
Sodium Selenite Enhances Antibiotics Sensitivity of Pseudomonas aeruginosa and Deceases Its Pathogenicity by Inducing Oxidative Stress and Inhibiting Quorum Sensing System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Survival Assays of Antimicrobial Treatment
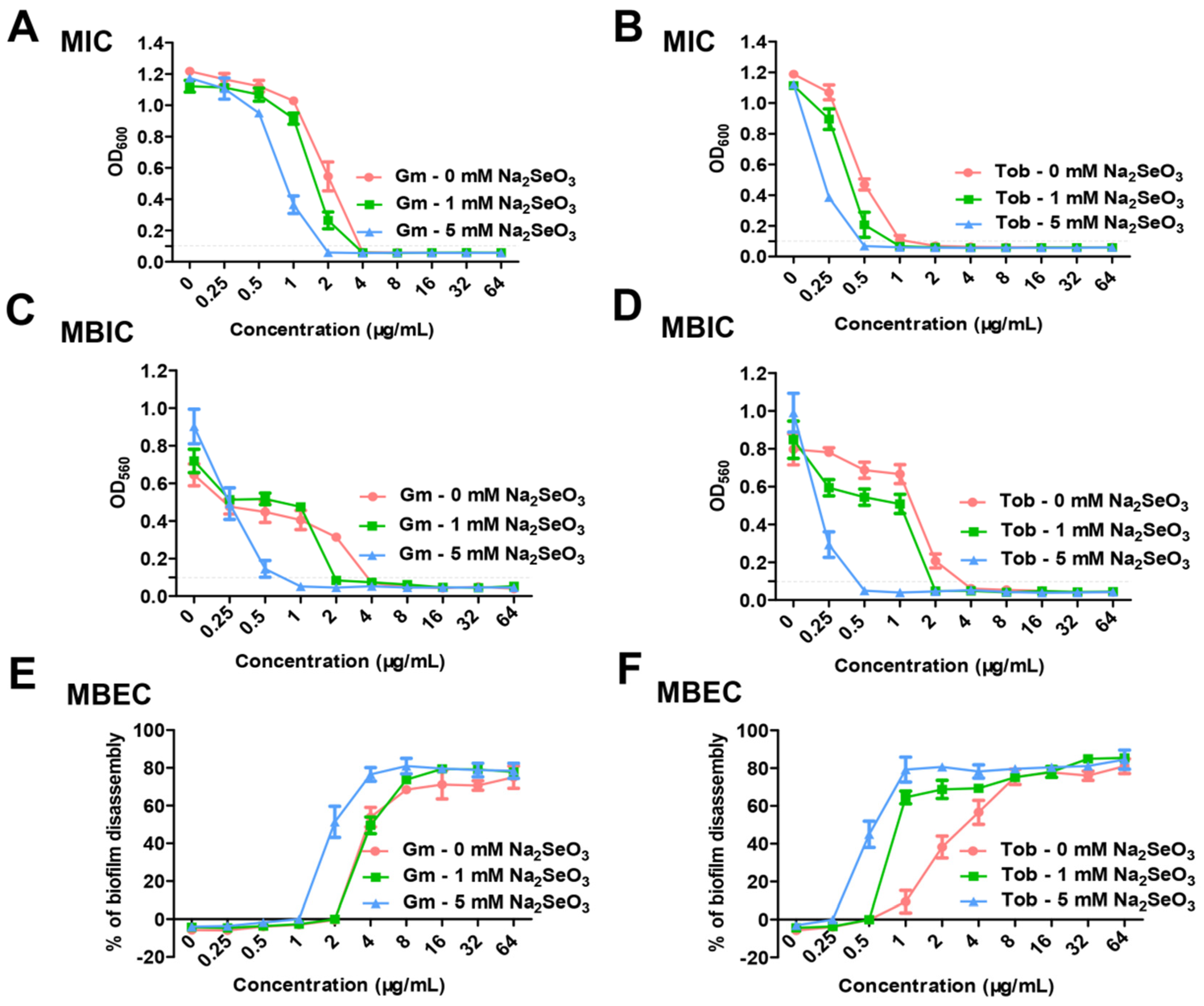
2.3. Determination of Minimal Inhibitory Concentration (MIC), Minimal Biofilm Inhibitory Concentration (MBIC), and Minimal Biofilm Eradication Concentration (MBEC) with the Combination of Sodium Selenite and Antibiotics
2.4. Biofilm Formation Measurement
2.5. Pyocyanin Production Assay
2.6. Bacterial Motility Examination
2.7. Rhamnolipid Quantification
2.8. Construction of the Promoter-Reporter Plasmids
2.9. Gene Expression Analysis
2.10. Measurement of Intracellular ROS Level
2.11. Superoxide Dismutase (SOD) Activity Assay
2.12. RNA-Seq Sample Preparation and Sequencing
2.13. Bioinformatics Analysis
2.14. Promoter Activity Assay
2.15. Chinese Cabbage Infection Assay
2.16. Drosophila Melanogaster Infection Assay
2.17. Statistical Analysis
3. Results
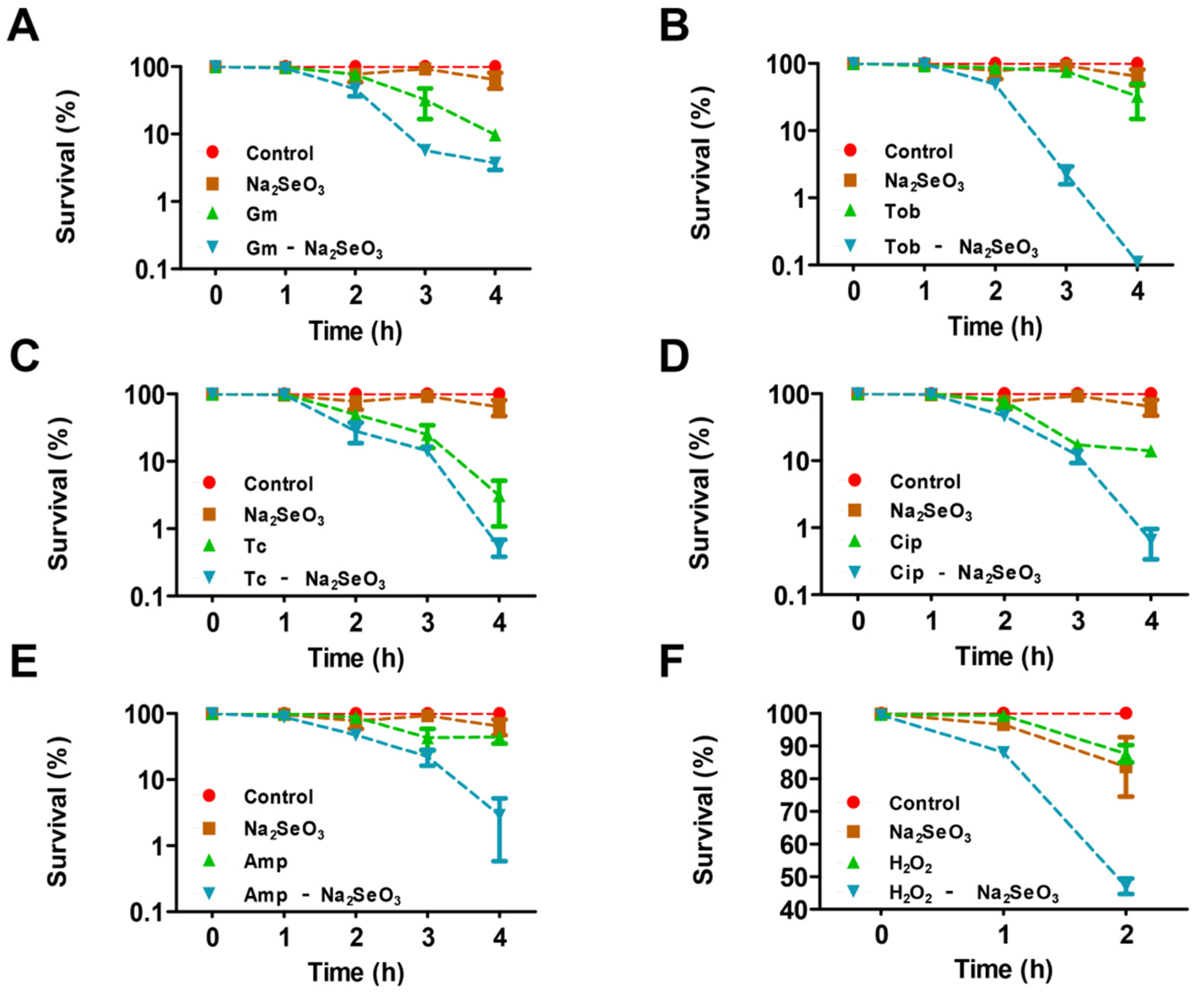
3.1. Sodium Selenite Increased the Lethality Rate of P. aeruginosa PAO1 with Antibiotics or H2O2 Treatment and the Antibiotics Susceptibility in Planktonic and Biofilm States
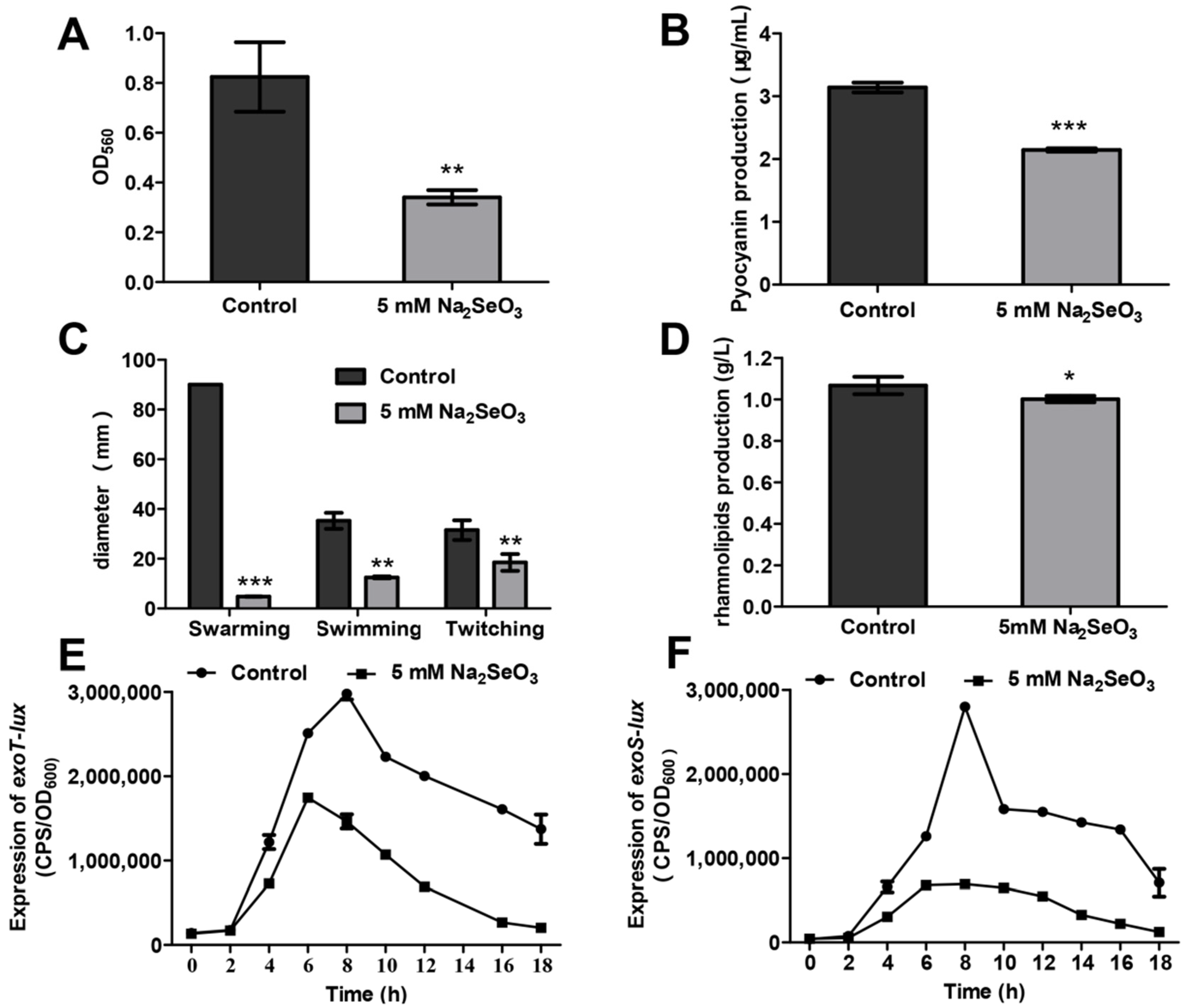
3.2. Sodium Selenite Inhibited Biofilm Formation, Pyocyanin Production, Bacterial Motilities, and the Type III Secretion System (T3SS)
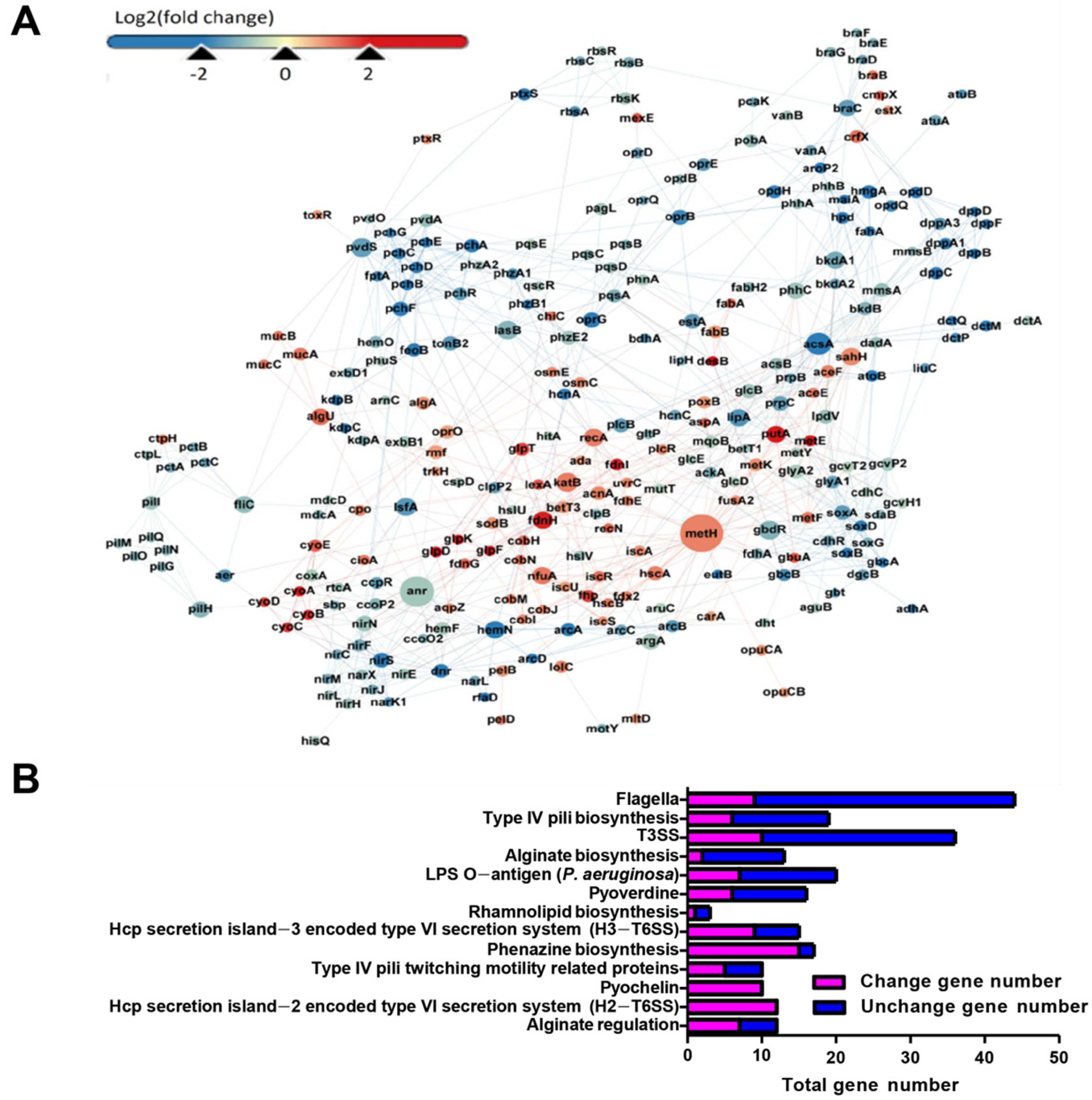
3.3. RNA-Seq Analysis Revealed the Global Response of P. aeruginosa in the Presence of 5 mM Sodium Selenite
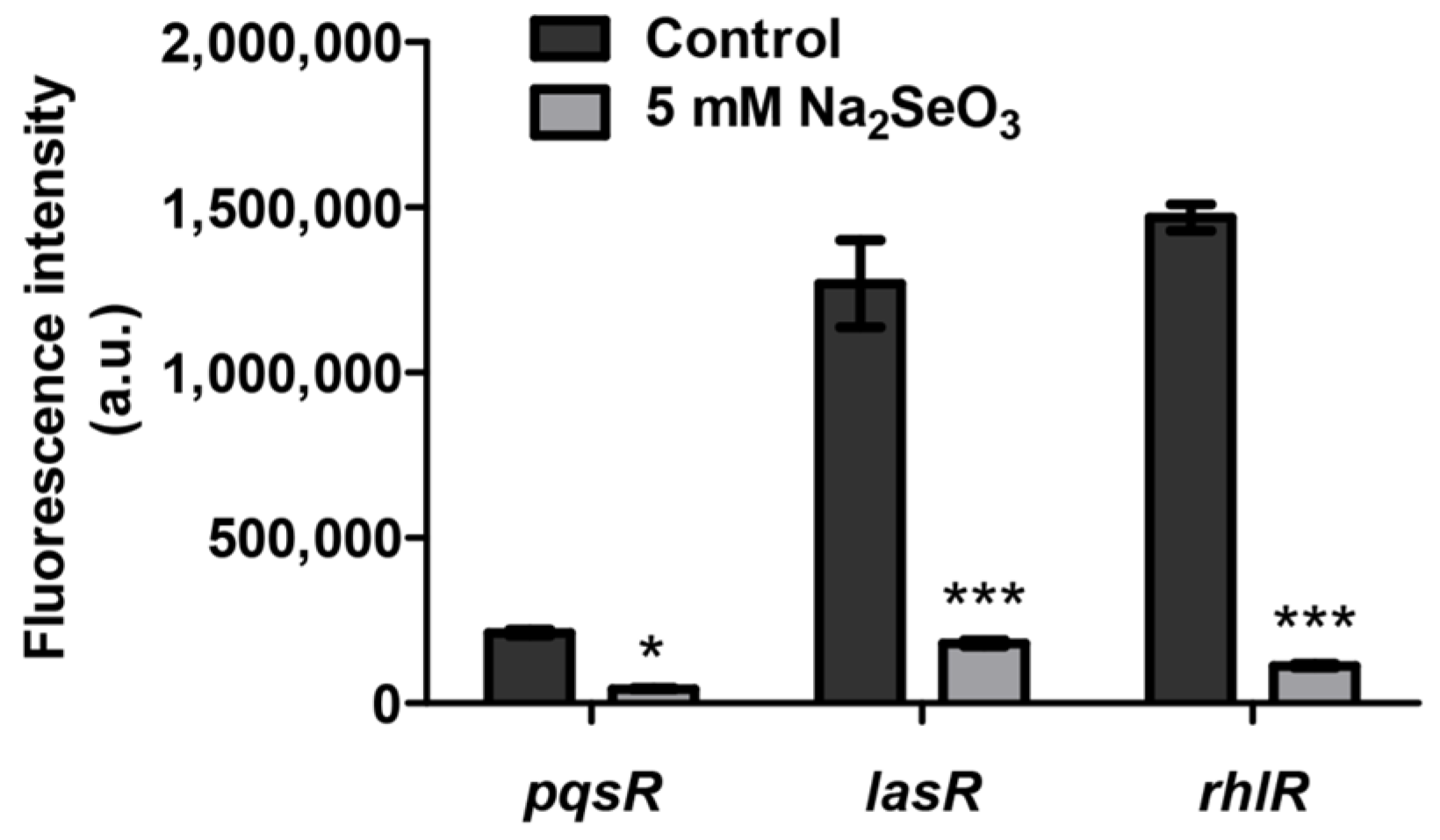
3.4. Sodium Selenite Inhibited the Expression of Quorum Sensing Genes
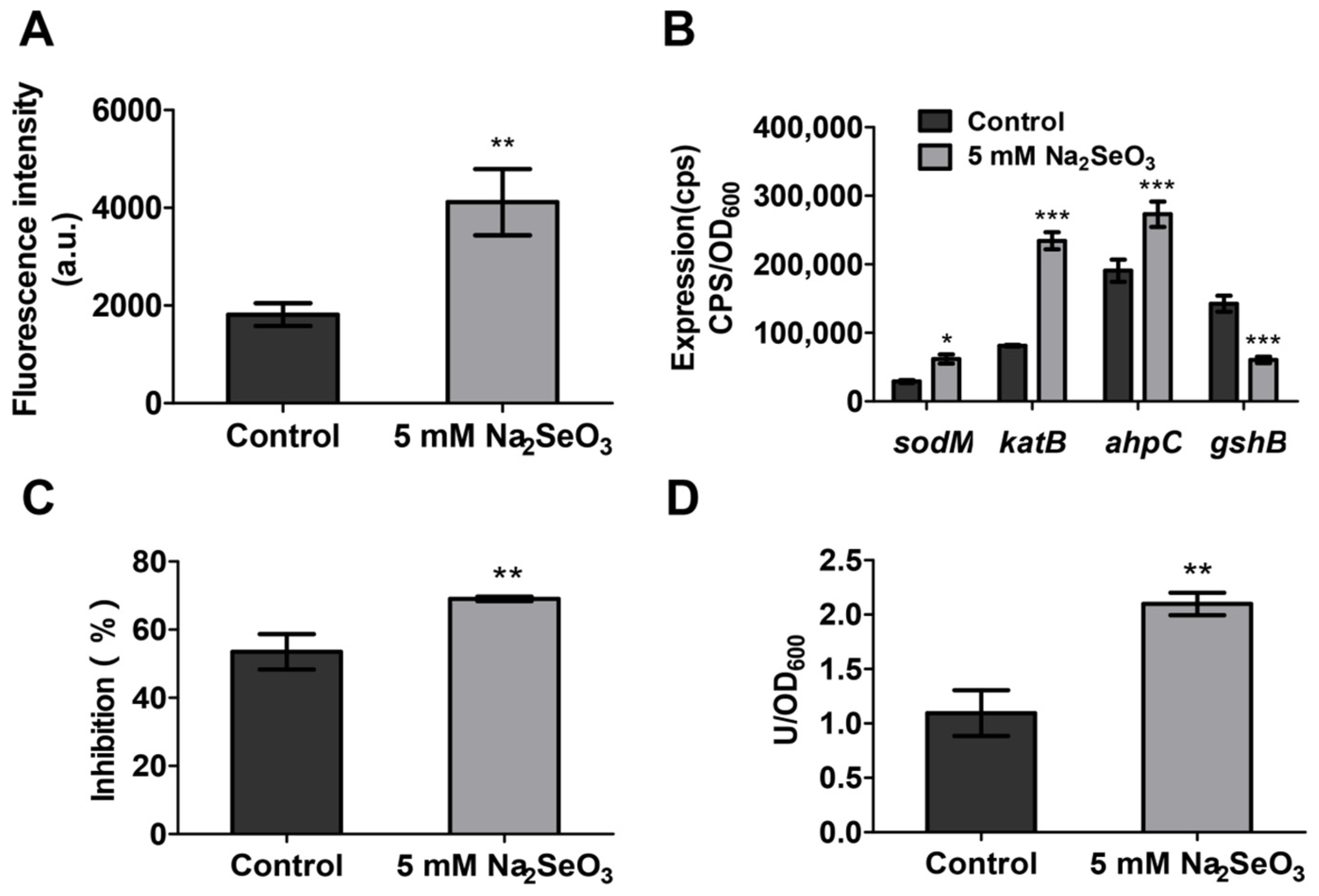
3.5. Sodium Selenite Increased the ROS Levels and the Expression of Oxidative Stress Related Genes
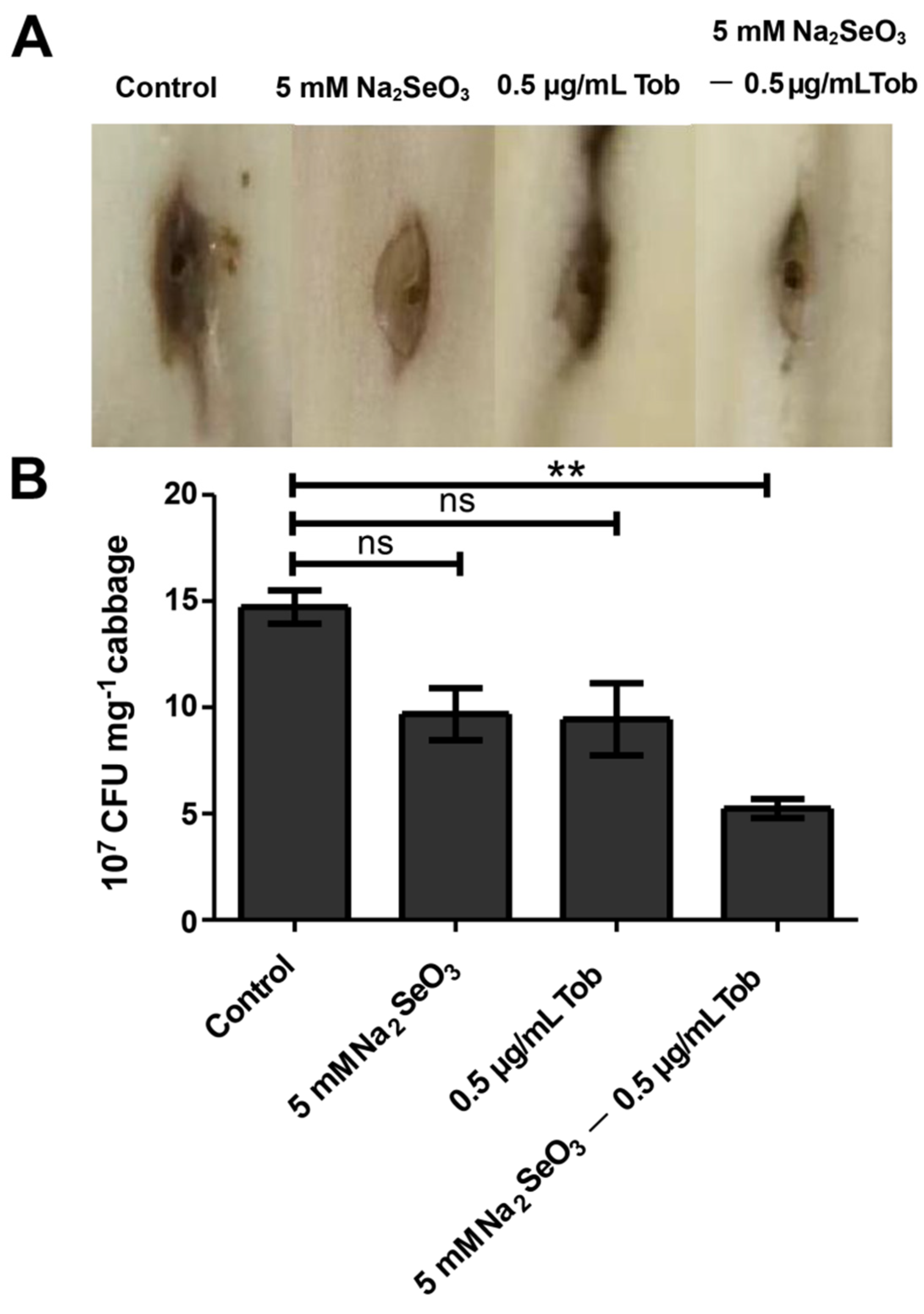
3.6. Sodium Selenite Reduced the Virulence of P. aeruginosa in Chinese Cabbage and D. melanogaster Infection Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Grosso-Becerra, M.V.; Croda-García, G.; Merino, E.; Servín-González, L.; Mojica-Espinosa, R.; Soberón-Chávez, G. Regulation of Pseudomonas aeruginosa virulence factors by two novel RNA thermometers. Proc. Natl. Acad. Sci. USA 2014, 111, 15562–15567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.W.; Luo, H.Z.; Jiang, H.; Jian, T.K.; Chen, Z.Q.; Jia, A.Q. Hordenine: A novel quorum sensing inhibitor and antibiofilm agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 2018, 66, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Anil Kumar, V.; Gopi Mohan, C.; Biswas, R. Antibiotic resistance in Pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Hurley, M.N.; Cámara, M.; Smyth, A.R. Novel approaches to the treatment of Pseudomonas aeruginosa infections in cystic fibrosis. Eur. Respir. J. 2012, 40, 1014–1023. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: Isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef]
- Bronesky, D.; Wu, Z.; Marzi, S.; Walter, P.; Geissmann, T.; Moreau, K.; Vandenesch, F.; Caldelari, I.; Romby, P. Staphylococcus aureus RNAIII and its regulon link quorum sensing, stress responses, metabolic adaptation, and regulation of virulence gene expression. Annu. Rev. Microbiol. 2016, 70, 299–316. [Google Scholar] [CrossRef]
- Barnard, A.M.; Bowden, S.D.; Burr, T.; Coulthurst, S.J.; Monson, R.E.; Salmond, G.P. Quorum sensing, virulence and secondary metabolite production in plant soft-rotting bacteria. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2007, 362, 1165–1183. [Google Scholar] [CrossRef] [Green Version]
- Hammer, B.K.; Bassler, B.L. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol. Microbiol. 2003, 50, 101–104. [Google Scholar] [CrossRef]
- García-Contreras, R. Is quorum sensing interference a viable alternative to treat pseudomonas aeruginosa infections? Front. Microbiol. 2016, 7, 1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, K.; Foltzs, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. Nutrition 1999, 15, 225. [Google Scholar]
- Biswas, K.C.; Barton, L.L.; Tsui, W.L.; Shuman, K.; Eze, C.S. A novel method for the measurement of elemental selenium produced by bacterial reduction of selenite. J. Microbiol. Meth. 2011, 86, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Li, D.B.; Cheng, Y.Y.; Wu, C.; Li, W.W.; Li, N.; Yang, Z.C.; Tong, Z.H.; Yu, H.Q. Selenite reduction by Shewanella oneidensis MR-1 is mediated by fumarate reductase in periplasm. Sci. Rep. 2014, 4, 3735. [Google Scholar] [CrossRef] [PubMed]
- Spallholz, J.E. On the nature of selenium toxicity and carcinostatic activity. Free Radic. Biol. Med. 1994, 17, 45–64. [Google Scholar] [CrossRef]
- Cihalova, K.; Chudobova, D.; Michalek, P.; Moulick, A.; Guran, R.; Kopel, P.; Adam, V.; Kizek, R. Staphylococcus aureus and MRSA growth and biofilm formation after treatment with antibiotics and SeNPs. Int. J. Mol. Sci. 2015, 16, 24656–24672. [Google Scholar] [CrossRef]
- Misra, S.; Boylan, M.; Selvam, A.; Spallholz, J.E.; Björnstedt, M. Redox-active selenium compounds—From toxicity and cell death to cancer treatment. Nutrients 2015, 7, 3536–3556. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.P.; Gandin, V. Selenium compounds as therapeutic agents in cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef]
- Hatfield, D.L. Redox pioneer: Professor Vadim N. gladyshev. Antioxid. Redox Signal. 2016, 25, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Firoz, A.M.; Safhi, M.M.; Sivagurunathan, M.S.; Aamena, J. In vitro antibacterial spectrum of sodium selenite against selected human pathogenic bacterial strains. Scientifica 2016, 2016, 9176273. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yu, S.; Zhang, Z.; Wei, Q.; Yan, L.; Ai, G.; Liu, H.; Ma, L.Z. Coordination of swarming motility, biosurfactant synthesis, and biofilm matrix exopolysaccharide production in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2014, 80, 6724–6732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brynildsen, M.P.; Winkler, J.A.; Spina, C.S.; MacDonald, I.C.; Collins, J.J. Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production. Nat. Biotechnol. 2013, 31, 160–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockerill, F.R.; Wiker, M.A.; Alder, J.; Dudley, M.N.; Eliopoulos, G.M.; Ferraro, M.J.; Hardy, D.J.; Hecht, D.W.; Hindler, J.A.; Patel, J.B. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 9th ed.; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Ma, L.; Jackson, K.D.; Landry, R.M.; Parsek, M.R.; Wozniak, D.J. Analysis of Pseudomonas aeruginosa conditional psl variants reveals roles for the psl polysaccharide in adhesion and maintaining biofilm structure postattachment. J. Bacteriol. 2006, 188, 8213–8221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leoni, L.; Rampioni, G. A coculture-based approach for screening campaigns aimed at identifying novel Pseudomonas aeruginosa quorum sensing inhibitors. Methods Mol. Biol. 2018, 1673, 287–296. [Google Scholar]
- Kurachi, M. Studies on the biosynthesis of pyocyanine. (II): Isolation and determination of pyocyanine. Bull. Inst. Chem. Res. Kyoto Univ. 1958, 36, 174–187. [Google Scholar]
- Chayabutra, C.; Wu, J.; Ju, L.K. Rhamnolipid production by Pseudomonas aeruginosa under denitrification: Effects of limiting nutrients and carbon substrates. Biotechnol. Bioeng. 2001, 72, 25–33. [Google Scholar] [CrossRef]
- Duan, K.; Dammel, C.; Stein, J.; Rabin, H.; MG, S. Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol. Microbiol. 2003, 50, 1477–1491. [Google Scholar] [CrossRef] [Green Version]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, P3. [Google Scholar] [CrossRef]
- Fito-Boncompte, L.; Chapalain, A.; Bouffartigues, E.; Chaker, H.; Lesouhaitier, O.; Gicquel, G.; Bazire, A.; Madi, A.; Connil, N.; Veron, W.; et al. Full virulence of Pseudomonas aeruginosa requires OprF. Infect. Immun. 2011, 79, 1176–1186. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, X.; Liu, H.; Zhang, L.; Guo, Y.; Yu, S.; Wozniak, D.J.; Ma, L.Z. The exopolysaccharide Psl-eDNA interaction enables the formation of a biofilm skeleton in Pseudomonas aeruginosa. Environ. Microbiol. Rep. 2015, 7, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Singh, S.P.; Häder, D.P.; Sinha, R.P. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2′,7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem. Biophys. Res. Commun. 2010, 397, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Jayaraman, P.; Sakharkar, M.K.; Lim, C.S.; Tang, T.H.; Sakharkar, K.R. Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. Int. J. Biol. Sci. 2010, 6, 556–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, P.A.; Webster, T.J. Selenium nanoparticles inhibit Staphylococcus aureus growth. Int. J. Nanomed. 2011, 6, 1553–1558. [Google Scholar] [CrossRef] [Green Version]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival, and persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Fang, X.; Wood, T.K.; Huang, Z.J. A systems-level approach for investigating Pseudomonas aeruginosa biofilm formation. PLoS ONE 2013, 8, e57050. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Manefield, M. Pyocyanin promotes extracellular DNA release in Pseudomonas aeruginosa. PLoS ONE 2012, 7, e46718. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, G.F.; Sartorio, R.; Lee, S.H.; Rogers, C.J.; Meijler, M.M.; Moss, J.A.; Clapham, B.; Brogan, A.P.; Dickerson, T.J.; Janda, K.D. Revisiting quorum sensing: Discovery of additional chemical and biological functions for 3-oxo-N-acylhomoserine lactones. Proc. Natl. Acad. Sci. USA 2005, 102, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Nickzad, A.; Déziel, E. The involvement of rhamnolipids in microbial cell adhesion and biofilm development—An approach for control? Lett. Appl. Microbiol. 2014, 58, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Conrad, J.C. Physics of bacterial near-surface motility using flagella and type IV pili: Implications for biofilm formation. Res. Microbiol. 2012, 163, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, D.J. Oxidative stress responses of the yeast Saccharomyces cerevisiae. Yeast 1998, 14, 1511–1527. [Google Scholar] [CrossRef]
- Smirnova, G.V.; Oktyabrsky, O.N. Glutathione in bacteria. Biochemistry 2005, 70, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Vasil, M.L.; Alsabbagh, E.; Parvatiyar, K.; Hassett, D.J. Role of the Pseudomonas aeruginosa oxyR-recG operon in oxidative stress defense and DNA repair: OxyR-dependent regulation of katB-ankB, ahpB and ahpC-ahpF. J. Bacteriol. 2000, 182, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Woo, E.R.; Lee, D.G. Phytol has antibacterial property by inducing oxidative stress response in Pseudomonas aeruginosa. Free Radic. Res. 2016, 50, 1309–1318. [Google Scholar] [CrossRef]
- Wongsaroj, L.; Saninjuk, K.; Romsang, A.; Duang-Nkern, J.; Trinachartvanit, W.; Vattanaviboon, P.; Mongkolsuk, S. Pseudomonas aeruginosa glutathione biosynthesis genes play multiple roles in stress protection, bacterial virulence and biofilm formation. PLoS ONE 2018, 13, e0205815. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Zhang, Y.; Catron, B.; Chan, Q.; Hu, Q.; Caruso, J.A. Identification of selenium compounds using HPLC-ICPMS and nano-ESI-MS in selenium-enriched rice via foliar application. J. Anal. At. Spectrom. 2009, 24, 1657–1664. [Google Scholar] [CrossRef]
- Cegelski, L.; Marshall, G.R.; Eldridge, G.R.; Hultgren, S.J. The biology and future prospects of antivirulence therapies. Nat. Rev. Microbiol. 2008, 6, 17–27. [Google Scholar] [CrossRef]
- Benko, I.; Nagy, G.; Tanczos, B.; Ungvari, E.; Sztrik, A.; Eszenyi, P.; Prokisch, J.; Banfalvi, G. Subacute toxicity of nano-selenium compared to other selenium species in mice. Environ. Toxicol. Chem. 2012, 31, 2812–2820. [Google Scholar] [CrossRef]
- Zonaro, E.; Lampis, S.; Turner, R.J.; Qazi, S.J.; Vallini, G. Biogenic selenium and tellurium nanoparticles synthesized by environmental microbial isolates efficaciously inhibit bacterial planktonic cultures and biofilms. Front. Microbiol. 2015, 6, 584. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdi, M.H.; Mahdavi, M.; Varastehmoradi, B.; Faramarzi, M.A.; Shahverdi, A.R. The immunostimulatory effect of biogenic selenium nanoparticles on the 4T1 breast cancer model: An in vivo study. Biol. Trace Elem. Res. 2012, 149, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Forootanfar, H.; Adeli-Sardou, M.; Nikkhoo, M.; Mehrabani, M.; Amir-Heidari, B.; Shahverdi, A.R.; Shakibaie, M. Antioxidant and cytotoxic effect of biologically synthesized selenium nanoparticles in comparison to selenium dioxide. J. Trace Elem. Med. Biol. 2014, 28, 75–79. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names 1 | Relevant Characteristics | Accession Number of the Involved Genes 2 | Source |
|---|---|---|---|
| Strains | |||
| E. coli DH5α | F– φ80lacZ ΔM15 Δ(lacZYA-argF)U169 recA1 endA1 hsdR17 (rk, mk+) phoA supE44 thi-1 gyrA96 relA1 tonA | - | Tiangen |
| P. aeruginosa PAO1 | P. aeruginosa wild type | - | this lab |
| PAO1 (pHERD20T- pqsR-gfp) | PAO1 containing pqsR-gfp reporter plasmid pHERD20T (Cbr) | 879994 | this lab |
| PAO1 (pHERD20T- lasR-gfp) | PAO1 containing lasR-gfp reporter plasmid pHERD20T (Cbr) | 881789 | this lab |
| PAO1 (pHERD20T- rhlR-gfp) | PAO1 containing rhlR-gfp reporter plasmid pHERD20T (Cbr) | 878968 | this lab |
| Plasmids | |||
| CTX-exoS | Integration plasmid, CTX6.1 with a fragment of pKD-exoS containing exoS promoter region and luxCDABE gene; Knr, Tmpr, Tcr | 879837 | this lab |
| CTX-exoT | Integration plasmid, CTX6.1 with a fragment of pKD-exoT containing exoT promoter region and luxCDABE gene; Knr, Tmpr, Tcr | 878350 | this lab |
| pMS402 | Expression reporter plasmid carrying the promoterless luxCDABE gene; Knr, Tmpr | - | [21] |
| pKD-sodM | pMS402 containing sodM promoter region; Knr, Tmpr | 881040 | this study |
| pKD-katB | pMS402 containing katB promoter region; Knr, Tmpr | 881120 | this study |
| pKD-ahpC | pMS402 containing ahpC promoter region; Knr, Tmpr | 879431 | this study |
| pKD-gshB | pMS402 containing gshB promoter region; Knr, Tmpr | 878223 | this study |
| Primers | Sequence (5′→3′) | Restriction sites | |
| sodM-S | TATCTCGAGTGGTCGAGTCGATGATGG | 881040 | Xho I |
| sodM-A | ATAGGATCCGACATGGCAACCTCACCA | 881040 | Bam HI |
| katB-S | GATCTCGAGTCACTCCCTGTATTTCGC | 881120 | Xho I |
| katB-A | GTAGGATCCAGGGTTCATGGAAGAGCT | 881120 | Bam HI |
| ahpC-S | GATCTCGAGGGCAGGTTCTTCGATTAG | 879431 | Xho I |
| ahpC-A | GTTGGATCCTCAGGGACATCAGTCGTT | 879431 | Bam HI |
| gshB-S | GATCTCGAGCACTTTCAAACCGTCGGA | 878223 | Xho I |
| gshB-A | GAAGGATCCCGTACGCTCATGGGAATT | 878223 | Bam HI |
| Locus Tag | Gene Name | Fold Change | Padj | Product |
|---|---|---|---|---|
| PA0996 | pqsA | 0.3252 | 1.01 × 10−47 | PqsA |
| PA0997 | pqsB | 0.3648 | 3.40 × 10−38 | PqsB |
| PA0998 | pqsC | 0.3866 | 6.23 × 10−32 | PqsC |
| PA0999 | pqsD | 0.3954 | 1.53 × 10−33 | 3-oxoacyl-[acyl-carrier-protein] synthase III |
| PA1000 | pqsE | 0.4951 | 3.45 × 10−18 | quinolone signal response protein |
| PA1432 | lasI | 0.8302 | 9.27 × 10−2 | autoinducer synthesis protein LasI |
| PA1430 | lasR | 0.9847 | 8.73 × 10−3 | transcriptional regulator LasR |
| PA3476 | rhlI | 0.6924 | 2.36 × 10−5 | autoinducer synthesis protein RhlI |
| PA3477 | rhlR | 0.7597 | 3.37 × 10−4 | transcriptional regulator RhlR |
| Locus Tag | Gene Name | Fold Change | Padj | Product |
|---|---|---|---|---|
| PA2273 | soxR | 1.0377 | 7.98 × 10−1 | SoxR |
| PA3183 | zwf | 1.1942 | 1.68 × 10−2 | Glucose-6-phosphate 1-dehydrogenase |
| PA4366 | sodB | 2.2508 | 7.69 × 10−23 | superoxide dismutase |
| PA1562 | acnA | 2.5174 | 1.32 × 10−40 | aconitate hydratase 1 |
| PA5344 | oxyR | 0.9082 | 2.24 × 10−1 | OxyR |
| PA4613 | katB | 2.6860 | 2.03 × 10−39 | catalase |
| PA2147 | katE | 1.9478 | 2.38 × 10−4 | catalase HPII |
| PA2185 | katN | 1.8293 | 3.23 × 10−3 | non-heme catalase KatN |
| PA0139 | ahpC | 1.1545 | 4.69 × 10−2 | alkyl hydroperoxide reductase subunit C |
| PA0140 | ahpF | 1.6734 | 3.49 × 10−13 | alkyl hydroperoxide reductase subunit F |
| PA0848 | ahpB | 2.6370 | 2.96 × 10−28 | alkyl hydroperoxide reductase |
| PA2025 | gor | 1.3427 | 7.19 × 10−5 | glutathione reductase |
| PA5203 | gshA | 0.8096 | 4.88 × 10−3 | glutamate--cysteine ligase |
| PA0407 | gshB | 0.7360 | 6.81 × 10−5 | glutathione synthetase |
| PA4210 | phzA1 | 0.1995 | 1.20 × 10−9 | probable phenazine biosynthesis protein |
| PA1899 | phzA2 | 0.4405 | 1.28 × 10−2 | probable phenazine biosynthesis protein |
| PA4214 | phzE1 | 0.4272 | 1.17 × 10−12 | phenazine biosynthesis protein PhzE |
| PA1903 | phzE2 | 0.4272 | 1.17 × 10−12 | phenazine biosynthesis protein PhzE |
| PA3812 | iscA | 2.2484 | 1.52 × 10−28 | probable iron-binding protein IscA |
| PA3813 | iscU | 2.1258 | 4.92 × 10−28 | probable iron-binding protein IscU |
| PA4615 | fprB | 2.8740 | 1.41 × 10−46 | FprB |
| PA2356 | msuD | 1.9334 | 2.24 × 10−1 | methanesulfonate sulfonatase MsuD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, W.; Tian, Q.; Yang, Q.; Liu, Y.; Wang, G.; Cao, Y.; Wang, L.; Xia, S.; Sun, Y.; Zhao, C.; et al. Sodium Selenite Enhances Antibiotics Sensitivity of Pseudomonas aeruginosa and Deceases Its Pathogenicity by Inducing Oxidative Stress and Inhibiting Quorum Sensing System. Antioxidants 2021, 10, 1873. https://doi.org/10.3390/antiox10121873
Kong W, Tian Q, Yang Q, Liu Y, Wang G, Cao Y, Wang L, Xia S, Sun Y, Zhao C, et al. Sodium Selenite Enhances Antibiotics Sensitivity of Pseudomonas aeruginosa and Deceases Its Pathogenicity by Inducing Oxidative Stress and Inhibiting Quorum Sensing System. Antioxidants. 2021; 10(12):1873. https://doi.org/10.3390/antiox10121873
Chicago/Turabian StyleKong, Weina, Qianqian Tian, Qiaoli Yang, Yu Liu, Gongting Wang, Yanjun Cao, Liping Wang, Sizhe Xia, Yanmei Sun, Cheng Zhao, and et al. 2021. "Sodium Selenite Enhances Antibiotics Sensitivity of Pseudomonas aeruginosa and Deceases Its Pathogenicity by Inducing Oxidative Stress and Inhibiting Quorum Sensing System" Antioxidants 10, no. 12: 1873. https://doi.org/10.3390/antiox10121873