
Redox Regulation in Diazotrophic Bacteria in Interaction with Plants
by
 , ,
, ,
Karine Mandon
1, 
Fanny Nazaret
1,
Davoud Farajzadeh
2,3,
Geneviève Alloing
1 and
Pierre Frendo
1,* 1
Université Côte d’Azur, INRAE, CNRS, ISA, 06903 Sophia Antipolis, France
2
Department of Biology, Faculty of Basic Sciences, Azarbaijan Shahid Madani University, Tabriz 5375171379, Iran
3
Center for International Scientific Studies and Collaboration (CISSC), Ministry of Science, Research and Technology, Tehran 158757788, Iran
*
Author to whom correspondence should be addressed.
Antioxidants 2021, 10(6), 880; https://doi.org/10.3390/antiox10060880
Submission received: 9 May 2021
/
Revised: 26 May 2021
/
Accepted: 26 May 2021
/
Published: 30 May 2021
(This article belongs to the Special Issue Redox Biology in Microorganisms)
Abstract
:Plants interact with a large number of microorganisms that greatly influence their growth and health. Among the beneficial microorganisms, rhizosphere bacteria known as Plant Growth Promoting Bacteria increase plant fitness by producing compounds such as phytohormones or by carrying out symbioses that enhance nutrient acquisition. Nitrogen-fixing bacteria, either as endophytes or as endosymbionts, specifically improve the growth and development of plants by supplying them with nitrogen, a key macro-element. Survival and proliferation of these bacteria require their adaptation to the rhizosphere and host plant, which are particular ecological environments. This adaptation highly depends on bacteria response to the Reactive Oxygen Species (ROS), associated to abiotic stresses or produced by host plants, which determine the outcome of the plant-bacteria interaction. This paper reviews the different antioxidant defense mechanisms identified in diazotrophic bacteria, focusing on their involvement in coping with the changing conditions encountered during interaction with plant partners.
1. Introduction
Diazotrophic rhizobacteria that interact positively with plants involve endophytic bacteria and rhizobial endocellular symbionts. Their benefit on plant growth has been attributed to a variety of single or combined mechanisms, including phosphate solubilization, siderophore synthesis, production, and secretion of several phytohormones as well as biological nitrogen (N2) fixation (BNF), leading to enhanced plant nutrition [1]. Nitrogen (N) is one of the major macronutrients for plants, which plays a fundamental role in protein synthesis and energy metabolism through photosynthesis, and its deficiency affects plant development [2]. The addition of chemical fertilizers is a major source of N for crops, which not only requires fossil fuels for their production but increases water pollution as well. Contributing to more than half of total crop N demand, BNF is a possible alternative to fertilizers [3]. The BNF is exclusively performed by prokaryotes, archaea, and bacteria, collectively known as diazotrophs. For archaea, BNF is performed by diazotrophic methanogens, which produce methane [4]. Diazotrophic bacteria encode a specific protein complex, the nitrogenase that reduces the atmospheric N2 to ammonia (NH3). The diazotrophs such as Azotobacter sp., Azospirillum sp., Bacillus sp., Cyanobacteria sp., and Clostridum sp. are free-living bacteria present in the rhizosphere, which can form endophytic interactions with a wide variety of plants (see Figure 1) [5]. In parallel, endosymbiotic diazotrophs reside intracellularly in living plant cells and induce the formation of differentiated plant structures as in the case of Nostoc sp. with the Gunneraceae family [6], Frankia sp. with the actinorhizal plants [7], and the Rhizobia sp. with the Fabaceae family [8]. In the two latter cases, the bacteria are hosted in the stem or root nodules that are the neoformed plant organs involved in BNF.
In plants and microbes interactions, the plant immune system is differentially modulated by the microorganisms, depending on the type of interaction [10,11]. Upon infection, plant receptors recognize a wide range of conserved bacterial molecules, referred to as microbe-associated molecular patterns (MAMPS) allowing the induction of plant defense mechanisms [12]. As part of the defense responses that follow MAMP receptor activation, oxidative and nitrosative bursts occur, both to control microorganism colonization and to spread a systemic defense signal throughout the plant [13]. The interacting bacteria have evolved systems to suppress plant defense reactions; they concomitantly have set up systems to cope with plant antimicrobial compounds, in particular non-enzymatic antioxidant molecules and Reactive Oxygen Species (ROS)/Reactive Nitrogen Species (RNS) detoxification enzymes (review in [14]).
Whereas oxidative burst during plant-pathogen interactions has been extensively described, studies concerning the modulation of redox activities in plants interacting with beneficial microbes remain sparse. However, modulation of antioxidant enzyme activities has been observed during some interactions between plants and plant growth-promoting rhizobacteria (PGPR) [15,16]. Analysis of sugarcane root transcriptome showed that many genes involved in the redox process were upregulated and that peroxidase and catalase enzyme activities were also significantly upregulated in response to the Burkholderia anthina MYSP113 [16]. Similarly, up-regulation of genes encoding ROS scavenging enzymes was observed in Arabidopsis, rice, and tomato roots inoculated with Azospirillum brasilense [17,18]; peroxidase accumulation was also observed in rice roots inoculated with Sinorhizobium meliloti [19]. In contrast, a decrease in the expression of superoxide dismutase, catalase and ascorbate peroxidase genes is detected in maize roots inoculated with A. brasilense [20], and inoculation of wheat with A. brasilense leads to limited production of O2•− from the root plant [21]. Altogether, these data suggest a complex pattern of adaptation during plant colonization as observed for the rhizobial infection of legumes.
Besides the endophytic interactions, plant-rhizobia interactions involving an intracellular symbiosis have been more deeply characterized. The mutualistic interaction of rhizobia with legume plants involves two major biological processes. The first one is the entry of the bacteria into the plant, followed by the bacterial transfer to the nodule primordia, then entering into plant cells in the nodule infection zone. In compatible interactions, the nodulation (Nod) factors excreted by the bacterial partner are recognized by the host plant receptors and elicit host responses such as root hair curling and invasion by the bacteria. During these early steps, there is production of ROS involved in the specific recognition of the bacteria [22,23]. The production of ROS has been detected a few minutes after Nod factor treatment [24]. The transfer to the inner cortex cells is performed through the infection thread (IT), a transcellular apoplastic compartment, which grows in parallel with the bacterial cell division [25]. Inhibition of ROS production impairs the root hair curling and the IT formation [26]. Respiratory Burst Oxidase Homolog Gene (RBOH) A is crucial for rhizobium infection in the common bean, suggesting that NADPH oxidases are responsible for the ROS production in the symbiotic interactions [27]. ROS production is also involved in the oxidative cross-linking of the IT matrix that allows an efficient transfer of the dividing bacteria from the plant epidermis to the inner cortex [28,29]. In addition to this general infection model, some rhizobia enter the plant through a mechanism called crack entry, which is a characteristic feature of some subtropical legumes belonging to the dalbergoid/genistoid clades [30]. This infection process involves the formation of infection pockets that was shown to be associated with localized plant cell death and the production of large amounts of ROS [31].
The second process is the differentiation of plant and bacterial cells, which occurs in the infected cells of the nodule to allow the setup of BNF. In the plant cell, bacteria are surrounded by the symbiosome membrane, which derives from the infected cell plasma membrane. At this time point, bacteria undergo a differentiation process allowing the formation of N-fixing bacteroids that release ammonia into the host cell in exchange for reduced carbon and other nutrients from the plant. Some leguminous plants, such as the ones from the inverted repeat-lacking clade, including Medicago truncatula, induce terminal differentiation of their symbionts, which prevents their division before they fix nitrogen. In this plant model, the production of hydrogen peroxide (H2O2) has been observed in the infection zone [32]. Besides the production of ROS in the infection zone, modification of the bacterial redox balance is also associated with the production of Nodule Cysteine Rich (NCR) peptides that are specifically expressed by the plant partners in the nodule infection and nitrogen-fixing zones [8,33]. The biological activity of these peptides is regulated by their redox state [34,35], and specific plant thioredoxins targeted to the bacteroids are able to modulate their redox state and the bacteroid differentiation [36].
Once differentiated, the bacteroid reduces N2 to NH3 in the nodule nitrogen-fixing zone (NFZ), where microoxic conditions prevail to allow nitrogenase functioning. These conditions involve the presence of a structural barrier in the nodule cortex and the expression of leghemoglobins in the NFZ [37]. The lower level of oxygen does not abolish the functioning of NADPH Oxidase in this NFZ. In M. truncatula, MtRBOHA is specifically expressed in the NFZ, and its activity was involved in nitrogen fixation efficiency [38]. Recently, four RBOHs from soybean (Glycine max) showed strong expression in nodules, pointing to their probable involvement in the nodulation process [39]. The importance of redox regulation in the plant symbiotic partner is also noticeable by the significantly higher level of antioxidant molecules in nodules than in roots [40]. Modification of the glutathione content in the plant partner affects both the nodule formation and functioning [41,42] showing that the plant antioxidant defense is crucial for the setup and the functioning of the nitrogen-fixing symbiosis. Similarly, methionine sulfoxide reductase B has been shown to play an essential role in the nodule development and functioning in the symbiotic interaction between Astragalus sinicus and Mesorhizobium [43].
To cope with the redox modifications that occur during endophytic and endosymbiotic interactions with plants, bacteria need to adapt to their redox homeostasis. The review presents some general aspects of endogenous antioxidant systems in bacteria and then summarizes the recent knowledge concerning redox regulation mechanisms in diazotrophic bacteria during beneficial plant-bacterial interactions.
2. The Bacterial Response to ROS
Aerobic bacteria have to cope with endogenous ROS derived from their metabolism. Exposure to abiotic stresses such as heat, low pH or salt, can also induce the formation of intracellular ROS [44]. In addition to these endogenous sources of redox stress, bacteria interacting with eukaryotic hosts are exposed to ROS produced by the hosts. These molecules are deleterious to biological macromolecules, including proteins, lipids, and DNA, and their concentration must be tightly controlled [45]. When present at the proper concentrations, ROS can function as signal molecules and activate multiple signal transduction pathways within the bacterial cells [46,47]. To maintain their intracellular redox homeostasis, bacteria use enzymatic and non-enzymatic defense systems. Both constitutive and inducible mechanisms are triggered by bacteria to manage the cellular redox environment and adapt to different lifestyles.
Bacteria protect themselves from ROS by producing various scavenging enzymes. For instance, the metalloenzyme superoxide dismutase (SOD) converts O2•− to H2O2, which is reduced to H2O by enzymes such as monofunctional catalases, bifunctional catalase-peroxidase (Kat), or thiol peroxidases such as peroxiredoxins (Prx) (Figure 2). The peroxiredoxin family has been divided into two categories, the organic hydroperoxide resistance protein Ohr, which displays a high affinity for long-chain fatty acid peroxides, and the TSA/AhpC subgroup containing 1-cys, 2-cys peroxiredoxins, and the AhpCD system that have a broad range of substrate including H2O2, organic and fatty acid peroxides. ROS can also oxidize methionine residues of proteins to methionine sulfoxide, thus altering protein structure. The dedicated methionine sulfoxide reductases (Msr) enzymes reduce methionine sulfoxide residues back to methionine [48].
Low molecular weight thiol-redox buffers also function in the detoxification of ROS. Tripeptide glutathione (γ-glutamyl-cysteinyl-glycine, GSH), the major thiol reductant in eukaryotes and many bacteria, participates in the intracellular defense against ROS-induced oxidative damage and maintains the intracellular compartment in a reduced state. GSH can remove free radicals, e.g., superoxide anion, hydroxyl radical, nitric oxide, and carbon radicals via direct interaction. It is also a cofactor for various antioxidant enzymes such as glutaredoxins (Grx) and glutathione-S-transferase (GST) to detoxify ROS, xenobiotics and/or heavy metals [49,50]. The resulting oxidized form of glutathione (GSSG) is reduced to GSH by the NAPDH-using enzyme glutathione reductase (GR). GSH is synthesized in a two steps procedure. The first step, in which glutamate is ligated with cysteine to form γ-glutamylcysteine, is catalyzed by the γ-glutamylcysteine synthetase (γ-ECS) encoded by gshA. GSH synthesis is then completed by the addition of glycine to γ-glutamylcysteine, which is mediated by the glutathione synthetase (GSHS) encoded by gshB. Under oxidative-stress conditions, GSH can form reversible mixed disulfides with reactive protein thiols, protecting them against over-oxidation. This S-glutathionylation can serve to transduce a redox signal by modulating protein function [51]. S-deglutathionylation, that is, the reduction of GSH-protein mixed disulfides is catalyzed by Grxs in a coupled system with GR and NADPH as an electron donor. Grxs together with Trxs are also involved in the reduction of protein disulfides. The oxidized Trx is reduced by thioredoxin reductase, receiving electrons from NADPH (Figure 2).
3. Redox Regulation in Rhizosphere and Root Colonization
Endophytic and endosymbiotic bacteria live in the rhizosphere in close association with the root surface, where plant-derived compounds such as amino acids or carbohydrates favor the proliferation of heterotrophic aerobic bacteria. It is also a place where bacteria have to deal with abiotic stresses such as osmotic and pH alterations or the presence of toxic compounds. Some root exudates are specific to particular plant species, like original carbon and nitrogen sources or secondary metabolic compounds such as flavonoids [52]. Despite their capacity to attenuate the induction of the plant defense, endophytic and endosymbiotic bacteria have to face oxidative stress during root adhesion and penetration. Adaptation to the rhizosphere is thus critical for the efficiency of root colonization and the outcome of plant-bacteria interactions.
Azospirillum sp. is a well-characterized PGPB due to its capacity to colonize the roots of major cereal crops and grasses. Different genetic determinants have been proposed to contribute to the large host range of Azospirillum sp. and may account for the niche-specific adaptation of the bacteria to a given rhizosphere. In particular, genes related to ROS detoxification and multidrug efflux seem to be involved in A. lipoferum 4B adaptation to the rice rhizosphere [53,54]. A comparative analysis of Azospirillum sp. genomes shows that several enzymes involved in the oxidative stress response differ among Azospirillum species [55]. As an example, only one gene encoding a monofunctional catalase was identified in Azospirillum sp. B510, while two genes, one encoding catalase and the other encoding a bifunctional catalase peroxidase, were encountered in A. lipoferum 4B. A comparative genome-wide transcriptome analysis performed with the A. lipoferum strain 4B following inoculation on original host and non-natural host rice cultivars, highlights common and cultivar-dependent transcriptomic responses [53]. During interaction with both cultivars, genes involved in redox homeostasis encoding an Ohr-like protein, a Grx, and a GST, were upregulated. Similarly, the expression of genes encoding methionine sulfoxide reductases (msrA and msrB) and a 2Fe-2S ferredoxin-like protein was also enhanced. By contrast, a specific induction of thioredoxin-encoding genes (trxB and trxC) and sod-like gene was observed upon inoculation of the non-host cultivar. The latter result supports the idea that bacteria interacting with a less adapted host perceive stronger oxidative stress suggesting a better adaptation of A. lipoferum 4B towards its original host cultivar. RNA-seq analysis of A. brasilense FP2 on wheat revealed induction of a sod-like gene (sodB), suggesting the activation of the antioxidant defense during the interaction with plants [56]. In addition, the A. brasilense Sp245 genome contains an ahpC gene that was shown to be involved in resistance towards H2O2 and organic alkyl hydroperoxide stress [57]. However, the ahpC inactivated mutant is not impaired in the colonization of wheat roots, even in competitive experiments with the wild-type strain. The multiplicity of ROS scavenging enzymes in the bacteria might be the reason for the absence of phenotype. Indeed A. brasilense sp7 contains three catalase-encoding genes, katAI, katAII, and katN [58].
“Omics” analyses were also performed in endophytes other than Azospirillum sp, and results concerning the importance of the antioxidant defense of the studied bacterial strains are not always conclusive. The genome of Leifsonia sp. Ku-ls isolated from rice presents an elaborate antioxidant defense [59]. A proteome analysis has been performed on Herbaspirillum seropedica, a bacterium able to colonize roots of gramineous plants of economic importance, such as wheat, maize, rice, or sugarcane (review in [5]). As protein secretion plays a fundamental role in the adaptation of bacteria to its environment and during plant-bacteria interaction, the secretome of H. seropedicae was performed using 2D electrophoresis [60]. Among the 41 proteins, the most important secreted protein was an NADPH:quinone oxidoreductase, and to a lesser extent, a peroxiredoxin and a Fe-superoxide dismutase (SOD). However, the secretome was extracted from bacteria in laboratory culture conditions and the presence of Fe-SOD during plant-bacteria interaction has not been investigated. In Gluconoacetobacter diazotrophicus, ROS level is increased in nitrogen-fixing cells, where a concomitant induction of genes related to the antioxidant defense, sodA, katE, kat, katC, and genes involved in glutathione cycle was emphasized [61]. Following inoculation on rice, the expression of genes encoding catalases, SOD, and GR was shown to be induced from 30 min to 4 days post-inoculation, and the bacterial mutants sod and gr were poorly able to colonize rice (Table 1) [62].
Several transcriptomic approaches have also been performed to identify genes of endosymbiotic bacteria potentially involved in adaptation to the rhizosphere. Gene induction pattern in the rhizosphere or response to root secretions was compared against laboratory culture conditions, and numerous genes of the antioxidant defense in various rhizobia were thereby found to be upregulated. An efficient antioxidant system may be a prerequisite for symbiotic bacteria to deal with ROS in the rhizosphere and root surfaces, providing the required physiological adaptation to thrive in the plant root system. In one of these studies, the early adaptation of Rhizobium leguminosarum to the rhizospheres of pea (host-legume), alfalfa (non-host legume), and sugar beet (non-legume) was investigated in parallel [85]. This approach allows identifying genes specifically induced in the pea rhizosphere (138 genes), whereas others were found to be upregulated in all rhizospheres (106 genes). Some genes involved in response to oxidative stress belong to this common core of rhizosphere-induced genes, such as genes encoding MsrA and MsrB, a thioredoxin and a peroxiredoxin. Another example is a comparative analysis of the transcriptomes of three Mimosa symbionts, the α–proteobacterium Rhizobium mesoamericanum and the two β-proteobacteria Bukholderia phymatum, an ancient symbiont, and Cupriavidus taiwanensis, which has probably acquired symbiotic genes from a Burkholderia symbiont more recently. The upregulation of genes encoding putative peroxiredoxin, thioredoxin, and GST was shown to be part of the common response of the two β-proteobacteria to Mimosa root exudates, highlighting the importance of antioxidant defense in bacterial survival in the rhizosphere [86]. In addition to transcriptomic approaches, genome-wide approaches based on STM (signature-tagged mutagenesis) mutants were performed in S. meliloti and R. leguminosarum to identify genes important for rhizosphere colonization [72,87]. They showed that competitiveness in the rhizosphere is largely determined by metabolic functions and motility and by tolerance to abiotic stresses generated by root exudates as well [87]. Of the 170 R. leguminosarum genes involved in pea rhizosphere competitiveness, sodB is specifically involved in survival in the rhizosphere, a Grx-encoding gene is necessary for both rhizosphere bacterial growth and root colonization, while gshA and gshB are required for rhizosphere competitiveness and onward stages of symbiosis as well [72].
4. Antioxidant Defense in Legume Symbionts
4.1. ROS Scavenging Enzymes
A complex pattern of ROS production is required for root colonization by rhizobia and nodule development (reviewed in [88,89]). Accumulation of ROS during plant colonization, nodule development, and functioning has been highlighted in Sesbania rostrata, Phaseolus vulgaris and M. truncatula [31,90,91,92]. The microsymbiont also participates in the fine-tuning of ROS concentration during symbiosis. For example, using a pharmacological approach, Bradyrhizobium sp. peroxidase activity has been shown to be necessary for the early events of the interaction with Arachis hypogaea [93], while a S. meliloti strain that degrades H2O2 very efficiently is affected in plant colonization [28].
Many rhizobia contain multiple ROS detoxifying enzymes that are required for symbiosis (Table 1). The role of rhizobial antioxidant defense has been first characterized in S. meliloti. This bacterium possesses two SODs, SodA and SodM, three heme b-containing catalases in addition to one heme-chloroperoxidase [64,94,95]. The SodM protein is expressed in nitrogen-fixing bacteroids, and mutants inactivated in sodA displayed different symbiotic phenotypes depending on the genotype of the strain used. A periplasmic SodA protein was characterized in Rhizobium leguminosarum bv. viciae [96] and a SodA purified from bacteroids appeared the most abundant SOD isoenzyme in pea nodules [97]. In a transcriptomic analysis of B. japonicum exposed to H2O2, three distinct sod genes, sodF, sodM and a Fe/Mn-SOD-encoding gene were upregulated [98]; however, their roles in planta have not been established. In S. meliloti, the three catalase genes, katA, katB, and katC, are differentially expressed in culture and symbiosis. The H2O2 inducible katA gene is expressed in the nitrogen-fixing zone, while the stress-inducible katC is strongly transcribed in ITs [65]. The disruption of both katB and katC genes leads to a drastic phenotype, with aborted bacteroid differentiation following plant cell release. Besides, the double mutant katAC is able to differentiate into bacteroids that undergo premature senescence. In parallel, the overexpression of the bifunctional catalase-peroxidase KatB, thereby acting as a sink for H2O2 inside the ITs, results in a delayed nodulation phenotype associated with aberrant infection threads [28]. Catalases have been shown to be essential during symbiosis in rhizobia, such as Mezorhizobium loti, which has a monofunctional catalase KatE and a bifunctional KatG [66]. KatG is required in a free-living state for H2O2 scavenging during the exponential growth phase or H2O2 exposure, while KatE acting during the stationary phase is required to sustain optimal nitrogenase activity in symbiosis with Lotus japonicus. Peroxiredoxins act in concert with the catalases for degrading H2O2. S. meliloti contains an Ohr protein induced in presence of alkyl peroxide, not essential for symbiosis, and an H2O2-inducible AhpC-like protein present in bacteroid extracts [99,100,101]. In addition to the H2O2-inducible catalase KatG, B. japonicum contains multiple genes involved in redox homeostasis whose products are present in bacteroid extracts, two SOD, one catalase, the AhpCD system, and two non-heme chloroperoxidases [102,103]. In Mezorhizobium huakii/A. sinikus interaction, the peroxiredoxin PrxA is important for symbiosis as a prxA mutant elicits spherical nodules displaying a considerable reduction of nitrogen fixation activity associated with bacteria that fail to differentiate [68]. Besides, R. etli contains both a catalase KatG and a 2-cys peroxiredoxin PrxS, whose inactivation gives no phenotype [67,104]. However, disruption of both katG and the prxS results in a reduction of 40% in nitrogen fixation, suggesting that a compensation mechanism may occur between multiple antioxidant overlapping enzymes [67]. Azorhizobium caulinodans is unique among rhizobia, as the bacterium is both able to assimilate N2 in a free-living state and to induce the formation of stem and root nodules on the tropical legume S.rostrata. A. caulinodans possesses the catalase-peroxidase KatG and the alkylhydroperoxide reductase AhpCD to cope with H2O2 increase, which are differentially used depending on the growth phase and H2O2 concentration [69,70]. In addition, protection against organic hydroperoxide stress requires the Ohr protein [71]. The katG, ahpC, and ohr single mutants displayed a significant reduction in stem nodule number associated with a reduced nitrogen fixation ability. In contrast, the S. rostrata root nodules were not affected by the katG or the ahpC inactivation, suggesting that a specific adaptation of A. caulinodans is required in stem nodules to face the high O2 and derivatives produced by the photosynthetic cortical cells close to the infected cells. Lastly, hydroperoxidase detoxifying enzymes also appear to play a significant role in actinorhizal symbiosis. Indeed, a transcriptomic analysis of Frankia alni during symbiosis with Alnus glutinosa showed that genes encoding a Prx and a SOD were upregulated in nodules as compared to free-living conditions, while the katA gene was highly expressed in both conditions [105].
4.2. The Role of Bacterial GSH/Grx System
The GSH biosynthetic pathway plays a crucial role in rhizobia during both free-living and plant-associated lifestyles. GSH was first shown to be essential in S. meliloti, from which a gshA mutant could not grow in a minimal medium. Conversely, a gshB mutant was able to divide, even at a lower rate than WT, suggesting that γ-EC could partially compensate for GSH defect [74]. A growth defect in gshB mutants was also observed in other rhizobia [73,76,77]. GSH deficiency was linked to metabolic changes in R. etli, reducing glutamine uptake through the activity of Aap and Bra transporters [76]; in R. leguminosarum, a gshB mutant is affected in the uptake of several carbon source compounds such as glucose, succinate, glutamine, and histidine [73]. In addition, gshB inactivation causes constitutive oxidative stress associated with a higher catalase activity in S. meliloti and an increased sensitivity to peroxides in M. huakuii [74,78].The lack of bacterial GSH compromises the early steps of interaction. Specifically, gshB mutants have a lower capacity to colonize the host plant in the interaction between R. leguminosarum and P. sativum and are impaired in nodulation during S. meliloti/ Medicago sp., R. etli/P. vulgaris, R. tropici/P. vulgaris and M. huakuii/A. sinicus interactions. Furthermore, GSH has been shown to be crucial in nodule functioning. In most cases, gshB mutants produced abnormal nodules with a strong nitrogen-fixation deficiency, correlated with the early senescence of nodules and bacteroids [74,75,76,77,78]. In line with these findings, the recycling of GSH by glutathione reductase was also shown to be involved in nodulation and in nitrogen fixation ability [79]. In R. tropici/P. vulgaris interaction, GSH deficiency was also associated with increased production of superoxide anion [77]. Regarding the interaction between S. meliloti and M. truncatula, a combination of physiological, and molecular markers, microscopic and flow cytometry analyses, showed that a gshB mutant undergoes complete differentiation before being engaged in an early senescence process [75]. Totally, these results underline the importance of the bacterial GSH/GSSG redox couple for adaptation to the host-plant environment and survival inside the host cells (Table 1).
The contribution of GSH-dependent redox enzymes to symbiosis efficiency was highlighted by studies on different Grxs (Table 1). Grxs are divided into two classes based on the active site motifs: dithiol Grxs (class I) contain a typical CxxC motif, while monothiol Grxs (class II) possess a CxxS motif. Grxs of the two classes are present in different rhizobia, and their particular role has been investigated in S. meliloti and A. caulinodans during free-living and symbiotic lifestyles [80,81]. In S. meliloti on one side, dithiol Grx1 has been shown to contribute to the deglutathionylation of proteins. A grx1 mutant has an intense growth defect and an increased sensitivity to H2O2 in bacterial cultures. During the interaction with M. truncatula, the grx1 mutant induces abortive nodules with infected cells containing no differentiated bacteroids. On the other side, the monothiol Grx2 of S. meliloti is involved in Fe metabolism. A grx2 mutant compared with the WT strain grows more slowly and contains an increased concentration of free Fe and decreased activity of Fe-S cluster containing enzymes. Inactivation of grx2 affects nodulation with M. truncatula and the nitrogen-fixing capacity of bacteroids. Those bacteroids are fully differentiated, which suggests that the mutation has a direct effect on the Fe-S cluster of nitrogenase. Likewise, mutants in the suf operon, involved in Fe/S cluster formation, also have a lowered nitrogen fixation capacity in S. meliloti and A. caulinodans. Furthermore, the suf operon of F. alni is one of the most up-regulated clusters in nodule bacteria compared to free-living cells [105,106,107].
The genome of A. caulinodans contains two Grxs of the class II (Grx1 and Grx2) and two Grxs of the class I (Grx3 and Grx4). Using a combination of single, double, and quadruple mutants, Cao and colleagues showed that genes encoding either monothiol or dithiol Grxs exhibit functional redundancies under normal growth conditions and in the presence of H2O2 and organic hydroperoxides [81]. The specific inactivation of the two monothiol Grxs results in lower GSH/GSSG ratio and Fe content, suggesting that they participate in Fe and redox homeostasis. In the symbiosis with S. rostrata, grx1 grx2 and grx3 grx4 double mutants have a reduced nitrogen fixation capacity, and the quadruple mutant, even more, showing a collective contribution by the two Grxs classes to the N2 fixation efficiency.
5. Redox-Based Transcriptional Regulation in Endophytic and Endosymbiotic Bacteria
ROS can cause reversible post-translational modifications and induce conformational changes of proteins, often by targeting thiol groups of redox-active cysteines. This mechanism contributes to redox signaling, notably by triggering activation or inactivation of redox-sensing transcriptional regulators (TR) [108]. In general, TRs are essential for bacteria to rapidly respond to environmental changes, improving their adaptation plasticity, cellular homeostasis, and colonization of new niches. Thiol-based redox regulators particularly play an essential role in the control of oxidative stress response and adaptation to the host-environment. Several have been characterized in rhizobacteria, and their role in symbiotic interaction was underlined in some cases.
OxyR is a redox-sensitive regulator of the LysR family, which is conserved in Gram+ and Gram- bacteria and regulates the response to oxidative stress. OxyR binds as a tetramer to promoter regions of target genes and acts as an activator or a repressor depending on whether it is oxidized or reduced. In many bacteria, OxyR regulates the expression of genes involved in peroxide detoxification pathways, protein repair, and other oxidative defense-related genes. OxyR in S. meliloti was initially shown to contribute to H2O2 adaptation and both represses and activates the expression of katA depending on its oxidation state [109]. The OxyR regulon has been determined in a whole-genome transcriptional analysis of WT strain and oxyR mutant exposed to H2O2 [101]. It contains over 100 genes, of which some encoding antioxidant functions such as the already known KatA, an AhpC-type alkyl hydroperoxidase, and two chloroperoxidases. OxyR is overexpressed during symbiosis in the nitrogen-fixing zone [109]. An oxyR mutant in A. caulinodans also has an increased sensitivity to H2O2 [69]. The regulator positively controls the expression of the katG gene encoding a bifunctional catalase-hydroperoxidase, involved in detoxification of exogenous H2O2. Both OxyR and KatG are critical for nodulation and nitrogen fixation with S. rostrata [69]. In other rhizobia, oxyR is similarly localized in front of the katG gene and most likely controls its H2O2-dependent expression [110].
The LsrB TR of S. meliloti is also a LysR-like regulator. An lsrB mutant was originally selected for its symbiotic defect during the interaction with M. sativa, and induced ineffective, early senescing nodules with abnormal bacteroid differentiation [111]. Thereafter, LsrB inactivation was found to increase strain sensitivity to oxidative stress. This TF positively regulates the expression of genes involved in GSH homeostasis (gshA, gshB, gor gene encoding GR) and the biosynthesis of lipopolysaccharide (lrp3-lpsCDE operon), which promote both IT formation and bacteroid survival in mature nodules [112]. LsrB was shown to bind to the promoters of gshA and lrp3, and its activity to be modulated by the redox state of three reactive cysteines [113].
In many bacteria, the regulators of the MarR family control the expression of genes involved in the resistance against various environmental and cellular toxic compounds such as antibiotics, detergents, or ROS [114]. Members of the MarR/OhrR subfamily act as dimeric repressors, which are inactivated by thiol-oxidation and dissociate from the promoter DNA of target genes. They respond to organic hydroperoxides and control the expression of peroxidase-encoding genes, often located in the immediate vicinity of the OhrR regulator. In S. meliloti and A. caulinodans, OhrR was shown to regulate ohr by direct binding to its promoter region and OhrR oxidation to prevent this binding and promote ohr expression [70,100]. During S. meliloti/M.sativa interaction, ohr transcripts were detected in nodules and ohr inactivation affects the efficiency of A. caulinodans/ S. rostrata symbiosis, suggesting that the sensing of organic peroxides by OhrR plays a role in the adaptation to the host environment [70,115].
In addition to redox-sensing TR, alternative sigma factors of the RNA polymerase may also contribute to the oxidative stress response. Alternative sigma factors enable bacteria to drive the transcription of genes of shared function in response to environmental stimuli. They are particularly abundant in bacteria experiencing a variety of lifestyles, such as rhizobia and endophytic bacteria. For example, the S. meliloti genome encodes 14 alternative sigma factors, among which RpoHI is required for nitrogen fixation [107,116]. RpoHI controls gene expression in response to pH stress, heat shock, and stationary phase. The RpoHI regulon was defined by transcriptomic studies in free-living bacteria exposed to heat shock or low pH; it includes various genes of the antioxidant defense (gshA, gshB, gor, grx1, trxA, trxB, msrA1, msrA2, msrA3, msrB1, msrB2) [117,118]. Moreover, RpoHI was recently shown to play a role in maintaining the cellular redox status [101]. In R. etli, RpoH1 is mainly involved in heat-shock and oxidative responses. An rpoHI mutant has an increased sensitivity to various oxidants in free-living conditions and forms early senescing nodules with bean [119]. Some other sigma factors that belong to the extracytoplasmic function (ECF) family were also involved in response to various stress conditions, including oxidative stress [53,58,120]. In particular, RpoE4 in R. etli, which contributes to tolerance against osmotic and oxidative stresses, controls the expression of the catalase gene katG; and RpoE2, a major global regulator of the general stress response in S. meliloti, controls the expression of the catalase gene katC [121,122,123].
Several data also concern the transcriptional response to oxidative stress in the endophytic bacterium A. brasilense. Among the ten alternative RpoE sigma factors that A. brasilense possesses, RpoE1 and RpoE2 are activated by photooxidative or oxidative stress through dissociation from their cognate anti-sigma [124]. This leads to indirect induction of the carotenoid biosynthetic pathway via a subset of alternative RpoH sigma factors [58,120,125]. In addition, the regulation of a few genes involved in ROS scavenging has been analyzed. The ahpC gene is located on the chromosome, adjacent to an oxyR-like gene (oxyR1), and its promoter region displays a putative RpoE2-dependent motif [55,124]. The expression of ahpC is negatively regulated by OxyR1 under reducing conditions and is positively regulated by RpoE2 [126]. Another oxyR copy (oxyR2), located adjacent to the katAII gene and in opposite orientation, is necessary for the H2O2 induction of katAII gene expression [58].
6. ROS Transcriptional and Post-Translational Control of Nodule Metabolism during Nitrogen Fixation
BNF is a highly energy-demanding process whereby 16 ATP are hydrolyzed per mol of reduced N2 [127]. However, the Mo-Fe-S complex nitrogenase enzyme is irreversibly inactivated by O2. The high aerobic respiratory turnover, in particular, the one maintained by the plant carbohydrates in bacteroids to sustain nitrogenase activity, as well as respiratory protection used by free-living diazotroph to decrease the internal oxygen concentration, are assumed to generate ROS as a side product. Accordingly, the nitrogenase is inactivated by a subtoxic increase of ROS level, as observed for the sugarcane isolate Gluconobacter diazotrophicus [61]. In addition to a central regulation exerted by O2 level, the nitrogen-fixing bacteria have evolved ROS-dependent regulation of nitrogen fixation metabolism. In endophytic diazotrophs, the genes involved in the synthesis and function of nitrogenase (nif genes) are coordinately regulated via the NifLA proteins [128]. NifL is a flavoprotein that modulates the activity of the TR NifA by direct protein-protein binding in response to changes in O2 and N2 concentrations, the ATP/ADP ratio, and the redox status of the cells [129,130]. In K. pneumoniae, the reduced quinone pool generated by the respiratory chain has been proposed to reduce the FAD domain of NifL under anaerobic conditions, leading to the release of NifA and subsequent induction of nif genes [131]. In diazotrophic bacteria that lack NifL, like rhizobia or free-living bacteria such as Herbaspirillum, the O2 regulation of NifA activity involves an invariant motif Cys-X4-Cys that have been proposed to be redox-sensitive [60,132]. In rhizobia, the FixLJ-FixK cascade positively regulates the nif genes together with genes involved in adaptation to microoxic conditions. The FixL protein contains a heme moiety, which can bind oxygen, thus inactivating the protein [128]. A correlation between the negative redox potential shift provoked by O2 binding to heme and FixL inactivation has been emphasized in R. etli [133]. In B. japonicum, the TR FixK2 amplifies the response under a moderate decrease of oxygen level to approximately 200 target genes. Its activity depends on a redox control, involving a critical cysteine residue near the DNA-binding motif whose oxidation results in FixK2 inactivation [134]. In parallel, a second hierarchical cascade RegSR-nifA, controls the expression of nif genes irrespectively of the oxygen conditions [135]. RegSR is a redox-sensitive two-component regulator system, where the sensor component RegS contains a conserved quinone binding site and a conserved redox-active cysteine, suggesting that the membrane-localized quinone pool and/or the active cysteine mediate the redox signal [134,136].
The post-translational regulation of protein activity has been highlighted for various proteins of nitrogen-fixing bacteroids. Among the 20 sulfenylated proteins identified in S. meliloti bacteroids, the NifK and NifH components of the nitrogenase were susceptible to sulfenylation [137]. In addition, a large proportion of the sulfenylated protein is involved in the TCA cycle allowing a coordinated regulation between the cellular redox state, the nitrogenase activity, and the enzymes of cellular energy metabolism. A fine control of energy demand and reducing power produced via the TCA cycle might involve polyhydroxybutyrate (PHB), a polyhydroxyalkanoate polymer that acts as carbon storage compounds, and regulates the cellular NAD(P)H pool, thereby being involved in cellular redox balance [138]. The PHB synthesis depends on a three-step pathway involving successively the β-ketothiolase PhbA, the acetoacetylcoA reductase PhbB, and the PHB synthase PhbC, while PhaZ initiates the depolymerization of PHB. Genes involved in PHB synthesis have been characterized in different endophytic bacteria and rhizobia, such as A. brasilense, H. seropedicae, S. meliloti, R. etli, R. leguminosarum, B. japonicum, or A. caulinodans [82,83,84,139,140,141,142]. The level of ROS was approximately 50% higher in a phbC mutant of H. seropedicae. This feature leads to the inactivation of the FixK homolog, FnR, a [Fe-S] containing protein that should be maintained in its reduced form to induced genes involved in the adaptation to microaerophilia [140]. The H. seropedicae phbC mutant fails to promote plant growth, although it conserves the ability to colonize plant roots as the wild type [63]. The PHB synthase is also not essential for the colonization of A. brasilense [143]. In A. caulinodans, a phbC mutant was deprived of nitrogenase activity, depleted in ATP and displayed an increase in reduced NADH [83]. Moreover, the nifA expression, controlled by the FixLJK cascade, was totally abolished, and nodules induced by the PHB mutant were devoid of bacteroids (Table 1). Similarly, the S. meliloti phbC mutation results in the formation of numerous nodules that are unable to fix nitrogen [82]. However, even if the PHB accumulation in bacteroids occurs in soybean and common bean nodules, and not in pea, alfalfa, and clover, [144], the phbC mutation of B. japonicum, R. leguminosarum, and R. etli does not impair nitrogen fixation [84,141]. Terpolilli and colleagues proposed that bacteroids used multiple lipid sinks, as fatty acid or glycolipids, in addition to PHB, for excess reductants and regulation of the redox balance in fixing symbiosis [145]. Thus, the importance of the PHB cycle depends on the physiology of the nodule.
7. Conclusions
During the last years, the characterization of many redox regulatory systems has shown the importance of redox regulation in the plant-bacteria symbiotic recognition and the functioning of the symbiotic interactions. The specific redox regulation in the symbiotic interaction may have a crucial role in differentiating pathogens from symbiotic microorganisms and to allowing the proper functioning of the interaction. The development of omics data analyses gives us a more general view of the redox network regulation and the expression of different members depending on the physiological state of the organisms. However, these advances are revealing the complexity of the regulatory mechanisms and an increased number of key regulatory actors depending on the stage of the analysis and the biological material. Many lines of research remain to be opened. One of them is to compare the different redox regulation occurring in the different symbiotic interactions to identify similar and different processes. These analyses may allow defining a regulatory hub used by partners in different types of symbiotic relations. Another crucial question is how the organisms regulate their responses in front of multiple stimuli, including biotic and abiotic environments and the metabolic processes. Finally, the screening of potential candidates involved in improving the plant-microbe symbiotic interactions to increase the efficiency of plant development and defense is of high interest to develop more sustainable agriculture.
Funding
This work was supported by “l’Institut National de Recherche pour l’Agriculture, l’Alimentation et l’Environnement”, the “Centre National de la Recherche Scientifique”, the University of Nice–Sophia Antipolis, the French Government (National Research Agency, ANR) through the “Investments for the Future” LABEX SIGNALIFE: program reference # ANR-11-LABX-0028-01 and the Hubert Curien Gundishapur program (program reference 42938YA).
Acknowledgments
We thank the Microscopy and the Analytical Biochemistry Platforms—Sophia Agrobiotech Institut—INRA 1355—UNS—CNRS 7254—INRA PACA-Sophia Antipolis for access to instruments. All figures were created with Biorender.com (assessed on 7 May 2021).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 420. [Google Scholar] [CrossRef]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef] [Green Version]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Welte, C.U. Revival of Archaeal Methane Microbiology. mSystems 2018, 3, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.L.G.; Balsemão-Pires, E.; Saraiva, R.M.; Ferreira, P.C.G.; Hemerly, A.S. Nitrogen signalling in plant interactions with associative and endophytic diazotrophic bacteria. J. Exp. Bot. 2014, 65, 5631–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, U.; Johansson, C.; Bergman, B. Early communication in the Gunnera-Nostoc symbiosis: Plant-induced cell differentiation and protein synthesis in the cyanobacterium. Mol. Plant Microbe Interact. 1994, 7, 696–702. [Google Scholar] [CrossRef]
- Pawlowski, K.; Demchenko, K.N. The diversity of actinorhizal symbiosis. Protoplasma 2012, 249, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [Green Version]
- Chelius, M.K.; Triplett, E.W. Immunolocalization of dinitrogenase reductase produced by Kiebsiella pneumoniae in association with Zea mays L. Appl. Environ. Microbiol. 2000, 66, 783–787. [Google Scholar] [CrossRef] [Green Version]
- Frederickson Matika, D.E.; Loake, G.J. Redox Regulation in Plant Immune Function. Antioxid. Redox Signal. 2014, 21, 1373–1388. [Google Scholar] [CrossRef]
- Syska, C.; Brouquisse, R.; Alloing, G.; Pauly, N.; Frendo, P.; Bosseno, M.; Dupont, L.; Boscari, A. Molecular Weapons Contribute to Intracellular Rhizobia Accommodation Within Legume Host Cell. Front. Plant Sci. 2019, 10, 1496. [Google Scholar] [CrossRef]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of Host Immunity by Beneficial Microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelly, M.J.; Loake, G.J. Synthesis of redox-active molecules and their signaling functions during the expression of plant disease resistance. Antioxid. Redox Signal. 2013, 19, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; Pedrosa, F.O.; Müller-Santos, M.; de Souza, E.M. What Did We Learn from Plant Growth-Promoting Rhizobacteria (PGPR)-Grass Associations Studies through Proteomic and Metabolomic Approaches? Front. Sustain. Food Syst. 2020, 4, e607343. [Google Scholar] [CrossRef]
- Malviya, M.K.; Li, C.-N.; Solanki, M.K.; Singh, R.K.; Htun, R.; Singh, P.; Verma, K.K.; Yang, L.-T.; Li, Y.-R. Comparative analysis of sugarcane root transcriptome in response to the plant growth-promoting Burkholderia anthina MYSP113. PLoS ONE 2020, 15, e0231206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaepen, S.; Bossuyt, S.; Engelen, K.; Marchal, K.; Vanderleyden, J. Phenotypical and molecular responses of Arabidopsis thaliana roots as a result of inoculation with the auxin-producing bacterium Azospirillum brasilense. New Phytol. 2014, 201, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Lade, S.B.; Román, C.; Cueto-Ginzo, A.I.; Serrano, L.; Sin, E.; Achón, M.A.; Medina, V. Host-specific proteomic and growth analysis of maize and tomato seedlings inoculated with Azospirillum brasilense Sp7. Plant Physiol. Biochem. 2018, 129, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Park, J.Y.; Lee, S.H.; Hwang, C.H. Comparative proteomic analysis of Soybean nodulation using a supernodulation mutant, SS2-2. Biosci. Biotechnol. Biochem. 2010, 74, 2396–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 2017, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Gómez, M.; Castro-Mercado, E.; Alexandre, G.; García-Pineda, E. Oxidative and antioxidative responses in the wheat-Azospirillum brasilense interaction. Protoplasma 2016, 253, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Damiani, I.; Puppo, A.; Frendo, P. Redox Changes during the Legume–Rhizobium Symbiosis. Mol. Plant 2009, 2, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.K.; Andrio, E.; Marino, D.; Pauly, N.; Dunand, C. Reactive Oxygen Species during Plant-microorganism Early Interactions. J. Integr. Plant Biol. 2010, 52, 195–204. [Google Scholar] [CrossRef]
- Cárdenas, L.; Martínez, A.; Sánchez, F.; Quinto, C. Fast, transient and specific intracellular ROS changes in living root hair cells responding to Nod factors (NFs). Plant J. 2008, 56, 802–813. [Google Scholar] [CrossRef]
- Fournier, J.; Timmers, A.C.J.; Sieberer, B.J.; Jauneau, A.; Chabaud, M.; Barker, D.G. Mechanism of infection thread elongation in root hairs of Medicago truncatula and dynamic interplay with associated rhizobial colonization. Plant Physiol. 2008, 148, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Peleg-Grossman, S.; Volpin, H.; Levine, A. Root hair curling and Rhizobium infection in Medicago truncatula are mediated by phosphatidylinositide-regulated endocytosis and reactive oxygen species. J. Exp. Bot. 2007, 58, 1637–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthikala, M.K.; Montiel, J.; Sánchez-López, R.; Nava, N.; Cárdenas, L.; Quinto, C. Respiratory burst oxidase homolog gene a is crucial for Rhizobium infection and nodule maturation and function in common bean. Front. Plant Sci. 2017, 8, 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamet, A.; Mandon, K.; Puppo, A.; Hérouart, D.; Hérouart, D. H2O2 is required for optimal establishment of the Medicago sativa/Sinorhizobium meliloti symbiosis. J. Bacteriol. 2007, 189, 8741–8745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gucciardo, S.; Rathbun, E.A.; Shanks, M.; Jenkyns, S.; Mak, L.; Durrant, M.C.; Brewin, N.J. Epitope Tagging of Legume Root Nodule Extensin Modifies Protein Structure and Crosslinking in Cell Walls of Transformed Tobacco Leaves. Mol. Plant Microbe Interact. 2005, 18, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Bhattacharyya, S.; Kumar, R.; Kumar, A.; Ibañez, F.; Wang, J.; Guo, B.; Sudini, H.K.; Gopalakrishnan, S.; DasGupta, M.; et al. Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- D’Haeze, W.; de Rycke, R.; Mathis, R.; Goormachtig, S.; Pagnotta, S.; Verplancke, C.; Capoen, W.; Holsters, M. Reactive oxygen species and ethylene play a positive role in lateral root base nodulation of a semiaquatic legume. Proc. Natl. Acad. Sci. USA 2003, 100, 11789–11794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrio, E.; Marino, D.; Marmeys, A.; de Segonzac, M.D.; Damiani, I.; Genre, A.; Huguet, S.; Frendo, P.; Puppo, A.; Pauly, N. Hydrogen peroxide-regulated genes in the Medicago truncatula-Sinorhizobium meliloti symbiosis. New Phytol. 2013, 198, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Mergaert, P. Terminal bacteroid differentiation in the Medicago–Rhizobium interaction—A tug of war between plant and bacteria. In The Model Legume Medicago Truncatula; de Bruijn, F., Ed.; Wiley: New York, NY, USA, 2020; pp. 600–616. [Google Scholar]
- Haag, A.F.; Kerscher, B.; Dall’Angelo, S.; Sani, M.; Longhi, R.; Baloban, M.; Wilson, H.M.; Mergaert, P.; Zanda, M.; Ferguson, G.P. Role of cysteine residues and disulfide bonds in the activity of a legume root nodule-specific, cysteine-rich peptide. J. Biol. Chem. 2012, 287, 10791–10798. [Google Scholar] [CrossRef] [Green Version]
- Shabab, M.; Arnold, M.F.F.; Penterman, J.; Wommack, A.J.; Bocker, H.T.; Price, P.A.; Griffitts, J.S.; Nolan, E.M.; Walker, G.C. Disulfide cross-linking influences symbiotic activities of nodule peptide NCR247. Proc. Natl. Acad. Sci. USA 2016, 113, 10157–10162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, C.W.; Baldacci-Cresp, F.; Pierre, O.; Larousse, M.; Benyamina, S.; Lambert, A.; Hopkins, J.; Castella, C.; Cazareth, J.; Alloing, G.; et al. Regulation of Differentiation of Nitrogen-Fixing Bacteria by Microsymbiont Targeting of Plant Thioredoxin s1. Curr. Biol. 2017, 27, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Becana, M.; Ibaliz Larrainzar, E.; Villar, I.; Rubio, M.C.; Erez-Rontom, E.C.P.; Huertas, R.; Sato, S.; Mun, J.-H. Research review Hemoglobins in the legume-Rhizobium symbiosis. New Phytol. 2020, 228, 472–484. [Google Scholar] [CrossRef]
- Marino, D.; Andrio, E.; Danchin, E.G.J.; Oger, E.; Gucciardo, S.; Lambert, A.; Puppo, A.; Pauly, N. A Medicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. New Phytol. 2011, 189, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, Y.; Feng, X.; Luo, Z.; Kong, S.; Zhang, C.; Gong, A.; Yuan, H.; Cheng, L.; Wang, X. Genomic, molecular evolution, and expression analysis of NOX genes in soybean (Glycine max). Genomics 2019, 111, 619–628. [Google Scholar] [CrossRef]
- Frendo, P.; Matamoros, M.A.; Alloing, G.; Becana, M. Thiol-based redox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 2013, 4, e376. [Google Scholar] [CrossRef] [Green Version]
- Frendo, P.; Harrison, J.; Norman, C.; Jiménez, M.J.H.; van de Sype, G.; Gilabert, A.; Puppo, A. Glutathione and homoglutathione play a critical role in the nodulation process of Medicago truncatula. Mol. Plant Microbe Interact. 2005, 18, 254–259. [Google Scholar] [CrossRef] [Green Version]
- El Msehli, S.; Lambert, A.; Baldacci-Cresp, F.; Hopkins, J.; Boncompagni, E.; Smiti, S.A.; Hérouart, D.; Frendo, P. Crucial role of (homo)glutathione in nitrogen fixation in Medicago truncatula nodules. New Phytol. 2011, 192, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Guan, N.; Zhou, Y.; Mei, L.; Li, Y.; Li, Y. A Methionine Sulfoxide Reductase B Is Required for the Establishment of Astragalus sinicus-Mesorhizobium Symbiosis. Plant Cell Physiol. 2020, 61, 1631–1645. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; Abee, T. Primary and secondary oxidative stress in Bacillus. Environ. Microbiol. 2011, 13, 1387–1394. [Google Scholar] [CrossRef]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [Green Version]
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Mishra, S.; Imlay, J. Why do bacteria use so many enzymes to scavenge hydrogen peroxide? Arch. Biochem. Biophys. 2012, 525, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Deponte, M. The Incomplete Glutathione Puzzle: Just Guessing at Numbers and Figures? Antioxid. Redox Signal. 2017, 27, 1130–1161. [Google Scholar] [CrossRef] [Green Version]
- Van Loi, V.; Rossius, M.; Antelmann, H. Redox regulation by reversible protein S-thiolation in bacteria. Front. Microbiol. 2015, 6, e187. [Google Scholar] [CrossRef] [Green Version]
- Tadra-Sfeir, M.Z.; Souza, E.M.; Faoro, H.; Müller-Santos, M.; Baura, V.A.; Tuleski, T.R.; Rigo, L.U.; Yates, M.G.; Wassem, R.; Pedrosa, F.O.; et al. Naringenin regulates expression of genes involved in cell wall synthesis in Herbaspirillum seropedicae. Appl. Environ. Microbiol. 2011, 77, 2180–2183. [Google Scholar] [CrossRef] [Green Version]
- Drogue, B.; Sanguin, H.; Borland, S.; Prigent-Combaret, C.; Wisniewski-Dyé, F. Genome wide profiling of Azospirillum lipoferum 4B gene expression during interaction with rice roots. FEMS Microbiol. Ecol. 2014, 87, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Lozano, L.; Acosta-Cruz, E.; Borland, S.; Drogue, B.; Prigent-Combaret, C.; Rouy, Z.; Barbe, V.; Herrera, A.M.; González, V.; et al. Genome sequence of Azospirillum brasilense CBG497 and comparative analyses of Azospirillum core and accessory genomes provide insight into niche adaptation. Genes 2012, 3, 576–602. [Google Scholar] [CrossRef] [PubMed]
- Camilios-Neto, D.; Bonato, P.; Wassem, R.; Tadra-Sfeir, M.Z.; Brusamarello-Santos, L.C.; Valdameri, G.; Donatti, L.; Faoro, H.; Weiss, V.A.; Chubatsu, L.S.; et al. Dual RNA-seq transcriptional analysis of wheat roots colonized by Azospirillum brasilense reveals up-regulation of nutrient acquisition and cell cycle genes. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasim, M.; Bible, A.N.; Xie, Z.; Alexandre, G. Alkyl hydroperoxide reductase has a role in oxidative stress resistance and in modulating changes in cell-surface properties in Azospirillum brasilense Sp245. Microbiology 2009, 155, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Singh, S.; Dwivedi, S.K.; Srivastava, A.; Pandey, P.; Kumar, S.; Singh, B.N.; Tripathi, A.K. Catalase Expression in Azospirillum brasilense Sp7 Is Regulated by a Network Consisting of OxyR and Two RpoH Paralogs and Including an RpoE1→RpoH5 Regulatory Cascade. Appl. Environ. Microbiol. 2018, 84, e01787-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battu, L.; Ulaganathan, K. Whole genome sequencing and identification of host-interactive genes in the rice endophytic Leifsonia sp. ku-ls. Funct. Integr. Genom. 2020, 20, 237–243. [Google Scholar] [CrossRef]
- Chaves, D.F.S.; de Souza, E.M.; Monteiro, R.A.; de Oliveira Pedrosa, F. A two-dimensional electrophoretic profile of the proteins secreted by Herbaspirillum seropedicae strain Z78. J. Proteom. 2009, 73, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Alquéres, S.M.C.; Oliveira, J.H.M.; Nogueira, E.M.; Guedes, H.V.; Oliveira, P.L.; Câmara, F.; Baldani, J.I.; Martins, O.B. Antioxidant pathways are up-regulated during biological nitrogen fixation to prevent ROS-induced nitrogenase inhibition in Gluconacetobacter diazotrophicus. Arch. Microbiol. 2010, 192, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alquéres, S.; Meneses, C.; Rouws, L.; Rothballer, M.; Baldani, I.; Schmid, M.; Hartmann, A. The Bacterial Superoxide Dismutase and Glutathione Reductase Are Crucial for Endophytic Colonization of Rice Roots by Gluconacetobacter diazotrophicus PAL5. Mol. Plant Microbe Interact. 2013, 26, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, L.P.S.; do Amaral, F.P.; Kim, D.; Bom, M.T.; Gavídia, M.P.; Teixeira, C.S.; Holthman, F.; de Oliveira Pedrosa, F.; de Souza, E.M.; Chubatsu, L.S.; et al. Importance of poly-3-hydroxybutyrate metabolism to the ability of Herbaspirillum seropedicae to promote plant growth. Appl. Environ. Microbiol. 2019, 85, e02586-18. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.; Herouart, D.; Puppo, A.; Touati, D. Critical protective role of bacterial superoxide dismutase in Rhizobium-legume symbiosis. Mol. Microbiol. 2000, 38, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Jamet, A.; Sigaud, S.; van de Sype, G.; Puppo, A.; Hérouart, D. Expression of the bacterial catalase genes during Sinorhizobium meliloti-Medicago sativa symbiosis and their crucial role during the infection process. Mol. Plant Microbe Interact. 2003, 16, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Hanyu, M.; Fujimoto, H.; Tejima, K.; Saeki, K. Functional differences of two distinct catalases in Mesorhizobium loti MAFF303099 under free-living and symbiotic conditions. J. Bacteriol. 2009, 191, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Dombrecht, B.; Heusdens, C.; Beullens, S.; Verreth, C.; Mulkers, E.; Proost, P.; Vanderleyden, J.; Michiels, J. Defence of Rhizobium etli bacteroids against oxidative stress involves a complexly regulated atypical 2-Cys peroxiredoxin. Mol. Microbiol. 2005, 55, 1207–1221. [Google Scholar] [CrossRef]
- Wang, S.; Lu, T.; Xue, Q.; Xu, K.; Cheng, G. Antioxidation and symbiotic nitrogen fixation function of prxA gene in Mesorhizobium huakuii. Microbiologyopen 2019, 8, e889. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Nickels, L.M.; Wang, H.; Ling, J.; Zhong, Z.; Zhu, J. OxyR-regulated catalase activity is critical for oxidative stress resistance, nodulation and nitrogen fixation in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2016, 363, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.; Yang, J.; Li, X.; Cao, Y.; Liu, X.X.; Ling, J.; Wang, H.; Zhong, Z.; Zhu, J. Alkyl hydroperoxide reductase is important for oxidative stress resistance and symbiosis in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2019, 366, 1–9. [Google Scholar] [CrossRef]
- Si, Y.; Guo, D.; Deng, S.; Lu, X.; Zhu, J.; Rao, B.; Cao, Y.; Jiang, G.; Yu, D.; Zhong, Z.; et al. Ohr and ohrR are critical for organic peroxide resistance and symbiosis in Azorhizobium caulinodans ORS571. Genes 2020, 11, 335. [Google Scholar] [CrossRef] [Green Version]
- Wheatley, R.M.; Ford, B.L.; Li, L.; Aroney, S.T.N.; Knights, H.E.; Ledermann, R.; East, A.K.; Ramachandran, V.K.; Poole, P.S. Lifestyle adaptations of Rhizobium from rhizosphere to symbiosis. Proc. Natl. Acad. Sci. USA 2020, 117, 23823–23834. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Karunakaran, R.; East, A.K.; Munoz-Azcarate, O.; Poole, P.S. Glutathione affects the transport activity of Rhizobium leguminosarum 3841 and is essential for efficient nodulation. FEMS Microbiol. Lett. 2017, 364, 1–7. [Google Scholar] [CrossRef]
- Harrison, J.; Jamet, A.; Muglia, C.I.; van de Sype, G.; Aguilar, O.M.; Puppo, A.; Frendo, P. Glutathione plays a fundamental role in growth and symbiotic capacity of Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; El Msehli, S.; Benyamina, S.; Lambert, A.; Hopkins, J.; Cazareth, J.; Pierre, O.; Hérouart, D.; Achi-Smiti, S.; Boncompagni, E.; et al. Glutathione Deficiency in Sinorhizobium meliloti Does Not Impair Bacteroid Differentiation But Induces Early Senescence in the Interaction With Medicago truncatula. Front. Plant Sci. 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Taté, R.; Cermola, M.; Riccio, A.; Diez-Roux, G.; Patriarca, E.J. Glutathione Is Required by Rhizobium etli for Glutamine Utilization and Symbiotic Effectiveness. Mol. Plant Microbe Interact. 2012, 25, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Muglia, C.; Comai, G.; Spegazzini, E.; Riccillo, P.M.; Aguilar, O.M. Glutathione produced by Rhizobium tropici is important to prevent early senescence in common bean nodules. FEMS Microbiol. Lett. 2008, 286, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Yin, J.; Peng, Y.; Xie, J.; Wu, H.; He, D.; Li, X.; Cheng, G. Glutathione is Involved in Detoxification of Peroxide and Root Nodule Symbiosis of Mesorhizobium huakuii. Curr. Microbiol. 2020, 77, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Li, N.; Liu, Y.; Yu, L.; Yan, J.; Luo, L. Sinorhizobium meliloti Glutathione Reductase Is Required for both Redox Homeostasis and Symbiosis. Appl. Environ. Microbiol. 2017, 84, e01937-17. [Google Scholar] [CrossRef] [Green Version]
- Benyamina, S.M.; Baldacci-Cresp, F.; Couturier, J.; Chibani, K.; Hopkins, J.; Bekki, A.; de Lajudie, P.; Rouhier, N.; Jacquot, J.P.; Alloing, G.; et al. Two Sinorhizobium meliloti glutaredoxins regulate iron metabolism and symbiotic bacteroid differentiation. Environ. Microbiol. 2013, 15, 795–810. [Google Scholar] [CrossRef]
- Cao, Y.; Jiang, G.; Li, M.; Fang, X.; Zhu, D.; Qiu, W.; Zhu, J.; Yu, D.; Xu, Y.; Zhong, Z.; et al. Glutaredoxins Play an Important Role in the Redox Homeostasis and Symbiotic Capacity of Azorhizobium caulinodans ORS571. Mol. Plant Microbe Interact. 2020, 33, 1381–1393. [Google Scholar] [CrossRef]
- Wang, C.; Saldanha, M.; Sheng, X.; Shelswell, K.J.; Walsh, K.T.; Sobral, B.W.S.; Charles, T.C. Roles of poly-3-hydroxybutyrate (PHB) and glycogen in symbiosis of Sinorhizobium meliloti with Medicago sp. Microbiology 2007, 153, 388–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandon, K.; Michel-Reydellet, N.; Encarnación, S.; Kaminski, P.A.; Leija, A.; Cevallos, M.A.; Elmerich, C.; Mora, J. Poly-β-hydroxybutyrate turnover in Azorhizobium caulinodans is required for growth and affects nifA expression. J. Bacteriol. 1998, 180, 5070–5076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quelas, J.I.; Mongiardini, E.J.; Perez-Gimenez, J.; Parisi, G.; Lodeiro, A.R. Analysis of Two Polyhydroxyalkanoate Synthases in Bradyrhizobium japonicum USDA 110. J. Bacteriol. 2013, 195, 3145–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, V.K.; East, A.K.; Karunakaran, R.; Downie, J.A.; Poole, P.S. Adaptation of Rhizobium leguminosarum to pea, alfalfa and sugar beet rhizospheres investigated by comparative transcriptomics. Genome Biol. 2011, 12, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klonowska, A.; Melkonian, R.; Miché, L.; Tisseyre, P.; Moulin, L. Transcriptomic profiling of Burkholderia phymatum STM815, Cupriavidus taiwanensis LMG19424 and Rhizobium mesoamericanum STM3625 in response to Mimosa pudica root exudates illuminates the molecular basis of their nodulation competitiveness and symbiotic evolutionary history. BMC Genom. 2018, 19, 1–22. [Google Scholar] [CrossRef]
- Salas, M.E.; Lozano, M.J.; López, J.L.; Draghi, W.O.; Serrania, J.; Torres Tejerizo, G.A.; Albicoro, F.J.; Nilsson, J.F.; Pistorio, M.; del Papa, M.F.; et al. Specificity traits consistent with legume-rhizobia coevolution displayed by Ensifer meliloti rhizosphere colonization. Environ. Microbiol. 2017, 19, 3423–3438. [Google Scholar] [CrossRef] [Green Version]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen Peroxide and Nitric Oxide: Key Regulators of the Legume—Rhizobium and Mycorrhizal Symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Alloing, G.; Mandon, K.; Frendo, P. Redox regulation of differentiation in symbiotic nitrogen fixation. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1469–1478. [Google Scholar] [CrossRef]
- Montiel, J.; Nava, N.; Cárdenas, L.; Sánchez-López, R.; Arthikala, M.K.; Santana, O.; Sánchez, F.; Quinto, C. A Phaseolus vulgaris NADPH Oxidase Gene is Required for Root Infection by Rhizobia. Plant Cell Physiol. 2012, 53, 1751–1767. [Google Scholar] [CrossRef] [Green Version]
- Tani, A.; Takai, Y.; Suzukawa, I.; Akita, M.; Murase, H.; Kimbara, K. Practical application of methanol-mediated mutualistic symbiosis between methylobacterium species and a roof greening moss, Racomitrium japonicum. PLoS ONE 2012, 7, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Damiani, I.; Pauly, N.; Puppo, A.; Brouquisse, R.; Boscari, A. Reactive oxygen species and nitric oxide control early steps of the legume—Rhizobium symbiotic interaction. Front. Plant Sci. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Muñoz, V.; Ibáñez, F.; Figueredo, M.S.; Fabra, A. An oxidative burst and its attenuation by bacterial peroxidase activity is required for optimal establishment of the Arachis hypogaea-Bradyrhizobium sp. symbiosis. J. Appl. Microbiol. 2016, 121, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Barloy-Hubler, F.; Chéron, A.; Hellégouarch, A.; Galibert, F. Smc01944, a secreted peroxidase induced by oxidative stresses in Sinorhizobium meliloti 1021. Microbiology 2004, 150, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroux, C.; Washburn, B.K.; Davis, O.M.; Stewart, J.; Brewer, T.E.; Lyons, M.R.; Jones, K.M. A comparative genomics screen identifies a Sinorhizobium meliloti 1021 sodM-like gene strongly expressed within host plant nodules. BMC Microbiol. 2012, 12, e74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krehenbrink, M.; Edwards, A.; Downie, J.A. The superoxide dismutase SodA is targeted to the periplasm in a SecA-dependent manner by a novel mechanism. Mol. Microbiol. 2011, 82, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Asensio, A.C.; Marino, D.; James, E.K.; Ariz, I.; Arrese-Igor, C.; Aparicio-Tejo, P.M.; Arredondo-Peter, R.; Moran, J.F. Expression and localization of a Rhizobium-derived cambialistic superoxide dismutase in pea (Pisum sativum) nodules subjected to oxidative stress. Mol. Plant Microbe Interact. 2011, 24, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.M.; Lee, H.I.; Donati, A.J.; So, J.S.; Emerich, D.W.; Chang, W.S. Whole-genome expression profiling of Bradyrhizobium japonicum in response to hydrogen peroxide. Mol. Plant Microbe Interact. 2011, 24, 1472–1481. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, M.A.; Chen, H.C.; Natera, S.; van Noorden, G.; Menzel, C.; Taylor, S.; Renard, C.; Geiger, O.; Weiller, G.F. A Global Analysis of Protein Expression Profiles in Sinorhizobium meliloti: Discovery of New Genes for Nodule Occupancy and Stress Adaptation. Mol. Plant Microbe Interact. 2003, 16, 508–524. [Google Scholar] [CrossRef] [Green Version]
- Fontenelle, C.; Blanco, C.; Arrieta, M.; Dufour, V.; Trautwetter, A. Resistance to organic hydroperoxides requires ohr and ohrR genes in Sinorhizobium meliloti. BMC Microbiol. 2011, 11, e100. [Google Scholar] [CrossRef] [Green Version]
- Lehman, A.P.; Long, R. OxyR-Dependent Transcription Response of Sinorhizobium meliloti to Oxidative Stress. J. Bacteriol. 2018, 200, e00622-17. [Google Scholar] [CrossRef] [Green Version]
- Panek, H.R.; O’Brian, M.R. KatG is the primary detoxifier of hydrogen peroxide produced by aerobic metabolism in Bradyrhizobium japonicum. J. Bacteriol. 2004, 186, 7874–7880. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, N.; Ahrens, C.H.; Knief, C.; Qeli, E.; Koch, M.; Fischer, H.-M.; Vorholt, J.A.; Hennecke, H.; Pessi, G. An integrated proteomics and transcriptomics reference data set provides new insights into the Bradyrhizobium japonicum bacteroid metabolism in soybean root nodules. Proteomics 2010, 10, 1391–1400. [Google Scholar] [CrossRef]
- del Carmen Vargas, M.; Encarnación, S.; Dávalos, A.; Reyes-Pérez, A.; Mora, Y.; García-de los Santos, A.; Brom, S.; Mora, J. Only one catalase, katG, is detectable in Rhizobium etli, and is encoded along with the regulator OxyR on a plasmid replicon. Microbiology 2003, 149, 1165–1176. [Google Scholar] [CrossRef] [Green Version]
- Alloisio, N.; Queiroux, C.; Fournier, P.; Pujic, P.; Normand, P.; Vallenet, D.; Médigue, C.; Yamaura, M.; Kakoi, K.; Kucho, K.I. The frankia alni symbiotic transcriptome. Mol. Plant Microbe Interact. 2010, 23, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Minamisawa, K.; Mitsui, H. A Sinorhizobium meliloti RpoH-regulated gene is involved in iron-sulfur protein metabolism and effective plant symbiosis under intrinsic iron limitation. J. Bacteriol. 2016, 198, 2297–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Aono, T.; Lee, K.B.; Suzuki, T.; Liu, C.T.; Miwa, H.; Wakao, S.; Iki, T.; Oyaizu, H. Rhizobial factors required for stem nodule maturation and maintenance in Sesbania rostrata-Azorhizobium caulinodans ORS571 symbiosis. Appl. Environ. Microbiol. 2007, 73, 6650–6659. [Google Scholar] [CrossRef] [Green Version]
- Hillion, M.; Antelmann, H. Thiol-based redox switches in prokaryotes. Biol. Chem. 2015, 396, 415–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamet, A.; Kiss, E.; Batut, J.; Puppo, A.; Hérouart, D. The katA catalase gene is regulated by OxyR in both free-living and symbiotic Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 376–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakjarung, K.; Mongkolsuk, S.; Vattanaviboon, P. The oxyR from Agrobacterium tumefaciens: Evaluation of its role in the regulation of catalase and peroxide responses. Biochem. Biophys. Res. Commun. 2003, 304, 41–47. [Google Scholar] [CrossRef]
- Luo, L.; Yao, S.-Y.; Becker, A.; Rüberg, S.; Yu, G.-Q.; Zhu, J.-B.; Cheng, H.-P. Two New Sinorhizobium meliloti LysR-type Transcriptional Regulators Required for Nodulation. J. Bacteriol. 2005, 187, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Wang, Y.; Luo, L. Transcriptional regulator LsrB of Sinorhizobium meliloti positively regulates the expression of genes involved in lipopolysaccharide biosynthesis. Appl. Environ. Microbiol. 2014, 80, 5265–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Xing, S.; Wang, S.; Yu, L.; Li, X.; Staehelin, C.; Yang, M.; Luo, L. Regulation of cysteine residues in LsrB proteins from Sinorhizobium meliloti under free-living and symbiotic oxidative stress. Environ. Microbiol. 2017, 19, 5130–5145. [Google Scholar] [CrossRef]
- Wilkinson, S.P.; Grove, A. Ligand-responsive transcriptional regulation by members of the MarR family of winged helix proteins. Curr. Issues Mol. Biol. 2006, 8, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, B.; Rodde, N.; Jardinaud, M.F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrère, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Oke, V.; Rushing, B.G.; Fisher, E.J.; Moghadam-Tabrizi, M.; Long, S.R. Identification of the heat-shock sigma factor RpoH and a second RpoH-like protein in Sinorhizobium meliloti. Microbiology 2001, 147, 2399–2408. [Google Scholar] [CrossRef] [Green Version]
- De Lucena, D.K.C.; Pühler, A.; Weidner, S. The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti. BMC Microbiol. 2010, 10, e265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, M.J.; Bittner, A.N.; Toman, C.J.; Oke, V.; Long, S.R. Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium. J. Bacteriol. 2012, 194, 4983–4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Salazar, J.M.; Sandoval-Calderón, M.; Guo, X.; Castillo-Ramírez, S.; Reyes, A.; Loza, M.G.; Rivera, J.; Alvarado-Affantranger, X.; Sánchez, F.; González, V.; et al. The Rhizobium etli RpoH1 and RpoH2 sigma factors are involved in different stress responses. Microbiology 2009, 155, 386–397. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Gupta, A.; Kumar, S.; Mishra, R.; Singh, C.; Tripathi, A.K. Cross-Talk Between Cognate and Noncognate RpoE Sigma Factors and Zn2+-Binding Anti-Sigma Factors Regulates Photooxidative Stress Response in Azospirillum brasilense. Antioxid. Redox Signal. 2014, 20, 42–59. [Google Scholar] [CrossRef]
- Sauviac, L.; Philippe, H.; Phok, K.; Bruand, C. An extracytoplasmic function sigma factor acts as a general stress response regulator in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 4204–4216. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Salazar, J.M.; Salazar, E.; Encarnación, S.; Ramírez-Romero, M.A.; Rivera, J. Role of the extracytoplasmic function sigma factor RpoE4 in oxidative and osmotic stress responses in Rhizobium etli. J. Bacteriol. 2009, 191, 4122–4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flechard, M.; Fontenelle, C.; Trautwetter, A.; Ermel, G.; Blanco, C. Sinorhizobium meliloti rpoE2 is necessary for H2O2 stress resistance during the stationary growth phase. FEMS Microbiol. Lett. 2009, 290, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Kumar, S.; Mishra, M.N.; Tripathi, A.K. A constitutively expressed pair of rpoE2–chrR2 in Azospirillum brasilense Sp7 is required for survival under antibiotic and oxidative stress. Microbiology 2013, 159, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Dubey, A.P.; Kumar, S.; Dutta, D.; Mishra, M.N.; Singh, B.N.; Tripathi, A.K. Carotenoid Biosynthetic Pathways Are Regulated by a Network of Multiple Cascades of Alternative Sigma Factors in Azospirillum brasilense Sp7. J. Bacteriol. 2016, 198, 2955–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Dwivedi, S.K.; Singh, V.S.; Tripathi, A.K. Expression of alkyl hydroperoxide reductase is regulated negatively by OxyR1 and positively by RpoE2 sigma factor in Azospirillum brasilense Sp7. Microbiology 2016, 162, 1870–1883. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Rees, D.C. Nitrogenase and Biological Nitrogen Fixation. Biochemistry 1994, 33, 389–397. [Google Scholar] [CrossRef]
- Rutten, P.J.; Poole, P.S. Oxygen regulatory mechanisms of nitrogen fixation in rhizobia. In Advances in Microbial Physiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 75, pp. 325–389. ISBN 9780128177143. [Google Scholar]
- Hill, S.; Austin, S.; Eydmann, T.; Jones, T.; Dixon, R. Azotobacter vinelandii NIFL is a flavoprotein that modulates transcriptional activation of nitrogen-fixation genes via a redox-sensitive switch. Proc. Natl. Acad. Sci. USA 1996, 93, 2143–2148. [Google Scholar] [CrossRef] [Green Version]
- Das, H.K. Azotobacters as biofertilizer. Adv. Appl. Microbiol. 2019, 108, 1–43. [Google Scholar] [CrossRef]
- Grabbe, R.; Schmitz, R.A. Oxygen control of nif gene expression in Klebsiella pneumoniae depends on NifL reduction at the cytoplasmic membrane by electrons derived from the reduced quinone pool. Eur. J. Biochem. 2003, 270, 1555–1566. [Google Scholar] [CrossRef] [Green Version]
- Fischer, H.M.; Bruderer, T.; Hennecke, H. Essential and non-essential domains in the Bradyrhizobium japonicum NifA protein: Identification of indispensable cysteine residues potentially involved in redox reactivity and/or metal binding. Nucleic Acids Res. 1988, 16, 2207–2224. [Google Scholar] [CrossRef]
- Honorio-Felício, N.; Carepo, M.S.P.; de F Paulo, T.; de França Lopes, L.G.; Sousa, E.H.S.; Diógenes, I.C.N.; Bernhardt, P.V. The Heme-Based Oxygen Sensor Rhizobium etli FixL: Influence of Auxiliary Ligands on Heme Redox Potential and Implications on the Enzyme Activity. J. Inorg. Biochem. 2016, 164, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Mesa, S.; Reutimann, L.; Fischer, H.M.; Hennecke, H. Posttranslational control of transcription factor FixK2, a key regulator for the Bradyrhizobium japonicum—Soybean symbiosis. Proc. Natl. Acad. Sci. USA 2009, 106, 21860–21865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, E.; Kaspar, T.; Fischer, H.-M.; Hennecke, H. Expression of the fixR-nifA Operon in Bradyrhizobium japonicum Depends on a New Response Regulator, RegR. J. Bacteriol. 1998, 180, 3853–3863. [Google Scholar] [CrossRef] [Green Version]
- Bueno, E.; Mesa, S.; Bedmar, E.J.; Richardson, D.J.; Delgado, M.J. Bacterial Adaptation of Respiration from Oxic to Microoxic and Anoxic Conditions: Redox Control. Antioxid. Redox Signal. 2012, 16, 819–852. [Google Scholar] [CrossRef]
- Oger, E.; Marino, D.; Guigonis, J.M.; Pauly, N.; Puppo, A. Sulfenylated proteins in the Medicago truncatula-Sinorhizobium meliloti symbiosis. J. Proteom. 2012, 75, 4102–4113. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.; Poole, P.S. Transport and Metabolism in Legume-Rhizobia Symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadouri, D.; Burdman, S.; Jurkevitch, E.; Okon, Y. Identification and isolation of genes involved in poly(β-hydroxybutyrate) biosynthesis in Azospirillum brasilense and characterization of a phbC mutant. Appl. Environ. Microbiol. 2002, 68, 2943–2949. [Google Scholar] [CrossRef] [Green Version]
- Batista, M.B.; Teixeira, C.S.; Sfeir, M.Z.T.T.; Alves, L.P.S.S.; Valdameri, G.; de Oliveira Pedrosa, F.; Sassaki, G.L.; Steffens, M.B.R.; de Souza, E.M.; Dixon, R.; et al. PHB biosynthesis counteracts redox stress in Herbaspirillum seropedicae. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, M.A.; Encarnacion, S.; Leija, A.; Mora, Y.; Mora, J. Genetic and physiological characterization of a Rhizobium etli mutant strain unable to synthesize poly-β-hydroxybutyrate. J. Bacteriol. 1996, 178, 1646–1654. [Google Scholar] [CrossRef] [Green Version]
- Walshaw, D.L.; Wilkinson, A.; Mundy, M.; Smith, M.; Poole, P.S. Regulation of the TCA cycle and the general amino acid permease by overflow metabolism in Rhizobium leguminosarum. Microbiology 1997, 143, 2209–2221. [Google Scholar] [CrossRef] [Green Version]
- Kadouri, D.; Jurkevitch, E.; Okon, Y. Involvement of the reserve material poly-β-hydroxybutyrate in Azospirillum brasilense stress endurance and root colonization. Appl. Environ. Microbiol. 2003, 69, 3244–3250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trainer, M.A.; Charles, T.C. The role of PHB metabolism in the symbiosis of rhizobia with legumes. Appl. Microbiol. Biotechnol. 2006, 71, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Terpolilli, J.J.; Masakapalli, S.K.; Karunakaran, R.; Webb, I.U.C.C.; Green, R.; Watmough, N.J.; Kruger, N.J.; Ratcliffe, R.G.; Poole, P.S. Lipogenesis and redox balance in nitrogen-fixing pea bacteroids. J. Bacteriol. 2016, 198, 2864–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Colonization patterns of endosymbiotic and endophytic bacteria in interaction with the plant. (A) Schematic representation of bacterial distribution within the plant. Rhizospheric bacteria (green) spread into the rhizosphere but do not directly interact with plant tissues. Endophytic bacteria (red) colonize the plant surface and intercellular space of plant tissues. Endosymbiotic bacteria (purple) are able to induce the formation of the stem (A1) or root (A2) nodules containing nitrogen-fixing bacteroids. In exchange for the ammonia provided by these endosymbiotic bacteria, plant transfer carbohydrates derived from photosynthesis. N2: atmospheric nitrogen; NH3: ammonia; C: carbohydrates; RE: root exudates. (B) Pictures of endosymbiotic and endophytic bacterial distribution within plants. For endosymbiotic bacteria, Live/Dead microscopic analysis of mature nodules (B1) and infected cells in zone III (B2) formed during M. truncatula/S. meliloti symbiosis is represented. For endophytic bacteria, longitudinal sections of a maize root colonized by GFP-labelled Klebsiella pneumoniae (B3; B4) are represented reprint from reference [9] with kind permission from the American Society for Microbiology. Scale bar: 100 µm (B1), 10 µm (B2; B4), or 20 µm (B3).
Figure 1.
Colonization patterns of endosymbiotic and endophytic bacteria in interaction with the plant. (A) Schematic representation of bacterial distribution within the plant. Rhizospheric bacteria (green) spread into the rhizosphere but do not directly interact with plant tissues. Endophytic bacteria (red) colonize the plant surface and intercellular space of plant tissues. Endosymbiotic bacteria (purple) are able to induce the formation of the stem (A1) or root (A2) nodules containing nitrogen-fixing bacteroids. In exchange for the ammonia provided by these endosymbiotic bacteria, plant transfer carbohydrates derived from photosynthesis. N2: atmospheric nitrogen; NH3: ammonia; C: carbohydrates; RE: root exudates. (B) Pictures of endosymbiotic and endophytic bacterial distribution within plants. For endosymbiotic bacteria, Live/Dead microscopic analysis of mature nodules (B1) and infected cells in zone III (B2) formed during M. truncatula/S. meliloti symbiosis is represented. For endophytic bacteria, longitudinal sections of a maize root colonized by GFP-labelled Klebsiella pneumoniae (B3; B4) are represented reprint from reference [9] with kind permission from the American Society for Microbiology. Scale bar: 100 µm (B1), 10 µm (B2; B4), or 20 µm (B3).

Figure 2.
General bacterial antioxidant defense. Bacteria protect themselves from ROS thanks to enzymes (SOD: superoxide dismutase; Kat: catalase; Px: peroxidase; Prx: peroxiredoxin; Trx: thioredoxin; Msr: methionine sulfoxide reductase) and low molecular weight thiol glutathione (GSH). GSH can be used by glutathione-S-transferase (GST) or glutaredoxin (Grx) to detoxify ROS. Glutathione reductase (GR) allows the regeneration of GSH from the oxidized form GSSG. Grx works with Trx in order to reduce protein disulfides, and Msr reduces methionine sulfoxide residues to methionine. GR, Trx, and Msr use NAD(P)H as an electron donor.
Figure 2.
General bacterial antioxidant defense. Bacteria protect themselves from ROS thanks to enzymes (SOD: superoxide dismutase; Kat: catalase; Px: peroxidase; Prx: peroxiredoxin; Trx: thioredoxin; Msr: methionine sulfoxide reductase) and low molecular weight thiol glutathione (GSH). GSH can be used by glutathione-S-transferase (GST) or glutaredoxin (Grx) to detoxify ROS. Glutathione reductase (GR) allows the regeneration of GSH from the oxidized form GSSG. Grx works with Trx in order to reduce protein disulfides, and Msr reduces methionine sulfoxide residues to methionine. GR, Trx, and Msr use NAD(P)H as an electron donor.


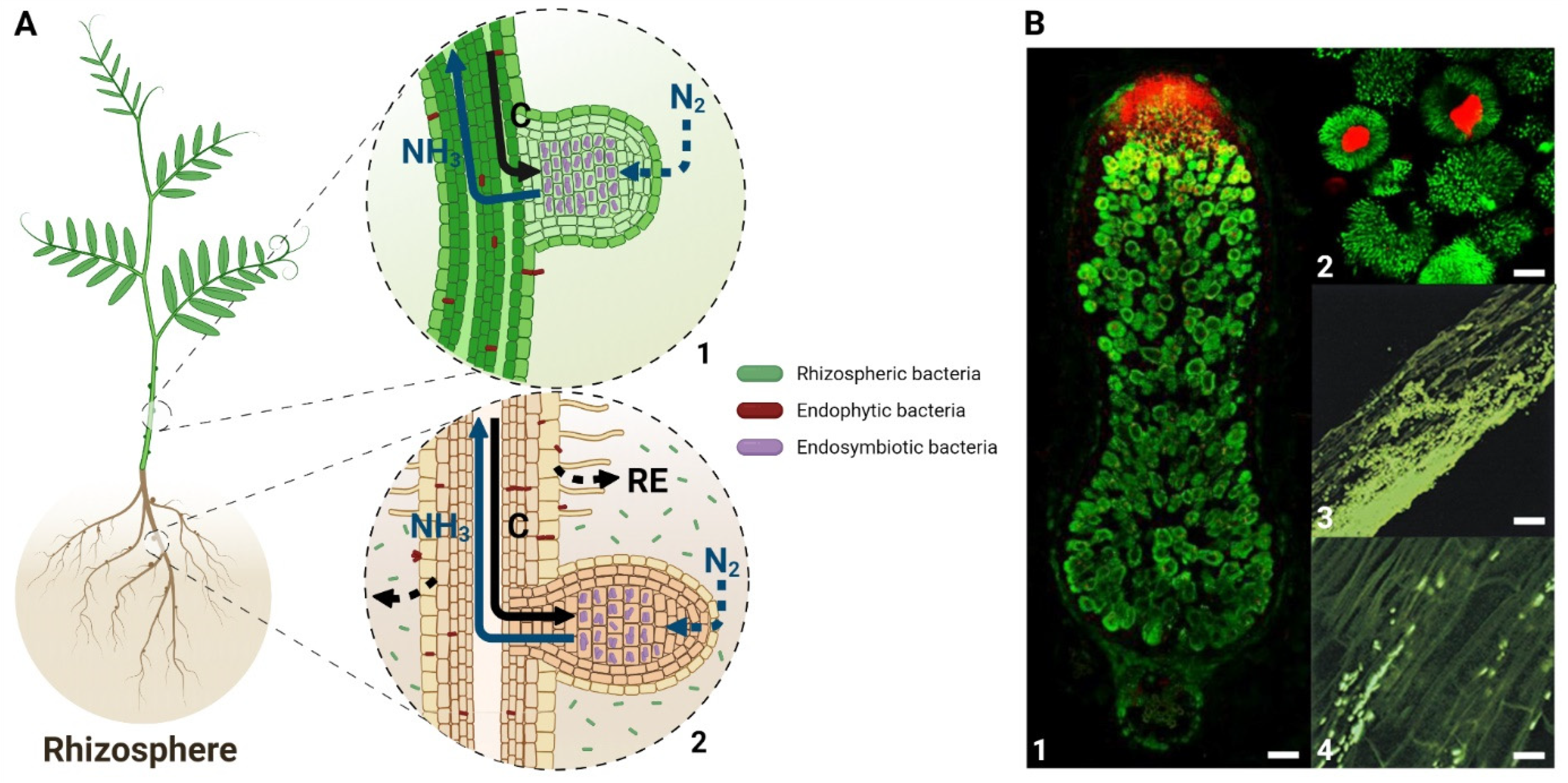{kind=link}
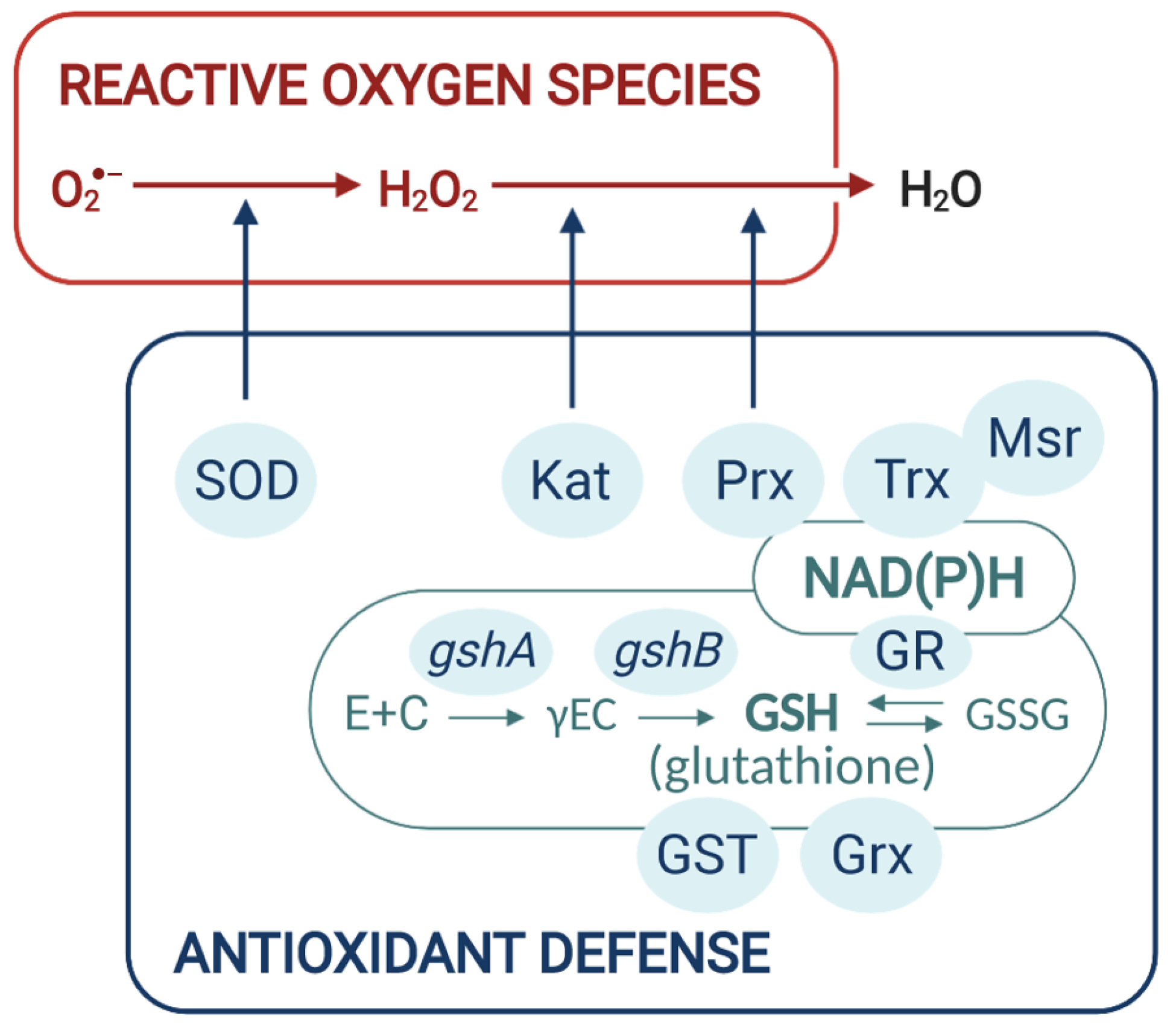{kind=link}
Table 1.
Genes of diazotrophic rhizobia involved in cellular redox homeostasis and in the efficiency of endophytic/endosymbiotic interaction. Phenotypes in planta are the results of gene inactivation except for S. meliloti katB mutant that overexpresses katB. PHB, polyhydroxybutyrate.
Table 1.
Genes of diazotrophic rhizobia involved in cellular redox homeostasis and in the efficiency of endophytic/endosymbiotic interaction. Phenotypes in planta are the results of gene inactivation except for S. meliloti katB mutant that overexpresses katB. PHB, polyhydroxybutyrate.
| Genes | Bacterium | Mutant Phenotype In planta | References |
|---|---|---|---|
| ENDOPHYTES | |||
| sod | G. diazotrophicus | impaired root colonization | [62] |
| gr | G. diazotrophicus | impaired root colonization | [62] |
| phbC | H. seropedicae | loss of plant growth promoting ability | [63] |
| ENDOSYMBIONTS | |||
| ROS scavenging | |||
| sodA | S. meliloti | depending on the S. meliloti strain genotype | [64] |
| katB | S. meliloti | delayed nodulation; aberrant and enlarged infection threads | [28] |
| katB/katC | S. meliloti | altered bacteroid differentiation | [65] |
| katA/katC | S. meliloti | early nodule senescence | [65] |
| katE | M. loti | lower nitrogen fixation capacity | [66] |
| katG/prxS | R. etli | lower nitrogen fixation capacity | [67] |
| prxA | M. huakii | lower nitrogen fixation capacity | [68] |
| katG | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [69] |
| ahpCD | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [70] |
| ohr | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [71] |
| GSH/Grx system | |||
| gshA | R. leguminosarum | impaired root colonization, lower nitrogen fixation capacity | [72] |
| gshB | R. leguminosarum | impaired root colonization, lower nitrogen fixation capacity | [72,73] |
| gshB | S. meliloti | delayed nodulation, early nodule senescence | [74,75] |
| gshB | R. etli | delayed nodulation, early nodule senescence | [76] |
| gshB | R. tropici | low competitiveness for nodule occupancy, early nodule senescence | [77] |
| gshB | M. huakii | early nodule senescence | [78] |
| gor | S. meliloti | low competitiveness for nodule occupancy, lower nitrogen fixation capacity | [79] |
| grx (RL2615) | R. leguminosarum | impaired root colonization | [72] |
| grx1 | S. meliloti | altered bacteroid differentiation | [80] |
| grx2 | S. meliloti | early nodule senescence | [80] |
| grx1/grx2 | A. caulinodans | lower nitrogen fixation capacity | [81] |
| grx3/grx4 | A. caulinodans | lower nitrogen fixation capacity | [81] |
| PHB metabolism | |||
| phbC | S. meliloti | lower nitrogen fixation capacity | [82] |
| phbC | A. caulinodans | loss of nitrogen fixation ability | [83] |
| phbC | B. japonicum | low competitiveness for root colonization | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mandon, K.; Nazaret, F.; Farajzadeh, D.; Alloing, G.; Frendo, P. Redox Regulation in Diazotrophic Bacteria in Interaction with Plants. Antioxidants 2021, 10, 880. https://doi.org/10.3390/antiox10060880
AMA Style
Mandon K, Nazaret F, Farajzadeh D, Alloing G, Frendo P. Redox Regulation in Diazotrophic Bacteria in Interaction with Plants. Antioxidants. 2021; 10(6):880. https://doi.org/10.3390/antiox10060880
Chicago/Turabian StyleMandon, Karine, Fanny Nazaret, Davoud Farajzadeh, Geneviève Alloing, and Pierre Frendo. 2021. "Redox Regulation in Diazotrophic Bacteria in Interaction with Plants" Antioxidants 10, no. 6: 880. https://doi.org/10.3390/antiox10060880
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.