
Dexamethasone Administration in Mice Leads to Less Body Weight Gain over Time, Lower Serum Glucose, and Higher Insulin Levels Independently of NRF2

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Model
2.2. Dexamethasone Treatment of Mice
2.3. Intraperitoneal Glucose Tolerance Test (IPGTT) and Blood Chemistries Measurement
2.4. Micro-CT
2.5. RNA Preparation and Quantitative Real-Time PCR
2.6. Statistics
3. Results
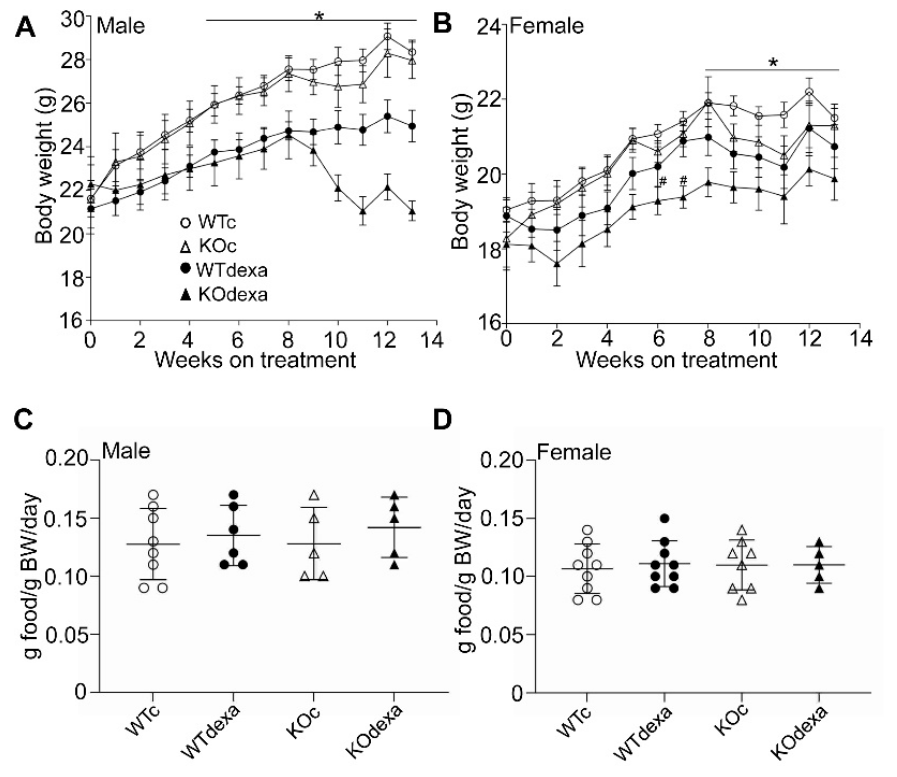
3.1. Chronic Dexamethasone Treatment Led to Less Weight Gain over Time in Male and Female Mice in an Nrf2-Independent Manner
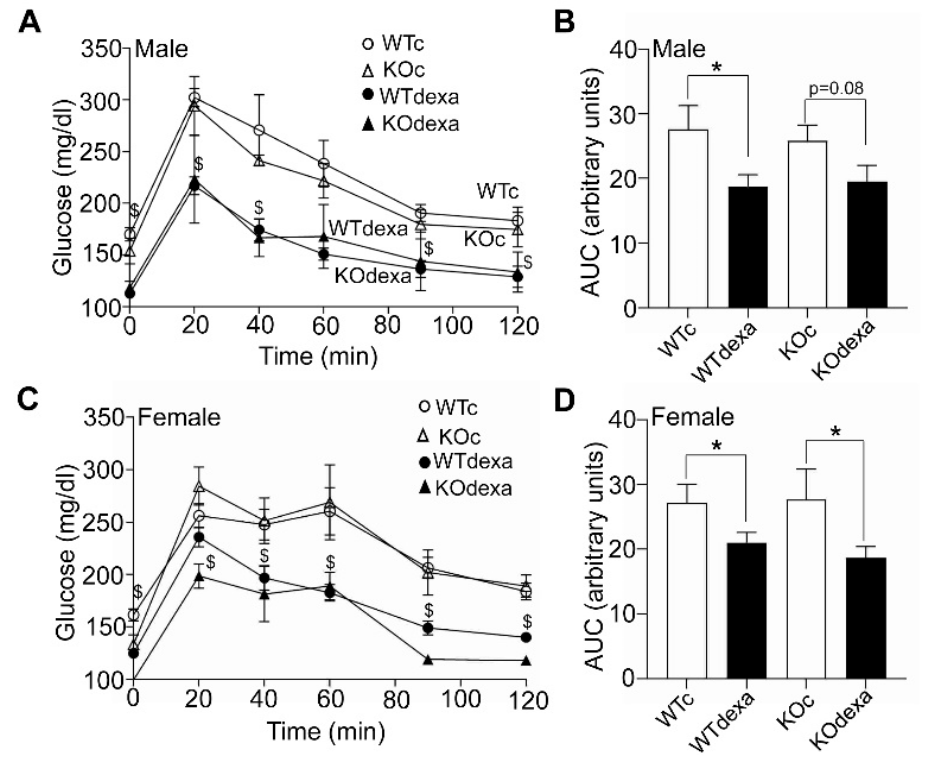
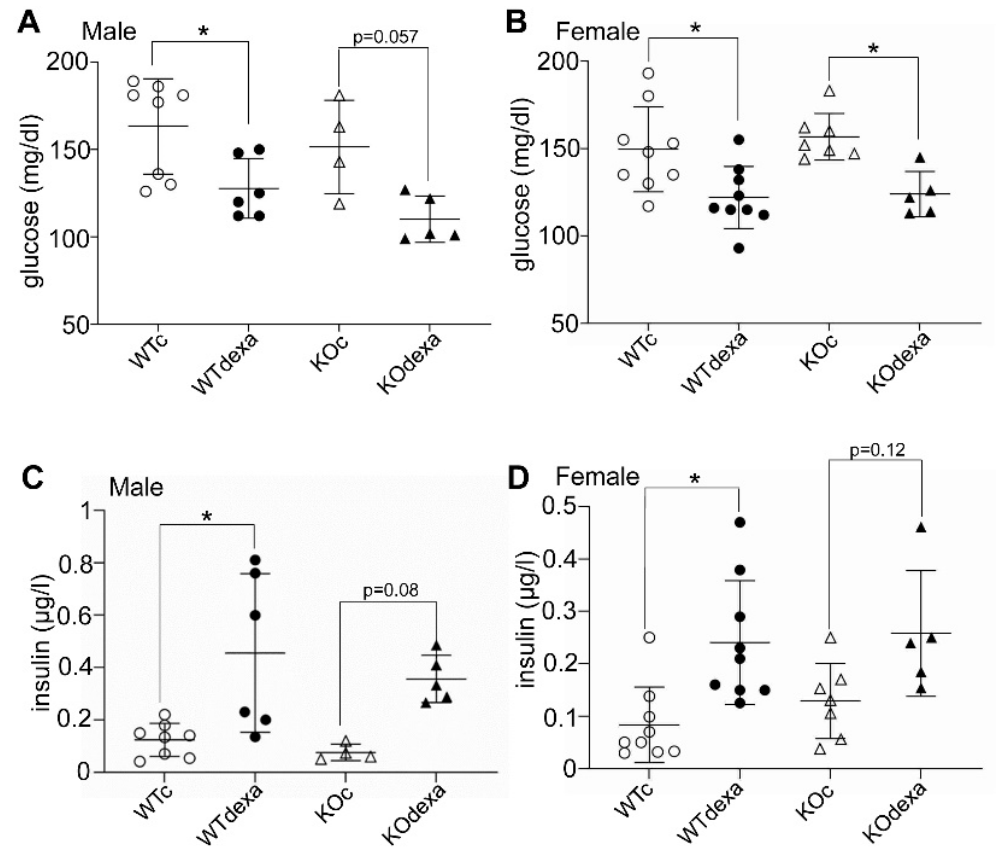
3.2. Dexamethasone-Treated Mice Showed Lower Baseline Blood Glucose Levels and Better Glucose Tolerance along with Higher Insulin Levels Irrespectively of Genotype
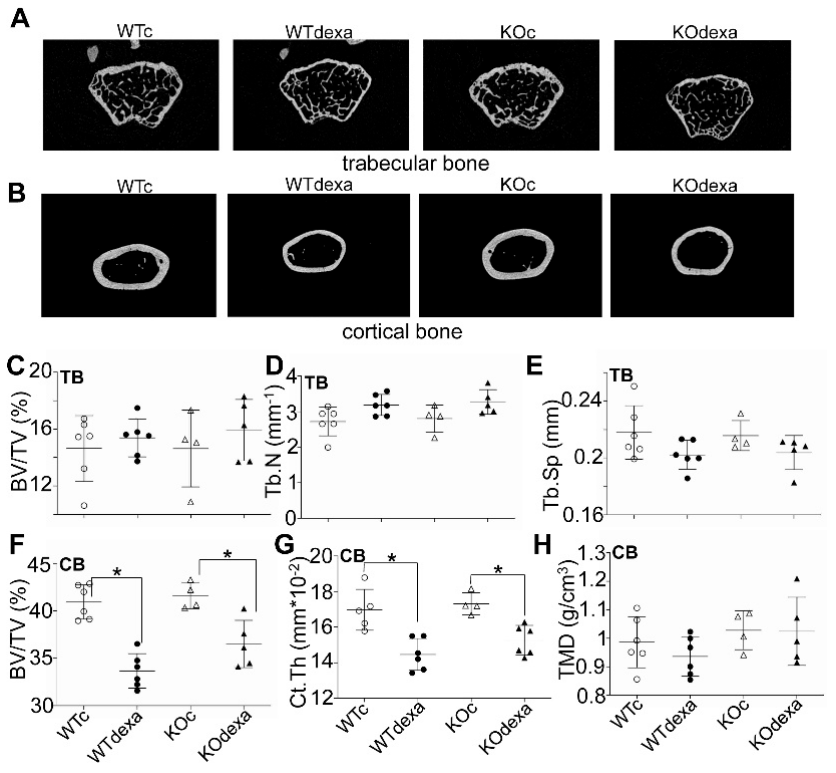
3.3. Dexamethasone Treatment Decreased Cortical Bone Volume in Male Mice Independently of Nrf2
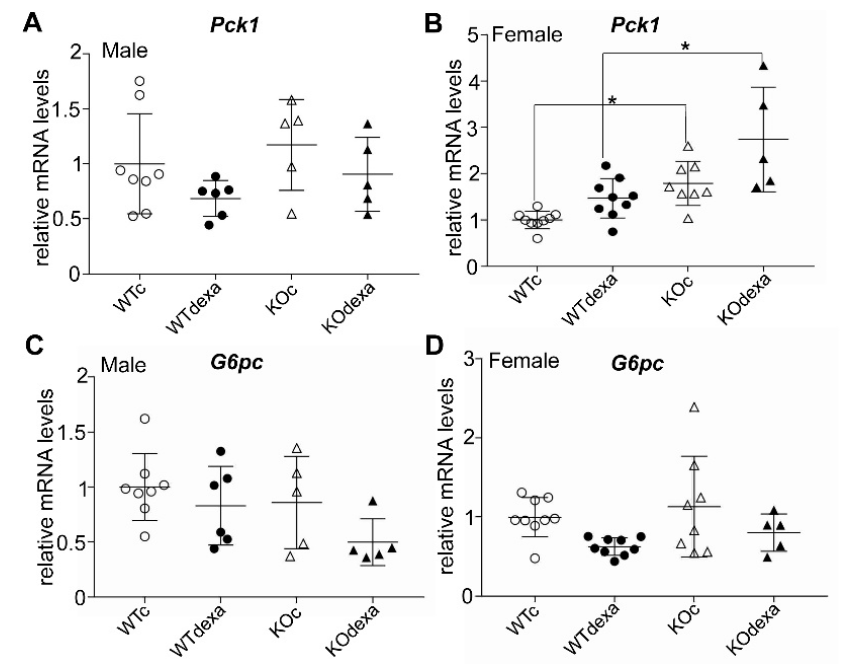
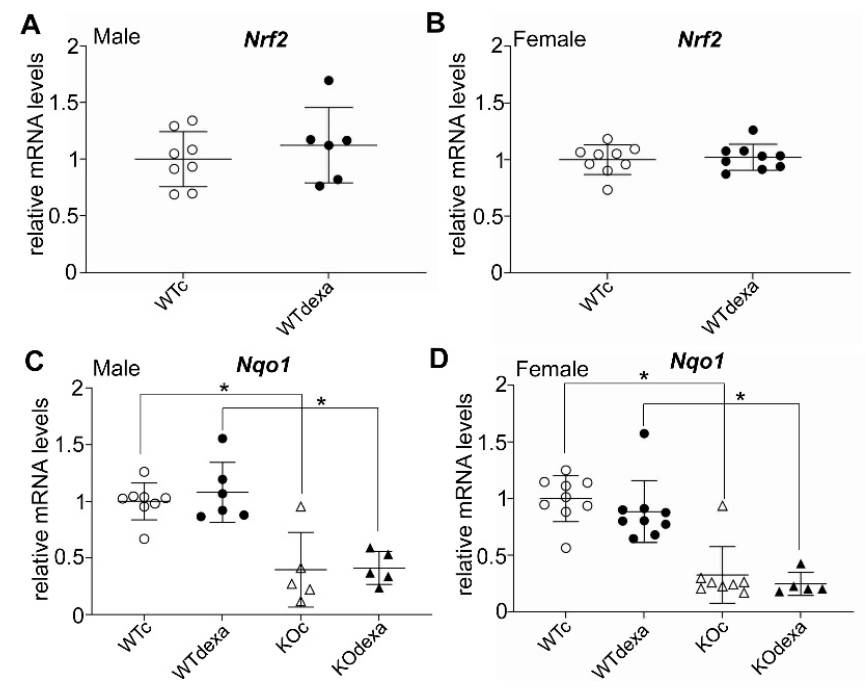
3.4. Chronic Dexamethasone Exposure Does Not Affect the mRNA Expression Levels of Key Gluconeogenic Enzymes and NRF2 Signaling-Related Genes in Liver
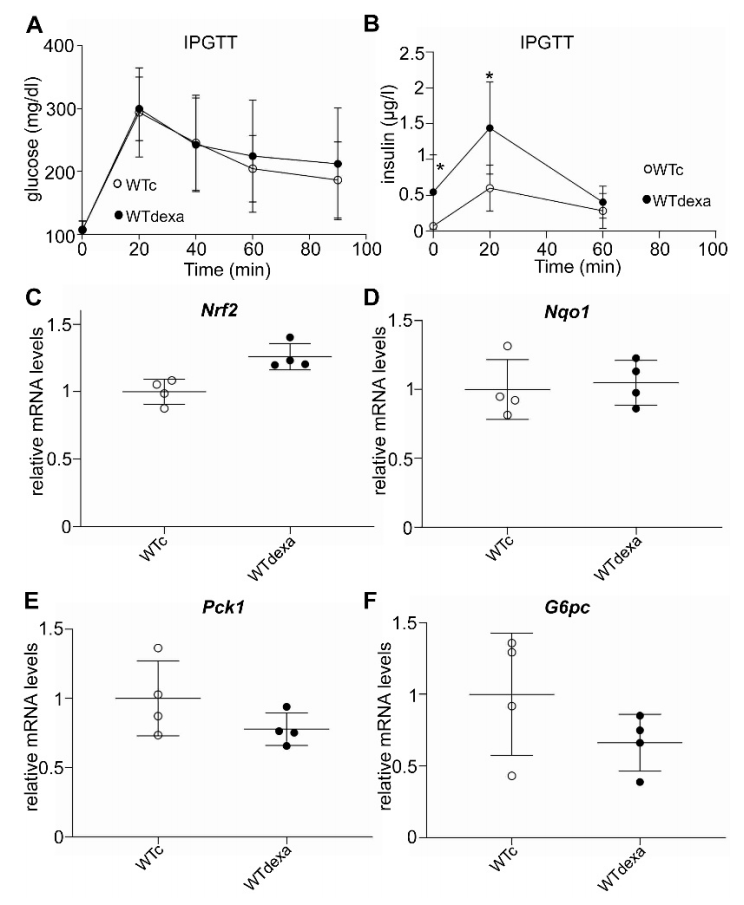
3.5. Acute Dexamethasone Treatment Caused Hyperinsulinemia without Affecting Gluconeogenic Gene Expression or Nrf2 Signaling
4. Discussion
5. Conclusions
- Dexamethasone induced hyperinsulinemia with concomitant low glucose levels independently of NRF2.
- Acute or chronic dexamethasone treatment did not affect NRF2 signaling.
- All the aforementioned effects of dexamethasone were independent of gender.
- Dexamethasone-induced osteoporosis in male mice was independent of NRF2.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Overman, R.A.; Yeh, J.Y.; Deal, C.L. Prevalence of oral glucocorticoid usage in the United States: A general population perspective. Arthritis Care Res. 2013, 65, 294–298. [Google Scholar] [CrossRef]
- Katsuyama, T.; Sada, K.E.; Namba, S.; Watanabe, H.; Katsuyama, E.; Yamanari, T.; Wada, J.; Makino, H. Risk factors for the development of glucocorticoid-induced diabetes mellitus. Diabetes Res. Clin. Pract. 2015, 108, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Compston, J. Management of glucocorticoid-induced osteoporosis. Nat. Rev. Rheumatol. 2010, 6, 82–88. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Robertson, R.P.; Harmon, J.; Tran, P.O.; Tanaka, Y.; Takahashi, H. Glucose toxicity in beta-cells: Type 2 diabetes, good radicals gone bad, and the glutathione connection. Diabetes 2003, 52, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Yagishita, Y.; Fahey, J.W.; Dinkova-Kostova, A.T.; Kensler, T.W. Broccoli or Sulforaphane: Is It the Source or Dose That Matters? Molecules 2019, 24, 3593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Waddington, J.C.; Tailor, A.; Lister, A.; Hamlett, J.; Berry, N.; Park, B.K.; Sporn, M.B. CDDO-imidazolide Targets Multiple Amino Acid Residues on the Nrf2 Adaptor, Keap1. J. Med. Chem. 2020, 63, 9965–9976. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap‘n’collar transcription factors in aging and human disease. Sci. Signal. 2010, 3, re3. [Google Scholar] [CrossRef] [Green Version]
- Yagishita, Y.; Fukutomi, T.; Sugawara, A.; Kawamura, H.; Takahashi, T.; Pi, J.; Uruno, A.; Yamamoto, M. Nrf2 protects pancreatic beta-cells from oxidative and nitrosative stress in diabetic model mice. Diabetes 2014, 63, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Yagishita, Y.; Uruno, A.; Chartoumpekis, D.V.; Kensler, T.W.; Yamamoto, M. Nrf2 represses the onset of type 1 diabetes in non-obese diabetic mice. J. Endocrinol. 2019, 240, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Uruno, A.; Furusawa, Y.; Yagishita, Y.; Fukutomi, T.; Muramatsu, H.; Negishi, T.; Sugawara, A.; Kensler, T.W.; Yamamoto, M. The Keap1-Nrf2 system prevents onset of diabetes mellitus. Mol. Cell. Biol. 2013, 33, 2996–3010. [Google Scholar] [CrossRef] [Green Version]
- Slocum, S.L.; Skoko, J.J.; Wakabayashi, N.; Aja, S.; Yamamoto, M.; Kensler, T.W.; Chartoumpekis, D.V. Keap1/Nrf2 pathway activation leads to a repressed hepatic gluconeogenic and lipogenic program in mice on a high-fat diet. Arch. Biochem. Biophys. 2016, 591, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, I.; Stephenson, E.J.; Redd, J.R.; Tran, Q.T.; Hochberg, I.; Qi, N.; Bridges, D. Glucocorticoid-Induced Metabolic Disturbances Are Exacerbated in Obese Male Mice. Endocrinology 2018, 159, 2275–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uto, A.; Miyashita, K.; Endo, S.; Sato, M.; Ryuzaki, M.; Kinouchi, K.; Mitsuishi, M.; Meguro, S.; Itoh, H. Transient Dexamethasone Loading Induces Prolonged Hyperglycemia in Male Mice With Histone Acetylation in Dpp-4 Promoter. Endocrinology 2021, 162, bqab193. [Google Scholar] [CrossRef]
- Boortz, K.A.; Syring, K.E.; Lee, R.A.; Dai, C.; Oeser, J.K.; McGuinness, O.P.; Wang, J.C.; O’Brien, R.M. G6PC2 Modulates the Effects of Dexamethasone on Fasting Blood Glucose and Glucose Tolerance. Endocrinology 2016, 157, 4133–4145. [Google Scholar] [CrossRef] [Green Version]
- Kusuda, Y.; Kondo, Y.; Miyagi, Y.; Munemasa, T.; Hori, Y.; Aonuma, F.; Tsuka, S.; Mukaibo, T.; Masaki, C.; Hosokawa, R. Long-term dexamethasone treatment diminishes store-operated Ca(2+) entry in salivary acinar cells. Int. J. Oral Sci. 2019, 11, 1. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Shin, S.; Slocum, S.L.; Agoston, E.S.; Wakabayashi, J.; Kwak, M.K.; Misra, V.; Biswal, S.; Yamamoto, M.; Kensler, T.W. Regulation of notch1 signaling by nrf2: Implications for tissue regeneration. Sci. Signal. 2010, 3, ra52. [Google Scholar] [CrossRef] [Green Version]
- Rinotas, V.; Niti, A.; Dacquin, R.; Bonnet, N.; Stolina, M.; Han, C.Y.; Kostenuik, P.; Jurdic, P.; Ferrari, S.; Douni, E. Novel genetic models of osteoporosis by overexpression of human RANKL in transgenic mice. J. Bone Miner. Res. 2014, 29, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, M.; Rinotas, V.; Violitzi, F.; Thireou, T.; Panayotou, G.; Samiotaki, M.; Douni, E. New Insights for RANKL as a Proinflammatory Modulator in Modeled Inflammatory Arthritis. Front. Immunol. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Spandidos, A.; Wang, H.; Seed, B. PrimerBank: A PCR primer database for quantitative gene expression analysis, 2012 update. Nucleic Acids Res. 2012, 40, D1144–D1149. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Okazaki, K.; Nguyen, L.T.T.; Ota, N.; Kitamura, H.; Murakami, S.; Shima, H.; Igarashi, K.; Sekine, H.; Motohashi, H. Glucocorticoid receptor signaling represses the antioxidant response by inhibiting histone acetylation mediated by the transcriptional activator NRF2. J. Biol. Chem. 2017, 292, 7519–7530. [Google Scholar] [CrossRef] [Green Version]
- Cummings, B.P.; Bremer, A.A.; Kieffer, T.J.; D’Alessio, D.; Havel, P.J. Investigation of the mechanisms contributing to the compensatory increase in insulin secretion during dexamethasone-induced insulin resistance in rhesus macaques. J. Endocrinol. 2013, 216, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Bonisch, C.; Irmler, M.; Brachthauser, L.; Neff, F.; Bamberger, M.T.; Marschall, S.; Hrabe de Angelis, M.; Beckers, J. Dexamethasone treatment alters insulin, leptin, and adiponectin levels in male mice as observed in DIO but does not lead to alterations of metabolic phenotypes in the offspring. Mamm. Genome 2016, 27, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, S.P.; Paquin, T.; Pritsker, A.; Haber, R.S. Glucocorticoid-induced insulin resistance: Dexamethasone inhibits the activation of glucose transport in rat skeletal muscle by both insulin- and non-insulin-related stimuli. Diabetes 1995, 44, 441–445. [Google Scholar] [CrossRef]
- Ruzzin, J.; Wagman, A.S.; Jensen, J. Glucocorticoid-induced insulin resistance in skeletal muscles: Defects in insulin signalling and the effects of a selective glycogen synthase kinase-3 inhibitor. Diabetologia 2005, 48, 2119–2130. [Google Scholar] [CrossRef] [Green Version]
- Sood, A.; Ismail-Beigi, F. Effect of dexamethasone on insulin secretion: Examination of underlying mechanisms. Endocr. Pract. 2010, 16, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.C.; Halter, J.B.; Best, J.D.; Pfeifer, M.A.; Porte, D., Jr. Dexamethasone-induced insulin resistance enhances B cell responsiveness to glucose level in normal men. Am. J. Physiol. 1984, 247, E592–E596. [Google Scholar] [CrossRef]
- Rafacho, A.; Marroqui, L.; Taboga, S.R.; Abrantes, J.L.; Silveira, L.R.; Boschero, A.C.; Carneiro, E.M.; Bosqueiro, J.R.; Nadal, A.; Quesada, I. Glucocorticoids in vivo induce both insulin hypersecretion and enhanced glucose sensitivity of stimulus-secretion coupling in isolated rat islets. Endocrinology 2010, 151, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Poggioli, R.; Ueta, C.B.; Drigo, R.A.; Castillo, M.; Fonseca, T.L.; Bianco, A.C. Dexamethasone reduces energy expenditure and increases susceptibility to diet-induced obesity in mice. Obesity 2013, 21, E415–E420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkawi, A.K.; Alzoubi, K.H.; Jacob, M.; Matic, G.; Ali, A.; Al Faraj, A.; Almuhanna, F.; Dasouki, M.; Abdel Rahman, A.M. Metabolomics Based Profiling of Dexamethasone Side Effects in Rats. Front. Pharmacol. 2018, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Macedo, A.G.; Krug, A.L.; Souza, L.M.; Martuscelli, A.M.; Constantino, P.B.; Zago, A.S.; Rush, J.W.; Santos, C.F.; Amaral, S.L. Time-course changes of catabolic proteins following muscle atrophy induced by dexamethasone. Steroids 2016, 107, 30–36. [Google Scholar] [CrossRef] [Green Version]
- De Vos, P.; Saladin, R.; Auwerx, J.; Staels, B. Induction of ob gene expression by corticosteroids is accompanied by body weight loss and reduced food intake. J. Biol. Chem. 1995, 270, 15958–15961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, C.Y.; Chih-Yuan Ho, K.; Lee, O.K.; Blunn, G.W.; Su, Y. Restoration of bone mass and strength in glucocorticoid-treated mice by systemic transplantation of CXCR4 and cbfa-1 co-expressing mesenchymal stem cells. J. Bone Miner. Res. 2009, 24, 837–848. [Google Scholar] [CrossRef]
- Yao, W.; Cheng, Z.; Pham, A.; Busse, C.; Zimmermann, E.A.; Ritchie, R.O.; Lane, N.E. Glucocorticoid-induced bone loss in mice can be reversed by the actions of parathyroid hormone and risedronate on different pathways for bone formation and mineralization. Arthritis Rheum. 2008, 58, 3485–3497. [Google Scholar] [CrossRef] [Green Version]
- Buckley, L.; Humphrey, M.B. Glucocorticoid-Induced Osteoporosis. N. Engl. J. Med. 2018, 379, 2547–2556. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, L.; Ferrandiz, M.L.; Brines, R.; Guede, D.; Cuadrado, A.; Alcaraz, M.J. Effects of Nrf2 deficiency on bone microarchitecture in an experimental model of osteoporosis. Oxid. Med. Cell. Longev. 2014, 2014, 726590. [Google Scholar] [CrossRef]
- Liu, Z.; Hou, Y.; Li, L.; Yang, Y.; Jia, J.; Hong, Z.; Li, T.; Xu, Y.; Fu, J.; Sun, Y.; et al. Nrf2 deficiency aggravates the increase in osteoclastogenesis and bone loss induced by inorganic arsenic. Toxicol. Appl. Pharmacol. 2019, 367, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Uruno, A.; Yumoto, A.; Taguchi, K.; Suzuki, M.; Harada, N.; Ryoke, R.; Naganuma, E.; Osanai, N.; Goto, A.; et al. Nrf2 contributes to the weight gain of mice during space travel. Commun. Biol. 2020, 3, 496. [Google Scholar] [CrossRef]
- Chartoumpekis, D.V.; Ziros, P.G.; Psyrogiannis, A.I.; Papavassiliou, A.G.; Kyriazopoulou, V.E.; Sykiotis, G.P.; Habeos, I.G. Nrf2 represses FGF21 during long-term high-fat diet-induced obesity in mice. Diabetes 2011, 60, 2465–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.K.; Wu, K.C.; Liu, J.; Klaassen, C.D. Nrf2 deficiency improves glucose tolerance in mice fed a high-fat diet. Toxicol. Appl. Pharmacol. 2012, 264, 305–314. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene NCBI ID | Forward Primer | Reverse Primer |
|---|---|---|---|
| G6pc | 14377 | CTAGCTTTGATCTGGTTGTCAG | GTTGAACCAGTCTCCGACCA |
| Gusb | 110006 | ACTGACACCTCCATGTATCCCAAG | CAGTAGGTCACCAGCCCGATG |
| Nrf2 | 18024 | CTTTAGTCAGCGACAGAAGGAC | AGGCATTCTTGTTTGGGAATGTG |
| Nqo1 | 18104 | CATTCTGAAAGGCTGGTTTGA | CTAGCTTTGATCTGGTTGTCAG |
| Pck1 | 18534 | CTGCATAACGGTCTGGACTTC | CAGCAACTGCCCGTACTCC |
| Ppia | 268373 | CAGACGCCACTGGTCGCTTT | TGTCTTTGGAACTTTGTCTGCAA |
| Rps29 | 20090 | TCTACTGGAGTCACCCACGGAA | GGAAGCACTGGCGGCACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippopoulou, F.; Habeos, G.I.; Rinotas, V.; Sophocleous, A.; Sykiotis, G.P.; Douni, E.; Chartoumpekis, D.V. Dexamethasone Administration in Mice Leads to Less Body Weight Gain over Time, Lower Serum Glucose, and Higher Insulin Levels Independently of NRF2. Antioxidants 2022, 11, 4. https://doi.org/10.3390/antiox11010004
Filippopoulou F, Habeos GI, Rinotas V, Sophocleous A, Sykiotis GP, Douni E, Chartoumpekis DV. Dexamethasone Administration in Mice Leads to Less Body Weight Gain over Time, Lower Serum Glucose, and Higher Insulin Levels Independently of NRF2. Antioxidants. 2022; 11(1):4. https://doi.org/10.3390/antiox11010004
Chicago/Turabian StyleFilippopoulou, Fotini, George I. Habeos, Vagelis Rinotas, Antonia Sophocleous, Gerasimos P. Sykiotis, Eleni Douni, and Dionysios V. Chartoumpekis. 2022. "Dexamethasone Administration in Mice Leads to Less Body Weight Gain over Time, Lower Serum Glucose, and Higher Insulin Levels Independently of NRF2" Antioxidants 11, no. 1: 4. https://doi.org/10.3390/antiox11010004