Google Scholar]
- Kaur, K.; Asthir, B.; Verma, D.K. Biosynthesis, bioavailability, and metabolism of plant polyphenols: Biological activities and their potential benefits in human health. In Phytochemicals in Food and Health: Perspectives for Research and Technological Development, 1st ed.; Verma, D.K., Thakur, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 233–258. [Google Scholar]
- Huang, R.; Xu, C. An overview of the perception and mitigation of astringency associated with phenolic compounds. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1036–1074. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yang, H.; Capanoglu, E.; Cao, H.; Xiao, J. Technological aspects and stability of polyphenols. In Polyphenols: Properties, Recovery, and Applications, 1st ed.; Galanakis, C.M., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 295–323. [Google Scholar]
- Guerra-Valle, M.; Lillo-Pérez, S.; Petzold, G.; Orellana-Palma, P. Effect of freeze crystallization on quality properties of two endemic Patagonian berries juices: Murta (Ugni molinae) and arrayan (Luma apiculata). Foods 2021, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Palma, P.; Tobar-Bolaños, G.; Casas-Forero, N.; Zúñiga, R.N.; Petzold, G. Quality attributes of cryoconcentrated calafate (Berberis microphylla) juice during refrigerated storage. Foods 2020, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-mediated gut microbiota modulation: Toward prebiotics and further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, M.; Sundralingam, U.; Palanisamy, U.D. Polyphenols as prebiotics in the management of high-fat diet-induced obesity: A systematic review of animal studies. Foods 2021, 10, 299. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the future of probiotics and prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Fruit polyphenols: A review of anti-inflammatory effects in humans. Crit. Rev. Food Sci. Nutr. 2016, 56, 419–444. [Google Scholar] [CrossRef]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef]
- Shaw, P.; Chattopadhyay, A. Nrf2–ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Mayol, I.; Guerra-Valle, M.; Casas-Forero, N.; Sobral, M.M.C.; Viegas, O.; Alarcón-Enos, J.; Pinho, O. Western dietary pattern antioxidant intakes and oxidative stress: Importance during the SARS-CoV-2/COVID-19 Pandemic. Adv. Nutr. 2021, 12, 670–681. [Google Scholar] [CrossRef]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.N.; Tang, G.Y.; Li, S.; Li, H.B. Natural products for the prevention and management of Helicobacter pylori infection. Compr. Rev. Food Sci. Food Saf. 2018, 17, 937–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.E.; Khoi, P.N.; Xia, Y.; Park, J.S.; Joo, Y.E.; Kim, K.K.; Choi, S.Y.; Jung, Y.D. Helicobacter pylori and interleukin-8 in gastric cancer. World J. Gastroenterol. 2013, 45, 8192–8202. [Google Scholar] [CrossRef] [PubMed]
- Keates, S.; Hitti, Y.S.; Upton, M.; Kelly, C.P. Helicobacter pylori infection activates NF-kappa B in gastric epithelial cells. Gastroenterology 1997, 113, 1099–1109. [Google Scholar] [CrossRef]
- Seo, J.H.; Lim, J.W.; Kim, H.; Kim, K.H. Helicobacter pylori in a Korean isolate activates mitogen-activated protein kinases, AP-1, and NF-kB and induces chemokine expression in gastric epithelial AGS cells. Lab. Investig. 2003, 84, 49–62. [Google Scholar] [CrossRef]
- Song, M.Y.; Lee, D.Y.; Kim, E.H. Anti-inflammatory and anti-oxidative effect of Korean propolis on Helicobacter pylori-induced gastric damage in vitro. J. Microbiol. 2020, 58, 878–885. [Google Scholar] [CrossRef]
- Tominaga, K.; Kawahara, T.; Sano, T.; Toida, K.; Kuwano, Y.; Sasaki, H.; Kawai, T.; Teshima-Kondo, S.; Rokutan, K. Evidence for cancer-associated expression of NADPH oxidase 1 (Nox1)-based oxidase system in the human stomach. Free Radic. Biol. Med. 2007, 43, 1627–1638. [Google Scholar] [CrossRef]
- Khan, H.; Sureda, A.; Belwal, T.; Çetinkaya, S.; Süntar, İ.; Tejada, S.; Ullah, H.; Devkota, H.P.; Aschner, M.; Aschner, M. Polyphenols in the treatment of autoimmune diseases. Autoimmun. Rev. 2019, 18, 647–657. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Castilho, P.C.M.F.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Sanches-Silva, A.; et al. Targeting NF-κB signaling pathway in cancer by dietary polyphenols. Crit. Rev. Food Sci. Nutr. 2020, 60, 2790–2800. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Rolt, A.; Cox, L.S. Structural basis of the anti-ageing effects of polyphenolics: Mitigation of oxidative stress. BMC Chem. 2020, 14, 1–13. [Google Scholar] [CrossRef]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Flavonoids in inflammatory bowel disease: A review. Nutrients 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyzowska, A. Plant extracts rich in polyphenols: Antibacterial agents and natural preservatives for meat and meat products. Crit. Rev. Food Sci. Nutr. 2021, 61, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Chen, Y. Polyphenol supplementation benefits human health via gut microbiota: A systematic review via meta-analysis. J. Funct. Foods 2020, 66, 103829. [Google Scholar] [CrossRef]
- Burkholder-Cooley, N.; Rajaram, S.; Haddad, E.; Fraser, G.E.; Jaceldo-Siegl, K. Comparison of polyphenol intakes according to distinct dietary patterns and food sources in the Adventist Health Study-2 cohort. Br. J. Nutr. 2016, 115, 2162–2169. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Goñi, I. Antioxidant capacity of the Spanish Mediterranean diet. Food Chem. 2006, 94, 442–447. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Touillaud, M. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Huang, D. Dietary antioxidants and health promotion. Antioxidants 2018, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Tsugawa, H.; Suzuki, H. Oxidative stress in stomach cancer. In Cancer: Oxidative Stress and Dietary Antioxidants, 2nd ed.; Preedy, V.R., Patel, V.B., Eds.; Academic Press: New York, NY, USA, 2021; pp. 49–54. [Google Scholar]
- Capparelli, R.; Iannelli, D. Genetics of host protection against Helicobacter pylori infections. Int. J. Mol. Sci. 2021, 22, 3192. [Google Scholar] [CrossRef]
- Jain, U.; Saxena, K.; Chauhan, N. Helicobacter pylori induced reactive oxygen species: A new and developing platform for detection. Helicobacter 2021, 26, e12796. [Google Scholar] [CrossRef] [PubMed]
- Öztekin, M.; Yilmaz, B.; Ağagündüz, D.; Capasso, R. Overview of Helicobacter pylori Infection: Clinical Features, treatment, and nutritional aspects. Diseases 2021, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Judaki, A.; Rahmani, A.; Feizi, J.; Asadollahi, K.; Hafezi Ahmadi, M.R. Curcumin in combination with triple therapy regimes ameliorates oxidative stress and histopathologic changes in chronic gastritis-associated Helicobacter pylori infection. Arq. Gastroenterol. 2017, 54, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N.L.; Shabib, S.; Sherman, P.M. Capsaicin as an inhibitor of the growth of the gastric pathogen Helicobacter pylori. FEMS Microbiol. Lett. 1997, 146, 223–227. [Google Scholar] [CrossRef]
- Lee, I.O.; Lee, K.H.; Pyo, J.H.; Kim, J.H.; Choi, Y.J.; Lee, Y.C. Anti-inflammatory effect of capsaicin in Helicobacter pylori-infected gastric epithelial cells. Helicobacter 2007, 12, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, Y.; Yamamoto, Y.; Hara, Y.; Shimamura, T. A combination effect of epigallocatechin gallate, a major compound of green tea catechins, with antibiotics on Helicobacter pylori growth in vitro. Curr. Microbiol. 2003, 47, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.F.; Pogačnik, L. Polyphenols from food and natural products: Neuroprotection and safety. Antioxidants 2020, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.; Perez-Gregorio, R.; Mateus, N.; de Freitas, V. Interactions of dietary polyphenols with epithelial lipids: Advances from membrane and cell models in the study of polyphenol absorption, transport and delivery to the epithelium. Crit. Rev. Food Sci. Nutr. 2021, 61, 3007–3030. [Google Scholar] [CrossRef]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between polyphenols and microorganisms, especially gut microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Martínez, I.; Wallace, G.; Zhang, C.; Legge, R.; Benson, A.K.; Carr, T.P.; Walter, J. Diet-induced metabolic improvements in a hamster model of hypercholesterolemia are strongly linked to alterations of the gut microbiota. Appl. Environ. Microbiol. 2009, 75, 4175–4184. [Google Scholar] [CrossRef] [Green Version]
- Hervert-Hernández, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Filosa, S.; di Meo, F.; Crispi, S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen. Res. 2018, 13, 154–169. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci. Technol. 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Bartolomé, B. A survey of modulation of gut microbiota by dietary polyphenols. BioMed Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Kumar-Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar-Rana, H.; Gupta, A.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Dey, P. Gut microbiota in phytopharmacology: A comprehensive overview of concepts, reciprocal interactions, biotransformations and mode of actions. Pharmacol. Res. 2019, 147, 104367. [Google Scholar] [CrossRef] [PubMed]
- Danneskiold-Samsøe, N.B.; Barros, H.D.D.F.Q.; Santos, R.; Bicas, J.L.; Cazarin, C.B.B.; Madsen, L.; Junior, M.R.M. Interplay between food and gut microbiota in health and disease. Food Res. Int. 2019, 115, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The modulatory effect of polyphenols from green tea, oolong tea and black tea on human intestinal microbiota in vitro. J. Food Sci. Technol. 2018, 55, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Milutinović, M.; Dimitrijević-Branković, S.; Rajilić-Stojanović, M. Plant extracts rich in polyphenols as potent modulators in the growth of probiotic and pathogenic intestinal microorganisms. Front. Nutr. 2021, 8, 688843. [Google Scholar] [CrossRef]
- Chiu, H.F.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C.K. Gastroprotective Effects of polyphenols against various gastro-intestinal disorders: A mini-review with special focus on clinical evidence. Molecules 2021, 26, 2090. [Google Scholar] [CrossRef]
- Mandici, A.; Covrig, A.A.; Macovei, I. Food phytochemicals against Helicobacter pylori. Rom. J. Pharm. Pract. 2021, 14, 11–17. [Google Scholar] [CrossRef]
- Chung, J.G. Inhibitory actions of ellagic acid on growth and arylamine N-acetyltransferase activity in strains of Helicobacter pylori from peptic ulcer patients. Microbios 1998, 93, 115–127. [Google Scholar]
- Martini, S.; d’Addario, C.; Colacevich, A.; Focardi, S.; Borghini, F.; Santucci, A.; Figura, N.; Rossi, C. Antimicrobial activity against Helicobacter pylori strains and antioxidant properties of blackberry leaves (Rubus ulmifolius) and isolated compounds. Int. J. Antimicrob. Agents 2009, 34, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jiang, A.; Qi, B.; Ma, Z.; Xiong, Y.; Dou, J.; Wang, J. Resveratrol protects against Helicobacter pylori-associated gastritis by combating oxidative stress. Int. J. Mol. Sci. 2015, 16, 27757–27769. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Yeo, M.; Choue, J.S.; Jin, J.H.; Park, S.J.; Cheong, J.Y.; Lee, K.J.; Kim, J.H.; Hahm, K.B. Protective mechanism of epigallocatechin-3-gallate against Helicobacter pylori-induced gastric epithelial cytotoxicity via the blockage of TLR-4 signaling. Helicobacter 2004, 9, 632–642. [Google Scholar] [CrossRef]
- Takeuchi, H.; Trang, V.T.; Morimoto, N.; Nishida, Y.; Matsumura, Y.; Sugiura, T. Natural products and food components with anti-Helicobacter pylori activities. World J. Gastroenterol. 2014, 20, 8971–8978. [Google Scholar] [CrossRef] [PubMed]
- Escandón, R.A.; del Campo, M.; López-Solis, R.; Obreque-Slier, E.; Toledo, H. Antibacterial effect of kaempferol and (-)-epicatechin on Helicobacter pylori. Eur. Food Res. Technol. 2016, 242, 1495–1502. [Google Scholar] [CrossRef]
- Chua, C.S.; Yang, K.C.; Chen, J.H.; Liu, Y.H.; Hsu, Y.H.; Lee, H.C.; Huang, S.Y. The efficacy of blueberry and grape seed extract combination on triple therapy for Helicobacter pylori eradication: A randomized controlled trial. Int. J. Food Sci. Nutr. 2016, 67, 177–183. [Google Scholar] [CrossRef]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Foods 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Park, J.U.; Cho, J.S.; Kim, J.S.; Kim, H.K.; Jo, Y.H.; Rahman, M.A.A.; Lee, Y.I. Synergistic effect of Rubus crataegifolius and Ulmus macrocarpa against Helicobacter pylori clinical isolates and gastritis. Front. Pharmacol. 2020, 11, 4. [Google Scholar] [CrossRef]
- Torres, E.; García, A.; Aranda, M.; Saéz, V.; Zúñiga, F.; Alarcón, J.; Pastene, E. One-step purification of two semi-synthetic epicatechin adducts prepared from avocado peels procyanidins by centrifugal partition chromatography and evaluation of their anti-inflammatory effects on adenocarcinoma gastric cells infected with Helicobacter pylori. J. Chil. Chem. Soc. 2018, 63, 4222–4228. [Google Scholar] [CrossRef] [Green Version]
- Spósito, L.; Oda, F.B.; Vieira, J.H.; Carvalho, F.A.; dos Santos Ramos, M.A.; de Castro, R.C.; Chorilli, M. In vitro and in vivo anti-Helicobacter pylori activity of Casearia sylvestris leaf derivatives. J. Ethnopharmacol. 2019, 233, 1–12. [Google Scholar] [CrossRef]
- Betoret, E.; Betoret, N.; Arilla, A.; Bennár, M.; Barrera, C.; Codoñer, P.; Fito, P. No invasive methodology to produce a probiotic low humid apple snack with potential effect against Helicobacter pylori. J. Food Eng. 2012, 110, 289–293. [Google Scholar] [CrossRef]
- Barrera, C.; Burca, C.; Betoret, E.; García-Hernández, J.; Hernández, M.; Betoret, N. Improving antioxidant properties and probiotic effect of clementine juice inoculated with Lactobacillus salivarius spp. salivarius (CECT 4063) by trehalose addition and/or sublethal homogenisation. Int. J. Food Sci. Technol. 2019, 54, 2109–2122. [Google Scholar] [CrossRef]
- Gao, T.; Hou, M.; Zhang, B.; Pan, X.; Liu, C.; Sun, C.; Jia, M.; Lin, S.; Xiong, K.; Ma, A. Effects of cranberry beverage on oxidative stress and gut microbiota in subjects with Helicobacter pylori infection: A randomized, double-blind, placebo-controlled trial. Food Funct. 2021, 12, 6878–6888. [Google Scholar] [CrossRef]
- Jahan, S.; Mahmud, M.H.; Khan, Z.; Alam, A.; Khalil, A.A.; Rauf, A.; Tareq, A.M.; Nainu, F.; Tareq, S.M.; Emran, B.T.; et al. Health promoting benefits of pongamol: An overview. Biomed. Pharmacother. 2021, 142, 112109. [Google Scholar] [CrossRef] [PubMed]
- Mishra, L.K.; Sarkar, D.; Mentreddy, R.; Shetty, K. Evaluation of phenolic bioactive-linked anti-hyperglycemic and Helicobacter pylori inhibitory activities of Asian Basil (Ocimum spp.) varieties. J. Herb. Med. 2020, 20, 100310. [Google Scholar] [CrossRef]
- Amaral, O.; Fernandes, I.; Veiga, N.; Pereira, C.; Chaves, C.; Nelas, P.; Silva, D. Living conditions and Helicobacter pylori in adults. Biomed Res. Int. 2017, 2017, 9082716. [Google Scholar] [CrossRef] [Green Version]
- Mard, S.A.; Haghighian, H.K.; Sebghatulahi, V.; Ahmadi, B. Dietary factors in relation to Helicobacter pylori infection. Gastroenterol. Res. Pract. 2014, 2014, 826910. [Google Scholar] [CrossRef] [Green Version]
- Ashaolu, T.J. Immune boosting functional foods and their mechanisms: A critical evaluation of probiotics and prebiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef]
- Fatima, N.; Akhtar, T.; Sheikh, N. Prebiotics: A novel approach to treat hepatocellular carcinoma. Can. J. Gastroenterol. Hepatol. 2017, 2017, 6238106. [Google Scholar] [CrossRef]
- Pathak, S.; Kesavan, P.; Banerjee, A.; Banerjee, A.; Celep, G.S.; Bissi, L.; Marotta, F. Metabolism of dietary polyphenols by human gut microbiota and their health benefits. In Polyphenols: Mechanisms of Action in Human Health and Disease, 2nd ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: New York, NY, USA, 2018; pp. 347–359. [Google Scholar]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [Green Version]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Vamanu, E.; Gatea, F. Correlations between microbiota bioactivity and bioavailability of functional compounds: A mini-review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The two way polyphenols-microbiota interactions and their effects on obesity and related metabolic diseases. Front. Nutr. 2019, 20, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashwin, K.; Pattanaik, A.K.; Howarth, G.S. Polyphenolic bioactives as an emerging group of nutraceuticals for promotion of gut health: A review. Food Biosci. 2021, 44, 101376. [Google Scholar] [CrossRef]
- Havlik, J.; Marinello, V.; Gardyne, A.; Hou, M.; Mullen, W.; Morrison, D.J.; Preston, T.; Combet, E.; Edwards, C.A. Dietary fibres differentially impact on the production of phenolic acids from rutin in an in vitro fermentation model of the human gut microbiota. Nutrients 2020, 12, 1577. [Google Scholar] [CrossRef]
Figure 1.
Helicobacter pylori colonization of the human stomach.

Figure 2.
Helicobacter pylori colonization of the human stomach.


{kind=link}
{kind=link}
Table 1.
Biotransformation of polyphenols by gut microbiota and main metabolites (Adapted from Marin et al. [76]).
Table 1.
Biotransformation of polyphenols by gut microbiota and main metabolites (Adapted from Marin et al. [76]).
| Polyphenol Group | Compound | Gut Microbiome | Metabolites |
|---|---|---|---|
| Anthocyanins | Cyanidin | Bifidobacterium lactis BB-12 | 3,4-Dihydroxybenzoic Acid |
| Malvidin | Lactobacillus acidophilus LA-5 | 3,4-Dimethocybenzoic Acid | |
| Ponidin Pelargonidin | Lactobacillus casei Lactobacillus plantarum | 3-Methoxy-4-Hydroxybenzoic Acid 4-Hydroxybenzoic Acid | |
| Flavan-3-ols | Catechin | Clostridium cocoides | 3-(3-Hydroxyphenyl)-Propionic Acid |
| Epicatechin Epigallocatechin | Bifidobacterium spp. | 5-(3′,4′-Dihydroxyphenil)-Γ-Valerolactone 5-(3,4-Dihydroxyphenyl)-Valeric Acid 3-(3,4-Dihydroxyphenyl)-Propionic Acid | |
| 5-(3′,4′-Dihydroxyphenyl)-Γ-Valerolactone 5-(3′,5′-Dihydroxyphenyl)-Γ-Valerolactone | |||
| Flavanones | Naringenin | Clostridum Strains | 3-(4-hydroxyphenyl)-propionic acid |
| Eubacterium ramulus | |||
| Flavones | Luteolin | Clostridium orbiscindens | 3-(3,4-Dihydroxyphenyl)-Propionic Acid |
| Apigenin | Enterococcus avium | 3-(4-Dihydroxyphenyl)-Propionic Acid 3-(3-Dihydroxyphenyl)-Propionic Acid 4-Hydroxycinnamic Acid | |
| Flavonols | Kaempferol | Clostridium orbiscindens | 2-(4-Hydroxyphenyl)-Propionic Acid |
| Quercetin Myricetin | Clostridium orbiscindens Eubacterium oxidoreducens Eubacterium ramulus Enterococcus casseliflavus Clostridium orbiscindens Eubacterium oxidoreducens | 2-(3,4-Dihydroxyphenyl)-Acetic Acid 2-(3-Hydroxyphenyl)-Acetic Acid 3-(3,4-Dihydroxyphenyl)-Propionic Acid 3-(3-Hydroxyphenyl)-Acetic Acid 2-(3,5-Dihydrosyphenyl)-Acetic Acid 2-(3-Hydroxyphenyl)-Acetic Acid | |
| Isoflavones | Daidzein | Bacteroides ovatus | (S)-Equol |
| Streptococcus intermedius Rumnococcus products Eggerthella sp. Julong 732 Enterococcus faecium EPI1 Lactobacillus mucosae EPI2 Finegoldia magna EPI3 Clostridium sp. HGHA136 | O-Demethylangolesin |
Table 2.
Juices, plant extracts, and polyphenols with anti-H. pylori effects.
| Sample | Test Type | Anti-Helicobacter pylori Effect | Reference |
|---|---|---|---|
| Bergamot Juice | In vitro | Inhibited the Growth and Reduced the Viability | [17] |
| Citrus Fruit Extract | In vitro | Antimicrobial Activity, Affecting the Urease Activity | [17] |
| 6-Shogaol, 6-Gingerol, 8-Gingerol 10-Gingerol Curcumin Propolis Muscadine Grapes | In vitro | Antibacterial Activity Inhibited the Growth and Damaged Its Cytoplasmic Membrane | [89] |
| Kaempferol And (-)-Epicatechin | Inhibition Halo Test | Suppressed the Growth, also proved the Inhibitory Effect depends on the Pathogen Exposure Time to the Polyphenol | [90] |
| Blueberry and Grape Seed Extract | Standard Triple Therapy Plus | There was no Significant Difference between this Therapy and one Standard Triple Therapy Plus Placebo | [91] |
| Aqueous Extract from Fragaria Vesca Leaves | Antibacterial Activity Evaluation | Revealed an Intense Activity Due to the Source of Ellagitannins | [92] |
| Red Raspberry Elm Tree | In vitro | Inhibited (Alone or in Combination) the Growth of H. Pylori | [93] |
| Avocado | Anti-Inflamatory Activity | Lowered the Early Inflammatory Signals | [94] |
| Caseria Sylvestris Leaves | In vitro and in vivo | Inhibited the Growth of H. Pylori | [95] |
| Low Humidity Apple Snack Impregnated with Mandarin and Pineapple/Grape Juice, and Inoculated with Lactobacillus Salivarius spp. Salivarius | In vivo | No Evidence Suggested Whether Bioactive Compounds Present in the Juices Affected the Eradication Rate | [96] |
| Clementine Juice Inoculated with Lactobacillus Salivarius spp. Salivarius | In vitro | Inhibited the Growth of H. Pylori | [97] |
| Cranberry Juice | Standard Triple Therapy Plus | Inhibited the Growth of H. Pylori | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guerra-Valle, M.; Orellana-Palma, P.; Petzold, G. Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota. Antioxidants 2022, 11, 109. https://doi.org/10.3390/antiox11010109
AMA Style
Guerra-Valle M, Orellana-Palma P, Petzold G. Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota. Antioxidants. 2022; 11(1):109. https://doi.org/10.3390/antiox11010109
Chicago/Turabian StyleGuerra-Valle, María, Patricio Orellana-Palma, and Guillermo Petzold. 2022. "Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota" Antioxidants 11, no. 1: 109. https://doi.org/10.3390/antiox11010109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.