
Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Analysis of TMA–TMAO Pathway
2.3. Analysis of SCFAs
2.4. Gut Microbiota Compositions
2.5. Analysis of NO Pathway
2.6. Statistical Analysis
3. Results
3.1. Blood Pressure and Renal Function
3.2. TMA–TMAO Pathway
3.3. Fecal SCFA Levels
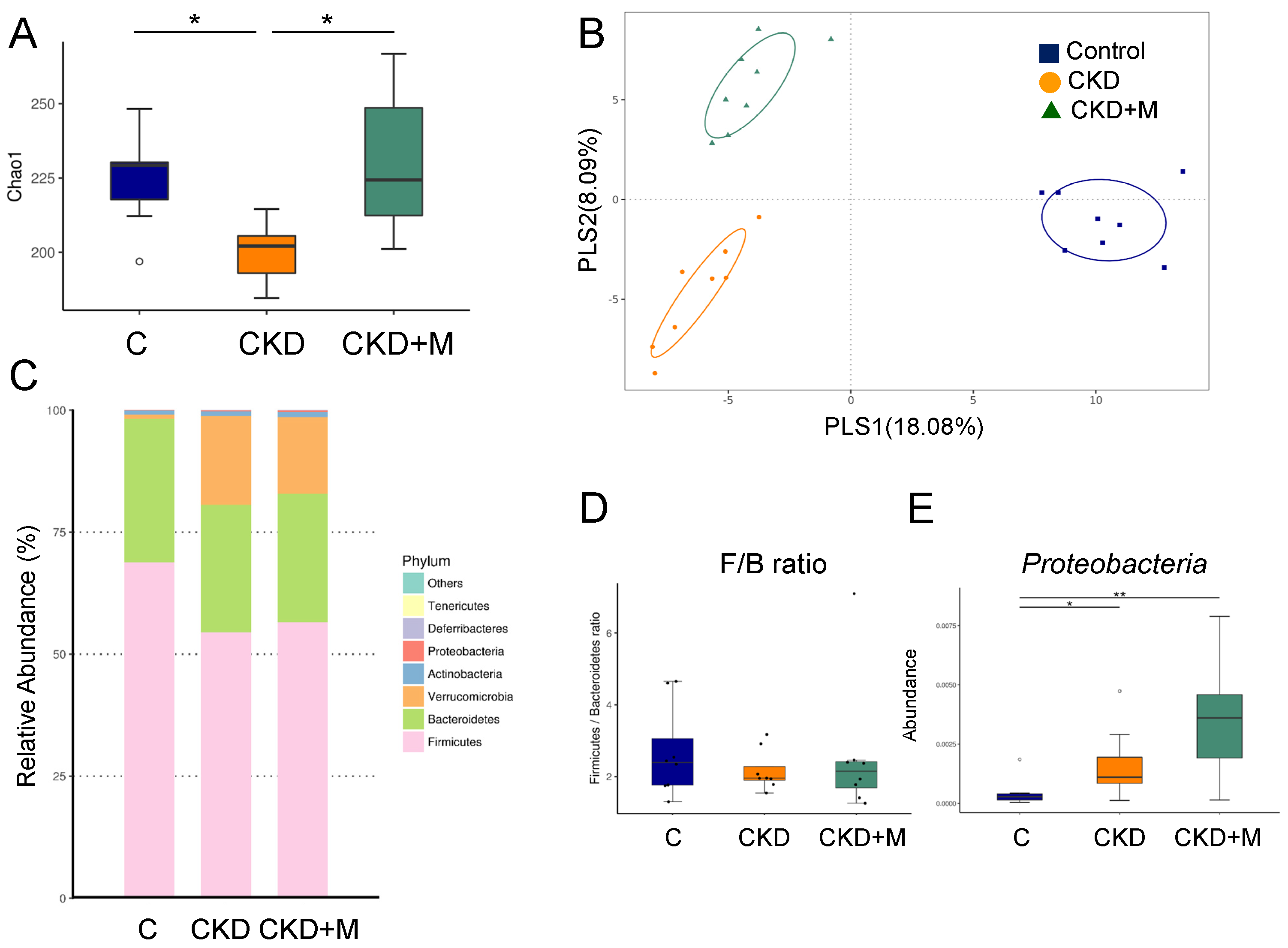
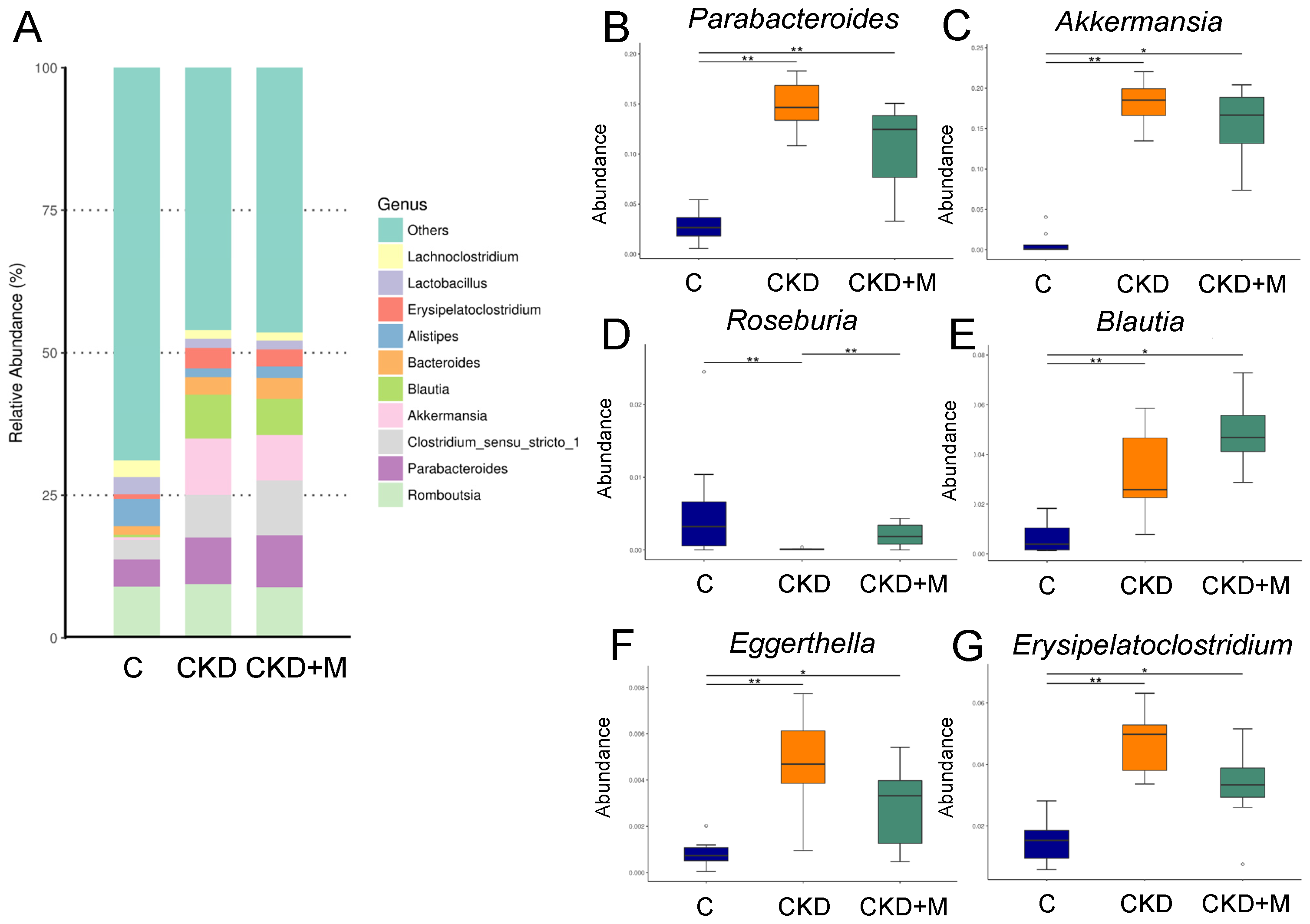
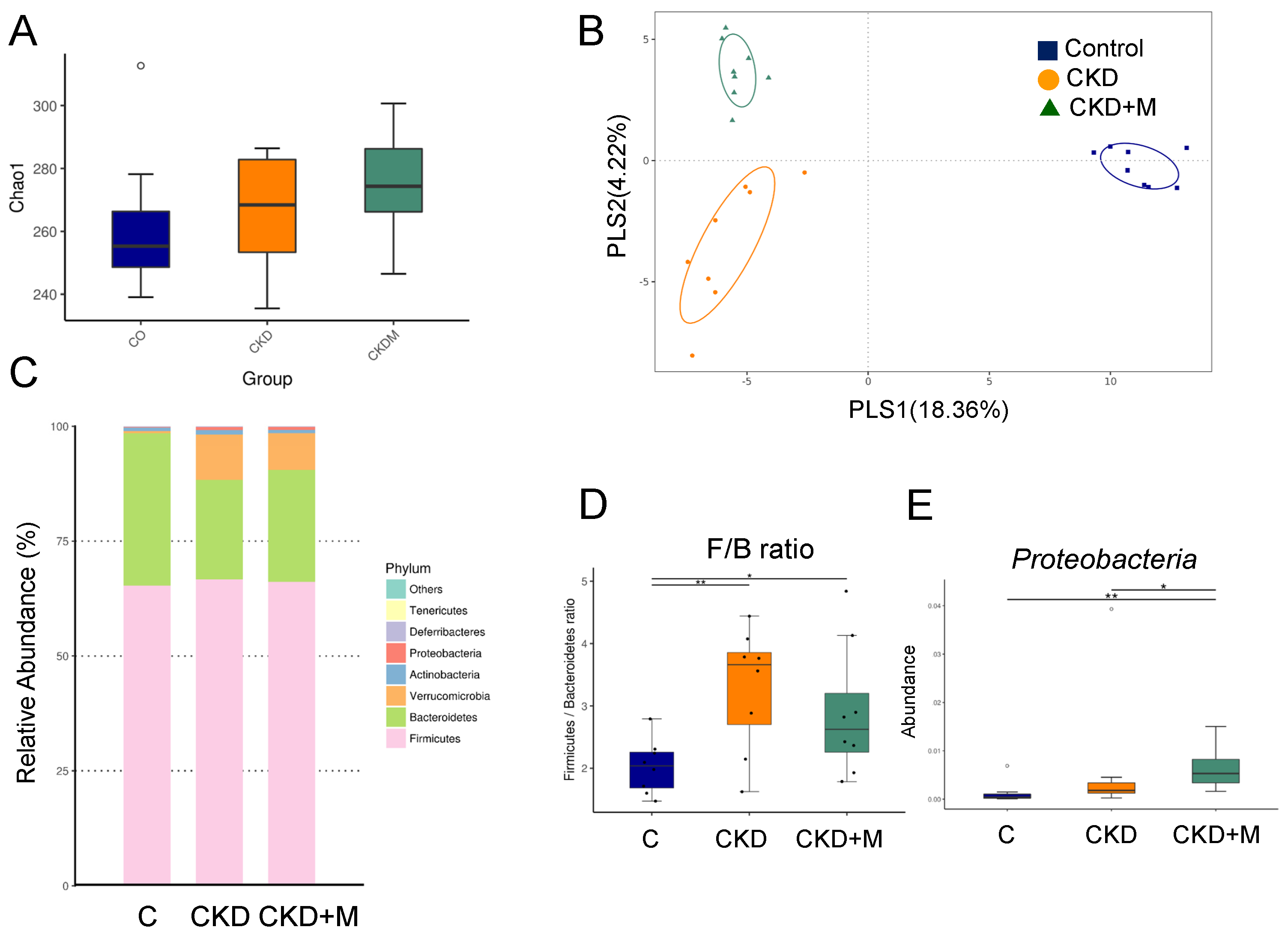
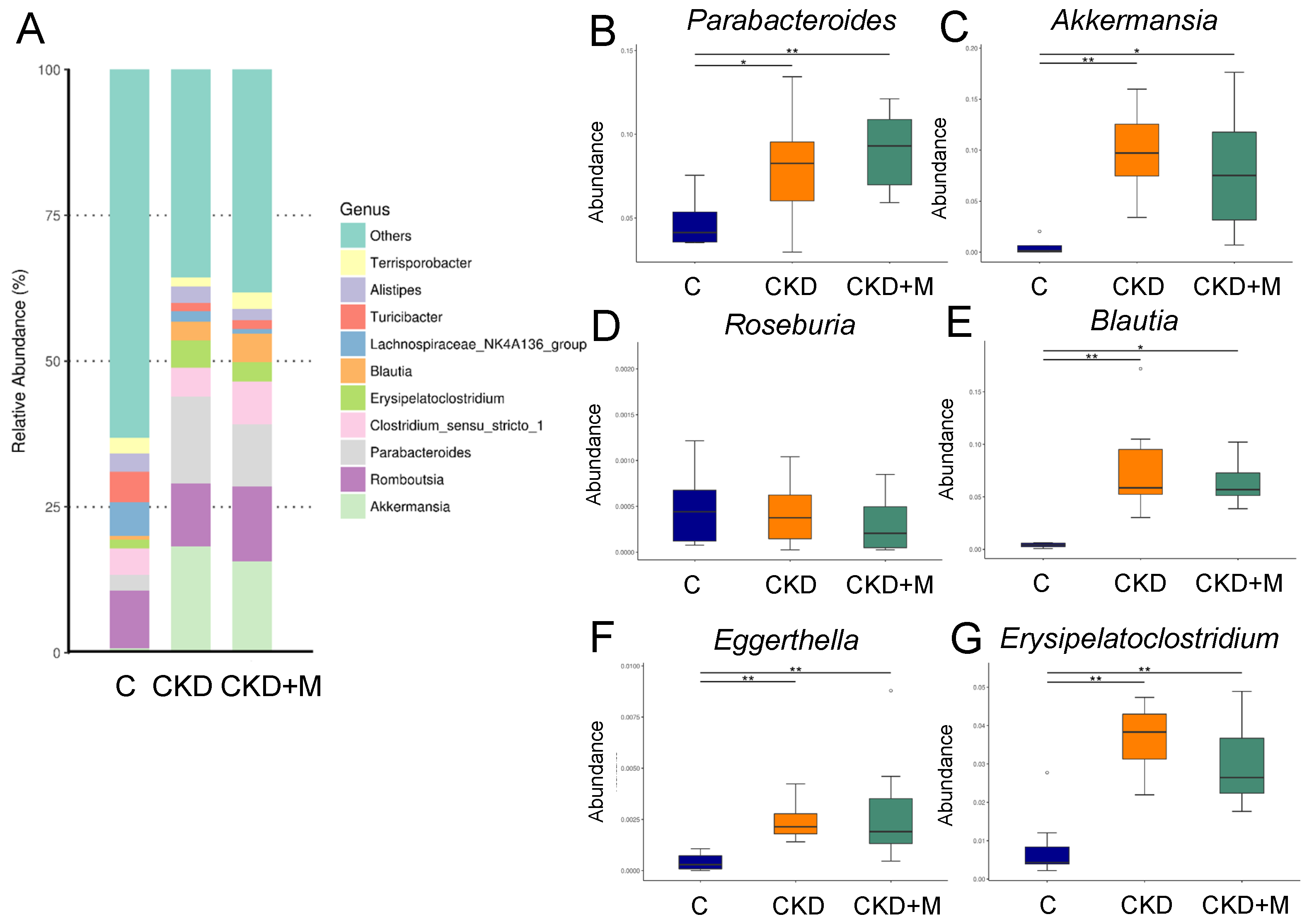
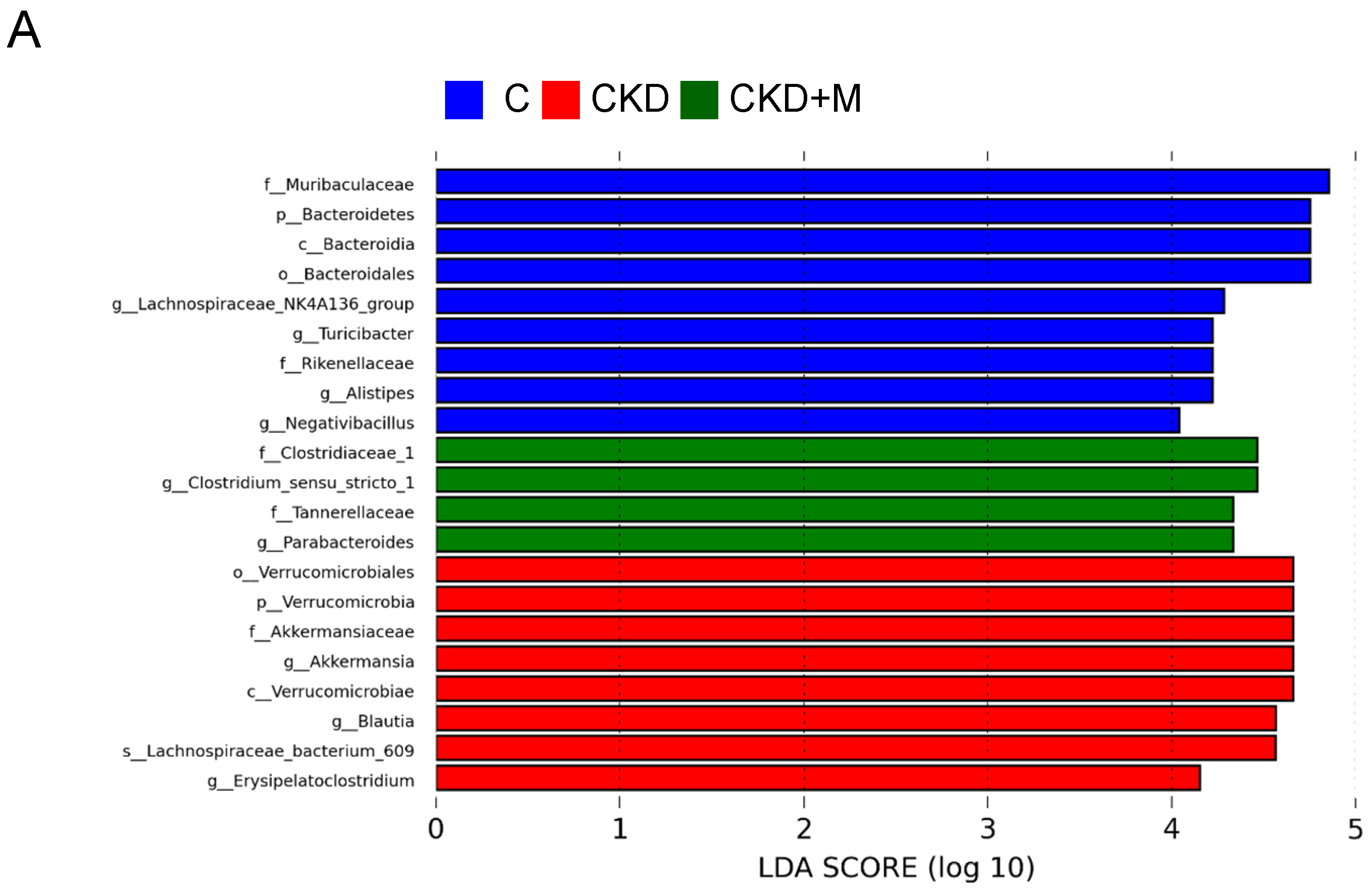
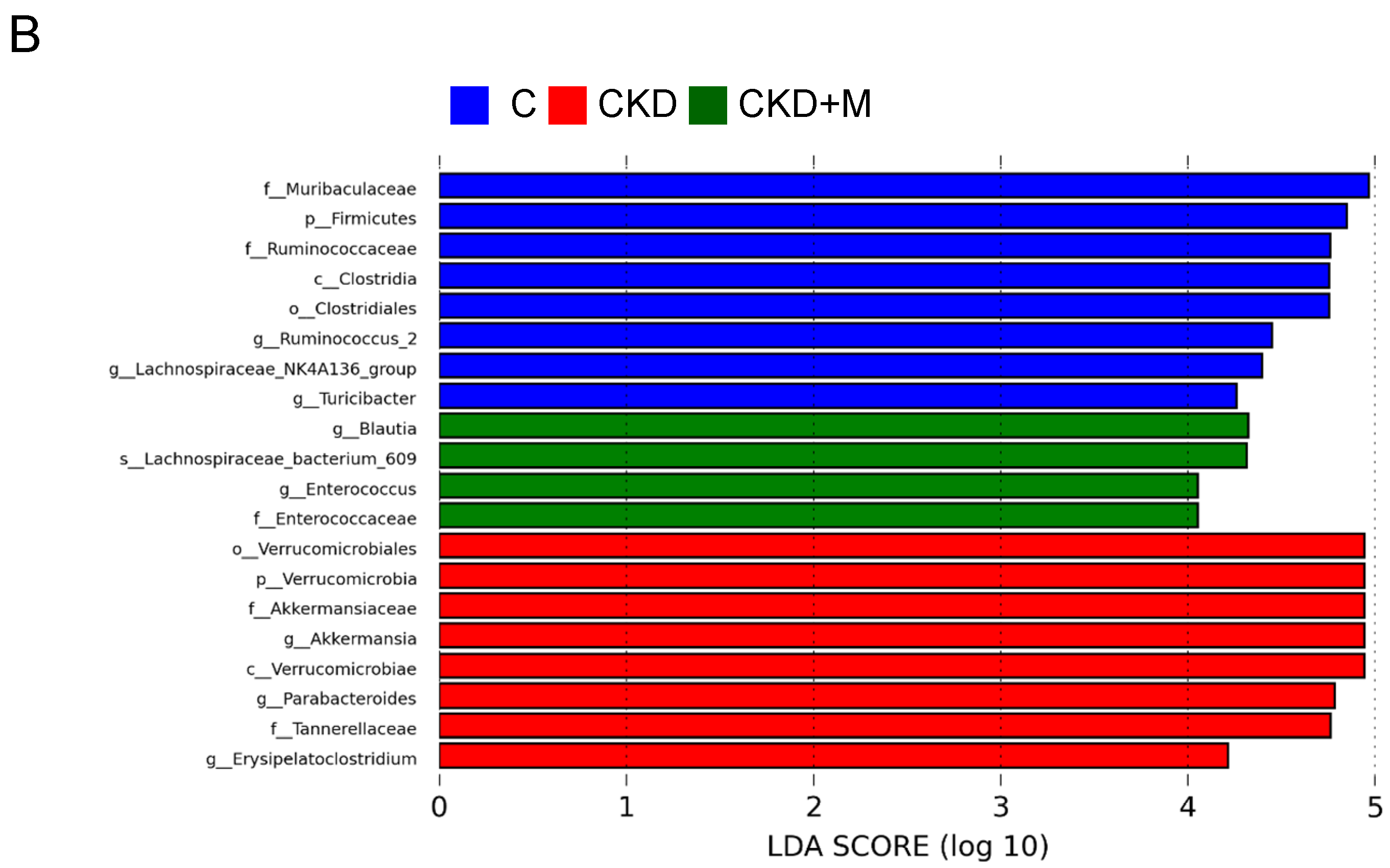
3.4. Gut Microbiota Compositions
3.5. NO-Related Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luyckx, V.A.; Tonelli, M.; Stanifer, J.W. The global burden of kidney disease and the sustainable development goals. Bull. World Health Organ. 2018, 96, 414–422. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Hsu, C.-N. Developmental Origins of Chronic Kidney Disease: Should We Focus on Early Life? Int. J. Mol. Sci. 2017, 18, 381. [Google Scholar] [CrossRef] [Green Version]
- Ingelfinger, J.R.; Kalantar-Zadeh, K.; Schaefer, F.; World Kidney Day Steering Committee. World Kidney Day 2016: Averting the legacy of kidney disease-focus on childhood. Pediatr. Nephrol. 2016, 31, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Diwan, V.; Brown, L.; Gobe, G.C. Adenine-induced chronic kidney disease in rats. Nephrology 2018, 23, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claramunt, D.; Gil-Peña, H.; Fuente, R.; Hernández-Frías, O.; Santos, F. Animal models of pediatric chronic kidney disease. Is adenine intake an appropriate model? Nefrologia 2015, 35, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, A.; Raj, D.S. The gut microbiome, kidney disease, and targeted interventions. J. Am. Soc. Nephrol. 2014, 25, 657–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graboski, A.L.; Redinbo, M.R. Gut-Derived Protein-Bound Uremic Toxins. Toxins 2020, 12, 590. [Google Scholar] [CrossRef]
- Schiattarella, G.G.; Sannino, A.; Toscano, E.; Giugliano, G.; Gargiulo, G.; Franzone, A.; Trimarco, B.; Esposito, G.; Perrino, C. Gut microbe-generated metabolite trimethylamine-N-oxide as cardiovascular risk biomarker: A systematic review and dose-response meta-analysis. Eur. Heart J. 2017, 38, 2948–2956. [Google Scholar] [CrossRef] [Green Version]
- Jaworska, K.; Hering, D.; Mosieniak, G.; Bielak-Zmijewska, A.; Pilz, M.; Konwerski, M.; Gasecka, A.; Kapłon-Cieślicka, A.; Filipiak, K.; Sikora, E.; et al. TMA, A Forgotten Uremic Toxin, but Not TMAO, Is Involved in Cardiovascular Pathology. Toxins 2019, 11, 490. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Maternal resveratrol therapy protected adult rat offspring against hypertension programmed by combined exposures to asymmetric dimethylarginine and trimethylamine N-oxide. J. Nutr. Biochem. 2021, 93, 108630. [Google Scholar] [CrossRef]
- Sun, S.; Lulla, A.; Sioda, M.; Winglee, K.; Wu, M.C.; Jacobs, D.R., Jr.; Shikany, J.M.; LloydJones, D.M.; Launer, L.J.; Fodor, A.A.; et al. Gut microbiota composition and blood pressure. Hypertension 2019, 73, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Adenine-Induced Chronic Kidney Disease Programs Hypertension in Adult Male Rat Offspring: Implications of Nitric Oxide and Gut Microbiome Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Regulation of Nitric Oxide Production in the Developmental Programming of Hypertension and Kidney Disease. Int. J. Mol. Sci. 2019, 20, 681. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Huang, L.T.; Lin, I.C.; Lau, Y.T.; Lin, C.Y. Melatonin prevents hypertension and increased asymmetric dimethylarginine in young spontaneous hypertensive rats. J. Pineal Res. 2010, 49, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Hrenak, J.; Paulis, L.; Repova, K.; Aziriova, S.; Nagtegaal, E.J.; Reiter, R.J.; Simko, F. Melatonin and renal protection: Novel perspectives from animal experiments and human studies (review). Curr. Pharm. Des. 2015, 21, 936–949. [Google Scholar] [CrossRef]
- Tain, Y.L.; Sheen, J.M.; Yu, H.R.; Chen, C.C.; Tiao, M.M.; Hsu, C.N.; Lin, Y.J.; Kuo, K.C.; Huang, L.T. Maternal Melatonin Therapy Rescues Prenatal Dexamethasone and Postnatal High-Fat Diet Induced Programmed Hypertension in Male Rat Offspring. Front. Physiol. 2015, 6, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal N(G)-Nitro-L-arginine-methyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636. [Google Scholar] [CrossRef]
- Hsu, C.N.; Chan, J.Y.H.; Wu, K.L.H.; Yu, H.R.; Lee, W.C.; Hou, C.Y.; Tain, Y.L. Altered Gut Microbiota and Its Metabolites in Hypertension of Developmental Origins: Exploring Differences between Fructose and Antibiotics Exposure. Int. J. Mol. Sci. 2021, 22, 2674. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Wagner, B.D.; Grunwald, G.K.; Zerbe, G.O.; Mikulich-Gilbertson, S.K.; Robertson, C.E.; Zemanick, E.T.; Harris, J.K. On the use of diversity measures in longitudinal sequencing studies of microbial communities. Front. Microbiol. 2018, 9, 1037. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Paulis, L.; Simko, F. Blood pressure modulation and cardiovascular protection by melatonin: Potential mechanisms behind. Physiol. Res. 2007, 56, 671–684. [Google Scholar]
- Baker, J.; Kimpinski, K. Role of melatonin in blood pressure regulation: An adjunct anti-hypertensive agent. Clin. Exp. Pharmacol. Physiol. 2018, 45, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.; Soares, J.M., Jr.; Gallo, C.C.; Furtado, A.; Cavaco, J.E.; Gonçalves, I.; Santos, C.R.A.; Quintela, T. The crosstalk between melatonin and sex steroid hormones. Neuroendocrinology 2021. [Google Scholar] [CrossRef] [PubMed]
- Iseki, K. Gender differences in chronic kidney disease. Kidney Int. 2008, 74, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Fan, Y.; Li, A.; Shen, Q.; Wu, J.; Ren, L.; Lu, H.; Ding, S.; Ren, H.; Liu, C.; et al. Alterations of the Human Gut Microbiome in Chronic Kidney Disease. Adv. Sci. 2020, 7, 2001936. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Ducatelle, R.; Eeckhaut, V.; Haesebrouck, F.; Van Immerseel, F. A review on prebiotics and probiotics for the control of dysbiosis: Present status and future perspectives. Animal 2015, 9, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Gu, Y.; Li, X.; Yang, W.; Jia, L.; Chen, C.; Han, X.; Huang, Y.; Zhao, L.; Li, P.; et al. Alterations of the Gut Microbiome in Hypertension. Front. Cell Infect. Microbiol. 2017, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, C.C.; Croyal, M.; Ene, L.; Aguesse, A.; Billon-Crossouard, S.; Krempf, M.; Lemoine, S.; Guebre-Egziabher, F.; Juillard, L.; Soulage, C.O. Elevation of Trimethylamine-N-Oxide in Chronic Kidney Disease: Contribution of Decreased Glomerular Filtration Rate. Toxins 2019, 11, 635. [Google Scholar] [CrossRef] [Green Version]
- Papandreou, C.; Moré, M.; Bellamine, A. Trimethylamine N-Oxide in Relation to Cardiometabolic Health-Cause or Effect? Nutrients 2020, 12, 1330. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Jameson, E.; Doxey, A.C.; Airs, R.; Purdy, K.J.; Murrell, J.C.; Chen, Y. Metagenomic data-mining reveals contrasting microbial populations responsible for trimethylamine formation in human gut and marine ecosystems. Microb. Genom. 2016, 2, e000080. [Google Scholar] [CrossRef] [Green Version]
- Pluznick, J.L. Microbial Short-Chain Fatty Acids and Blood Pressure Regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felizardo, R.J.F.; Watanabe, I.K.M.; Dardi, P.; Rossoni, L.V.; Câmara, N.O.S. The interplay among gut microbiota, hypertension and kidney diseases: The role of short-chain fatty acids. Pharmacol. Res. 2019, 141, 366–377. [Google Scholar] [CrossRef]
- Dan, X.; Mushi, Z.; Baili, W.; Han, L.; Enqi, W.; Huanhu, Z.; Shuchun, L. Differential Analysis of Hypertension-Associated Intestinal Microbiota. Int. J. Med. Sci. 2019, 16, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Palmu, J.; Salosensaari, A.; Havulinna, A.S.; Cheng, S.; Inouye, M.; Jain, M.; Salido, R.A.; Sanders, K.; Brennan, C.; Humphrey, G.C.; et al. Association Between the Gut Microbiota and Blood Pressure in a Population Cohort of 6953 Individuals. J. Am. Heart Assoc. 2020, 9, e016641. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Toxic Dimethylarginines: Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA). Toxins 2017, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.T. Melatonin therapy prevents programmed hypertension and nitric oxide deficiency in offspring exposed to maternal caloric restriction. Oxid. Med. Cell Longev. 2014, 2014, 283180. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.C.; Wu, T.H.; Huang, L.T.; Tain, Y.L. Renoprotective effects of melatonin in young spontaneously hypertensive rats with L-NAME. Pediatr. Neonatol. 2014, 55, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Huang, L.T.; Tain, Y.L. Perinatal Use of Melatonin for Offspring Health: Focus on Cardiovascular and Neurological Diseases. Int. J. Mol. Sci. 2019, 20, 5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, R.M.; Calhoun, D.A.; Bakris, G.L.; Brook, R.D.; Daugherty, S.L.; Dennison-Himmelfarb, C.R.; Egan, B.M.; Flack, J.M.; Gidding, S.S.; Judd, E.; et al. Resistant Hypertension: Detection, Evaluation, and Management: A Scientific Statement from the American Heart Association. Hypertension 2018, 72, e53–e90. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Reiter, R.J.; Paulis, L. Melatonin as a rational alternative in the conservative treatment of resistant hypertension. Hypertens. Res. 2019, 42, 1828–1831. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | C | CKD | CKD + M |
|---|---|---|---|
| Male | N = 8 | N = 8 | N = 8 |
| Body weight (BW) (g) | 360 ± 8 | 261 ± 11 * | 237 ± 8 * |
| Left kidney weight (g) | 1.78 ± 0.07 | 5.1 ± 0.44 * | 4.09 ± 0.27 *# |
| Left kidney weight/100 g BW | 0.49 ± 0.02 | 1.94 ± 0.12 * | 1.73 ± 0.01 *# |
| Systolic blood pressure (mmHg) | 128 ± 1 | 144 ± 1 * | 132 ± 1 # |
| Diastolic blood pressure (mmHg) | 86 ± 1 | 87 ± 1 | 87 ± 2 |
| Mean arterial pressure (mmHg) | 100 ± 1 | 106 ± 1 * | 102 ± 1 |
| Creatinine (μM) | 22.7 ± 1 | 322.7 ± 25.7 * | 235.9 ± 22.4 *# |
| Female | N = 8 | N = 8 | N = 8 |
| Body weight (BW) (g) | 244 ± 11 | 187 ± 6 * | 174 ± 5 * |
| Left kidney weight (g) | 1.12 ± 0.03 | 2.12 ± 0.12 * | 1.91 ± 0.13 * |
| Left kidney weight/100 g BW | 0.46 ± 0.02 | 1.14 ± 0.05 * | 1.09 ± 0.05 * |
| Systolic blood pressure (mmHg) | 118 ± 1 | 132 ± 1 * | 121 ± 1 # |
| Diastolic blood pressure (mmHg) | 75 ± 3 | 84 ± 3 * | 81 ± 1 * |
| Mean arterial pressure (mmHg) | 89 ± 2 | 100 ± 2 * | 94 ± 1 # |
| Creatinine (μM) | 23.4 ± 1.3 | 220.7 ± 23.5* | 190 ± 18.2 * |
| Groups | C | CKD | CKD + M |
|---|---|---|---|
| Male | N = 8 | N = 8 | N = 8 |
| TMA (ng/mL) | 455 ± 18 | 466 ± 26 | 786 ± 65 *# |
| TMAO (ng/mL) | 301 ± 18 | 1427 ± 92 * | 1376 ± 164 * |
| DMA (ng/mL) | 111 ± 5 | 902 ± 95 * | 809 ± 84 * |
| TMAO-to-TMA ratio | 0.67 ± 0.05 | 3.13 ± 0.27 * | 1.78 ± 0.19 *# |
| DMA-to-TMAO ratio | 0.38 ± 0.02 | 0.63 ± 0.05 * | 0.62 ± 0.07 * |
| Female | N = 8 | N = 8 | N = 8 |
| TMA (ng/mL) | 1653 ± 291 | 1877 ± 253 | 3543 ± 185 *# |
| TMAO (ng/mL) | 424 ± 68 | 2037 ± 683 * | 1453 ± 222 * |
| DMA (ng/mL) | 202 ± 34 | 755 ± 51 * | 931 ± 56 * |
| TMAO-to-TMA ratio | 0.3 ± 0.05 | 1.13 ± 0.31 * | 0.41 ± 0.06 # |
| DMA-to-TMAO ratio | 0.47 ± 0.02 | 0.5 ± 0.06 | 0.7 ± 0.07 * |
| Groups | C | CKD | CKD + M |
|---|---|---|---|
| Male | N = 8 | N = 8 | N = 8 |
| Acetate, mM/gm feces | 7.68 ± 0.53 | 5.49 ± 0.6 * | 5.25 ± 0.35 * |
| Propionate, mM/gm feces | 2.99 ± 0.22 | 1.45 ± 0.44 * | 1.49 ± 0.24 * |
| Butyrate, mM/gm feces | 3.82 ± 0.66 | 1.15 ± 0.24 * | 1.08 ± 0.15 * |
| Female | N = 8 | N = 8 | N = 8 |
| Acetate, mM/gm feces | 7.04 ± 0.95 | 3.93 ± 0.46 * | 3.95 ± 0.35 * |
| Propionate, mM/gm feces | 2.73 ± 0.35 | 0.91 ± 0.27 * | 0.54 ± 0.09 * |
| Butyrate, mM/gm feces | 5.1 ± 0.71 | 1.06 ± 0.14 * | 0.7 ± 0.09 * |
| Groups | C | CKD | CKD + M |
|---|---|---|---|
| Male | N = 8 | N = 8 | N = 8 |
| L-citrulline (µM) | 53.7 ± 1.6 | 47.6 ± 1.1 | 46.7 ± 1 |
| L-arginine (µM) | 165.8 ± 2.8 | 146.3 ± 4 | 118.8 ± 1.4 * |
| ADMA (µM) | 1.65 ± 0.02 | 2.4 ± 0.04 * | 1.96 ± 0.05 # |
| SDMA (µM) | 1.21 ± 0.01 | 1.69 ± 0.04 * | 1.61 ± 0.02 * |
| L-arginine-to-ADMA ratio (µM/µM) | 102.4 ± 2.9 | 66 ± 1.9 * | 58.8 ± 1.6 * |
| Female | N = 8 | N = 8 | N = 8 |
| L-citrulline (µM) | 56.1 ± 0.9 | 53.7 ± 0.8 | 55.8 ± 1.2 |
| L-arginine (µM) | 166.4 ± 2.5 | 160.9 ± 4.1 | 140.4 ± 3.2 |
| ADMA (µM) | 1.38 ± 0.03 | 2.1 ± 0.07 * | 2.16 ± 0.05 * |
| SDMA (µM) | 1.58 ± 0.07 | 2.13 ± 0.05 | 1.72 ± 0.02 |
| L-arginine-to-ADMA ratio (µM/µM) | 123.1 ± 3.4 | 83.2 ± 3.9 * | 65.8 ± 1.2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-N.; Yang, H.-W.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites. Antioxidants 2021, 10, 1211. https://doi.org/10.3390/antiox10081211
Hsu C-N, Yang H-W, Hou C-Y, Chang-Chien G-P, Lin S, Tain Y-L. Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites. Antioxidants. 2021; 10(8):1211. https://doi.org/10.3390/antiox10081211
Chicago/Turabian StyleHsu, Chien-Ning, Hung-Wei Yang, Chih-Yao Hou, Guo-Ping Chang-Chien, Sufan Lin, and You-Lin Tain. 2021. "Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites" Antioxidants 10, no. 8: 1211. https://doi.org/10.3390/antiox10081211